Introduction
A wide variety of functional appliances, such as the
activator, twin-block and Herbst types, are used to correct Class
II skeletal and occlusal disharmonies in patients undergoing growth
and development (1). Functional
appliances are particularly popular for correcting Class II
malocclusion in growing children, as well as for improving
undesirable facial profiles (2,3).
Clinically, functional and fixed appliances are used to treat Class
II division 1 malocclusions characterized by lower jaw inadequacy,
allowing the mandible to extend forward in a fixed position to
stimulate mandibular growth (4).
Several studies have reported that functional appliances promote
movement of the teeth and help to achieve proper facial muscle
function (5,6). With the use of functional appliances,
the patient's neuromuscular and skeletal systems undergo adaptive
modifications. However, despite the widespread use of such
functional appliances, the precise mechanism of action responsible
for these modifications, as well as the skeletal and dental
effects, remain unclear. The mechanisms underlying tensile muscle
structural adaptability in response to functional instruments have
received a great deal of interest. Research has suggested that
skeletal muscle proliferation, differentiation, migration and
apoptosis occur as a result of significant levels of mechanical
stretch (7). Mechanical stretching
of skeletal muscle initiates a series of cellular responses that
can cause stem or progenitor cells to enter the cell cycle, divide,
differentiate and fuse with other cells to repair damaged areas
(8,9), or alternatively to undergo apoptosis
(10,11). Previous studies have focused on the
response of cell proliferation and differentiation to adaptive
mechanical stretching of muscle cells (12,13).
However, as accumulating evidence has revealed that apoptosis plays
a major role in the adaptation of skeletal muscle function, more
attention has been given to stretch-induced apoptosis (14). Therefore, elucidating the mechanism
by which myoblast apoptosis is induced by mechanical stress is
essential for improving the understanding of the adaptive
mechanisms of skeletal muscle function. This in turn could help to
maximize stress-induced skeletal muscle remodeling.
Apoptosis is a distinctive and physiologically
important mode of programmed cell death, and plays a crucial role
in homeostasis, normal development and elimination of potentially
pathological cells from the body (15,16).
One important trigger of the apoptotic response is overload
mechanical stretch (17). The
duration and intensity of mechanical stretching determine whether a
cell will survive the stress by adapting, or instead undergo
autophagy or apoptosis due to unacceptably high levels of stress.
There are three main apoptotic pathways: The death receptor
depentent pathway (extrinsic pathway), the mitochondrial-mediated
pathway (intrinsic pathway) and the endoplasmic reticulum-mediated
pathway (14). Previous evidence
has implicated the PI3K/Akt pathway in apoptosis (18). However, the involvement of PI3K/Akt
signaling in mechanically induced muscle apoptosis has not been
clearly demonstrated.
The PI3K/Akt pathway is a critical signaling pathway
that mediates a variety of cellular functions, such as survival,
proliferation, migration and differentiation (19). PI3K can be activated by a wide
range of environmental stimuli (20). After activation of PI3K,
phosphoinositide-dependent protein kinase-1 recruits Akt to the
cell membrane and activates it (21). The activation of Akt controls a
variety of biological responses, such as the stimulation of cell
proliferation and inhibition of apoptosis (22,23).
The mitochondria play an important role in the regulation of
apoptosis and necrotic death, and the opening of the mitochondrial
permeability transition pore (MPTP) is considered to be a critical
regulator of cell death. However, the specific mechanism underlying
this remains unclear (24). The
PI3K/Akt pathway is involved in MPTP opening during cardiomyocyte
apoptosis induced by oxidative stress (23). To date, however, research
addressing the role of the PI3K/Akt pathway in stretch-induced
apoptosis of myoblasts has been limited.
In the present study, rat L6 cells were used to
explore stretch-induced apoptosis in skeletal muscles and
investigate the involvement of the PI3K/Akt pathway. Furthermore, a
specific inhibitor of PI3K/Akt was used to investigate the role of
the PI3K/Akt pathway in this process. L6 cell structure,
ultrastructure and levels of apoptosis were measured by microscopy,
and protein expression levels were determined by western blotting
(WB).
Materials and methods
Cell culture
L6 rat myoblasts (The Cell Bank of Type Culture
Collection of the Chinese Academy of Sciences) were cultured in a
humidified incubator at 37°C in an atmosphere containing 5%
CO2. Cultures were grown in DMEM (HyClone; GE Healthcare
Life Sciences), containing 10% fetal bovine serum (HyClone; GE
Healthcare Life Sciences), 100 U/ml penicillin and 100 µg/ml
streptomycin. Growth medium was replaced every other day. When the
cells reached 80% confluence, they were digested with 0.25% trypsin
and passaged at a split ratio of 1:2. Cells were then seeded in
growth medium into each well of a 6-well plate (BioFlex; Flexcell
International Corporation) at 3×105 cells/well.
Stretch loading
L6 cells were suspended in growth medium, seeded at
a density of 3×105 cells/well in flexible bottom 6-well
plates and assigned to one of the following groups: Control, 2, 6,
12 and 24 h. Except for stretching, the control group was treated
in the same way as the mechanically stretched group. Cells in the
2, 6, 12 and 24 h groups were subjected to a cyclic strain of 15%
elongation at a frequency of 10 cycles/min for the indicated
periods of time on a computer-controlled vacuum stretch apparatus
(FX-4000T™ Tension Plus System; Flexcell International
Corporation). Each cycle consisted of a 3-sec stretching period
followed by a 3-sec relaxation period. Experiments were repeated
three times. Cell growth was examined and imaged with an inverted
microscope using at a magnification of ×10× (Eppendorf).
Hoechst staining assay
Cells were stained using the Hoechst Staining Kit
(Beyotime Institute of Biotechnology), Briefly, L6 rat skeletal
muscle myoblasts were fixed for 10 min at room temperature or
overnight at 4°C with 0.5 ml/well fixation buffer, and then washed
twice in PBS. After addition of Hoechst 33258 (5 ml/well), the
cells were placed at 37°C in the dark to stain for 5 min and then
washed twice again with PBS. Apoptotic cells were visualized by a
fluorescence microscope (Eppendorf). The apoptotic index was
calculated as the percentage of apoptotic nuclei per total nuclei
number per field.
Assessment of MPTP opening
MPTP opening was assessed by using the
calcein-AM/cobalt method. The cells were seeded in 6-well plates;
after cyclic stretching, the cells were washed twice with PBS and
stained with 5 µl calcein-AM working solution (Cell Stain Buffer
1:500 diluted 1mM Calcein AM stock solution) for 30 min at 37°C.
After two washes with PBS, the cells were analyzed by flow
cytometry (Epics XL; Beckman-Coulter). The flow cytometer showed
the proportion of apoptotic area (V1-L) and Calcein fluorescence
area (V1-R) of myoblast in each group.
WB analysis
After cyclic stretching, the cells were harvested in
RIPA lysis buffer [50 mM Tris (pH 7.4), 1 mM EDTA, 1% TritonX-100,
150 mM NaCl, 1 mM phenylmethanesulfonyl fluoride and 1X protease
inhibitor]. Lysates were centrifuged at 12,000 × g for 10 min at
4°C, and protein concentrations were determined using the
bicinchoninic acid method. Protein extracts (30 µl/well) were
separated on a 10% gel via SDS-PAGE and transferred to PVDF
membranes. Membranes were blocked with 10% instant non-fat dry milk
for 1 h at room temperature and then incubated with primary
antibody purchased from Cell Signaling Technology, Inc.: Akt
(1:1,000; rabbit; cat. no. 9331), phosphorylated (p)-Akt (1:1,000;
rabbit; cat. no. 9271), GSK-3β (1:1,000; rabbit; cat. no. 9315),
p-GSK-3β (1:1,000; rabbit; cat. no. 9336) and GAPDH (1:1,000;
rabbit; cat. no. 5174) overnight at 4°C. The membranes were washed
three times for 5 min each with 10% TBS- 0.1% Tween 20 and then
incubated with the goat anti-rabbit IgG-HRP secondary antibody
conjugated with horseradish peroxidase (1:2,000; rabbit; cat. no.
CW0103S, ComWin Biotech Co.) for 45–60 min at room temperature.
Protein bands on the membranes were visualized on an enhanced
chemiluminescence machine (Bio-Rad Laboratories, Inc.). and
analyzed using Quantity One software version 4.6.6.(Bio-Rad
Lbaoratories, Inc.)
