Introduction
Female breast cancer is a major health problem that
accounts for 30% of cancer cases worldwide; therefore, breast
cancer is the most common type of cancer in women (1). The incidence of breast cancer varies
greatly worldwide, with 0.194 in East Africa and 0.897 in Western
Europe, exhibiting an overall increasing trend (2). Generally, breast cancer patients rely
on conventionally available chemotherapy; however, the results are
often unsatisfactory with a poor patient prognosis. There are many
risk factors associated with breast cancer, including the use of
hormonal contraceptives, sedentariness, and alcohol consumption;
however, its etiology and pathogenesis are still not clearly
understood. Therefore, there is an urgent need to develop new drugs
to improve the prognosis of breast cancer patients.
Natural products provide a valuable source of
anticancer compounds. Many commonly used chemotherapeutic drugs,
such as vinblastine, etoposide, paclitaxel, and camptothecin, are
derived directly or indirectly from natural products. Among the
various plant bioactive compounds, alkaloids appear to have the
most prominent anticancer effect. Camptothecin and vinblastine are
two of the most notable examples (3), and other alkaloids, such as berberine
and isoquinoline, also exhibit antitumor potential (4). Previous studies have reported that
dehydrocorydaline (DHC), an alkaloid isolated from Corydali
syanhusuo (WT Wang, 1985), presents anticancer potential.
However, there are very few studies on the use of DHC for breast
cancer treatment. Previous studies indicated that DHC exerts
anti-allergic and antitumor effects, and can inhibit the
proliferation of MCF-7 breast cancer cells in vitro
(5). However, the underlying
mechanism of action remains unclear.
Among the many metastasis-related molecules, CDK1,
CCND1, and MMP family members are known to be closely related to
cell proliferation, migration, and differentiation. In addition,
the BCL2 and caspase family proteins are involved in apoptosis
(6). These molecules may also play
a key role in the inhibition of breast cancer mediated by DHC. In
the present study, the effects of DHC treatment on cell
proliferation and migration, as well as on the expression of
apoptotic markers in vitro and in vivo were
evaluated, thus revealing the molecular mechanism of DHC against
cancer.
Materials and methods
Cell culture
For the present study, human breast cancer cells
MDA-MB-231 were obtained from the American Type Culture Collection
(ATCC). During the experimental protocol, cells were cultured in
the Dulbecco's modified Eagle medium-high glucose (H-DMEM)
supplemented with 10% fetal bovine serum (FBS), 100 U/ml
penicillin, and 100 µg/ml streptomycin (all from Gibco; Thermo
Fisher Scientific, Inc.). Cells were maintained at 37°C in a
humidified atmosphere supplemented with 5% CO2 in an
incubator. The culture medium was changed every ~2–3 days. Cells
were passaged when the cell confluency reached ~80-90%; cells from
different flasks were passaged independently.
Cell viability
The viability of MDA-MB-231 cells after treatment
with DHC (dissolved in DMSO) was assessed through a Cell Counting
Kit-8 (CCK-8) assay. After trypsinization (0.25%) at 37°C for 2
min, cells were seeded on 96-well plates at a cell density of
3×104 cells/cm2 and cultured for 24 h at
37°C, to allow adequate cell attachment. Then, the culture medium
was replaced with FBS-free H-DMEM for cell starvation. After 24 h
of starvation at 37°C, the medium was changed with fresh 10% FBS
H-DMEM supplemented with various concentrations of DHC (20, 30, 40,
50 or 100 µM). In addition, the DMSO-treatment group was set as the
blank group, and the non-treatment group was set as the control
group. All cells were cultured for 48 or 72 h at 37°C, then cell
viability was evaluated by a CCK-8 assay. A volume of 10 µl CCK-8
(Beijing Solarbio Science & Technology Co., Ltd.) was added in
each well, and the plates were incubated for 1 h in the dark. Then,
the absorbance was measured at 450 nm using a microplate
spectrophotometer.
