Introduction
Coronary artery disease (CAD) is a type of heart
disease caused by atherosclerotic lesions of coronary arteries,
which is characterized by stenosis or obstruction of the vascular
cavity that further leads to myocardial ischemia, hypoxia or
necrosis (1). Atherosclerosis, the
primary cause of cardiovascular disease, is a chronic inflammatory
process involving inflammatory plaques, different types of cells
with multiple complex interactions and abnormal lipoprotein
deposits in the arterial wall (2,3).
Hypertension, diabetes mellitus, tobacco consumption, alcohol
consumption, sex (male), obesity, dyslipidemia, genetic variance
and a lack of physical activity are commonly known risk factors for
CAD (4,5).
Long noncoding RNAs (lncRNAs) are a class of
noncoding RNAs with a transcript length >200 bp that lack
protein-coding potential. Due to their cell and tissue specificity,
lncRNAs in blood, plasma or urine can be used as biomarkers and
therapeutic targets for cancer and numerous other diseases
(6,7). An increasing number of studies have
shown that lncRNAs are abnormally expressed in a variety of
diseases and have important roles in cell proliferation, apoptosis
and invasion (8–10).
It has been demonstrated that lncRNAs are closely
associated with the occurrence and development of CAD, including
CAD-induced pathological changes such as vascular smooth muscle
cell proliferation, apoptosis, lipid metabolism and inflammation
(11,12). Various lncRNAs have been considered
promising regulatory genes or biomarkers of CAD, such as lncPPARδ,
H19 imprinted maternally expressed transcript, maternally expressed
3, antisense noncoding RNA in the INK4 locus (ANRIL),
mitochondrially encoded long non-coding cardiac associated RNA,
lincRNA-p21, CoroMarker, NONHSAT112178, Novlnc6, metastasis
associated lung adenocarcinoma transcript 1, myocardial infarction
associated transcript and steroid receptor RNA activator (SRA)
(13,14). In atherosclerosis, lincRNA-p21 has
been identified to inhibit proliferation and induce apoptosis in
vascular smooth muscle cells (15). In addition, the G-A-A-G haplotype
of lincRNA-p21 is correlated with a decreased risk of CAD (16).
CoroMarker (lncRNA AC100865.1), abhydrolase domain
containing 16A, and IL21R antisense RNA 1 were identified by
microarray and confirmed by reverse transcription-quantitative
(RT-q) PCR in peripheral blood mononuclear cells (PBMCs) (17). It has been shown that in CAD the
CoroMarker is significantly upregulated and has high specificity
and sensitivity (17). Studies
have also demonstrated that CoroMarker can positively regulate foam
cell formation, arteriosclerosis, inflammation and the associated
immune responses, and can inhibit cell apoptosis (17,18).
A number of lncRNAs have been shown to be associated
with CAD, however key lncRNAs in patients with CAD from different
regions and different ethnicities are likely to differ, so further
studies are required. In the present study, high-throughput
sequencing was adopted to identify differentially expressed lncRNAs
and identify a biomarker for CAD according to the local resident
(Suzhou, Jiangsu, China). Further study of the mechanism of the
lncRNA was performed to identify new targets for the early
diagnosis and treatment of clinical CAD. These results also
provided a theoretical basis for the further study of the molecular
mechanism of CAD.
Materials and methods
Clinical characteristics of
patients
A total of 187 patients with CAD, aged 42–85 years,
were enrolled in the Department of Cardiology of Nanjing Medical
University Affiliated Suzhou Hospital in China between October 2016
and August 2018. The diagnosis of CAD was based on angiographic
evidence of at least one segment of a major coronary artery,
including the left anterior descending, left circumflex and right
coronary arteries, with ≥50% stenosis (19). Exclusion criteria were as follows:
i) Patients with severe primary diseases, such as liver, kidney and
hematopoietic system; ii) patients with acute infection, trauma or
other operations; iii) patients with myocardial infarction,
cerebral infarction, cancer and other major diseases; iv) patients
with alcoholism and drug addictions; v) patients >85 years; and
vi) pregnant patients. During the same period, 150 healthy subjects
were selected from patients attending check-up appointments. The
routine blood test results (WBC: 4–10×109/L, HGB:
120–170 g/l (male), 110–150 g/l (female), PLT:
100–300×109/l), blood pressure (60-90/90-140 mmHg),
blood glucose (4-6.1 mmol/l), blood lipid (total cholesterol:
2.8–5.17 mmol/l, triglycerides: 0.56–1.7 mmol/l, cholesterol
lipids: 2.8–5.17 mmol/l) and electrocardiogram (ECG; physician
judgment) of those 150 healthy subjects were within the normal
ranges. The study was approved by the Ethics Committee of Nanjing
Medical University. Informed consent was obtained from the patients
and their families.
Collection and preparation of blood
samples
Peripheral blood samples (4–6 ml) were collected
from patients with CAD and healthy subjects using EDTA vacuum
anticoagulant blood vessels. After incubation for 30 min at 4°C,
the blood samples were centrifuged at 200 × g for 10 min (4°C).
Then, the plasma was recentrifuged at 1,600 × g for 10 min (4°C) to
remove debris. PBMCs were separated from the blood cells with
Ficoll-Paque PLUS density gradient media (GE Healthcare) as
previously described (20,21).
lncRNA-Seq high-throughput
sequencing
Plasma samples from 3 patients with CAD and 3
healthy subjects were subjected to lncRNA-Seq high-throughput
sequencing, followed by bioinformatics analysis performed by
CloudSeq Biotech. Total RNA (1 µg) was used to remove the ribosomal
(r)RNAs using Ribo-Zero rRNA Removal kits (Illumina, Inc.)
following the manufacturer's protocols.
Fragments per kilobase million (FPKM) of the
expression profiles of lncRNAs and mRNAs was calculated using
Cuffdiff software (v2.2.1, part of Cufflinks) (22). Fold change and P-value were
calculated based on FPKM to identify the differentially expressed
lncRNAs and mRNAs. Gene Ontology (GO) pathway enrichment analysis
was performed based on the differentially expressed mRNAs (23,24).
Competing endogenous (ce)RNA analysis
of lncRNAs
An RNA transcript with micro (mi)RNA response
elements (MREs) may serve as a competing endogenous (ce)RNA. ceRNAs
include pseudogene transcripts, lncRNAs, circular RNAs and mRNAs,
and these transcripts can compete for the same MREs to exert their
biological functions. Potential target miRNAs were predicted with
in-house miRNA target prediction software, based on TargetScan and
miRanda software (25–31). Cytoscape software (v3.7.0) was used
to visualize the ceRNA networks with the target miRNAs (32).
To further analyze the metabolic pathway of lncRNA
ENST00000416361, a ceRNA network map was constructed according to
the corresponding target miRNAs and mRNAs. The physiological effect
of lncRNA ENST00000416361 was studied in depth by analyzing the
biological functions of related miRNAs and mRNAs.
Cell culture
HUVECs (human umbilical vein endothelial cells) were
purchased from CoWin Biosciences. HUVECs were cultured in DMEM
(Thermo Fisher Scientific, Inc.) supplemented with 100 U/ml
streptomycin, 100 U/ml penicillin and 10% (v/v) FBS (Gibco; Thermo
Fisher Scientific, Inc.) under 37°C and 5% (v/v) CO2
conditions.