Inhibition of PI3K/Akt signaling using
LY294002
To determine whether PI3K/Akt signaling was involved
in stretch-induced apoptosis, cells were treated with the PI3K/Akt
signaling inhibitor, LY294002 (ApexBio). Cells were treated with 20
µM LY294002 (2 µl/well) for 2 h prior to cyclic stretching.
Apoptosis was assessed by Hoechst staining and opening of the MPTP,
and levels of Akt, phosphorylated (p)-Akt, GSK-3β and p-GSK-3β were
assessed by WB.
Statistical analysis
All experiments were performed ≥3 independent
experiments performed in duplicate, and data are expressed as mean
± SD. Statistical significance of multiple groups was determined by
two-way analysis of variance with post hoc Bonferroni test (SPSS
version 17.0). Differences were considered statistically
significant when P<0.05.
Results
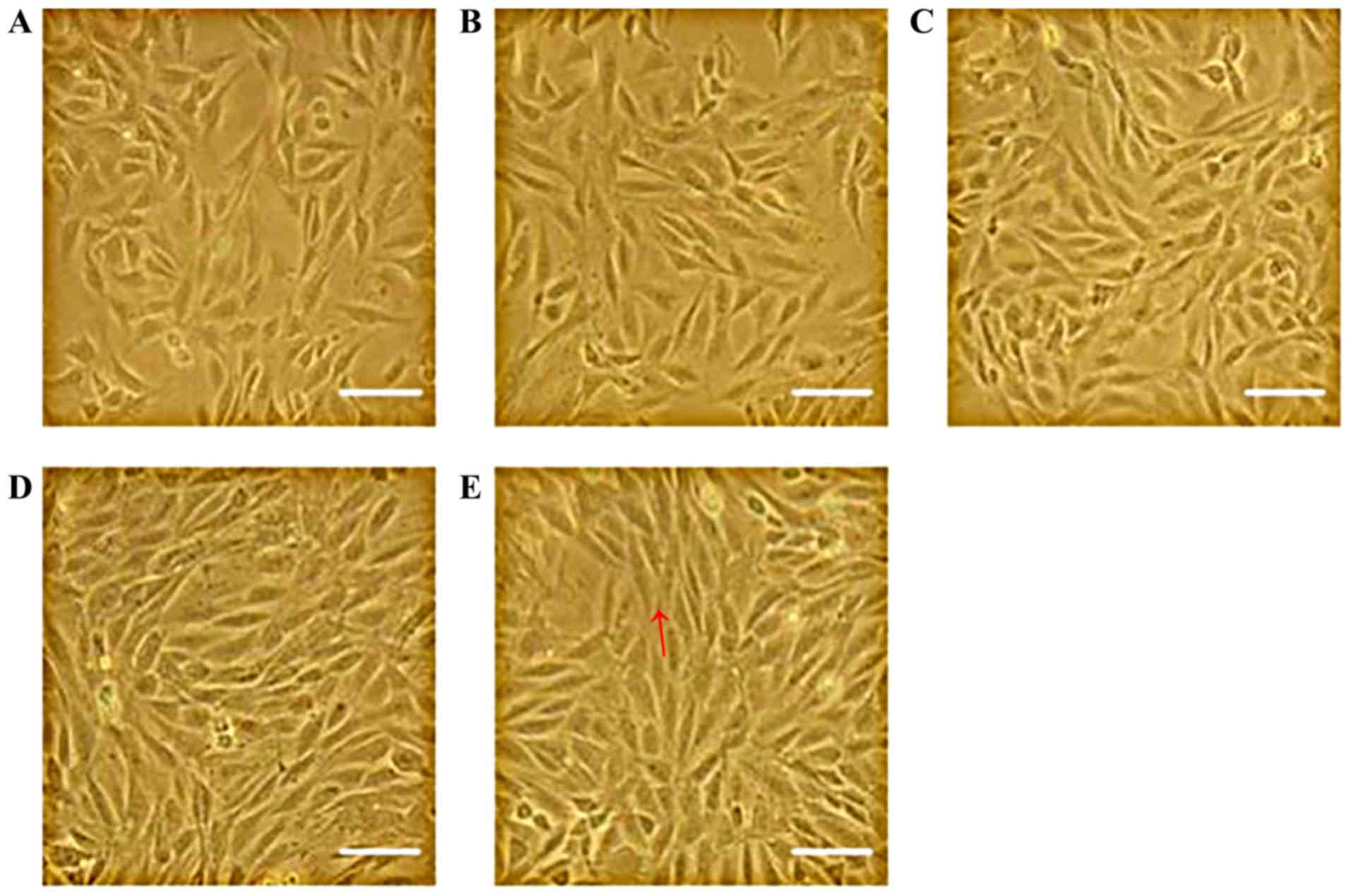
Effect of mechanical stretch on the
shape of L6 myoblasts
Images captured using an inverted microscope showed
that the unstretched, plate-attached cells mainly possessed a
spindle and irregular triangle-like morphology. However, after
mechanical stretch for ≥2 h, the cells were more tightly aligned in
the direction of the force field (Fig.
1). The longer the stretch interval, the more clear this
phenomenon was, suggesting that the cells were stretched
effectively.
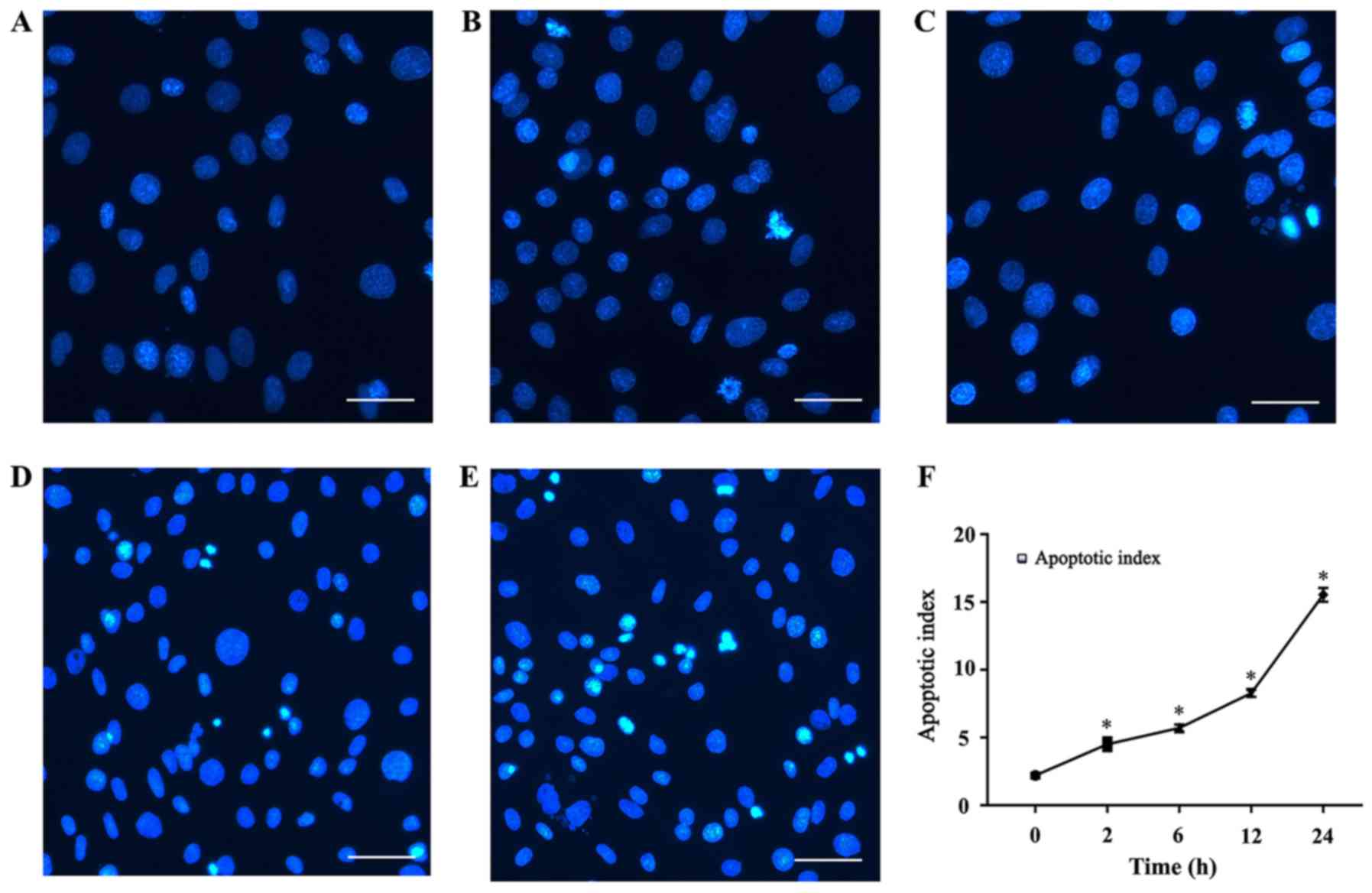
Effect of high levels of mechanical
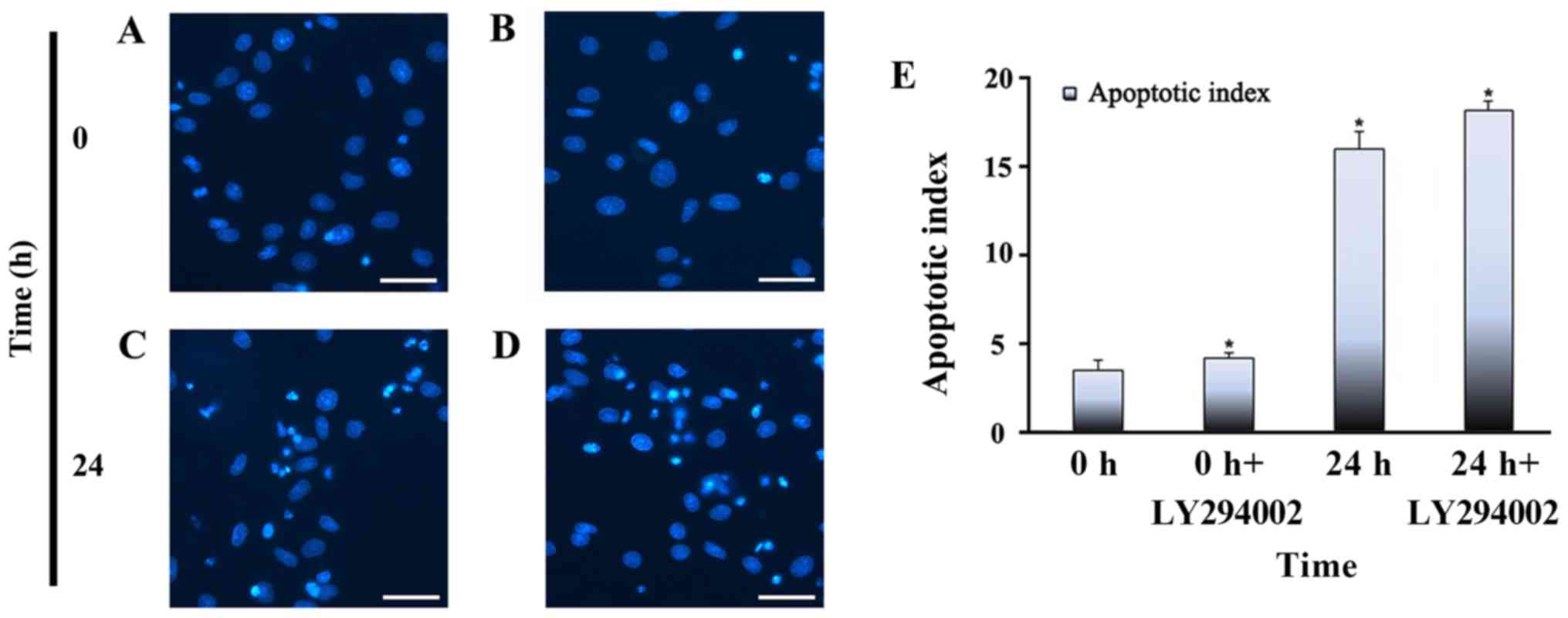
stretch on apoptosis
Hoechst 33258 is a convenient and popular nuclear
counterstain that emits blue fluorescence when it binds to DNA.
Accordingly, it is used for general counterstaining, as well as for
studies of apoptosis and the cell cycle. In this study, Hoechst
