Introduction
Colorectal cancer (CRC) is recognized as one of the
most common and deadly cancers worldwide (1). It occupies an increasingly important
position in the spectrum of cancer, and represents the leading
cause of cancer-related mortality worldwide (2,3). Due
to the lack of specific clinical symptoms and early screening
tests, a substantial number of patients with CRC are first
diagnosed at an advanced stage with metastasis (4). Furthermore, even if a tumor is
resected, CRC recurrence and metastasis may occur after surgery;
therefore, the prognosis of CRC remains poor (5). Thus, it is of great importance to
understand the molecular mechanisms underlying CRC development and
to identify novel tumor-associated biomarkers.
Bioinformatics analysis provides a deeper and more
comprehensive understanding of targeted genes, and allows
researchers to identify functional genes for further analysis. The
Cancer Genome Atlas (TCGA) generates comprehensive,
multi-dimensional maps of the key genomic changes in cancer, and
provides researchers with a variety of genetic information on
different types of cancer (6).
Kyoto Encyclopedia of Genes and Genomes (KEGG) (7) and Gene Ontology (GO) (8) analyses have enabled the
identification of metabolic signaling pathways and major biological
functions of differentially expressed genes. Therefore, TCGA, data
collection, and GO and KEGG pathway enrichment analyses were
performed in this study.
Cell division cycle associated protein 7 (CDCA7) is
located on chromosome 2q31 and encodes a nuclear protein containing
371 amino acids (9). CDCA7 is a
cell division cycle-associated protein that was first discovered in
Myc-transfected fibroblasts, which is commonly overexpressed in
various types of human cancer (10). CDCA7 is periodically expressed
during the human cell cycle, with the highest expression level
found in the G1 to S phase (11).
Deregulation of cell cycle proteins often leads to the increased
risk of tumor occurrence (12,13).
Previous reports have demonstrated that CDCA7 is a c-Myc-responsive
gene that participates in cancer transformation and tumorigenesis
(9). Thus far, several studies
have reported that CDCA7 is overexpressed in different types of
cancer, including lymphoma, ovarian cancer, retinoblastoma, breast
cancer, acute myeloid leukemia and esophageal cancer (10,14–18).
These findings provide evidence that there may be an association
between CDCA7 and tumor development. However, to the best of our
knowledge, the relationship between CDCA7 and the development of
colorectal disease has not yet been reported on. Therefore, the
clinical relevance and underlying mechanisms of CDCA7 in the
occurrence and development of CRC remain largely unclear. The
present study aimed to investigate the expression levels and
clinical role of CDCA7 in CRC.
Materials and methods
Bioinformatics analysis
Collection of CDCA7 expression data from
TCGA
microRNA (miR/miRNA) expression matrix was retrieved
from TCGA (https://portal.gdc.cancer.gov). RNA-seq data from 647
CRC tissues (cancer group) and 51 normal adjacent tissues (normal
group) were downloaded from TCGA database (dataset nos. TCGA-COAD
and TCGA-READ). Differentially expressed levels of CDCA7 between
the cancer group and normal group were identified using the
Ballgown (https://github.com/alyssafrazee/ballgown) package with
the following criteria: P<0.05 and fold-change >2.
Gene functions and pathways
KEGG (http://www.genome.jp/kegg/pathway.html) and GO
(http://geneontology.org/) were used for the
enrichment analysis of dysregulated genes, in order to determine
the changes in functions and pathways associated with CRC. The
conditional criterion was P<0.05.
Research hotspots
According to the gene annotation in the database,
genetic studies were evaluated based on five different
perspectives: i) Relevant literature of CDCA7 (total number of
articles reported); ii) pathways (number of pathways that CDCA7 was
involved in according to the KEGG database); iii) functions (number
of functions that involved the CDCA7 gene according to the GO
database); iv) validated targeted miRNAs (number of miRNAs
associated with CDCA7 that have been published in literature); and
v) diseases (number of diseases associated with CDCA7 that have
been published in the literature), in order to identify research
hotspots of CDCA7 through the KEGG and GO databases, and published
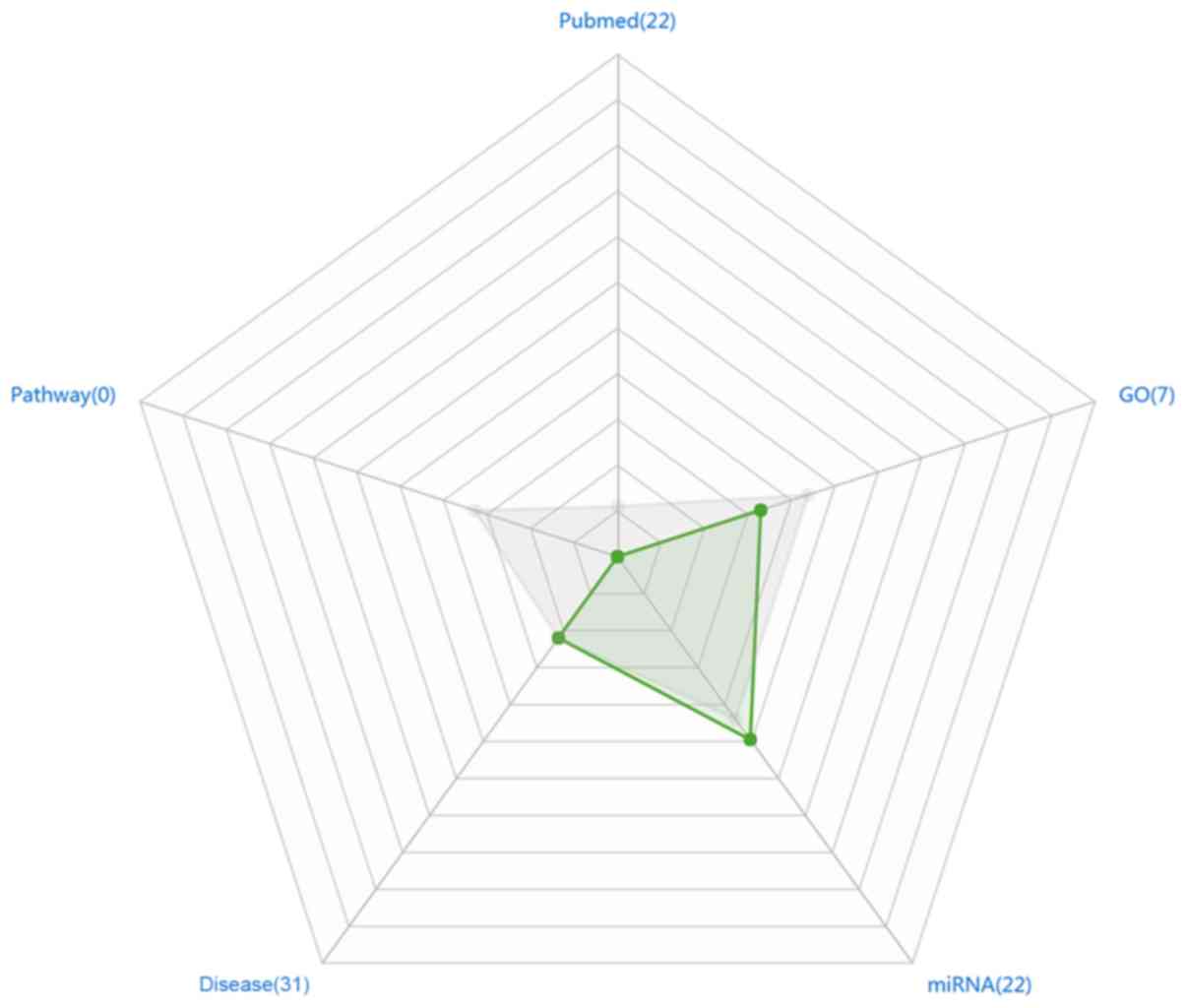
literature. A radar chart was produced using Microsoft Office 2013
software to demonstrate the research hotspots of CDCA7 (Fig. 1).