Design of small interfering (si)RNAs
and cell transfection
The siRNAs were designed and synthesized by Suzhou
GenePharma Co., Ltd. The sequences of si-ENST00000416361 were:
Sense (5′-3′), CCCAACAGCUCAUUGAGAATT and antisense (5′-3′),
UUCUCAAUGAGCUGUUGGGTT. The sequences of siRNA-NC were: Sense
(5′-3′), UUCUCCGAACG UGUCACGUTT and antisense (5′-3′),
ACGUGACACGUUCGG AGAATT. A total of 100 nM siRNA was used for each
transfection. A negative control with fluorescein was used to
identify cell transfection efficiency. Cell transfection was
performed using Lipofectamine™ 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) following the manufacturer's protocol. Knockdown
efficiency and related gene expression were measured after 12
h.
RNA isolation and RT-qPCR
Plasma RNA was extracted by TRIzol™ LS Reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), and 1 µg RNA was
transcribed into cDNA by 4 µl 5X PrimeScript™ RT Master Mix
(RR036A) (Takara Bio, Inc.), up to 20 µl with RNase free
dH2O. RNAiso Plus (Takara Bio, Inc.) was used to extract
RNA from PBMCs and HUVECs, followed by reverse transcription to
cDNA with PrimeScript™ RT Master Mix (RR036A) (Takara Bio, Inc.).
RNA quality was analyzed with an ultra-micro nucleic acid analyzer
(Hangzhou Allsheng Instruments Co., Ltd.).
RT-qPCR was performed using SYBR® Premix
Ex Taq™ II (Takara Bio, Inc.) to detect relative expression using
the standard protocols on the LightCycler® 480
Instrument II Real-Time PCR Detection system (Roche Diagnostics).
β-actin was used as the internal reference. PCR reaction conditions
were: 95°C for 30 sec; 40 cycles of 95°C for 5 sec and 60°C for 20
sec; 95°C for 0 sec, 65°C for 15 sec and 95°C for 0 sec. The
percentage of agarose gel was 2%. The primers for lncRNAs, the
lipid-associated genes SREBP1, FAS, ACC1, DGAT2, SREBP2, ApoE,
ApoA-I, Cpt1a (33) and β-actin
used in the RT-qPCR were synthesized by Genewiz, Inc. The primer
sequences are listed in Table I.
All measurements were performed in triplicate. Relative expression
data were analyzed with the formula 2−∆∆Cq (34). GraphPad Prism 7 (GraphPad Software,
Inc.) was used to analyze the differences in expression.
 | Table I.The primer sequences. |
Table I.
The primer sequences.
| Gene | Primer
sequence |
|---|
|
ENST00000416361 |
5′-CTCATTGAGAACGGGCCATG-3′ |
|
|
5′-CATGTTTCAGAGAGCACCGG-3′ |
| SREBP1 |
5′-GGAGGGGTAGGGCCAACGGCCT-3′ |
|
|
5′-CATGTCTTCGAAAGTGCAATCC-3′ |
| SREBP2 |
5′-CAAGATGCACAAGTCTGGCG-3′ |
|
|
5′-GCTTCAGCACCATGTTCTCCTG-3′ |
| FAS |
5′-ACAGGGACAACCTGGAGTTCT-3′ |
|
|
5′-CTGTGGTCCCACTTGATGAGT-3′ |
| ACC1 |
5′-GTTGCACAAAAGGATTTCAG-3′ |
|
|
5′-CGCATTACCATGCTCCGCAC-3′ |
| DGAT2 |
5′-CGGTCCCCAATCACCTCATC-3′ |
|
|
5′-GGGATGTTCCAGTTCTGCCA-3′ |
| ApoE |
5′-CCCAGGTCACCCAGGAACT-3′ |
|
|
5′-TTCCGATTTGTAGGCCTTCAA-3′ |
| ApoA-I |
5′-ATCGAGTGAAGGACCTGGC-3′ |
|
|
5′-AGCTTGCTGAAGGTGGAGGT-3′ |
| Cpt1a |
5′-TGCTTTACAGGCGCAAACTG-3′ |
|
|
5′-TGGAATCGTGGATCCCAAA-3′ |
| β-actin |
5′-CACGAAACTACCTTCAACTCC-3′ |
|
|
5′-CATACTCCTGCTTGCTGATC-3′ |
Cytokine detection
The human Th1/Th2 subgroup test kit (CellGene
Biotech Co., Ltd.) was used to detect the levels of the
inflammatory cytokines interleukin (IL)-2, IL-4, IL-6, IL-10,
interferon-γ and tumor necrosis factor-α (TNF-α). The standard
solution and standard curves were prepared according to the
manufacturer's protocols. The mixed microsphere solution was
centrifuged at 200 × g for 5 min (4°C). Subsequently, the same
volume of microsphere buffer solution was added and incubated for
30 min in the dark at room temperature. For each tube, 25 µl
microsphere mixture, 25 µl sample and 25 µl fluorescent detection
reagent was added, and the solution was incubated for 2.5 h at room
temperature. After washing with 1 ml of PBS, the mixture was
centrifuged at 200 × g for 5 min at room temperature, and the
precipitate was diluted in 100 µl of PBS solution. Finally,
cytokine detection was performed using BD FACSCalibur™
(Becton-Dickinson and Company).
Statistical analysis
GraphPad Prism 7 software was used for statistical
analysis. All data were obtained from at least 3 independent
experiments and are expressed as the mean ± SD. One-way ANOVA
followed by Dunnett's test was performed to analyze multiple group
comparisons of quantitative data and Student's t-test was performed
for analyzing two-group comparisons of quantitative data. A
Chi-square test was used for category data. Correlation was
confirmed by Pearson's correlation analysis. P<0.05 was
considered statistically significant.
Results
Pathological differences between
patients with CAD and healthy subjects
A total of 187 patients with CAD (diagnosed with at
least one vascular ≥50% stenosis) and 150 healthy subjects were
selected for the experiment. The clinical characteristics of the
enrolled patients and healthy controls are presented in Table II. The data showed that 82.9% of
patients with CAD had hypertension, 28.9% had diabetes, 45% had a
history of smoking and were male, all of which were significantly
increased compared with those in the control group. In addition,
high-density lipoprotein (HDL) levels were significantly decreased
in patients with CAD relative to the controls.
| Table II.Clinical characteristics of patients
with CAD and controls. |
Table II.
Clinical characteristics of patients
with CAD and controls.