staining was used to observe the effects of different stretch times
on apoptosis. In the present study, cells were cyclically stretched
for 2, 6, 12 or 24 h. Stretch loading consisted of 15% elongation
at a frequency of 10 cycles/min. As shown in Fig. 2, the number of apoptotic cells in
the mechanical stretch-treated group increased in quantity relative
to the control group. Stained apoptotic cells exhibited dense,
bright fluorescent chromatin in their nucleus, whereas the nuclei
of normal cells appeared oval or round, and emitted a uniformly
dispersed blue fluorescence. Nuclear condensation, fragmentation
and apoptotic bodies gradually increased in frequency starting
after 2 h, peaking after 24 h, indicating that the cells were
undergoing apoptosis. The apoptotic index, defined as the
percentage of nuclei in each region that were apoptotic, increased
with time; after continuous stretch for 24 h, apoptotic bodies
could be observed in the stretch-treated group but were less
abundant in the control group.
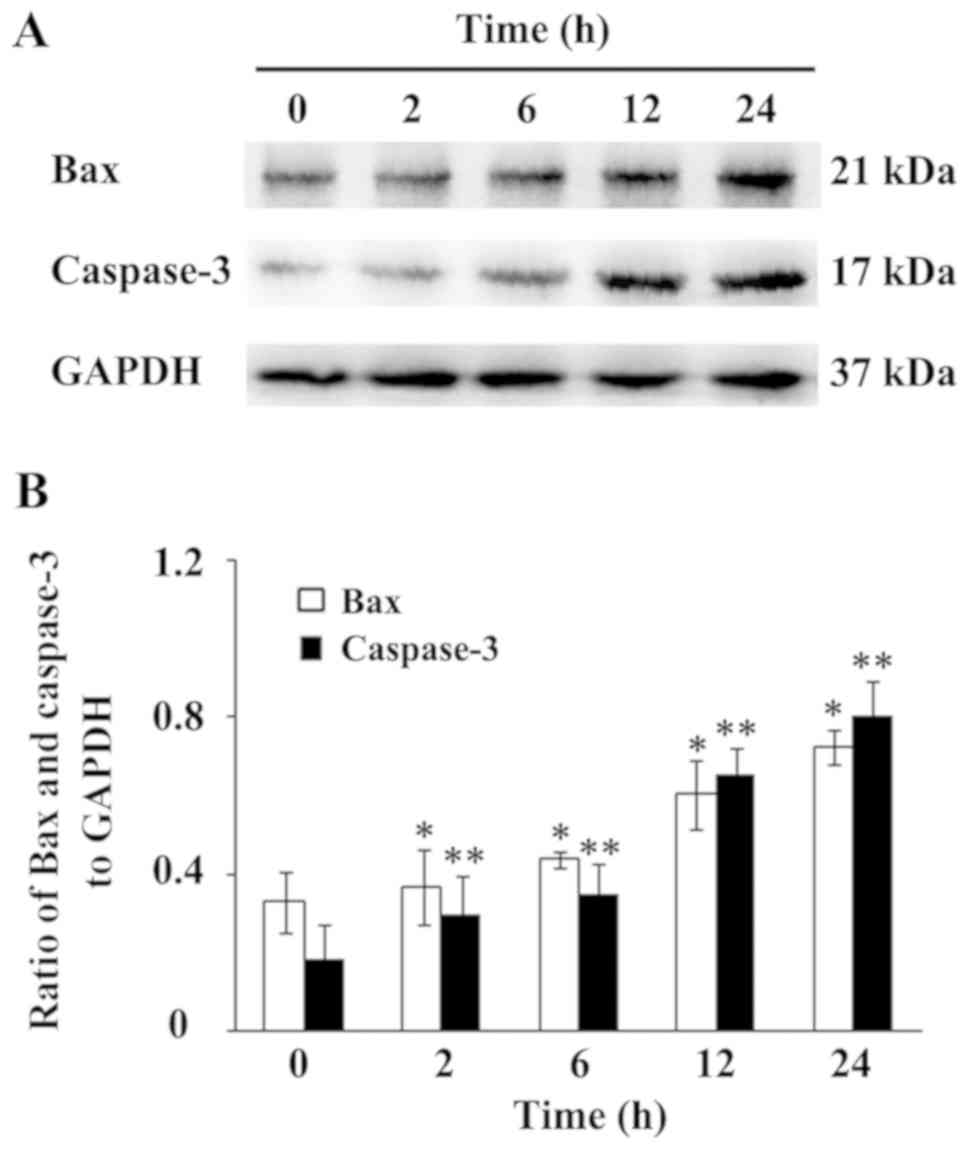
In apoptosis, Bax has been widely recognized as the
most representative proapoptotic protein of Bcl-2 family proteins
(25). Caspase-3 has also widely
accepted as a marker of most types of apoptosis (26). In the present study, protein
expression of Bax and Caspase-3 was detected via WB. As shown in
Fig. 3, an increase in Bax and
Caspase-3 protein expression was observed after 2 and 12 h of
mechanical loading, respectively, which increased gradually in a
time-dependent manner. The ratio of Bax and Caspase-3 to GAPDH are
displayed in Fig. 3B. Taken
together, these results suggested that the mechanical loading
condition promotes myoblast apoptosis in a time-dependent
manner.