Disease association
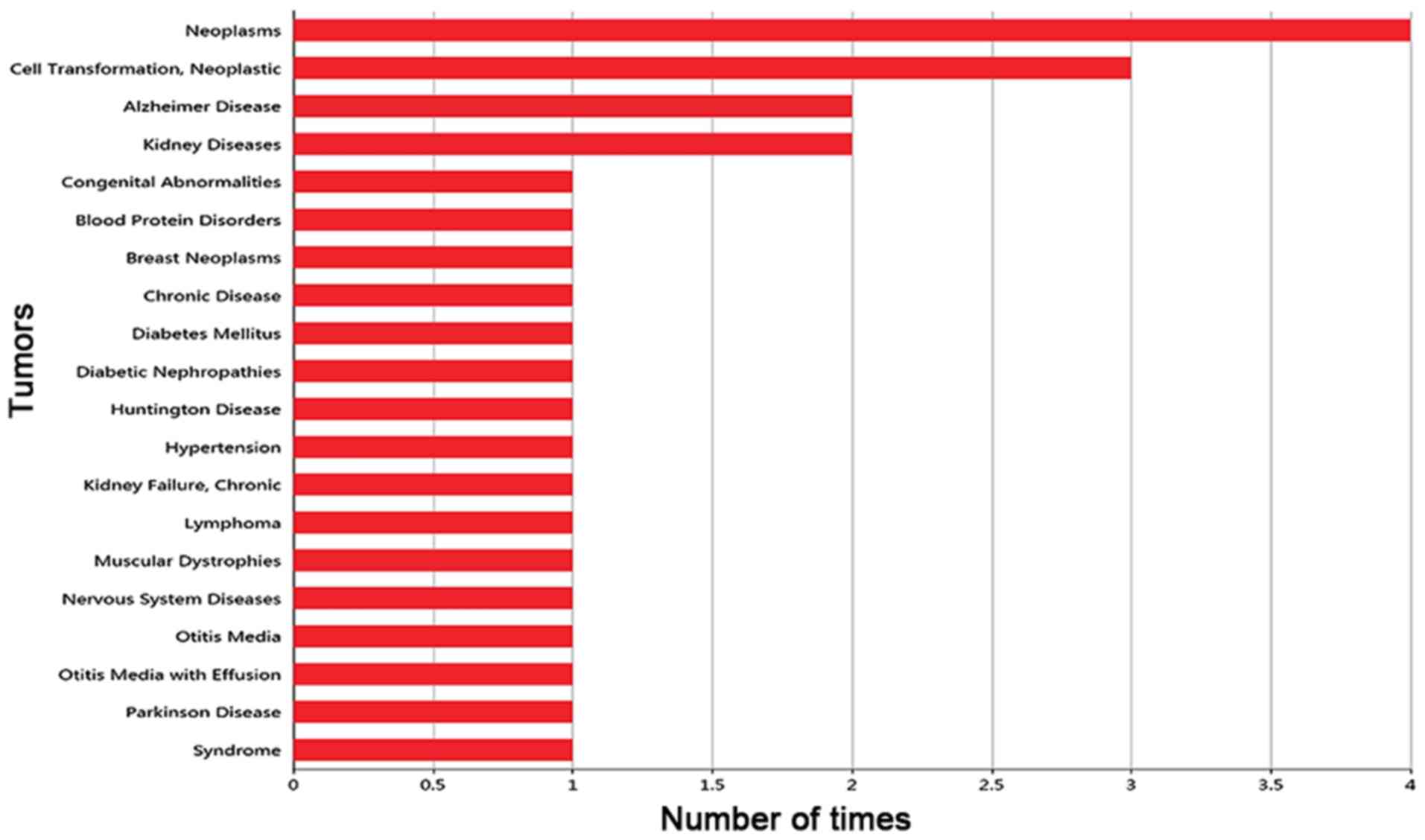
The frequency of the gene reported in different
diseases was mined from the PubMed database (https://www.ncbi.nlm.nih.gov/pubmed/) according to the
MeSH disease classification (https://www.ncbi.nlm.nih.gov/mesh). The top 20
diseases associated with the target gene were sought out.
Regulatory relationship
The regulatory relationship among genes was examined
based on four different perspectives: Gene-related transcription
factors, miRNAs, long non-coding (lnc)RNAs, as well as upstream and
downstream gene sequences, via published literature and the
databases of PubMed, MeSH and KEGG.
Transcription factor prediction
Transcription factors were predicted according to
the positional binding patterns. The integrated score based on the
two predicted scores in the TRANSFAC® database
(http://genexplain.com/transfac/#section3) were
assigned as the final predicted score. Binding regions of the
predicted transcription factors were annotated based on the DNA
methylation and single nucleotide polymorphism (SNP) information in
COSMIC (19) and dbSNP (https://www.ncbi.nlm.nih.gov/snp/) databases,
respectively. According to the predicted scores of the
TRANSFAC® database and whether the corresponding comment
information existed in the COSMIC database and the dbSNP database,
the recommendation degree was integrated. The recommendation degree
was used to represent the research value of the predicted
transcription factor; the higher the recommendation degree, the
better. Results with a high recommendation degree were selected for
use in the present study.
Gene expression in tumors
According to TCGA barcodes, all samples were
classified into normal and tumor tissues. Subsequently, the average
value and standard deviation of all samples were determined based
on the expression values in the normalized-results file of each
sample provided by RNA-SEQv2 data. Finally, the target gene
expression in human tumors was interpreted.
Tissue samples
CRC tissues and the paired adjacent non-tumor
tissues (n=104) in the form of pathological slices were collected
from patients (sex, 61 males and 43 females; age, 26–82 years;
average age, 56.42±12.87 years) who underwent surgical operation at
the Department of Colorectal-anal Surgery, The First Affiliated
Hospital of Guangxi Medical University (Guangxi, China) between
October 2012 and November 2013. In addition, CRC tissues were
collected from 15 patients (sex, 9 males and 6 females; age, 47–75
years; average age, 60.33±8.83 years) with pathological
confirmation of the diagnosis in November 2017. All tissue
specimens were collected within 30 min after surgical resection.
The 15 pairs of CRC tissues and normal adjacent tissues were
immediately snap-frozen in liquid nitrogen, and then stored at
−80°C until protein and RNA extraction. The survival status of
patients with CRC was verified by telephone interview. None of the
patients received pre-operative chemoradiotherapy or radiotherapy.
Clinicopathological characteristics of the patients were assessed
by reviewing patients' medical records and pathology reports.
The ethical approval for this study was obtained
from the Protection of Human Ethics Committee of The First
Affiliated Hospital of Guangxi Medical University. All specimens
were made anonymous and handled in accordance with the legal and
ethical standards for protecting human rights.
Cell lines and cell culture
Human normal colonic epithelial cell line NCM460 and
human CRC cell line SW620 were obtained from Wuhan Boster
Biological Technology, Ltd. Other human CRC cell lines, HT29,
HCT116 and RKO, were purchased from Shanghai R&S Biotechnology
Co., Ltd. These cell lines were cultured in DMEM (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with FBS (100 ml/l; Shanghai
ExCell Biology, Inc.) and maintained in 5% CO2 at 37°C.