Cell proliferation
The effect of DHC treatment on the proliferation of
MDA-MB-231 cells was evaluated by 5-ethynyl-2′-deoxyuridine (EdU)
staining and flow cytometry. For EdU staining, cells were seeded on
96-well plates at a cell density of 3×104
cells/cm2. After starvation at 37°C overnight, the cells
were treated with various concentrations of DHC (10, 50 or 100 µM)
for 22 or 46 h at 37°C, and incubated with 50 µM EdU for another 2
h at 37°C. Subsequently, the samples were fixed with 4%
paraformaldehyde at room temperature for 20 min. After 3×5 min
washes with PBS, cells were stained with 100 µl Apollo 567 stain
reaction buffer for 30 min at room temperature. Then, the cells
were washed again with PBS, and stained with DAPI (5 µg/ml) at room
temperature for 15 min. Finally, EdU-labelled cells were visualized
under a fluorescence microscope (magnification, ×200; five random
fields of view). Image-Pro Plus software (version 6.0; Media
Cybernetics, Inc.) was used to quantify EdU-labelled cells. The
ratio of EdU-positive cells was calculated using the following
formula: EdU add-in cells/DAPI-stained cells ×100%. For the flow
cytometric assay, MDA-MB-231 cells were treated with 100 µM DHC for
48 h at 37°C. Then, cells were washed once with pre-cooled PBS and
centrifuged (300 × g for 5 min at 4°C). Cells were fixed with 500
µl 70% ethanol at 4°C for 2 h. After fixation, the cells were
washed once with PBS, and the cell suspension was incubated with
100 µl RNase A at 37°C for 30 min. Propidium iodide (PI; 400 µl;
cat. no. C0080; Solarbio biotech, Beijing, cat no. C0080) was added
into the cell suspension, which was then incubated at 4°C for 30
min in the dark. Samples were analyzed using a BD FACSCalibur flow
cytometer (BD Biosciences) at 488 nm excitation wavelength and
FlowJo software (version 7.6.1; Tree Star, Inc.).
Cell apoptosis
In the present study, cell apoptosis was evaluated
by Annexin V-FITC and PI staining and flow cytometry. In brief,
MDA-MB-231 cells were seeded on 6-well plates and treated as
described in the previous sections. After cell starvation, 50 µM
DHC was added into the culture medium, and the cells were cultured
for another 48 h at 37°C. Subsequently, the apoptosis of MDA-MB-231
cells was evaluated using an Annexin V-FITC/PI kit (cat no. FAK012;
Neobioscience), according to the manufacturer's protocol. Cells
were washed twice in PBS by centrifugation (300 × g for 5 min at
4°C), and resuspended in 195 µl binding buffer. Subsequently, 5 µl
Annexin V-FITC and 10 µl PI were added to each cell suspension and
incubated for 10 min at RT in the dark. The samples were analyzed
using a BD FACSCalibur flow cytometer (BD Biosciences) and FlowJo
software (version 7.6.1; Tree Star, Inc.).
Colony formation
Alterations in the colony-forming activity of
MDA-MB-231 cells after treatment with DHC were also evaluated.
MDA-MB-231 cells were seeded on 6-well plates at a cell density of
1×103 cells/well. Subsequently, 50 or 100 µM DHC were
added into the culture medium, and the cells were cultured for 12
days at 37°C. The medium was changed every 3 days. Then, the cells
were stained using crystal violet solution (cat. C8470; Beijing
Solarbio Science & Technology, Co., Ltd.) at room temperature
for 15 min, images were captured, and colonies with >50 cells
were counted using a light microscope (magnification, ×100).
Scratch assay
The invasive capacity of the MDA-MB-231 cells was
verified through a scratch assay. MDA-MB-231 cells were seeded on
6-well plates at a cell density of 3×104
cells/cm2. When the cells reached confluence, a line of
10×1.4 mm (length × width) was created by scraping the cells with a
pipette tip. Then, the suspended cells were washed away, and 2% FBS
H-DMEM was added into the wells. After another 48 h at 37°C, the
migrated cells were imaged and counted using a light microscope
(magnification, ×100).