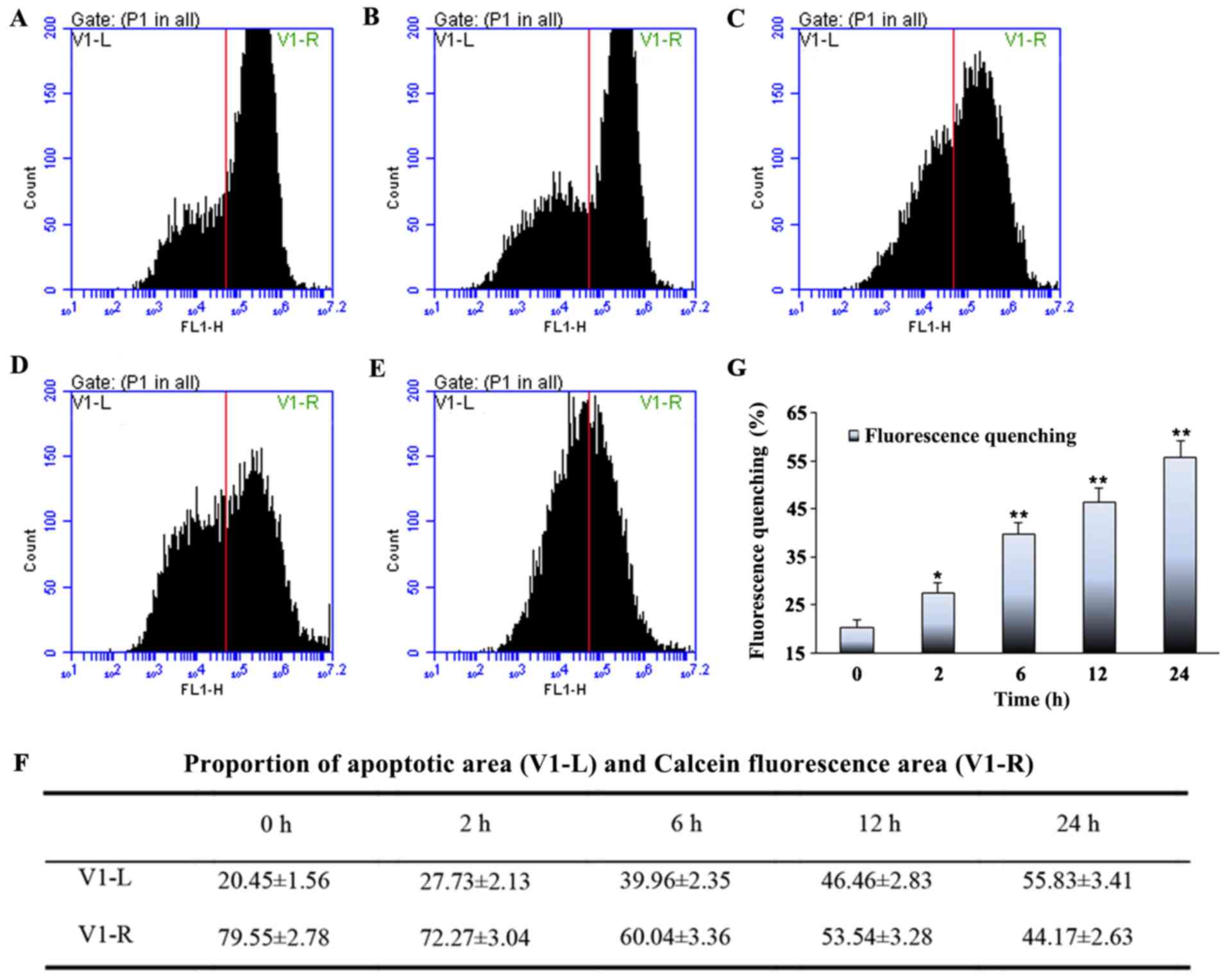
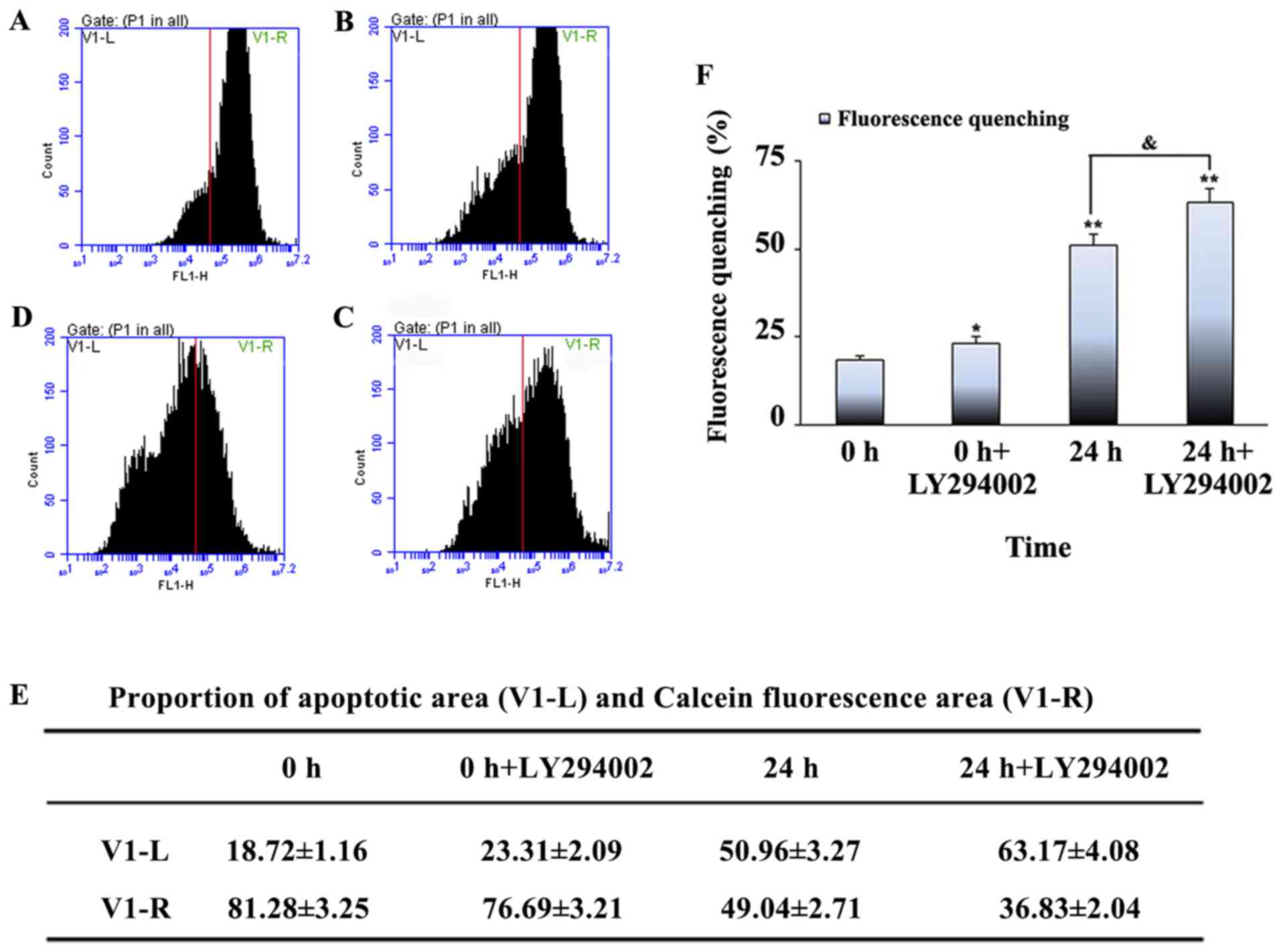
To further investigate the effects of mechanical
stretch on the regulation of apoptosis, the opening of the MPTP, a
useful method for monitoring the apoptotic rate in most cell types,
was tested. MPTP function can be measured by monitoring its
fluorescence intensity via flow cytometry. The number of cells in
the proportion of apoptotic area (V1-L) was the lowest in the
control group. As the mechanical stretch time increased, the number
of cells in V1-L also increased, reaching a maximum after 24 h
(Fig. 4). The proportion of cells
in V1-L represents the proportion of apoptotic cells; these
findings demonstrated that cyclic stretch can induce myoblast
apoptosis in a time-dependent manner.