Mycoplasma testing was then performed on all the cultured cell
lines. The four human CRC cell lines were referenced to the NCM460
cell line, which acted as the control group.
Immunohistochemistry
Immunohistochemistry was performed using SPlink
Detection kits (cat. no. SP-9000; OriGene Technologies, Inc.),
according to the manufacturer's protocol. Tissue specimens were
fixed in 10% formalin for 48 h at room temperature. After
dehydration and paraffin embedding, all the tissue specimens were
cut into 4-µm sections. Specifically, at room temperature, colon
tissue sections were routinely dewaxed with xylene and dehydrated
in a descending alcohol series, prior to being incubated in 0.01 M
citrate buffer at 100°C for 6 min for antigen retrieval. Following
being cooled down, the sections were washed three times in PBS for
5 min each and the sections were then incubated with 3%
H2O2 at room temperature for 12 min.
Subsequently, after being washed thrice, sections were incubated
with goat serum (provided by the kit) at room temperature for 12
min. Following being blocked, the tissue sections were incubated
with CDCA7 polyclonal primary antibody (1:100; cat. no. PA5-52165;
Invitrogen; Thermo Fisher Scientific, Inc.) at 4°C overnight. The
following day, the incubated tissue sections were washed three
times with PBS after being left at room temperature for ≥30 min.
The primary antibody was replaced with PBS as a negative control.
Subsequently, the tissue sections were incubated with a
biotin-conjugated secondary antibody (provided by the kit) at room
temperature for 12 min. Subsequently, after being washed with PBS
three times, the tissue sections were incubated with horseradish
peroxidase-conjugated streptomycin avidin (provided by the kit) at
room temperature for 12 min. The sections were then repeatedly
immersed 3 times in PBS for 5 min each. After drying the sections
at room temperature, the sections were stained with DAB reagent for
6 min, counterstained with hematoxylin for 30 sec and sealed with
neutral gum. Significant yellow or yellow-brown granules appeared
in the cell membrane, cytoplasm or nucleus, which was regarded as
positive streptavidin peroxidase (SP) staining. Semi-quantitative
analysis was conducted by integrating the staining intensity and
the percentage of positive cells. The stained tissue sections were
observed under an Olympus optical microscope (magnification, ×400).
The values of staining index (0–12) were calculated by multiplying
the two scores obtained (intensity and percentage). The staining
intensities of the positive cells were categorized as follows: 0
point (negative); 1 point (weak); 2 points (moderate); and 3 points
(strong). A total of five different fields were randomly observed
for each slice, and the percentages of positively-stained cells
were categorized as follows: 0 points (<5%); 1 point (5-25%); 2
points (26-50%); 3 points (51-75%); and 4 points (>75%). Final
staining scores of 0–7 indicated low CDCA7 expression, whereas
staining scores of 8–12 represented high CDCA7 expression. Each
slide was independently examined by two experienced
pathologists.
RNA extraction, cDNA synthesis and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from 30 samples of fresh CRC
tissues and adjacent normal colorectal tissues, as well as CRC cell
lines using Eastep® Super Total RNA Extraction kit
(Promega Corporation), according to the manufacturer's protocol.
Subsequently, cDNA was synthesized by the GoScript™ Reverse
Transcription Mix, Random Primers kit (Promega Corporation). The
following RT temperature protocol was used: 5 min at 25°C, 60 min
at 42°C and 15 min at 70°C, prior to being maintained at 4°C.
RT-qPCR was conducted with Applied Biosystems™ 7500 Real-Time PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.) using a
GoTaq® qPCR Master Mix kit (Promega Corporation). qPCR
cycling conditions were as follows: 10 min at 95°C, followed by 40
cycles of 15 sec at 95°C and 60 sec at 60°C, and a dissociation
cycle consisting of 15 sec at 95°C, 15 sec at 60°C and 15 sec at
95°C (ramping up at 0.2°C/sec). The expression levels of CDCA7 were
measured by 2−ΔΔCq method (20) using GAPDH as a standard reference,
and the relative expression value was set to 1 under unstimulated
conditions. The primer sequences (Takara Bio, Inc.) of GAPDH and
CDCA7 are presented in Table
I.
 | Table I.Primer sequences used for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences used for reverse
transcription-quantitative PCR.
| Gene | Primer sequences
(5′→3′) |
|---|
| GAPDH | F:
GCACCGTCAAGGCTGAGAAC |
|
| R:
TGGTGAAGACGCCAGTGGA |
| CDCA7 | F:
CCAGGCTCCGACTCACAATCAAG |
|
| R:
GTACTTATCCTCTTCCTCCTCCTCCTC |
Western blot analysis
Total protein was isolated from CRC tissues, normal
colorectal tissues and CRC cell lines using RIPA lysis buffer
containing 1% phosphatase inhibitor and 1% protease inhibitor.
After incubation at 0°C for 30 min, the lysed cells were washed
twice with ice-cold PBS, and then centrifuged at 12,000 × g and 4°C
for 15 min to eliminate potential cell debris. Total protein
concentration was determined by a bicinchoninic acid assay
(Beyotime Institute of Biotechnology). Subsequently, the quantified
proteins (50 µg) were denatured and subjected to SDS-PAGE on 12%
gels, and transferred onto 0.22 µm PVDF membranes. The membranes
were then blocked with non-fat milk (5%) for 1 h at 37°C. After
washing with Tris-buffered saline containing 5% Tween-20 (TBST)
three times, the membranes were incubated with rabbit polyclonal
CDCA7 antibody (1:250; cat. no. PA5-52165; Invitrogen; Thermo
Fisher Scientific, Inc.) and mouse monoclonal GAPDH antibody
(1:2,000; cat. no. 60004-1-lg; ProteinTech Group, Inc.) antibodies.
After incubation at 4°C overnight, the membranes were incubated
with IRDye® 680RD goat anti-rabbit immunoglobulin G
(IgG; 1:10,000; cat. no. 925-68071; LI-COR Biosciences) and goat
anti-mouse IgG (1:5,000; cat. no. BA1038; Wuhan Boster Biological
Technology, Ltd.) secondary antibodies, respectively, at room
temperature for 1 h. After washing with TBST three times, the
stained membranes were visualized using an Odyssey® CLx
imaging system (LI-COR Biosciences). The gray value of each protein
was determined by ImageJ version 1.4.3 software (National
Institutes of Health). GAPDH expression was used as the internal
reference. The relative expression level of CDCA7 was calculated by
the ratio of CDCA7 gray value to GAPDH gray value.
Statistical analysis
All analyses were carried out using SPSS Statistics
version 20.0 (IBM Corp.). Each experiment was repeated three times.
Student's paired t-test was performed to compare the expression
levels of CDCA7 between CRC tissues and adjacent normal colorectal
tissues. Expression levels of CDCA7 in CRC cell lines were compared
using a one-way ANOVA and Bonferroni's post hoc test. The strength
of association between CDCA7 expression and CRC clinicopathological
features was determined using a χ2 test or Fisher's
exact test. Overall survival data were analyzed by the Kaplan-Meier
method and log-rank test. Data are presented as the means ± SD.
P<0.05 was regarded as statistically significant.
Results
Bioinformatics prediction of CDCA7
mRNA expression in CRC
Based on the data obtained from TCGA database, under
the criteria of P<0.05 and |log2 fold change|>2,
the mRNA expression of CDCA7 was upregulated in CRC tissues
compared to normal tissues (P<0.05), and the fold change (T/N)
was 3.06.