|
| Sequencing
samples | All clinical
samples collected |
|---|
|
|
|
|
|---|
|
Characteristics | CAD (n=3) | CON (n=3) | CAD (n=187) | CON (n=150) | P-value |
|---|
| Age, years | 60.33 | 57.78 | 67.5±17.5 | 62.06±18.06 | >0.050 |
| Sex, male (%) | 2 (66.7) | 2 (66.7) | 115 (61.5) | 78 (52) | 0.083 |
| BMI
(kg/m2) | 24.32 | 23.99 | 25.13 | 25.68 | >0.050 |
| Diabetes mellitus
(%) | 0 (0) | 0 (0) | 54 (28.9) | 22 (14.7) | 0.002 |
| Hypertension
(%) | 1 (33.4) | 0 (0) | 155 (82.9) | 58 (38.7) | <0.001 |
| TC (mmol/l) | 3.56 | 3.69 | 4.14 | 4.589 | 0.066 |
| TG (mmol/l) | 1.57 | 1.77 | 1.7 | 1.56 | 0.679 |
| HDL (mmol/l) | 0.98 | 1.08 | 1.02 | 1.19 | 0.002 |
| LDL (mmol/l) | 2.13 | 2.21 | 2.56 | 2.89 | 0.171 |
| VLDL (mmol/l) | 0.34 | 0.5 | 0.56 | 0.53 | 0.425 |
| ApoA (g/l) | 1.32 | 1.11 | 1.21 | 1.25 | 0.776 |
| ApoB (g/l) | 0.71 | 0.69 | 0.82 | 0.83 | 0.421 |
| Lipoprotein A | 200.67 | 162.09 | 207.24 | 177 | 0.051 |
| CHE | 8.41 | 8.74 | 7.8 | 8.04 | 0.773 |
| TBA | 5.23 | 5.15 | 5.77 | 5.56 | 0.894 |
| Smoking (%) | 0 (0) | 0 (0) | 84 (45) | 40 (26.7) | 0.001 |
| Drinking (%) | 0 (0) | 0 (0) | 37 (19.8) | 20 (13.3) | 0.116 |
Differential expression profiles of
lncRNAs
Plasma samples from 3 patients with CAD and 3
healthy controls were examined via high-throughput sequencing
methods. The 3 patients with CAD were screened based on their
clinical records, in order to exclude the interference of other
diseases as well as to avoid including the effects of smoking and
drinking. In the 3 healthy controls, all the physical examination
indexes were in the normal range, and the interference of other
diseases was excluded. All the physical examination indexes were in
the normal range, and the interference of other diseases was
excluded of the normal group. By analyzing FPKM, fold changes and
P-values using the Cuffdiff software, 66 upregulated and 86
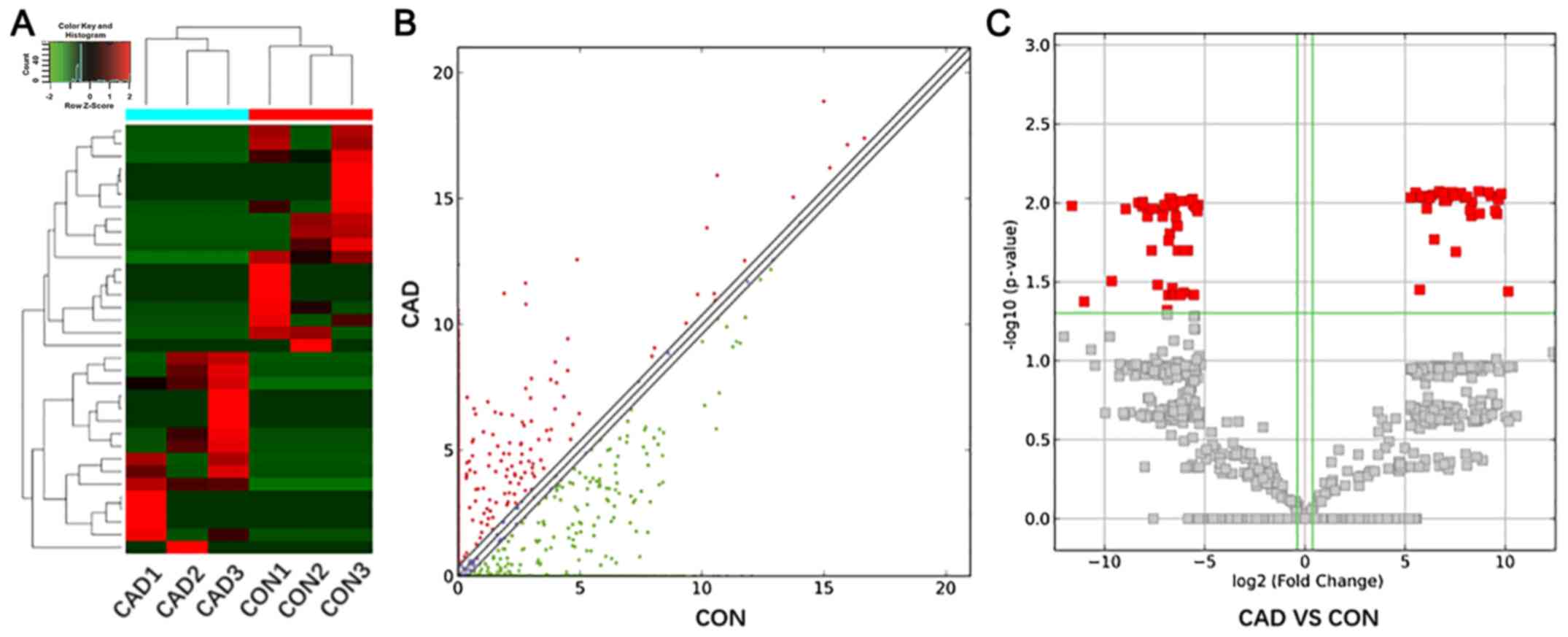
downregulated lncRNAs were identified. Cluster analysis, scatter
diagram and volcano plots show the differentially expressed lncRNAs
with a P≤0.05 and fold change ≥2.0 (Fig. 1).
GO pathway analysis of differentially
expressed lncRNAs
Target genes of lncRNAs were predicted based on
their proximity to the lncRNAs, and GO pathway analysis of
biological processes was performed based on the identified target
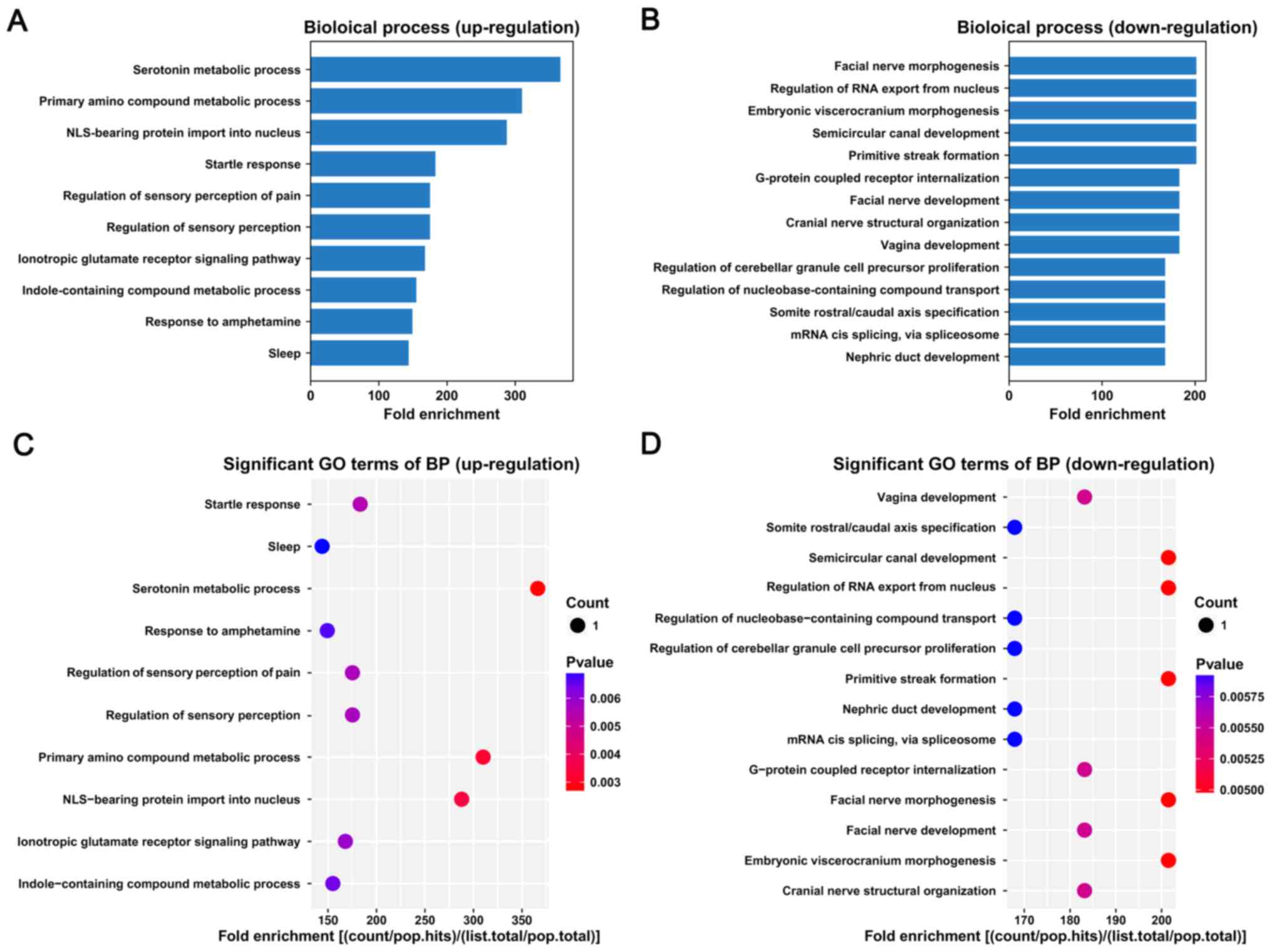
genes. The bar plot shows the top fold enrichment value of the
significantly enriched terms (Fig.