Activation of the PI3K/Akt signaling
pathway by mechanical stretch
Activation of PI3K is typically essential for cell
proliferation and plays a central role in cellular signaling,
leading to cell proliferation, survival, motility and secretion, as
well as specialized cell responses, such as the respiratory burst
of granulocytes (27). To
determine whether mechanical stretch affects the PI3K/Akt signaling
pathway in myoblasts, the protein expression levels of p-Akt and
Akt by were measured by WB. In certain samples, LY294002, an
inhibitor of the PI3K/Akt signaling pathway was added, and
apoptosis was monitored by staining with Hoechst 33258 and
detecting changes in the MPTP by flow cytometry. The aforementioned
proteins were also detected via WB.
Inhibition of PI3K/Akt signaling on
GSK3β dephosphorylation, mPTP opening and myoblast apoptosis
The apoptotic indices of each group of myoblasts are
presented in Fig. 5. After
stretching for 24 h and adding LY294002, myoblasts exhibited an
elevated apoptotic index. However, myoblasts that were not exposed
to stretch, or that were treated with LY294002 alone, had a lower
apoptotic index than stretch-treated myoblasts. The lowest
apoptotic index was observed in non-stretched cells without
LY294002 treatment.
GSK-3β is an important cell regulator of metabolism
and an important regulator of cell proliferation, migration, cell
death and immune function (28,29).
Moreover, it is a major downstream target of PI3K/Akt signaling
(30). To further assess the
functional role of PI3K/Akt in stretch-induced apoptosis, a
specific PI3K inhibitor LY294002 was used. The V1-L proportion was
lower in the unstretched cells + LY294002 treatment compared with
the unstretched cells without treatment, and higher in the 24 h
stretched cells + LY294002 treatment compared with the 24 h
stretched cells without treatment. The V1-L proportion was highest
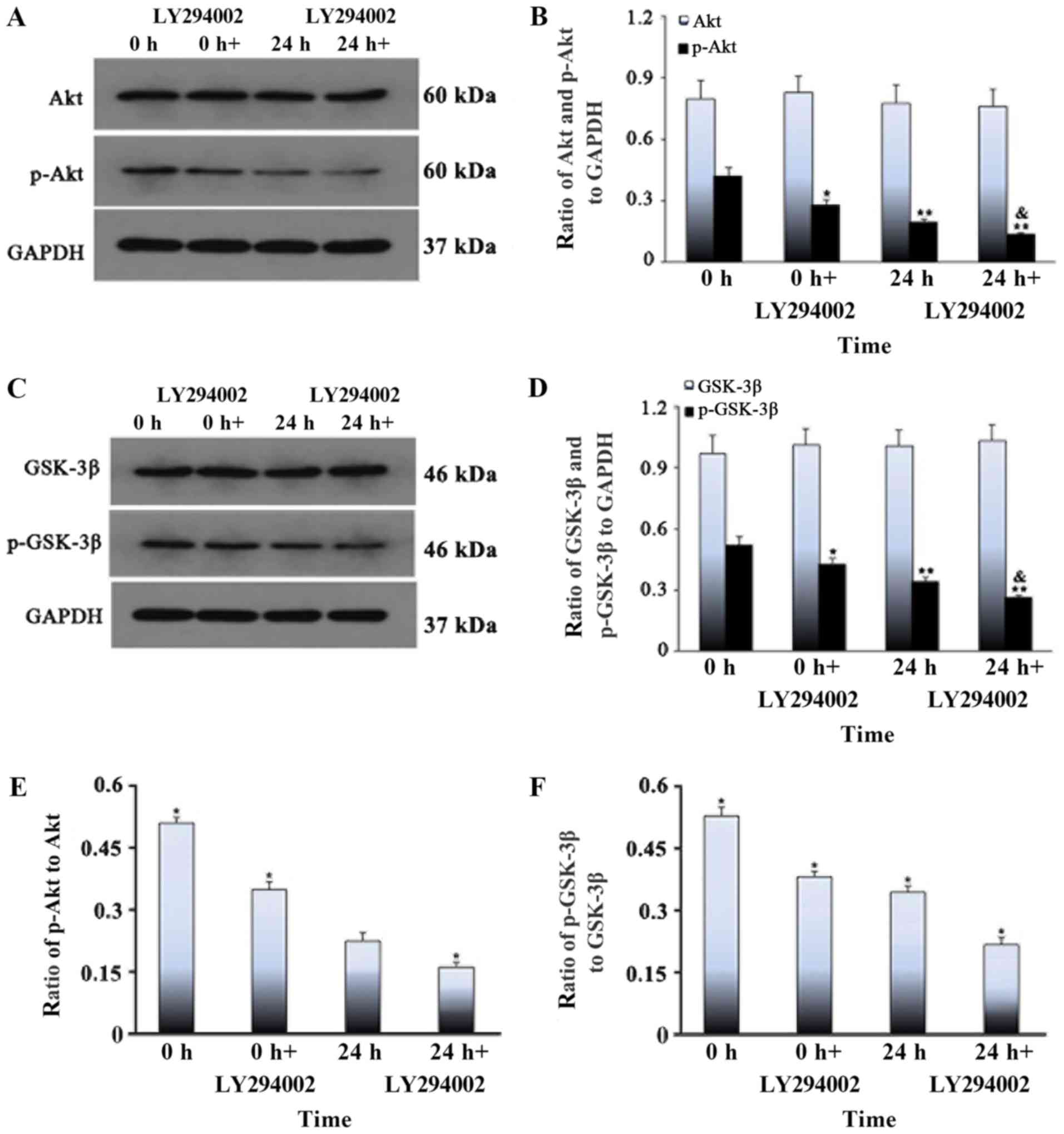
in the 24 h stretched cells + LY294002 treatment group (Fig. 6). The levels of total Akt and
GSK-3β did not change when the stretch time was extended, whereas
the levels of p-Akt and p-GSK-3β tended to decrease with stretch
time; the p-proteins were least abundant at 24 h (Fig. 7). After the addition of the PI3K
inhibitor, there was no difference between groups in the levels of
total Akt and GSK-3β, whereas the levels of p-Akt and p-GSK-3β were
lower in the inhibitor group than in the pure stretch group, and
the levels of p-proteins in the inhibitor group were lowest after
24 h treatment with the inhibitor (Fig. 8). It was speculated that LY294002
inhibited the phosphorylation of GSK-3β in myoblasts under cyclic
stress conditions, which led to the opening of MPTP and eventually
cell apoptosis. Therefore, the results suggested that inhibition of
the PI3K/Akt pathway plays a critical role in the dephosphorylation
of GSK-3β, MPTP opening and mitochondrial apoptosis of myoblast
cells under cyclic stress.