GO and KEGG pathway enrichment
analysis
The biological functions of CDCA7 in CRC were
determined by GO and KEGG pathway enrichment analyses. As shown in
Table II, according to GO term
analysis, it was noted that the most marked functions of CDCA7 in
CRC were apoptosis and transcription. However, no significant
finding was obtained for gene-involved pathways, as revealed by
KEGG pathway analysis.
| Table II.Gene function and pathway analysis of
CDCA7. |
Table II.
Gene function and pathway analysis of
CDCA7.
| Gene ID | GO function |
|---|
| GO:0006351 | Regulation, DNA
template |
| GO:0006355 | Transcriptional
regulation, DNA template |
| GO:0006915 | Apoptosis |
| GO:0042127 | Regulates cell
proliferation |
Research hotspots of CDCA7
As shown in Fig. 1,
a total of 22 articles were reported on CDCA7, in which CDCA7 was
associated with seven biological GO functions. However, according
to the KEGG database, no signaling pathway involving CDCA7 was
found. The total number of diseases associated with CDCA7 was 31,
and 22 different targeted miRNAs had been validated. Only 2
(disease and miRNA) of the 5 indicators of CDCA7 were higher than
average. Thus, the lack of findings surrounding CDCA7 indicated
that little research has been performed on this gene until
relatively recently.
Gene-disease association
Based on the PubMed database, the frequency with
which CDCA7 was reported to be associated with disease was
collected, and the histogram of the top 20 diseases associated with
CDCA7 is shown in Fig. 2. Notably,
CDCA7 was most commonly reported in neoplasms, but its association
with CRC has not yet been reported. Hence, there is still more
research that is needed to investigate the expression and
significance of CDCA7 in CRC.
Regulatory relationships
As shown in Table
III, there were numerous regulatory factors associated with
CDCA7, such as hsa-let-7b-5p and human leukocyte antigen (HLA)
complex P5 (HCP5), indicating that this gene may act as a
transcriptional regulator.
| Table III.miRNAs and lncRNAs targeted by cell
division cycle associated protein 7. |
Table III.
miRNAs and lncRNAs targeted by cell
division cycle associated protein 7.
| miRNA | lncRNA |
|---|
| hsa-let-7b-5p | HCP5 |
| hsa-miR-124-3p | TP53TG1 |
| hsa-miR-124-3p | KCNQ1OT1 |
| hsa-miR-1254 | DKFZP434K028 |
| hsa-miR-1254 | DKFZP434K028 |
|
hsa-miR-1271-3p | LINC00341 |
|
hsa-miR-1271-3p | BIN3-IT1 |
|
hsa-miR-193b-3p | DLGAP1-AS2 |
| hsa-miR-24-3p | LOC90768 |
| hsa-miR-299-5p | BAALC-AS2 |
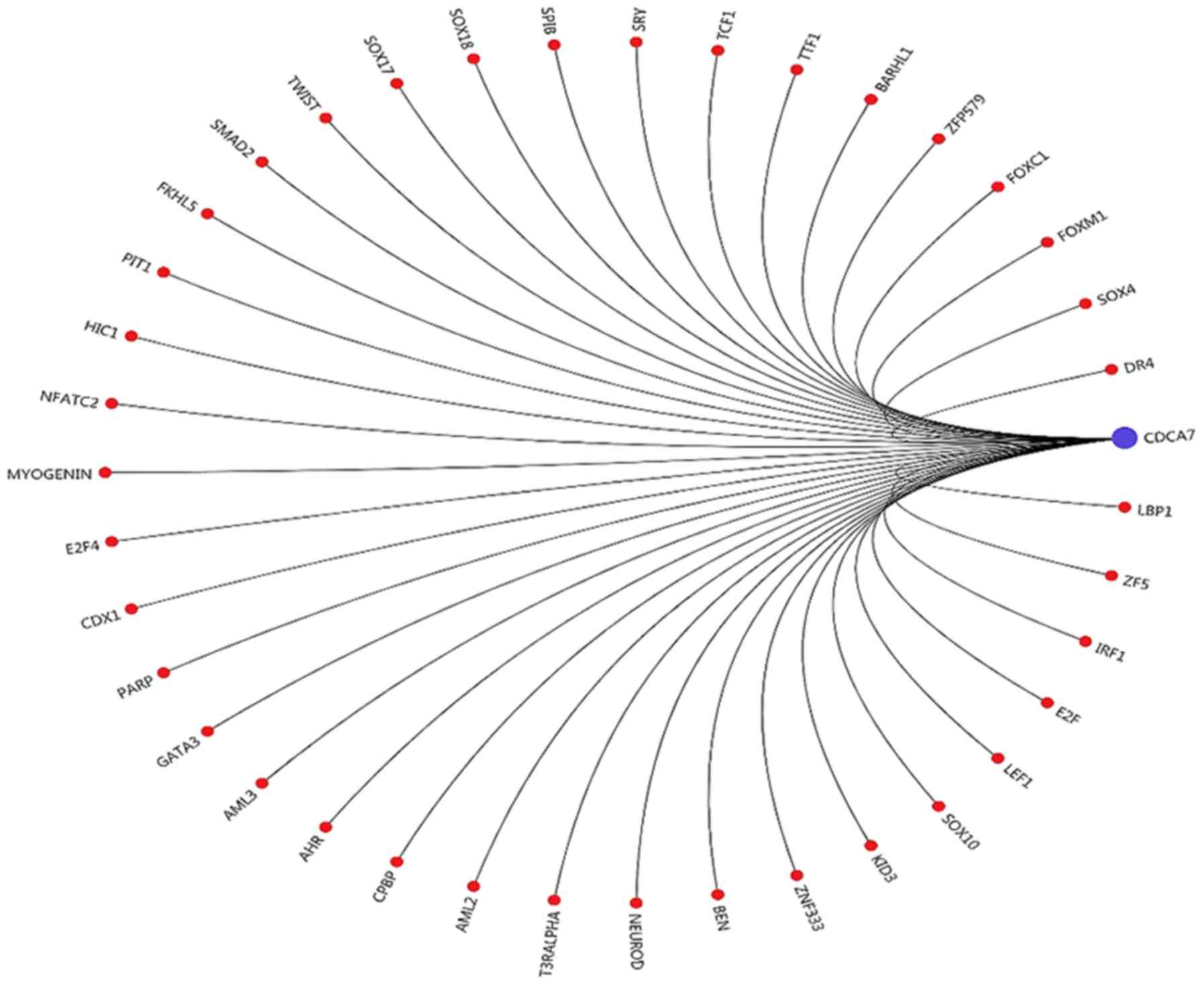
Transcription factor prediction
The transcription factors bound to the promoter
region of CDCA7 were predicted based on their positional binding
relationship. It was identified that upstream-binding protein 1
(LBP1), zinc finger 5 (ZF5) and interferon regulatory factor 1
(IRF1) were closely associated with CDCA7 (Fig. 3).