2). The target genes of upregulated lncRNAs were associated
with serotonin metabolic process, primary amino compound metabolic
process and nuclear localization signal (NLS)-bearing protein
import into nucleus (Fig. 2A and
C). The target genes of the downregulated lncRNAs were mainly
involved in primitive streak formation, semicircular canal
development, embryonic viscerocranium morphogenesis, regulation of
RNA export from nucleus and facial nerve morphogenesis (Fig. 2B and D).
Target lncRNA selection
The top 5 upregulated and downregulated lncRNAs with
P<0.01 were selected, and overlapping sequences were excluded
(Table III).
| Table III.Target lncRNA selection. |
Table III.
Target lncRNA selection.
| Transcript_ID | Fold change | P-value | FDR | Regulation | LncRNA_source | LncRNA_length
(bp) | Relationship |
|---|
|
ENST00000416361 | inf | 0.009 | 0.400733 | Up | Ensembl | 2,102 | Intergenic |
|
ENST00000565257 | inf | 0.010 | 0.400733 | Up | Ensembl | 1,406 | Intergenic |
|
ENST00000568324 | inf | 0.010 | 0.400733 | Up | Ensembl | 3,102 | Intergenic |
| uc003pxg.1 | inf | 0.009 | 0.400733 | Up | UCSC knowngene | 2,183 | Nidirectional |
|
ENST00000607001 | inf | 0.010 | 0.400733 | Up | Ensembl | 744 | Nidirectional |
|
ENST00000418006 | -inf | 0.009 | 0.400733 | Down | Ensembl | 2,350 | Intergenic |
|
ENST00000564481 | -inf | 0.010 | 0.400733 | Down | Ensembl | 992 | Nidirectional |
|
ENST00000537921 | -inf | 0.011 | 0.400733 | Down | Ensembl | 477 | Nidirectional |
|
ENST00000554522 | -inf | 0.009 | 0.400733 | Down | Ensembl | 1,701 | Intergenic |
|
ENST00000425077 | -inf | 0.010 | 0.400733 | Down | Ensembl | 422 | Intergenic |
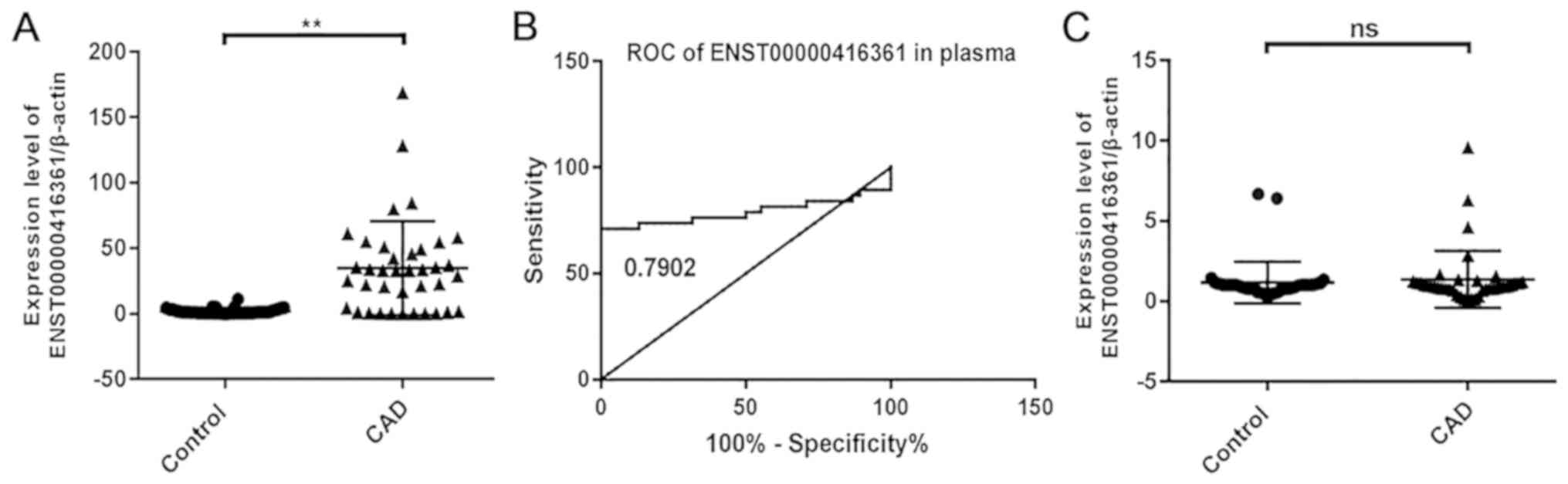
lncRNA ENST00000416361 is
significantly upregulated in the plasma of patients with CAD
A total of 3 pairs of primers were designed for each
lncRNA, and the results showed that ENST00000416361 was upregulated
in the peripheral blood plasma samples of patients with CAD
(Fig. 3A). However, the
upregulation trend of ENST00000416361 was not obvious in PBMCs
(Fig. 3C). The relative expression
levels of these lncRNAs in plasma and PBMC samples from 50 patients
with CAD and 50 healthy controls were verified by RT-qPCR. Then, 50
samples per group were randomly selected from the 187 patients with
CAD and 150 controls enrolled in the study. The results indicated
that the expression level of ENST00000416361 in patients with CAD
was 2.3-fold higher compared with that in the normal group, and
P<0.01 was used as a cut-off value to indicate a statistically
significant difference. A ROC curve was generated for analyzing the
early-stage screening and diagnostic potential of ENST00000416361
in CAD (Fig. 3B). The area under
the curve was 0.7902, indicating that the plasma level of
ENST00000416361 may represent a useful clinical diagnostic
biomarker for CAD.