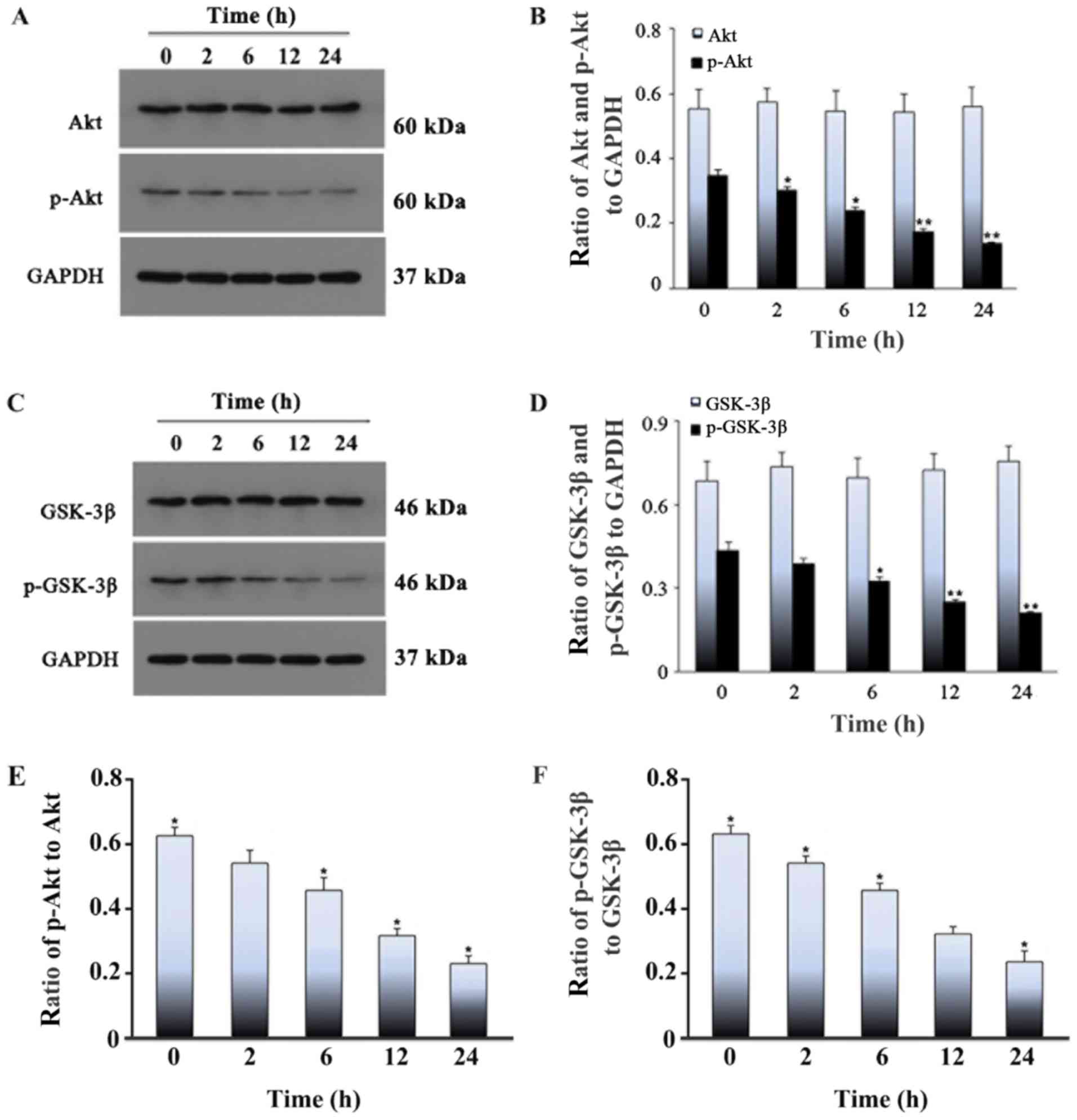 | Figure 7.Effect of cyclic stretch on the
levels of Akt, p-Akt, GSK-3β and p-GSK-3β in rat L6 myoblasts.
Myoblasts were exposed to cyclic stretch for 0, 2, 6, 12 or 24 h,
followed by western blotting. (A) Western blotting results of the
protein expression of (B) p-Akt was significantly decreased over
time, reaching a minimum level at 24 h, whereas the level of total
Akt remained stable. (C) Western blotting results of the protein
expression of (D) p-GSK-3β was significantly decreased over time,
reaching a minimum level at 24 h, whereas the level of total GSK-3β
protein remained stable. The ratio of (E) p-Akt to Akt and the
ratio of (F) p-GSK-3β to GSK-3β. Values represent mean ± SD of
three experiments. *P<0.05 vs. control; **P<0.01 vs. control.
GSK-3β, glycogen synthase kinase 3β; p, phosphorylated. |
Discussion
In the present study, the involvement of the
PI3K/Akt signaling pathway in apoptosis induced by cyclic stretch
in rat L6 myoblasts was investigated. First, it was demonstrated
that cyclic stretch consisting of 15% elongation at 10 cycles/min
affected the morphology and induced apoptosis of rat L6 myoblasts.
Second, it was shown that cyclic mechanical stretch can increase
the opening of the MPTP, thereby increasing mitochondrial
permeability. Furthermore, it was found that LY294002, a PI3K/Akt
signaling pathway inhibitor, partly promoted apoptosis caused by
cyclic stretch in rat L6 cells.
Overloading mechanical stretch may cause injury or
apoptosis in skeletal muscle (11). Several studies have shown that
excessive stretch can seriously damage the structure and
physiological function of cells (9). Our previous study reported that
apoptosis increased with stretch time (31). In the present study, to test the
hypothesis that excessive stretch would increase myoblast
apoptosis, histopathological and staining changes were observed in
cells subjected to cyclic stretch. Fluorescence microscopy revealed
that myoblasts tended to align in the direction of the force field,
and that this change became more pronounced as the stretch time
increased. Hoechst 33258 staining results displayed changes in cell
morphology. The structure of stretched L6 cells was markedly
different from those of the control group, and contained condensed
chromatin and apoptotic bodies. WB was also used to detect the
expression level of Bax and Caspase-3: The results showed that
cyclic mechanical stretch induces myoblast apoptosis in a
time-dependent manner.
Next, it was shown that the PI3K/Akt signaling
pathway was involved in mechanical stretch-induced myoblast
apoptosis, and that treatment with LY294002 inhibited the PI3K/Akt
signaling pathway. The PI3K/Akt signaling pathway plays a vital
role in diverse cellular functions, such as the suppression of
apoptosis and promotion of proliferation (32,33),
as well as cell cycle progression (34). Phosphorylation of Akt is an
important step in the process of apoptosis (35). Sussman (36) reported that phosphorylation of Akt
facilitates cell survival and proliferation, and regulates multiple
signaling pathways. On the other hand, GSK3β, an Akt substrate that
plays critical roles in oxidative stress-induced neuronal apoptosis
(37), is negatively regulated by
Akt activity.
PI3K is involved in the regulation of various
intracellular functions, such as glucose transport, cell
proliferation, differentiation and apoptosis (22). After PI3K is activated, three
products are produced. One of the products, phosphatidylinositol
(2,4,5)-triphosphate, acts as a second
messenger, binding to Akt to activate it and translocate it to the
cell membrane (38). Akt is a
serine/threonine protein kinase, also known as protein kinase B.
Activated Akt activates or inhibits downstream target proteins
through phosphorylation, thereby regulating cell proliferation,
differentiation and survival (22). Therefore, the signaling pathway
composed of PI3K and Akt plays a key role in regulating cell
proliferation and survival (38).
GSK-3β is also a serine/threonine protein kinase and is involved
glucose metabolism regulation, as well as numerous other functions
(39). A growing body of research
has found that GSK-3β phosphorylates a variety of substrates and is
involved in a variety of cellular processes, including regulation
of the cell cycle and apoptosis, regulation of gene transcription
and protein expression, and the maintenance of cytoskeletal
integrity (40). It has been shown
that activation of GSK-3β further activates apoptosis-associated
protein kinases (41). The
activity of GSK-3β is mainly dependent on its phosphorylation
status. Unlike most other kinases, p-GSK-3β is the inactive state
of GSK-3β (42). The
phosphorylation status of GSK-3β is mainly regulated by Akt, and
various stimulating factors activate the PI3K/Akt pathway,
phosphorylating GSK-3β to inactivate it and thereby inhibit
apoptosis (43). In models of
cardiac, cerebral and renal ischemia, ischemic treatment enhanced
the phosphorylation of Akt and GSK-3β (Ser9) by activating the
PI3K/Akt/GSK-3β signaling pathway and exerting its antiapoptotic
effect (44,45). Previous studies have also found
that activated GSK-3β can induce the opening of the MPTP, thereby
promoting cell apoptosis. By inhibiting the activity of GSK-3β, the
opening of the MPTP can be prevented, thereby inhibiting apoptosis
(46,47).
In the present study, a cell model of mechanical
stretch-induced myoblast apoptosis was established and used to
elucidate how PI3K/Akt signaling modulates this process.
Specifically, cells were treated with the pathway-specific
inhibitor, LY294002, to block PI3K/Akt signaling. Hoechst 33258
staining revealed that LY294002 increased apoptosis compared with
the 24 h stretch without inhibitor group, while the proportion of
apoptotic cells was highest in the 24 h stretch + inhibitor group.