Reverse transcription-quantitative PCR
(RT-qPCR)
Based on the aforementioned experiments, only the
effects of treatments with 20 and 50 µM DHC on the expression level
of various genes were verified. The expression levels of
proliferation-associated genes (CDK1 and CCND1) and
apoptosis-relevant genes (BCL2, caspases 3/8/9) were
assessed by RT-qPCR. Total RNA was extracted using the RNA simple
total RNA kit (Tiangen Biotech Co., Ltd.), according to the
manufacturer's instructions, from both the control and experimental
groups (cells treated with DHC for 48 h). Subsequently, isolated
mRNA was reverse-transcribed to cDNA using the RevertAid™ First
Strand cDNA synthesis kit (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. The mRNA expression
levels of the aforementioned genes and GAPDH were
quantified. qPCR was performed on the Bio-rad CFX96TM Real-Time PCR
system (Bio-Rad Laboratories, Inc.) using the PCR Master Mix kit
(Promega Corporation), according to the manufacturer's protocol.
The primer pairs used for qPCR are listed in Table I. The following thermocycling
conditions were used for qPCR: Initial denaturation at 95°C for 30
sec; followed by 39 cycles of 95°C for 5 sec and 60°C for 30 sec;
and a final extension at 72°C for 5 min. mRNA levels were
quantified using the 2−ΔΔCq method (7) and normalized to the internal
reference gene GAPDH.
 | Table I.Primers used for reverse
transcription-quantitative PCR. |
Table I.
Primers used for reverse
transcription-quantitative PCR.
|
| Sequence (5′→3′) |
|---|
|
|
|
|---|
| Gene | Forward | Reverse |
|---|
| CDK1 |
5′-CTGGCTCTTGGAAATTGAGCG-3′ |
5′-CTGGCAAGGCCAAAATCAGC-3′ |
| CCND1 |
5′-CTGATTGGACAGGCATGGGT-3′ |
5′-GTGCCTGGAAGTCAACGGTA-3′ |
| BCL2 |
5′-GGTGGGGTCATGTGTGTGG-3′ |
5′-CGGTTCAGGTACTCAGTCATCC-3′ |
| CASP3 |
5′-CATGGAAGCGAATCAATGGACT-3′ |
5′-CTGTACCAGACCGAGATGTCA-3′ |
| CASP8 |
5′-CGGACTCTCCAAGAGAACAGG-3′ |
5′-TCAAAGGTCGTGGTCAAAGCC-3′ |
| CASP9 |
5′-CTTCGTTTCTGCGAACTAACAGG-3′ |
5′-GCACCACTGGGGTAAGGTTT-3′ |
| GAPDH |
5′-GCACCGTCAAGGCTGAGAAC-3′ |
5′-TGGTGAAGACGCCAGTGGA-3′ |
Western blotting
Western blotting was performed to quantify the
protein expression levels of cleaved caspase 3/9 and MMP2/9. After
48 h of DHC treatment, cells were harvested and lysed in RIPA
buffer (Beyotime Institute of Biotechnology) supplemented with
protease inhibitors to extract total protein. Total protein was
quantified using a bicinchoninic acid assay. Subsequently, 35 µg
protein/lane were separated by SDS-PAGE on 10% polyacrylamide gels
and transferred to PVDF membranes. Subsequently, the membranes were
blocked with Tris-buffered saline (TBS) containing 5% non-fat dry
milk at 37°C for 30 min. The membranes were incubated overnight at
4°C with primary antibodies targeted against: MMP2 (cat. no.
ab92536; 1:1,000; Abcam), MMP9 (cat. no. ab76003; 1:1,000; Abcam),
cleaved caspase 3 (cat. no. ab2302; 1:1,000; Abcam), cleaved
caspase 9 (cat. no. ab185719; 1:1,000; Abcam) and GAPDH (cat. no.