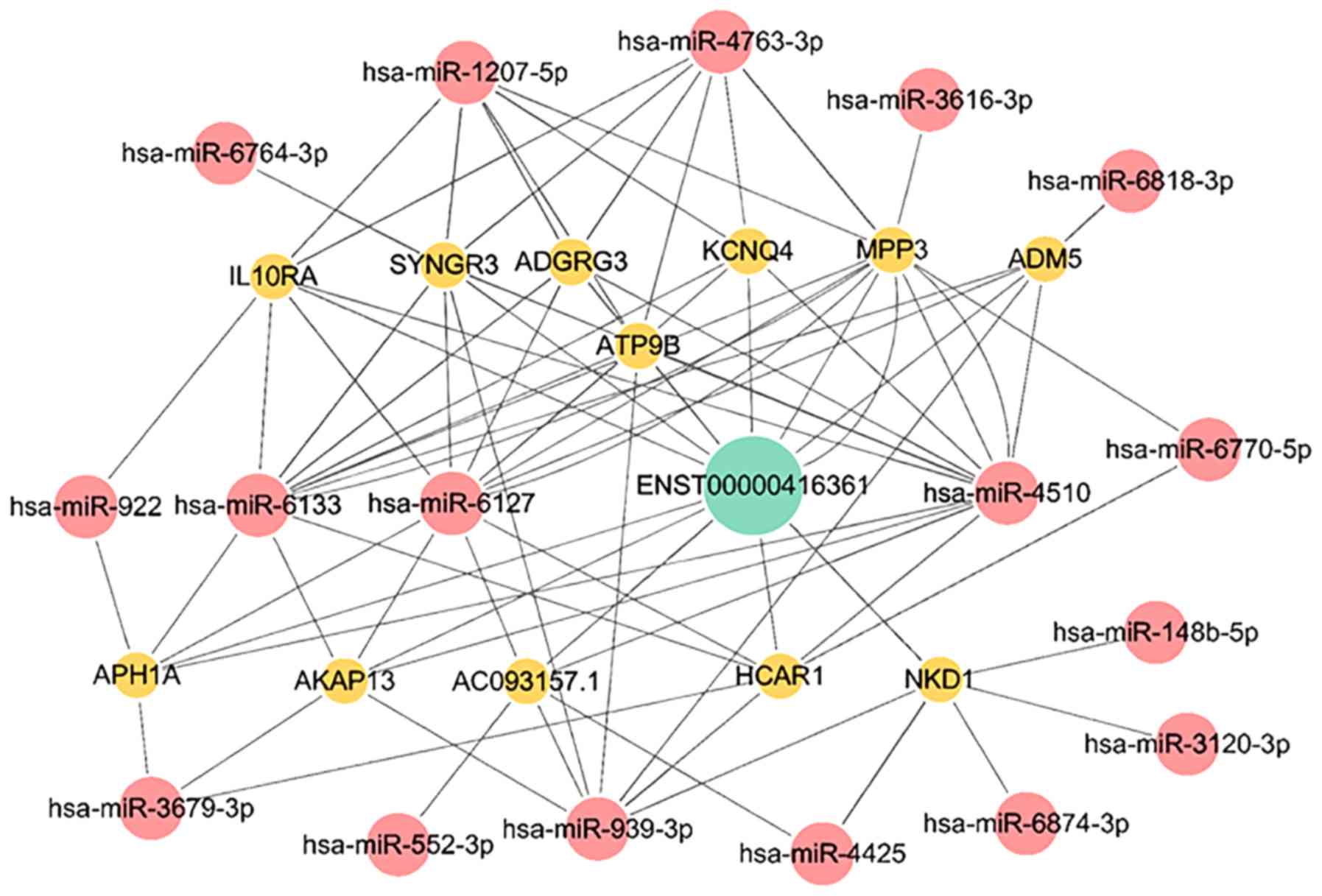
lncRNA ENST00000416361 serves as a
ceRNA
Bioinformatics analysis was conducted to explore the
gene metabolic pathway of lncRNA ENST00000416361. A ceRNA network
map was constructed with the relevant miRNAs and mRNAs (Fig. 4).
Among these related mRNAs, synaptogyrin 3, membrane
palmitoylated protein 3, adhesion G protein-coupled receptor G3,
aph-1 homolog A, γ-secretase subunit, hydroxycarboxylic acid
receptor 1, ATPase phospholipid transporting 9B and potassium
voltage-gated channel subfamily Q member 4 were associated with
membrane structures.
The miRNA-939 expression is downregulated in
patients with CAD. miRNA-939 inhibits angiogenesis by targeting
human-catenin proteins, thus destroying the integrity of blood
vessels. Overexpression of miRNA-939 in HUVECs can significantly
inhibit cell proliferation, adhesion and angiogenesis but increase
cell migration (35).
Inflammatory factors IL-6 and TNF-α
are significantly downregulated following lncRNA ENST00000416361
knockdown
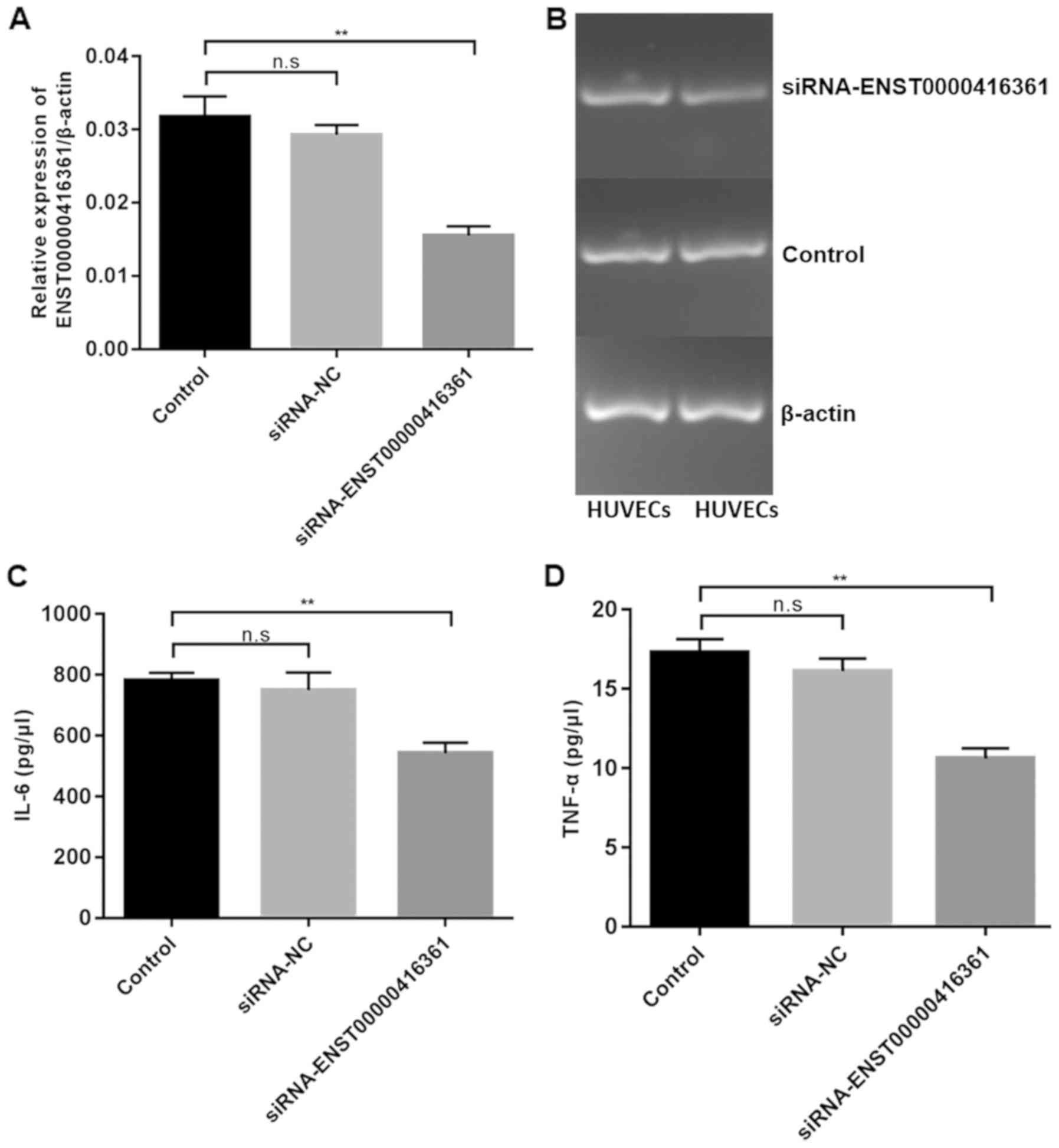
Transfection of si-ENST00000416361 in HUVECs
exhibited a 50% knockdown efficiency (Fig. 5A and B). Furthermore, transfection
of si-ENST00000416361 led to the downregulation of IL-6 (Fig. 5C) and TNF-α (Fig. 5D).