These observations indicated that LY294002 promoted cyclic
stretch-induced apoptosis, in accordance with the results of flow
cytometry assays. WB revealed that levels of the phosphorylated
proteins, p-Akt and p-GSK-3β were reduced in PI3K/Akt signaling,
indicating that PI3K/Akt signaling is involved in stretch-induced
apoptosis. No difference was observed between the total levels of
Akt and GSK-3β proteins after the addition of PI3K inhibitors.
Together, these findings suggested that myoblast apoptosis induced
by cyclic stretch is related to the phosphorylation levels of Akt
and GSK-3β, and that the phosphorylation level of both proteins is
negatively associated with myoblast apoptosis.
In addition, the opening state of the MPTP was
investigated. Mitochondrial damage induced by the opening of the
MPTP leads to the disruption of cellular functions and eventually
causes cell death by inducing the release of apoptogenic factors,
depolarizing the transmembrane potential and impairing oxidative
phosphorylation (48,49). Moreover, substances that promote
MPTP opening can induce apoptosis, whereas substances that prevent
its opening can prevent apoptosis (27). In general, changes in mitochondrial
membrane potential are used to indirectly detect mitochondrial
membrane permeability. However, the increase in mitochondrial
membrane permeability occurs before the change in membrane
potential. Therefore, direct detection of mitochondrial membrane
permeability enables the detection of apoptosis at an earlier stage
(50,51). In this experiment, flow cytometry
was used to detect the opening of the MPTP. As stretch loading time
increased, this may have led to an increase in the intracellular
concentration of LY294002, thereby stimulating a gradual increase
in mitochondrial membrane permeability and maintaining the MPTP in
an open state via the inhibition of PI3K/Akt signaling,
consequently leading to an increase in the proportion of apoptotic
cells over time. In support of this, it was shown that the
fluorescence quenching rate was higher in the inhibitor-treated
group than in cells that were simply stretched for 24 h, and was
highest in the 24 h stretch + LY294002 treatment group.
Furthermore, activation of the mitochondrial apoptosis pathway has
been previously associated with the MPTP (52). Collectively, these findings
suggested that LY294002 increased myoblast apoptosis by promoting
MPTP opening via the PI3K/Akt pathway.
In summary, it was shown in the present study that
mitochondria, as the main site of endogenous apoptosis, play
important roles in this process. The opening of the MPTP is a key
enabling event in mitochondria-mediated apoptosis, and the
subsequent decrease of membrane potential is an important feature
of early apoptosis. The PI3K/Akt signaling pathway is important for
cell proliferation and survival. In cell models under multiple
stimulation, the PI3K/Akt signaling pathway has been shown to be an
important factor in inhibiting apoptosis and promoting cell
survival (53). However, to the
best of our knowledge, the role of PI3K/Akt signaling in cyclic
stress-induced apoptosis has not been previouly reported. Further
investigation of this will help clarify the mechanisms underlying
adaptive reconstruction of facial and maxillary muscles during
functional correction, and thereby provide a theoretical basis for
clinical observations.
The limitations of the present study are as follows:
i) Although Akt, a target for PI3K kinase activation, is a key
signaling molecule for the PI3K/Akt/GSK-3β signaling pathway, thus
investigations are required to detect PI3K in subsequent
experiments; and ii) only one PI3K inhibitor was used to treat the
cells when observing the morphological changes and apoptosis of
myoblasts, and the specificity of this inhibitor may be lower than
that of RNAi technology. In future validation studies, it would be
beneficial to use RNAi technology to inhibit the PI3K/Akt/GSK-3β
pathway.
Acknowledgements
Not applicable.
Funding
The current study was supported by the National
Natural Science Foundation of China (grant no. 31870929).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XH, XYa, XYu, ML, QiaZ and YT conceived and designed
the experiments. YT, FW and XYa performed the experiments. QiaZ,
JC, QiZ acquired the data. JC and QiZ analyzed and interpreted the
data. ML, QiZ, YT and XH drafted the manuscript. QiaZ, ML and XYu
reviewed and edited the manuscript. QiaZ and XYu supervised the
study. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Proske U and Morgan DL: Muscle damage from
eccentric exercise: Mechanism, mechanical signs, adaptation and
clinical applications. J Physiol. 537:333–345. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thiruvenkatachari B, Harrison J,
Worthington H and O'Brien K: Early orthodontic treatment for Class
II malocclusion reduces the chance of incisal trauma: Results of a
Cochrane systematic review. Am J Orthod Dentofacial Orthop.
148:47–59. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
DiBiase AT, Cobourne MT and Lee RT: The
use of functional appliances in contemporary orthodontic practice.
Br Dent J. 218:123–128. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
O'Brien K, Wright J, Conboy F, Sanjie Y,
Mandall N, Chadwick S, Connolly I, Cook P, Birnie D, Hammond M, et
al: Effectiveness of treatment for Class II malocclusion with the
Herbst or twin-block appliances: A randomized, controlled trial. Am
J Orthod Dentofacial Orthop. 124:128–137. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yamin-Lacouture C, Woodside DG, Sectakof
PA and Sessle BJ: The action of three types of functional
appliances on the activity of the masticatory muscles. Am J Orthod
Dentofacial Orthop. 112:560–572. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tallgren A, Christiansen RL, Ash M Jr and
Miller RL: Effects of a myofunctional appliance on orofacial muscle
activity and structures. Angle Orthod. 68:249–258. 1998.PubMed/NCBI
|
|
7
|
Haga JH, Li YS and Chien S: Molecular
basis of the effects of mechanical stretch on vascular smooth
muscle cells. J Biomech. 40:947–960. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Seale P and Rudnicki MA: A new look at the
origin, function, and ‘stem-cell’ status of muscle satellite cells.
Dev Biol. 218:115–124. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hawke TJ and Garry DJ: Myogenic satellite
cells: Physiology to molecular biology. J Appl Physiol (1985).
91:534–551. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tan J, Kuang W, Jin Z, Jin F, Xu L, Yu Q,
Kong L, Zeng G, Yuan X and Duan Y: Inhibition of NFkappaB by
activated c-Jun NH2 terminal kinase 1 acts as a switch for C2C12
cell death under excessive stretch. Apoptosis. 14:764–770. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sun K, Liu F, Wang J, Guo Z, Ji Z and Yao
M: The effect of mechanical stretch stress on the differentiation
and apoptosis of human growth plate chondrocytes. In Vitro Cell Dev
Biol Anim. 53:141–148. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cheng CS, El-Abd Y, Bui K, Hyun YE, Hughes
RH, Kraus WE and Truskey GA: Conditions that promote primary human
skeletal myoblast culture and muscle differentiation in vitro. Am J
Physiol Cell Physiol. 306:C385–C395. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen R, Liu X, Huang W, Zeng H, Shi D, Cao
B and Liao H: Effects of mechanical stimulation on expression of
autoantigens in myoblasts. Zhongguo Xiu Fu Chong Jian Wai Ke Za
Zhi. 27:1128–1133. 2013.(In Chinese). PubMed/NCBI
|
|
14
|
Liu J, Liu J, Mao J, Yuan X, Lin Z and Li
Y: Caspase-3-mediated cyclic stretch-induced myoblast apoptosis via
a Fas/FasL-independent signaling pathway during myogenesis. J Cell
Biochem. 107:834–844. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kerr JF: History of the events leading to
the formulation of the apoptosis concept. Toxicology 181-182.
471–474. 2002. View Article : Google Scholar
|
|
16
|
Fiers W, Beyaert R, Declercq W and
Vandenabeele P: More than one way to die: Apoptosis, necrosis and
reactive oxygen damage. Oncogene. 18:7719–7730. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cheng W, Li B, Kajstura J, Li P, Wolin MS,
Sonnenblick EH, Hintze TH, Olivetti G and Anversa P:
Stretch-induced programmed myocyte cell death. J Clin Invest.
96:2247–2259. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang Y, Zhou H, Wu W, Shi C, Hu S, Yin T,
Ma Q, Han T, Zhang Y, Tian F and Chen Y: Liraglutide protects
cardiac microvascular endothelial cells against
hypoxia/reoxygenation injury through the suppression of the
SR-Ca(2+)-XO-ROS axis via activation of the
GLP-1R/PI3K/Akt/survivin pathways. Free Radic Biol Med. 95:278–292.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang J, Chen L, Yang J, Ding J, Rong H,
Dong W and Li X: High mobility group box-1 induces migration of
vascular smooth muscle cells via TLR4-dependent PI3K/Akt pathway
activation. Mol Biol Rep. 39:3361–3367. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee WJ: Insulin-like growth
factor-I-induced androgen receptor activation is mediated by the
PI3K/Akt pathway in C2C12 skeletal muscle cells. Mol Cells.