10494-1-AP; 1:5,000; ProteinTech Group, Inc.). Following primary
incubation, the membranes were incubated with a secondary antibody
(cat. no. ab6721; 1:5,000; Abcam) at 37°C for 1 h. Finally, the
immunoreactive bands were developed using the Supersignal West
Femto Maximum Sensitivity Substrate (Thermo Fisher Scientific,
Inc.), and the corresponding images were analyzed using the
Quantity One Software (version 4.6.6; Bio-Rad Laboratories, Inc.)
using GAPDH as the loading control.
Animal experiments
In the present study, 8 female SCID mice (age, 6–8
weeks; weight, 20 g) were kept in pathogen-free conditions at 25°C
with 12-h light dark cycles and access to food and water ad
libitum. Mice were randomly divided into two groups (control
and experimental). In the control group, MDA-MB-231 cells
(3×106 cells resuspended in 100 µl Matrigel) were
injected subcutaneously into both flanks of each mouse. In the
experimental group, after subcutaneous implantation of MDA-MB-231
cells, 500 µM DHC (50 µl) was injected locally daily during the
first 3 days, and every other day for the next 10 days. The animals
were euthanized by CO2 inhalation until they ceased
breathing completely and then cervical dislocation followed. The
tumor weight was measured 2 weeks after the implantation of breast
cancer cells. The tumors were snap-frozen in liquid nitrogen and
submitted for immunohistochemical (IHC) analyses. All the animal
experiments were carried out according to ethical principles and
protocols approved by the Fifth Hospital of Wuhan.
IHC analyses
All tumor samples were fixed in 4% paraformaldehyde
at 4°C overnight and washed with PBS later. The samples were
dehydrated, embedded in paraffin, and cut into 5 µm-thick sections
using a cryostat. H&E (Beijing Solarbio Science &
Technology Co., Ltd.) staining was performed on the sections,
according to the manufacturer's protocol. Protein expression levels
of Ki67, proliferating cell nuclear antigen (PCNA) were verified
through immunofluorescence (IF) staining. Firstly, antigen
retrieval was performed by incubation in 0.01 M sodium citrate
buffer solution (pH=6.0) for 25 min in a water bath at 95°C.
Sections were incubated with primary antibodies against Ki67 (cat.
no. ab16667; 1:100; Abcam), PCNA (cat. no. ab92552; 1:100; Abcam)
at 4°C overnight. Subsequently, the sections were incubated with an
Alexa Fluor 488-conjugated secondary antibody (cat. no. A32731;
1:1,000; Thermo Fisher Scientific, Inc.) for 1 h at room
temperature. Nuclei were stained with DAPI for 10 min at room
temperature. Cell proliferation and histological alterations in the
DHC group compared with the control were examined by fluorescence
microcopy (magnification, ×200; five random fields of view). The
images were analyzed using Image-Pro Plus software (version 6.0;
Media Cybernetics, Inc.).
Statistical analyses
All experiments were performed at least four times.
Data are presented as the mean ± SD. Statistical analysis was
performed using one-way ANOVA followed by Tukey's post hoc test.
Statistical analyses were performed using Origin software (version
8.0; OriginLab). P<0.05 was considered to indicate a
statistically significant difference.