Lipid-associated genes SREBP1 and
SREBP2 are positively correlated with the expression of
ENST00000416361
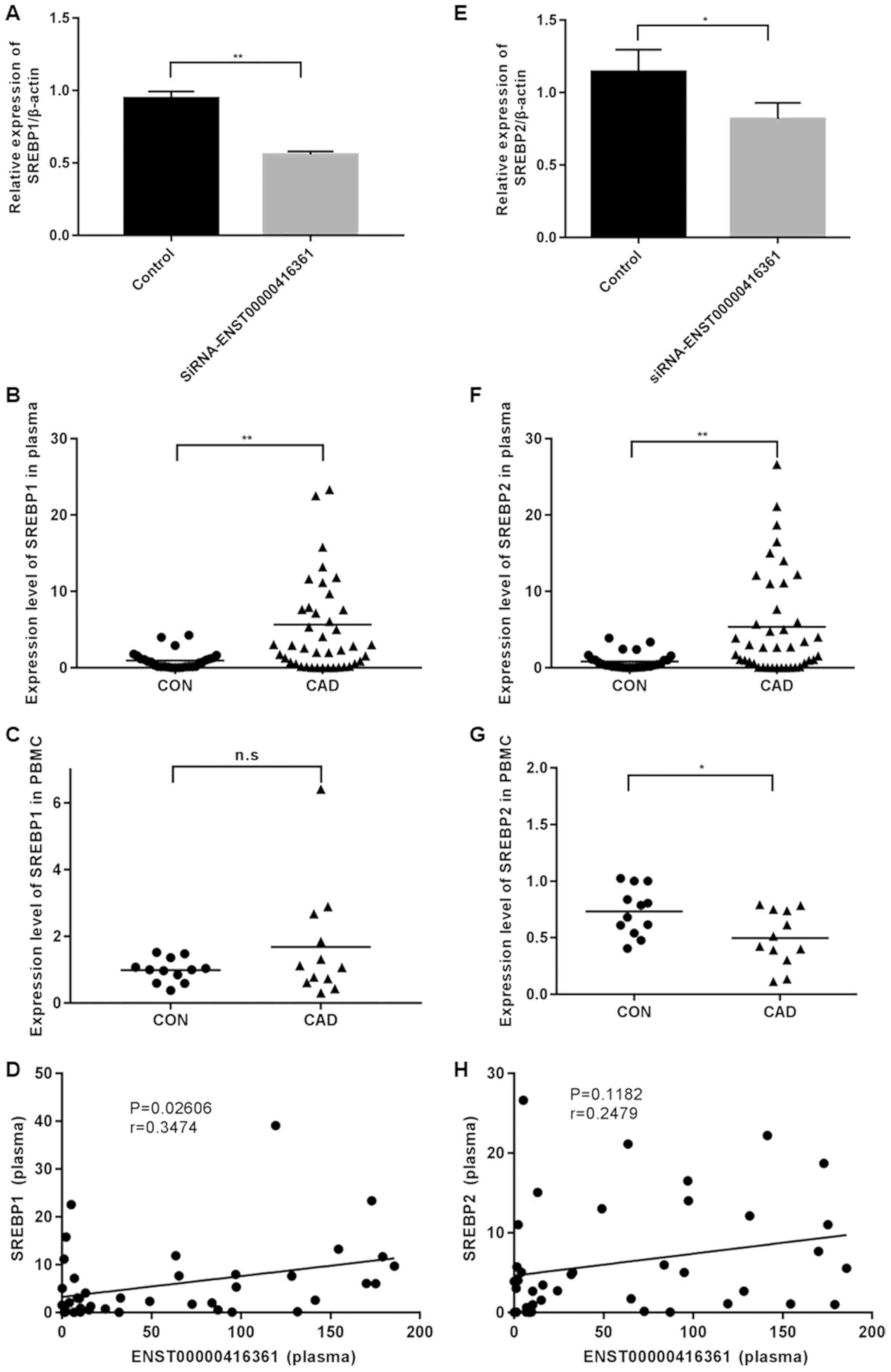
It was identified that SREBP1 and SREBP2 were
significantly upregulated in plasma samples of patients with CAD
(Fig. 6B and F), but not in PBMCs
(Fig. 6C and G). Knockdown of
ENST00000416361 led to the significant downregulation of SREBP1 and
SREBP2 (Fig. 6A and E). Pearson's
correlation scatter plots were generated based on the plasma levels
of ENST00000416361 and SREBP, and it was indicated that
ENST00000416361 could affect the occurrence and development of CAD
by interacting with SREBP. The levels of SREBP1 (P=0.02606,
r=0.3474) and SREBP2 (P=0.1182, r=0.2479) were both positively
correlated with the expression of ENST00000416361, which was more
pronounced in the former gene (Fig. 6D
and H).
Discussion
lncRNAs have been identified to be closely
associated with the occurrence and development of CAD (36). These differentially expressed
lncRNAs have attracted increasing attention due to their value in
disease diagnosis and prognosis. In the present study,
high-throughput sequencing and pathway analysis were conducted to
find the differentially expressed lncRNAs in the peripheral blood
of patients with CAD. The top upregulated lncRNAs were primarily
associated with serotonin metabolic process, primary amino compound
metabolic process and NLS-bearing protein import into nucleus. By
analyzing clinical indexes of patients with CAD and healthy
subjects, it was identified that HDL levels were significantly
decreased in patients with CAD. The expression pattern of lncRNA
ENST00000416361 was determined in plasma samples and PBMCs,
indicating that the plasma level of lncRNA ENST00000416361 may be a
potential biomarker for clinical CAD screening.
Knockdown of ENST00000416361 significantly decreased
the levels of the inflammatory factors IL-6 and TNF-α. A growing
number of studies have shown that IL-6 and TNF-α are closely
related to CAD progression. It has been demonstrated that the serum
levels of IL-6 and TNF-α are increased in patients with CAD and are
positively correlated with miR-342-5p expression (37). Decreased serum levels of C1q/TNF-α
related protein 12 (CTRP12) have been identified in patients with
CAD compared to controls, and IL-6 and TNF-α are negatively
correlated with CTRP12 in patients with CAD (38). It has been suggested that
inflammatory cytokines, particularly IL-6, may be elicited in
response to an acute inflammatory stressor, thus contributing to
the progression of CAD (39,40).