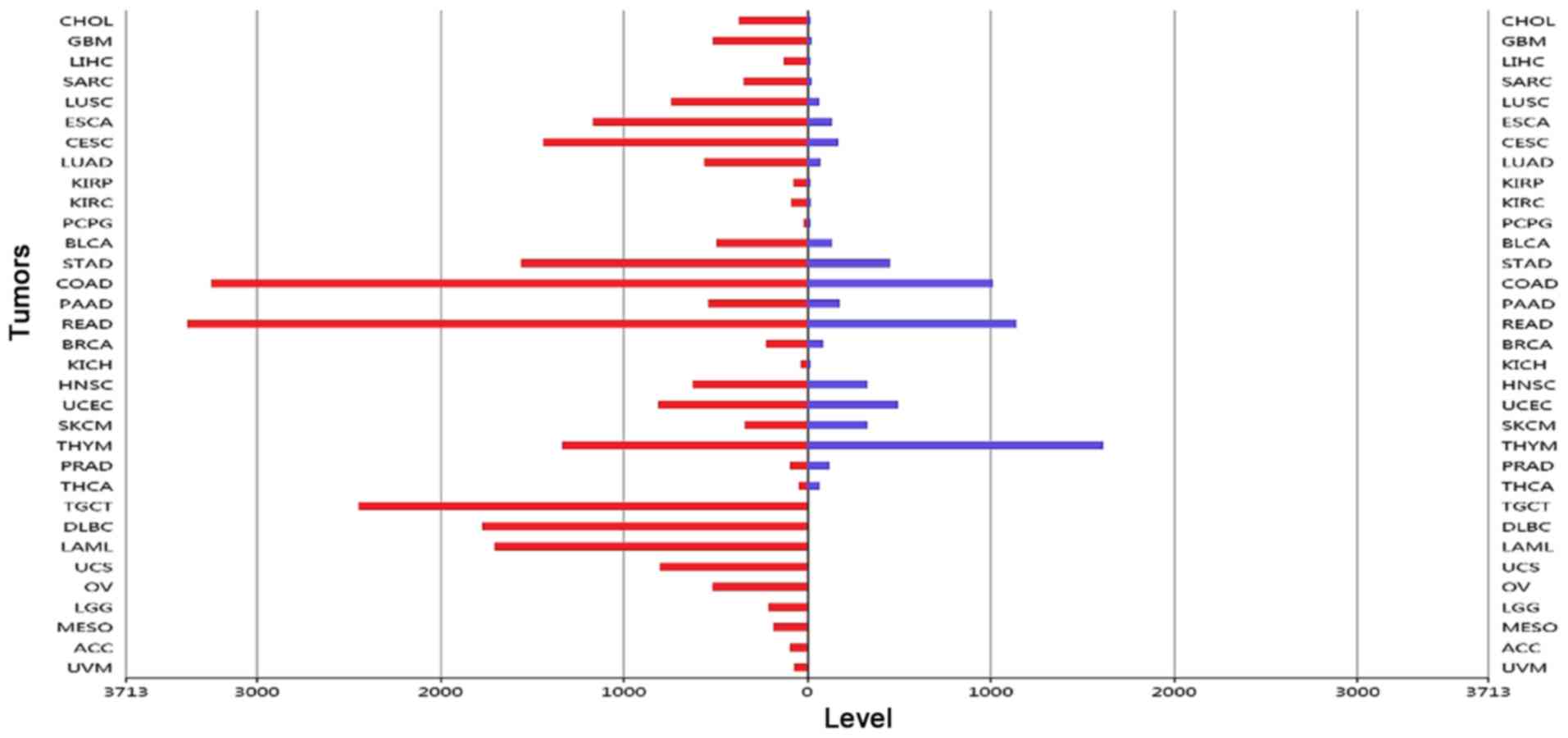
Gene expression in tumors
The expression levels of CDCA7 in each sample were
analyzed by the RNA-SEQv2 data derived from TCGA database. The
results showed that CDCA7 expression was mainly upregulated in
tumor tissues compared to adjacent normal tissues (Fig. 4).
High expression of CDCA7 in CRC
tissues
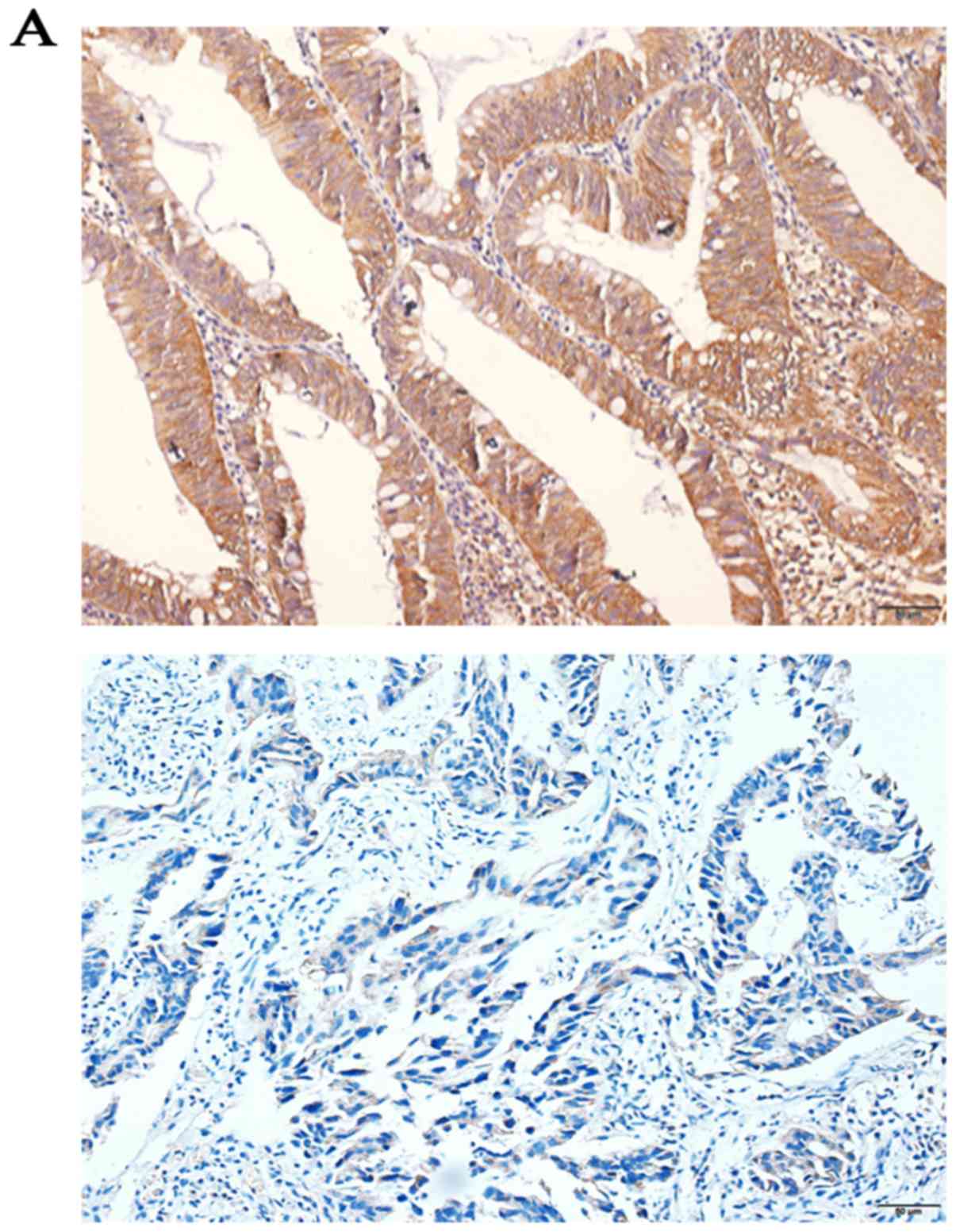
Immunohistochemistry SP staining revealed that the
expression levels of CDCA7 were higher in CRC tissues (75.96%)
compared with in adjacent normal colorectal tissues (26.92%;
P<0.0001; Fig. 5A). In
addition, the RT-qPCR results indicated that the expression levels
of CDCA7 were significantly upregulated among 15 CRC tissues
compared to adjacent normal colorectal tissues (P=0.044; Fig. 5B). Furthermore, western blotting
revealed that similar increased levels of CDCA7 protein were
observed in 15 CRC tissues (P=0.016; Fig. 5C and D).