28:495–499. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tang Y, Liu P, Tian Y, Xu Y, Ren F, Cui X
and Fan J: Overexpression of ribonuclease inhibitor defines good
prognosis and suppresses proliferation and metastasis in human
colorectal cancer cells via PI3K/AKT pathway. Clin Transl Oncol.
17:306–313. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shaw RJ and Cantley LC: Ras, PI(3)K and
mTOR signalling controls tumour cell growth. Nature. 441:424–430.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jie B, Zhang X, Wu X, Xin Y, Liu Y and Guo
Y: Neuregulin-1 suppresses cardiomyocyte apoptosis by activating
PI3K/Akt and inhibiting mitochondrial permeability transition pore.
Mol Cell Biochem. 370:35–43. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Giorgio V, von Stockum S, Antoniel M,
Fabbro A, Fogolari F, Forte M, Glick GD, Petronilli V, Zoratti M,
Szabó I, et al: Dimers of mitochondrial ATP synthase form the
permeability transition pore. Proc Natl Acad Sci USA.
110:5887–5892. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lindsay J, Esposti MD and Gilmore AP:
Bcl-2 proteins and mitochondrial-specificity in membrane targeting
for death. Biochim Biophys Acta. 80:593–614. 2011.
|
|
26
|
Riedl SJ and Shi Y: Molecular mechanisms
of caspase regulation during apoptosis. Nat Rev Mol Cell Biol.
5:897–907. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Claerhout S, Decraene D, Van Laethem A,
Van Kelst S, Agostinis P and Garmyn M: Akt delays the
early-activated apoptotic pathway in UVB-irradiated keratinocytes
via BAD translocation. J Invest Dermatol. 127:429–438. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jacobs KM, Bhave SR, Ferraro DJ, Jaboin
JJ, Hallahan DE and Thotala D: GSK-3β: A bifunctional role in cell
death pathways. Int J Cell Biol. 2012:9307102012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu D and Pan W: GSK3: A multifaceted
kinase in Wnt signaling. Trends Biochem Sci. 35:161–168. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nakayama M, Hisatsune J, Yamasaki E,
Isomoto H, Kurazono H, Hatakeyama M, Azuma T, Yamaoka Y, Yahiro K,
Moss J and Hirayama T: Helicobacter pylori VacA-induced inhibition
of GSK3 through the PI3K/Akt signaling pathway. J Biol Chem.
284:1612–1619. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu J, Liu J, Mao J, Yuan X, Lin Z and Li
Y: Caspase-3-mediated cyclic stretch-induced myoblast apoptosis via
a Fas/FasL- independent signaling pathway during myogenesis. J Cell
Biochem. 107:834–844. 2010. View Article : Google Scholar
|
|
32
|
Freudlsperger C, Burnett JR, Friedman JA,
Kannabiran VR, Chen Z and Van Waes C: EGFR-PI3K-AKT-mTOR signaling
in head and neck squamous cell carcinomas: Attractive targets for
molecular-oriented therapy. Expert Opin Ther Targets. 15:63–74.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ke F, Wang Z, Song X, Ma Q, Hu Y, Jiang L,
Zhang Y, Liu Y, Zhang Y and Gong W: Cryptotanshinone induces cell
cycle arrest and apoptosis through the JAK2/STAT3 and PI3K/Akt/NFκB
pathways in cholangiocarcinoma cells. Drug Des Devel Ther.
11:1753–1766. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chang F, Lee JT, Navolanic PM, Steelman
LS, Shelton JG, Blalock WL, Franklin RA and McCubrey JA:
Involvement of PI3K/Akt pathway in cell cycle progression,
apoptosis, and neoplastic transformation: A target for cancer
chemotherapy. Leukemia. 17:590–603. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gu X, Han D, Chen W, Zhang L, Lin Q, Gao
J, Fanning S and Han B: SIRT1-mediated FoxOs pathways protect
against apoptosis by promoting autophagy in osteoblast-like
MC3T3-E1 cells exposed to sodium fluoride. Oncotarget.
7:65218–65230. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sussman M: ‘AKT'ing lessons for stem
cells: Regulation of cardiac myocyte and progenitor cell
proliferation. Trends Cardiovasc Med. 17:235–240. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Long ZM, Zhao L, Jiang R, Wang KJ, Luo SF,
Zheng M, Li XF and He GQ: Valproic acid modifies synaptic structure
and accelerates neurite outgrowth via the glycogen synthase
kinase-3β signaling pathway in an Alzheimer's disease model. CNS
Neurosci Ther. 21:887–897. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Osaki M, Oshimura M and Ito H: PI3K-Akt
pathway: Its functions and alterations in human cancer. Apoptosis.
9:667–676. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ali A, Hoeflich KP and Woodgett JR:
Glycogen synthase kinase-3: Properties, functions, and regulation.
Chem Rev. 101:2527–2540. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cohen P and Frame S: The renaissance of
GSK3. Nat Rev Mol Cell Biol. 2:769–776. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang X, Jiang W, Zhou AL, Zhao M and
Jiang DR: Inhibitory effect of oxymatrine on hepatocyte apoptosis
via TLR4/PI3K/Akt/GSK-3β signaling pathway. World J Gastroenterol.
23:3839–3849. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Embi N, Rylatt DB and Cohen P: Glycogen
synthase kinase-3 from rabbit skeletal muscle. Separation from
cyclic-AMP-dependent protein kinase and phosphorylase kinase. Eur J
Biochem. 107:519–527. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cai Z and Semenza GL:
Phosphatidylinositol-3-Kinase signaling is required for
erythropoietin-mediated acute protection against myocardial
ischemia/reperfusion injury. Circulation. 109:2050–2053. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Linseman DA, Butts BD, Precht TA, Phelps
RA, Le SS, Laessig TA, Bouchard RJ, Florez-McClure ML and
Heidenreich KA: Glycogen synthase kinase-3beta phosphorylates Bax
and promotes its mitochondrial localization during neuronal
apoptosis. J Neurosci. 24:9993–10002. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sharples EJ, Patel N, Brown P, Stewart K,
Mota-Philipe H, Sheaff M, Kieswich J, Allen D, Harwood S, Raftery
M, et al: Erythropoietin protects the kidney against the injury and
dysfunction caused by ischemia-reperfusion. J Am Soc Nephrol.
15:2115–2124. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Pap M and Cooper GM: Role of translation
initiation factor 2B in control of cell survival by the
phosphatidylinositol 3-kinase/Akt/glycogen synthase kinase 3beta
signaling pathway. Mol Cell Biol. 32:578–586. 2002. View Article : Google Scholar
|
|
47
|
Bopassa JC, Ferra R, Gateau-Rosech O,
Couture-Lepetit E and Ovize M: PI 3-kinase regulates the
mitochondrial transition pore in controlled reperfusion and
postconditioning. Cardiovasc Res. 69:178–185. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu XH, Aksan A, Menze MA, Hand SC and
Toner M: Trehalose loading through the mitochondrial permeability
transition pore enhances desiccation tolerance in rat liver
mitochondria. Biochim Biophys Acta. 1717:21–26. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ly JD, Grubb DR and Lawen A: The
mitochondrial membrane potential (deltapsi(m)) in apoptosis; an
update. Apoptosis. 8:115–128. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Quan JH, Cha GH, Zhou W, Chu JQ, Nishikawa
Y and Lee YH: Involvement of PI 3 kinase/Akt-dependent Bad
phosphorylation in Toxoplasma gondii-mediated inhibition of host
cell apoptosis. Exp Parasitol. 133:462–471. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Gardai SJ, Hildeman DA, Frankel SK,
Whitlock BB, Frasch SC, Borregaard N, Marrack P, Bratton DL and
Henson PM: Phosphorylation of Bax Ser184 by Akt regulates its
activity and apoptosis in neutrophils. J Biol Chem.
279:21085–21095. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Fu M, Wan F, Li Z and Zhang F: 4SC-202
activates ASK1-dependent mitochondrial apoptosis pathway to inhibit
hepatocellular carcinoma cells. Biochem Biophys Res Commun.
471:267–273. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhou BH, Tan PP, Jia LS, Zhao WP, Wang JC
and Wang HW: PI3K/AKT signaling pathway involvement in
fluoride-induced apoptosis in C2C12 cells. Chemosphere.
199:297–302. 2018. View Article : Google Scholar : PubMed/NCBI
|