The C allele of rs1800795 is located on the IL-6 gene promoter, and
IL-6 rs1800795-tobacco smoking and IL-6 rs1800795-alcohol
consumption interactions have been associated with increased CAD
risk (41). lncRNA ANRIL affects
the progression of CAD by targeting miR-181b, activating the NF-κB
signal pathway and inducing the release of inflammatory factors
such as IL-6 and TNF-α (42).
In addition, SREBP1 and SREBP2, which were
significantly upregulated in CAD plasma samples, were downregulated
following the knockdown of lncRNA ENST00000416361. SREBPs are
important molecules in lipid synthesis and are encoded by 2 SREBP
genes, SREBP1 and SREBP2 (43).
SREBP1 is primarily distributed in the liver and adrenal gland and
it is closely associated with genes involved in fatty acid
metabolism (44). SREBP2 is widely
expressed and participates in cholesterol biosynthesis and
metabolism (44,45). Increasing studies have focused on
the correlation between SREBP and CAD. SREBP is upregulated in
patients with CAD and is a cardiovascular risk factor for the
severity of CAD and poor lipid control (46). Another study identified that the
recessive allele (TT vs. CT+CC) of SREBP1 rs9902941 is highly
expressed in patients with CAD compared with the normal control
(47). miRNA-33a and miRNA-33b are
intron miRNAs encoded by the intron regions of SREBP1 and SREBP2
genes (48). miRNA-122 affects CAD
by regulating genes associated with lipid synthesis and oxidation,
including SREBP, microsomal triglyceride transfer protein,
Krüppel-like factor 6 and cationic amino acid transporter 1
(49). Silencing of miRNA-122
leads to a decrease in total cholesterol and triglyceride in plasma
(49). At present, lncRNAs have
emerged as critical factors in the promotion of hepatic
gluconeogenesis and cholesterol biosynthesis via the SREBP pathway
(46). However, their functions in
CAD remain unknown, which highlights the importance of the results
of the present study.
In conclusion, a novel lncRNA ENST00000416361 was
first identified to be significantly upregulated in patients with
CAD by high-throughput sequencing in the present study. lncRNA
ENST00000416361 has potential as a biomarker for clinical screening
and diagnosis of CAD. The experimental results indicated that
ENST00000416361 had an association with 2 inflammatory cytokines
(IL-6 and TNF-α) and the lipid metabolism-associated genes (SREBP1
and SREBP2). These results suggested that ENST00000416361 may be
related to the pathogenesis of atherosclerosis, such as
inflammation and lipid metabolism. Nevertheless, CAD is often
accompanied by hypertension, and its occurrence and development are
also affected by numerous environmental factors. Although these
parameters are markedly associated with CAD, it is difficult to
independently evaluate the effect of each factor, therefore this
should be investigated in future studies. Therefore, the specific
mechanism underlying ENST00000416361 in the progression of CAD has
not been elucidated, which requires further exploration.
Acknowledgements
Not applicable.
Funding
This work was financially supported by the Special
Diagnosis Techniques for Clinical Key Diseases of Suzhou Municipal
Health and Family Planning Commission (grant no. LCZX201610),
Science and Technology Development Fund of Nanjing Medical
University (grant no. NMUB2018219) and the Suzhou Basic Research in
Medical and Health Application Grant (grant no. sys2018090).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
KS designed and funded this study. PL was the
principal experimenter and drafted the manuscript. XY assisted the
experiment and provided statistical analysis. QC designed the study
and revising this manuscript critically for important intellectual
content. GX, JW, and ZP performed the collection and interpretation
of data. JY, ML and LY conducted literature search and data
interpretation. All authors reviewed the results and approved the
final version of the manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Nanjing Medical University. Informed consent was obtained from the
patients and their families.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Pagliaro BR, Cannata F, Stefanini GG and
Bolognese L: Myocardial ischemia and coronary disease in heart
failure. Heart Fail Rev. 25:53–65. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sayols-Baixeras S, Luis-Ganella C, Lucas G
and Elosua R: Pathogenesis of coronary artery disease: focus on
genetic risk factors and identification of genetic variants. Appl
Clin Genet. 7:15–32. 2014.PubMed/NCBI
|
|
3
|
Glass CK and Witztum JL: Atherosclerosis.
The road ahead. Cell. 104:503–516. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dalen JE, Alpert JS, Goldberg RJ and
Weinstein RS: The epidemic of the 20(th) century: Coronary heart
disease. The American journal of medicine. 127:807–812. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dai X, Wiernek S, Evans JP and Runge MS:
Genetics of coronary artery disease and myocardial infarction.
World J Cardiol. 8:1–23. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Meseure D, Drak Alsibai K, Nicolas A,
Bieche I and Morillon A: Long noncoding RNAs as new architects in
cancer epigenetics, prognostic biomarkers, and potential
therapeutic targets. Biomed Res Int. 2015:3202142015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Esteller M: Non-coding RNAs in human
disease. Nat Rev Genet. 12:861–874. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hueso M, Cruzado JM, Torras J and Navarro
E: ALUminating the path of atherosclerosis progression:
Chaos theory suggests a role for Alu repeats in the
development of atherosclerotic vascular disease. Int J Mol Sci.
19:E17342018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang F, Xue X, Bi J, Zheng L, Zhi K, Gu Y
and Fang G: Long noncoding RNA CCAT1, which could be activated by
c-Myc, promotes the progression of gastric carcinoma. J Cancer Res
Clin Oncol. 139:437–445. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yuan JH, Yang F, Wang F, Ma JZ, Guo YJ,
Tao QF, Liu F, Pan W, Wang TT, Zhou CC, et al: A long noncoding RNA
activated by TGF-beta promotes the invasion-metastasis cascade in
hepatocellular carcinoma. Cancer Cell. 25:666–681. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Leung A and Natarajan R: Noncoding RNAs in
vascular disease. Curr Opin Cardiol. 29:199–206. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bitarafan S, Yari M, Broumand MA,
Ghaderian SM, Rahimi M, Mirfakhraie R, Azizi F and Omrani MD:
Association of increased levels of lncRNA H19 in PBMCs with risk of
coronary artery disease. Cell J. 20:564–568. 2019.PubMed/NCBI
|
|
13
|
Zhang Z, Gao W, Long QQ, Zhang J, Li YF,
Liu DC, Yan JJ, Yang ZJ and Wang LS: Increased plasma levels of
lncRNA H19 and LIPCAR are associated with increased risk of
coronary artery disease in a Chinese population. Sci Rep.
7:74912017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu D, Tang C, Liu P, Qian W and Sheng L:
Withdrawal notice: Targeting lncRNAs for cardiovascular
therapeutics in coronary artery disease. Curr Pharm Des. Jan
8–2018.(Epub ahead of print). doi:
10.2174/1381612824666180108120727. View Article : Google Scholar
|
|
15
|
Wu G, Cai J, Han Y, Chen J, Huang ZP, Chen
C, Cai Y, Huang H, Yang Y, Liu Y, et al: LincRNA-p21 regulates
neointima formation, vascular smooth muscle cell proliferation,
apoptosis, and atherosclerosis by enhancing p53 activity.
Circulation. 130:1452–1465. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tang SS, Cheng J, Cai MY, Yang XL, Liu XG,
Zheng BY and Xiong XD: Association of lincRNA-p21 haplotype with
coronary artery disease in a Chinese Han population. Dis Markers.