Results
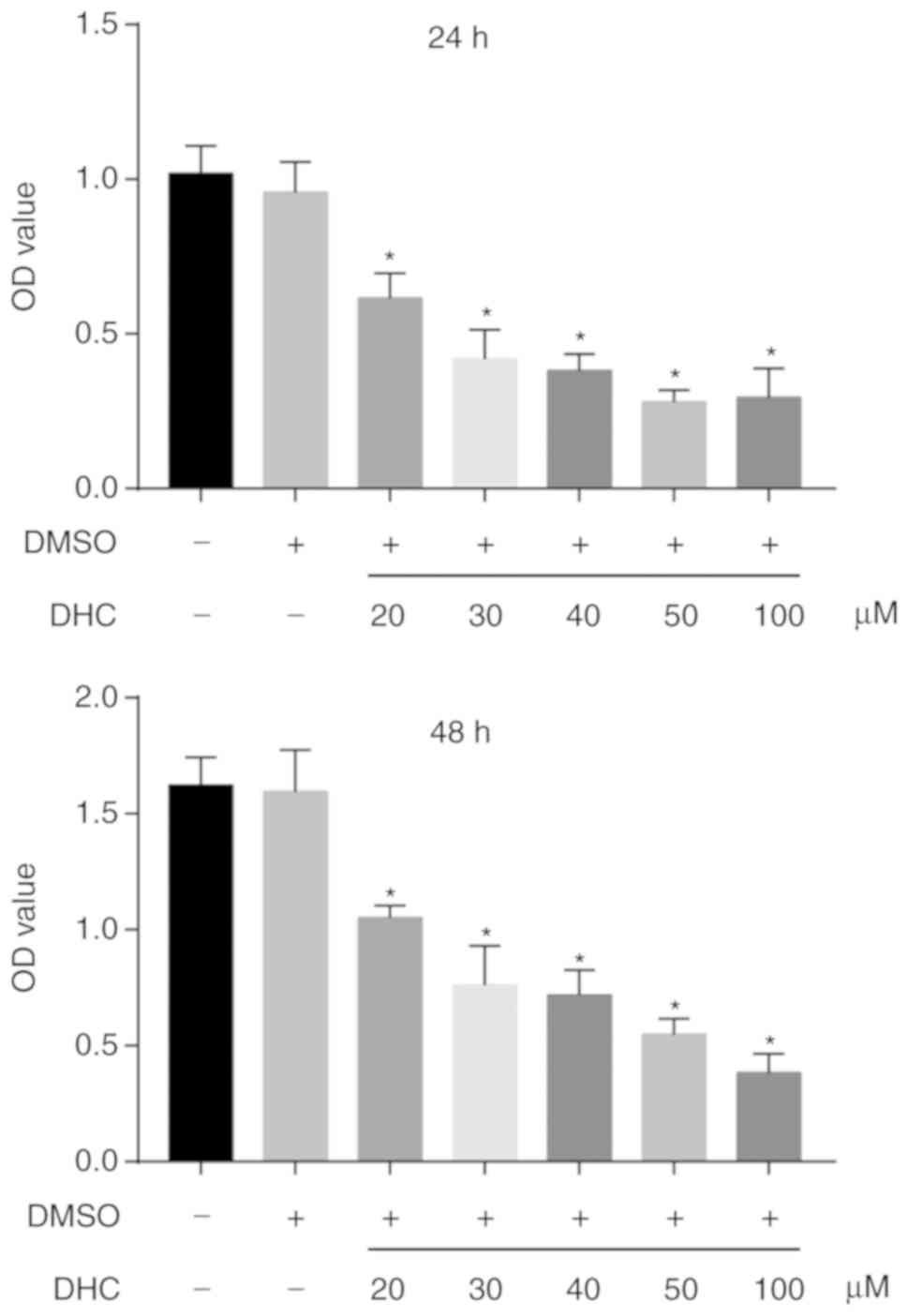
DHC inhibits the viability of
MDA-MB-231 cells
After treatment with various concentrations of DHC,
the viability of MDA-MB-231 cells decreased in a dose-dependent
manner. As revealed in the Fig. 1,
DMSO exerted no evident effect on the viability of MDA-MB-231
cells; cell viability was significantly decreased after treatment
with increasing concentrations of DHC. At 24 h, 20, 30, 40, 50, and
100 µM DHC decreased the viability of MDA-MB-231 cells from
101.02±9.04 to 61.24±8.62, 42.77±9.74, 40.13±5.37, 32.45±2.62, and
36.03±7.13, respectively (Fig. 1,
upper image). Moreover, treatment with 20, 30, 40, 50, and 100 µM
DHC for 48 h inhibited the viability of MDA-MB-231 cells from
159.72±9.87 to 103.58±1.92, 76.17±11.71, 75.06±9.32, 56.19±6.03,
and 46.18±7.74, respectively (Fig.
1, lower image).