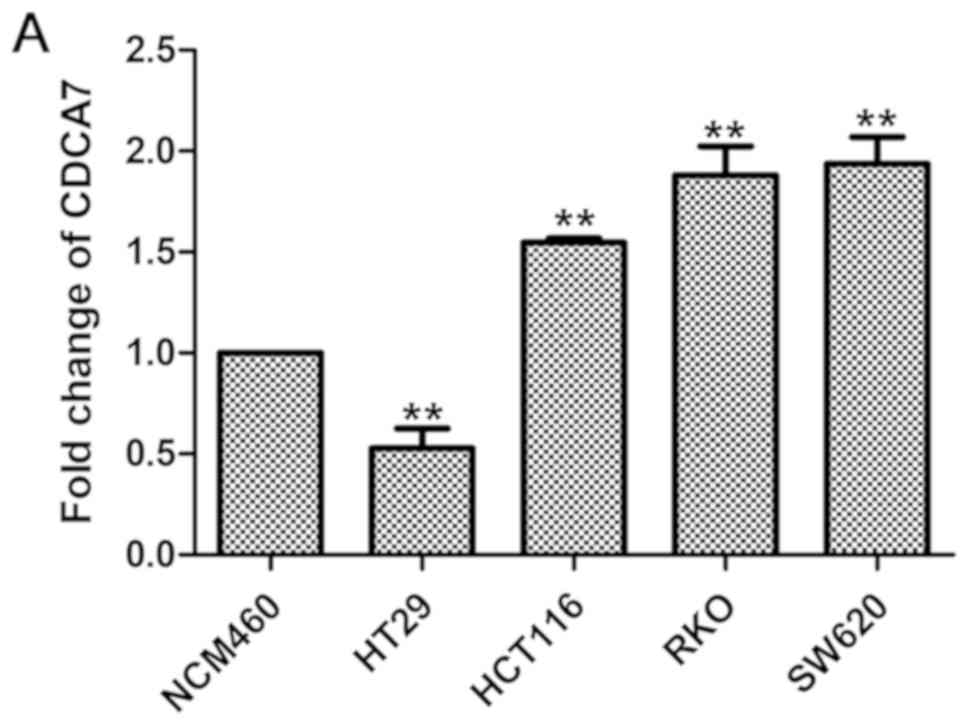
High expression of CDCA7 in CRC cell
lines
CDCA7 expression levels were relatively higher in
CRC cell lines compared to NCM460 (a normal colonic epithelial cell
line), as revealed by RT-qPCR (Fig.
6A) and western blotting (Fig. 6B
and C) analyses, suggesting that CDCA7 serves a role in
CRC.
CDCA7 expression is associated with
the clinical progression of CRC
The results of RT-qPCR and western blotting
indicated that CDCA7 expression levels were significantly increased
in human CRC. To further examine the potential association between
CDCA7 expression and CRC clinicopathological features, the
expression levels of CDCA7 were detected in 104 CRC tissues and
their paired adjacent non-tumor tissues via immunohistochemistry
staining. The positive expression rates of CDCA7 were 26.92%
(28/104) and 75.96% (79/104) in normal and CRC tissues,
respectively, with a statistically significant difference
(P<0.0001) (data not shown). CDCA7 expression levels in 104
pairs of CRC tissues were categorized into high-expression (n=79)
and low-expression (n=25) groups. As summarized in Table IV, high expression levels of CDCA7
were significantly associated with CRC invasion depth (P=0.002),
lymph node metastasis (P=0.038), tumor-node-metastasis stage
(P=0.001) and distant metastasis (P=0.019). However, no significant
differences were found in sex (P=0.533), age (P=0.369), tumor size
(P=0.217) and CRC differentiation (P=0.318) between high and low
CDCA7 expression groups. Collectively, these results indicated that
CDCA7 overexpression may be involved in the progression of CRC.
| Table IV.Association between CDCA7 expression
and clinicopathological characteristics of patients with CRC. |
Table IV.
Association between CDCA7 expression
and clinicopathological characteristics of patients with CRC.
|
|
| CDCA7 |
|
|---|
|
|
|
|
|
|---|
| Clinicopathological
characteristics | Number of
cases | Low | High | P-value |
|---|
| Sex |
|
|
| 0.533 |
|
Male | 61 | 16 | 45 |
|
|
Female | 43 | 9 | 34 |
|
| Age (years) |
|
|
| 0.369 |
|
<60 | 58 | 12 | 46 |
|
|
≥60 | 46 | 13 | 33 |
|
| Tumor size |
|
|
| 0.217 |
| <5
cm | 64 | 18 | 46 |
|
| ≥5
cm | 40 | 7 | 33 |
|
| Invasion depth |
|
|
| 0.002 |
|
Submucosal | 17 | 9 | 8 |
|
| Below
the muscle layer | 87 | 16 | 71 |
|
|
Differentiation |
|
|
| 0.318 |
|
High | 12 | 3 | 9 |
|
|
Moderate | 78 | 21 | 57 |
|
|
Poor | 14 | 1 | 13 |
|
| TNM stage |
|
|
|
|
|
I–II | 54 | 20 | 34 | 0.001 |
|
III–IV | 50 | 5 | 45 |
|
| Lymph node
metastasis |
|
|
| 0.038 |
|
Negative | 65 | 20 | 45 |
|
|
Positive | 39 | 5 | 34 |
|
| Distant
metastasis |
|
|
| 0.019 |
|
Negative | 89 | 25 | 64 |
|
|
Positive | 15 | 0 | 15 |
|
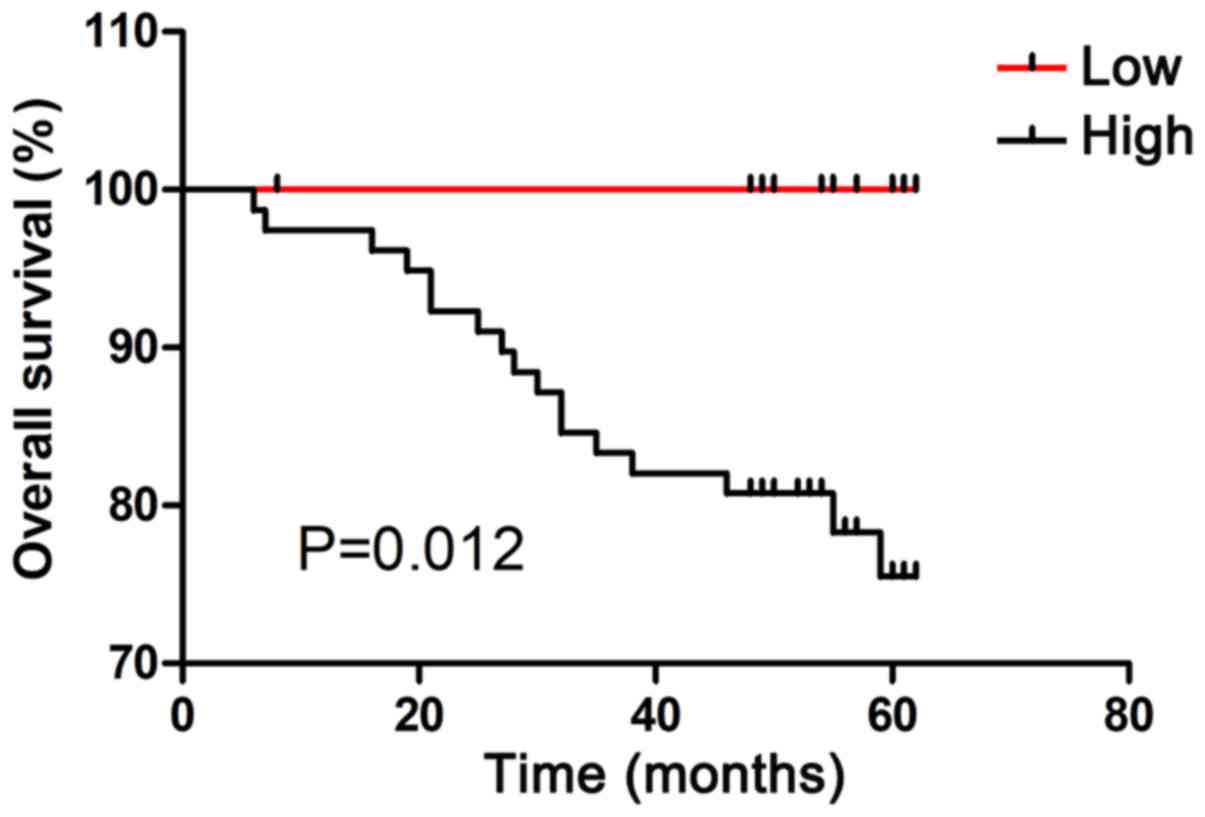
High expression of CDCA7 predicts poor
prognosis in patients with CRC
The association between CDCA7 expression and overall
survival in patients with CRC was evaluated by Kaplan-Meier
analysis. As shown in Fig. 7, high
CDCA7 expression group was significantly associated with shorter
survival times as compared with the low CDCA7 expression group
(P=0.012).