2016:91097432016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cai Y, Yang Y, Chen X, Wu G, Zhang X, Liu
Y, Yu J, Wang X, Fu J, Li C, et al: Circulating ‘lncRNA
OTTHUMT00000387022’ from monocytes as a novel biomarker for
coronary artery disease. Cardiovasc Res. 112:714–724. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang Y, Cai Y, Wu G, Chen X, Liu Y, Wang
X, Yu J, Li C, Chen X, Jose PA, et al: Plasma long non-coding RNA,
CoroMarker, a novel biomarker for diagnosis of coronary artery
disease. Clin Sci (Lond). 129:675–685. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wenger NK: 2011 ACCF/AHA focused update of
the guidelines for the management of patients with Unstable
Angina/Non-ST-Elevation Myocardial Infarction (updating the 2007
Guideline): highlights for the clinician. Clin Cardiol. 35:3–8.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jaatinen T and Laine J: Isolation of
mononuclear cells from human cord blood by Ficoll-Paque density
gradient. Curr Protoc Stem Cell Biol. Jun 1–2007.(Epub ahead of
print). doi: org/10.1002/9780470151808.sc02a01s1. View Article : Google Scholar
|
|
21
|
Jia Y, Xu H, Li Y, Wei C, Guo R, Wang F,
Wu Y, Liu J, Jia J, Yan J, et al: A Modified Ficoll-Paque gradient
method for isolating mononuclear cells from the peripheral and
umbilical cord blood of humans for biobanks and clinical
laboratories. Biopreserv Biobank. 16:82–91. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Trapnell C, Hendrickson DG, Sauvageau M,
Goff L, Rinn JL and Pachter L: Differential analysis of gene
regulation at transcript resolution with RNA-seq. Nat Biotechnol.
31:46–53. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The Gene
Ontology Consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
The Gene Ontology Consortium, . The Gene
Ontology Resource: 20 years and still GOing strong. Nucleic Acids
Res. 47:D330–D338. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang M, Zhong Z, Lv M, Shu J, Tian Q and
Chen J: Comprehensive analysis of differentially expressed profiles
of lncRNAs and circRNAs with associated co-expression and ceRNA
networks in bladder carcinoma. Oncotarget. 7:47186–47200.
2016.PubMed/NCBI
|
|
26
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
Elife. 4:e050052015. View Article : Google Scholar :
|
|
27
|
Garcia DM, Baek D, Shin C, Bell GW,
Grimson A and Bartel DP: Weak seed-pairing stability and high
target-site abundance decrease the proficiency of lsy-6 and other
microRNAs. Nat Struct Mol Biol. 18:1139–1146. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Grimson A, Farh KK-H, Johnston WK,
Garrett-Engele P, Lim LP and Bartel DP: MicroRNA targeting
specificity in mammals: determinants beyond seed pairing. Mol Cell.
27:91–105. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
John B, Enright AJ, Aravin A, Tuschl T,
Sander C and Marks DS: Human MicroRNA targets. PLoS Biol. 2:e363.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Iliopoulos D, Drosatos K, Hiyama Y,
Goldberg IJ and Zannis VI: MicroRNA-370 controls the expression of
microRNA-122 and Cpt1alpha and affects lipid metabolism. J Lipid
Res. 51:1513–1523. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hou S, Fang M, Zhu Q, Liu Y, Liu L and Li
X: MicroRNA-939 governs vascular integrity and angiogenesis through
targeting gamma-catenin in endothelial cells. Biochem Biophys Res
Commun. 484:27–33. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jaé N, Heumüller AW, Fouani Y and Dimmeler
S: Long non-coding RNAs in vascular biology and disease. Vascul
Pharmacol. 114:13–22. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ahmadi R, Heidarian E, Fadaei R, Moradi N,
Malek M and Fallah S: miR-342-5p Expression levels in coronary
artery disease patients and its association with inflammatory
cytokines. Clin Lab. 64:603–609. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fadaei R, Moradi N, Kazemi T, Chamani E,
Azdaki N, Moezibady SA, Shahmohamadnejad S, Fallah S, et al:
Decreased serum levels of CTRP12/adipolin in patients with coronary
artery disease in relation to inflammatory cytokines and insulin
resistance. Cytokine. 113:326–331. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Libby P, Tabas I, Fredman G and Fisher EA:
Inflammation and its resolution as determinants of acute coronary
syndromes. Circ Res. 114:1867–1879. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hammadah M, Sullivan S, Pearce B, Mheid
IA, Wilmot K, Ramadan R, Tahhan AS, O'Neal WT, Obideen M, Alkhoder
A, et al: Inflammatory response to mental stress and mental stress
induced myocardial ischemia. Brain Behav Immun. 68:90–97. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen H, Ding S, Liu X, Wu Y and Wu X:
Association of interleukin-6 genetic polymorphisms and environment
factors interactions with coronary artery disease in a Chinese Han
population. Clin Exp Hypertens. 40:514–517. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Guo F, Tang C, Li Y, Liu Y, Lv P, Wang W
and Mu Y: The interplay of LncRNA ANRIL and miR-181b on the
inflammation-relevant coronary artery disease through mediating
NF-kappaB signalling pathway. J Cell Mol Med. 22:5062–5075. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kuan YC, Hashidume T, Shibata T, Uchida K,
Shimizu M, Inoue J and Sato R: Heat shock protein 90 modulates
lipid homeostasis by regulating the stability and function of
sterol regulatory element-binding protein (SREBP) and SREBP
cleavage-activating protein. J Biol Chem. 292:3016–3028. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Brown MS and Goldstein JL: The SREBP
pathway: regulation of cholesterol metabolism by proteolysis of a
membrane-bound transcription factor. Cell. 89:331–340. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sato R: Sterol metabolism and SREBP
activation. Arch Biochem Biophys. 501:177–181. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Perez-Belmonte LM, Moreno-Santos I,
Cabrera-Bueno F, Sánchez-Espín G, Castellano D, Such M,
Crespo-Leiro MG, Carrasco-Chinchilla F, Alonso-Pulpón L,
López-Garrido M, et al: Expression of Sterol Regulatory
Element-Binding Proteins in epicardial adipose tissue in patients
with coronary artery disease and diabetes mellitus: Preliminary
study. Int J Med Sci. 14:268–274. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Abudesimu A, Adi D, Siti D, Xie X, Yang
YN, Li XM, Wang YH, Wang YT, Meng YJ, Liu F, et al: Association of
genetic variations in the lipid regulatory pathway genes FBXW7 and
SREBPs with coronary artery disease among Han Chinese and Uygur
Chinese populations in Xinjiang, China. Oncotarget. 8:88199–88210.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Price NL, Singh AK, Rotllan N, Goedeke L,
Wing A, Canfrán-Duque A, Diaz-Ruiz A, Araldi E, Baldán A, Camporez
JP, et al: Genetic ablation of miR-33 increases food intake,
enhances adipose tissue expansion, and promotes obesity and insulin
resistance. Cell Rep. 22:2133–2145. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Dong J, Liang YZ, Zhang J, Wu LJ, Wang S,
Hua Q and Yan YX: Potential role of lipometabolism-related
MicroRNAs in peripheral blood mononuclear cells as biomarkers for
coronary artery disease. J Atheroscler Thromb. 24:430–441. 2017.
View Article : Google Scholar : PubMed/NCBI
|