Discussion
CRC is the most common form of gastrointestinal
cancer, and one of the leading causes of cancer-related mortality
globally (21,22). Although clinicopathological
parameters have been used to predict the clinical outcome of CRC in
patients, these classification indicators are not precise enough to
predict the prognosis of patients with CRC. Therefore, it is
essential to identify novel tumor biomarkers and therapeutic
targets for early diagnosis and treatment of CRC. This study
focused on determining the expression levels and clinical role of
CDCA7 in CRC.
CDCA7 is a family member of the cell division cycle
proteins, which are mainly expressed in the cell nucleus (9). In the present study, the results of
immunohistochemical staining demonstrated that CDCA7 was expressed
primarily in the nucleus, and at least partly in cytoplasm. A
previous study has demonstrated that CDCA7 is a Myc target gene,
which participates in Myc-mediated tumor transformation and
eventually leads to tumor occurrence (9). Mitotic stimulation can induce the
activation of Myc to promote cell cycle progression (23,24).
Moreover, CDCA7 is positively expressed in solid tumors, along with
high Myc levels (17). These
findings indicated that CDCA7 may promote tumor growth progression
with Myc. In a previous study, CDCA7 was identified as a novel
transcription factor E2F transcription factor 1 (E2F1)-induced
protein, suggesting that the expression of CDCA7 is activated by
both E2F1 and Myc (25).
Additionally, other studies have shown that Myc and E2F1 can bind
to the CDCA7 promoter in order to upregulate the expression levels
of CDCA7 (9,25). These findings suggest that CDCA7
may participate in tumor occurrence via the activation of Myc and
E2F1. However, this statement needs further experimental
verification.
It is worth noting that disordered cell cycle
control is a basic feature of tumor pathogenesis, and there is a
large imbalance in cyclic protein regulation in tumor cells
(26–28). High CDCA7 expression has been
detected in different types of human cancer, indicating that CDCA7
may function as an oncogene. For example, Jiménez-P et al
(14) demonstrated that CDCA7
protein is upregulated in Burkitt lymphoma cell lines and tumor
tissues, and CDCA7 mRNA levels are significantly elevated in
numerous T and B lymphoma cell lines. In addition, a previous study
reported that CDCA7 is overexpressed in the YDOV-151 human ovarian
cancer cell line (>7-fold expression) compared with in human
ovarian surface epithelial cells (10). Moreover, CDCA7 has been reported to
be involved in the occurrence of retinoblastoma, which can be
utilized as a biomarker for early diagnosis and treatment of the
disease (15). Cheng et al
(18) demonstrated that the
expression level of CDCA7 is higher in esophageal squamous cell
carcinoma compared to normal esophageal tissue. Furthermore, Osthus
et al (17) reported that
CDCA7 is overexpressed in patients with acute myeloid leukemia,
leading to an increased risk of lymphoid malignancies in these
patients. Overall, CDCA7 is upregulated in a wide variety of human
tumors, and is likely to be associated with cancer progression.
Additionally, previous evidence has suggested that
CDCA7 is involved in the proliferation and apoptosis of tumor cells
(29). Recently, it has been shown
that the lncRNA FGD5-AS1 can promote the proliferation, migration
and invasion of CRC cells by upregulating CDCA7 via sponging
miR-302e (30). This study also
found that FGD5-AS1 can competitively bind with miR-302e to
modulate CDCA7, resulting in the induction of CRC cell apoptosis
(30). This indicated that CDCA7
exhibits a transcriptional regulatory function and by being
modulated by its upstream target lncRNA, it can affect the
progression of CRC. In the present study, GO term analysis revealed
that CDCA7 was related to cell proliferation and apoptosis. While a
recent report has focused on the molecular mechanisms of CDCA7 and
CRC (30), the exact relationship
between CDCA7 and CRC still remains largely unknown. Further
research into this relationship is needed in the future.
The results of the present study indicated that
CDCA7 expression was upregulated in human CRC tissues compared with
in adjacent normal tissues. In addition, it was shown that high
CDCA7 expression could contribute to advanced tumor progression in
patients with CRC. The results of Kaplan-Meier analysis
demonstrated that different expression levels of CDCA7 exhibited
significant effects on the prognosis of patients with CRC
(P=0.012). Hence, CDCA7 could be a reliable marker for predicting
tumor progression and survival prognosis in patients with CRC, but
further studies are needed in order to validate this. However, this
study is limited by the lack of clarification on the specific
mechanisms underlying the positive association between CDCA7 and
CRC progression. Therefore, further research and clinical trials
into CDCA7 are needed in the future.
In conclusion, the results of the present study
provided evidence that CDCA7 may be highly expressed in CRC tissues
and may be associated with advanced tumor progression. Notably, to
the best of our knowledge, this is the first study to investigate
the expression of CDCA7 in CRC tissues and cell lines, as well as
its relationship with the clinical parameters of tumor progression.
In addition, these results demonstrated that high CDCA7 expression
contributes to poor prognosis in patients with CRC. Taken together,
these results suggested that CDCA7 may serve as a novel biomarker
for CRC diagnosis and a reference indicator for the prognosis of
patients with CRC.
Acknowledgements
The authors would like to thank Dr S.Q. Liu, Dr Y.J.
Su, Mr. T,Y. Zhang and Mr. Y. Zhu at the Department of
Gastroenterology, The Second Affiliated Hospital of Guangxi Medical
University for their help and excellent technical support.
Funding
The current study was funded by the National Nature
Science Foundation of China (grant no. 81760516) and the Program
for Improvement Scientific Research Ability of Young and
Middle-Aged Teachers of Higher Education of Guangxi (grant no.
2017KY0093).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CL designed the study and revised the manuscript. MQ
and JZ contributed to sample collection and performed the
bioinformatics analysis. SL and JH performed the experiments, data
analysis and wrote the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
Informed consent was obtained from patients. The
present study was approved by the Protection of Human Ethics
Committee of the First Affiliated Hospital of Guangxi Medical
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Freddie B, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chiu HM, Hsu WF, Chang LC and Wu MH:
Colorectal cancer screening in Asia. Curr Gastroenterol Rep.
19:472017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Calon A, Espinet E, Palomo-Ponce S,
Tauriello DV, Iglesias M, Céspedes MV, Sevillano M, Nadal C, Jung
P, Zhang XH, et al: Dependency of colorectal cancer on a
TGF-β-driven program in stromal cells for metastasis initiation.
Cancer Cell. 22:571–584. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cui G, Cai F, Ding Z and Gao L: MMP14
predicts a poor prognosis in patients with colorectal cancer. Hum
Pathol. 83:36–42. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ogata H, Goto S, Sato K, Fujibuchi W, Bono
H and Kanehisa M: KEGG: Kyoto Encyclopedia of Genes and Genomes.
Nucleic Acids Res. 27:29–34. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Harris MA, Clark J, Ireland A, Lomax J,
Ashburner M, Foulger R, Eilbeck K, Lewis S, Marshall B, Mungall C,
et al: The Gene Ontology (GO) database and informatics resource.
Nucleic Acids Res. 32((Database Issue)): D258–D261. 2004.PubMed/NCBI
|
|
9
|
Prescott JE, Osthus RC, Lee LA, Lewis BC,
Shim H, Barrett JF, Guo Q, Hawkins AL, Griffin CA and Dang CV: A
novel c-Myc-responsive gene, JPO1, participates in neoplastic
transformation. J Biol Chem. 276:48276–48284. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cho H, Lim BJ, Kang ES, Choi JS and Kim
JH: Molecular characterization of a new ovarian cancer cell line,
YDOV-151, established from mucinous cystadenocarcinoma. Tohoku J
Exp Med. 218:129–139. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lewis BC, Shim H, Li Q, Wu CS, Lee LA,
Maity A and Dang CV: Identification of putative c-Myc-responsive
genes: Characterization of rcl, a novel growth-related gene. Mol
Cell Biol. 17:4967–4978. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Basu Baul TS, Longkumer I, Duthie A, Singh
P, Koch B and Guedes da Silva MFC: Triphenylstannyl
((arylimino)methyl) benzoates with selective potency that induce G1
and G2/M cell cycle arrest and trigger apoptosis via ROS in human
cervical cancer cells. Dalton Trans. 47:1993–2008. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang YC, Chang KC, Lin BW, Lee JC, Lai CH,
Lin LJ, Yen Y, Lin CS, Yang SJ, Lin PC, et al: The EGF/hnRNP Q1
axis is involved in tumorigenesis via the regulation of cell
cycle-related genes. Exp Mol Med. 50:1–14. 2018. View Article : Google Scholar
|
|
14
|
Jiménez-P R, Martín-Cortázar C, Kourani O,
Chiodo Y, Cordoba R, Domínguez-Franjo MP, Redondo JM, Iglesias T
and Campanero MR: CDCA7 is a critical mediator of lymphomagenesis
that selectively regulates anchorage-independent growth.
Haematologica. 103:1669–1678. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang QL, Chen X, Zhang MH, Shen QH and Qin
ZM: Identification of hub genes and pathways associated with
retinoblastoma based on co-expression network analysis. Genet Mol
Res. 14:16151–16161. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Albulescu R: Elevated cyclin B2 expression
in invasive breast carcinoma is associated with unfavorable
clinical outcome. BMC Cancer. 3:12013.
|
|
17
|
Osthus RC, Karim B, Prescott JE, Smith BD,
McDevitt M, Huso DL and Dang CV: The Myc target gene JPO1/CDCA7 is
frequently overexpressed in human tumors and has limited
transforming activity in vivo. Cancer Res. 65:5620–5627. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cheng C, Zhou Y, Li H, Xiong T, Li S, Bi
Y, Kong P, Wang F, Cui H, Li Y, et al: Whole-genome sequencing
reveals diverse models of structural variations in esophageal
squamous cell carcinoma. Am J Hum Genet. 98:256–274. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tate JG, Bamford S, Jubb HC, Sondka Z,
Beare DM, Bindal N, Boutselakis H, Cole CG, Creatore C, Dawson E,
et al: COSMIC: The catalogue of somatic mutations in cancer.
Nucleic Acids Res. 47:D941–D947. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bray F, Ferlay J, Soerjomataram I, Siegal
RL, Torre LA and Jemal A: Global Cancer Statistics 2018: GLOBOCAN
Estimates of Incidence and Mortality Worldwide for 36 Cancers in
185 Countries. CA Cancer J Clin. 68((6)): 394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
GBD 2015 Mortality and Causes of Death
Collaborators, . Global, regional, and national life expectancy,
all-cause mortality, and cause-specific mortality for 249 causes of
death, 1980–2015: a systematic analysis for the Global Burden of
Disease Study 2015. Lancet. 388((10053)): 1459–1544.
2016.PubMed/NCBI
|
|
23
|
Conover CA and Bale LK: Insulin-like
growth factor I induction of c-myc expression in bovine fibroblasts
can be blocked by antecedent insulin receptor activation. Exp Cell
Res. 238:122–127. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kitaura H, Shinshi M, Uchikoshi Y, Ono T,
Iguchi-Ariga SM and Ariga H: Reciprocal regulation via
protein-protein interaction between c-Myc and p21(cip1/waf1/sdi1)
in DNA replication and transcription. J Biol Chem. 275:10477–10483.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Goto Y, Hayashi R, Muramatsu T, Ogawa H,
Eguchi I, Oshida Y, Ohtani K and Yoshida K: JPO1/CDCA7, a novel
transcription factor E2F1-induced protein, possesses intrinsic
transcriptional regulator activity. Biochim Biophys Acta.
1759:60–68. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang J, Sun Z, Han Y, Yao R, Yue L, Xu Y
and Zhang J: Rnf2 knockdown reduces cell viability and promotes
cell cycle arrest in gastric cancer cells. Oncol Lett.
13:3817–3822. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee J, Choi BY and Keum YS: Acetonitrile
extract of Salvia miltiorrhizaRadix exhibits growth-inhibitory
effects on prostate cancer cells through the induction of cell
cycle arrest and apoptosis. Oncol Lett. 13:2921–2928. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lu J, Chen X, Qu S, Yao B, Xu Y, Wu J, Jin
Y and Ma C: Oridonin induces G2/M cell cycle arrest and
apoptosis via the PI3K/Akt signaling pathway in hormone-independent
prostate cancer cells. Oncol Lett. 13:2838–2846. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gill RM, Gabor TV, Couzens AL and Scheid
MP: The MYC-associated protein CDCA7 is phosphorylated by AKT to
regulate MYC-dependent apoptosis and transformation. Mol Cell Biol.
33:498–513. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li D, Jiang X, Zhang X, Cao G, Wang D and
Chen Z: Long noncoding RNA FGD5-AS1 promotes colorectal cancer cell
proliferation, migration, and invasion through upregulating CDCA7
via sponging miR-302e. In Vitro Cell Dev Biol Anim. 55:577–585.
2019. View Article : Google Scholar : PubMed/NCBI
|