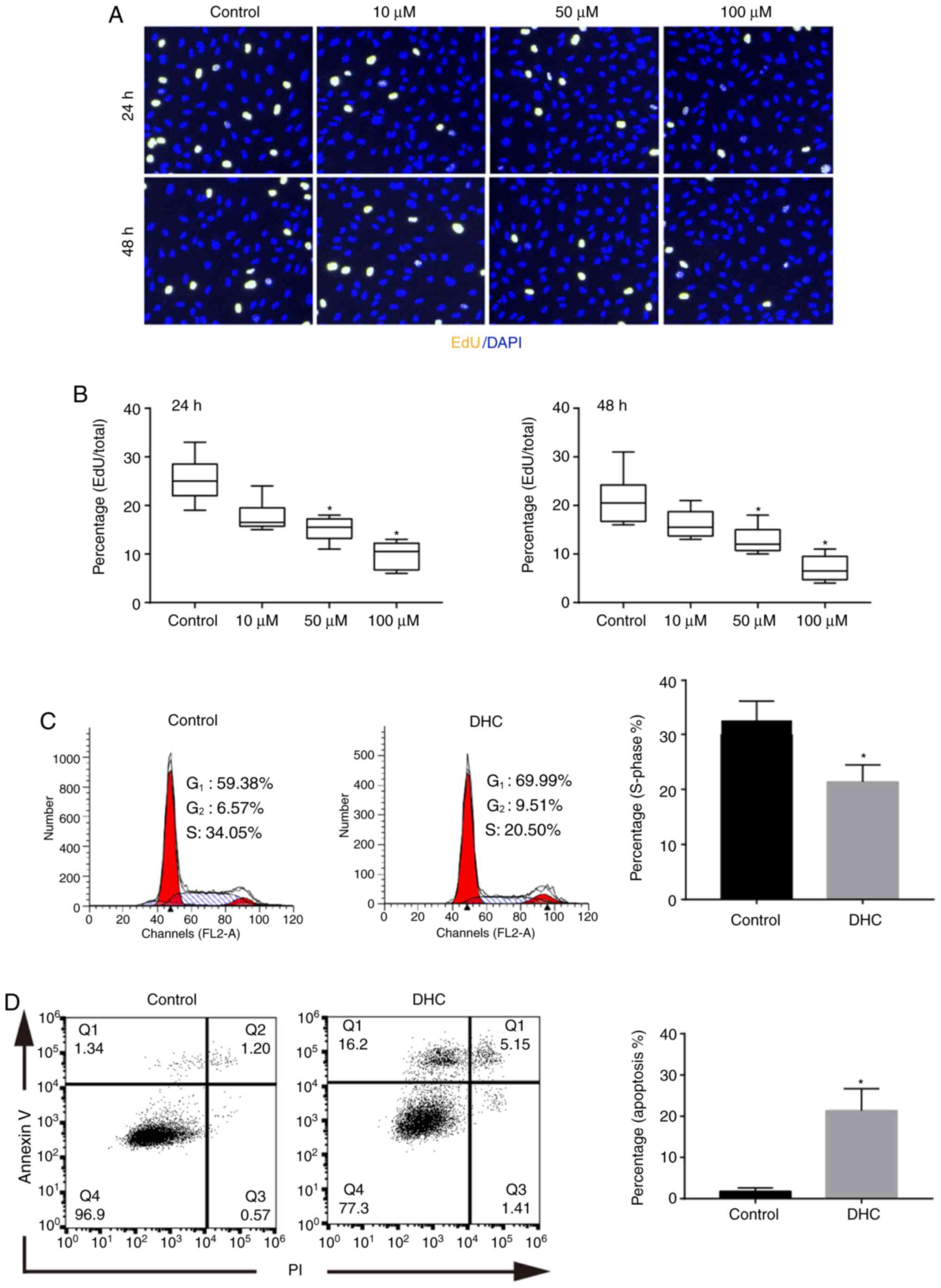
DHC inhibits cell proliferation and
promotes apoptosis of MDA-MB-231 cells
The results of the EdU staining and flow cytometry
both confirmed that DHC could effectively inhibit the proliferation
of MDA-MB-231 cells. After treatment with 10, 50, and 100 µM DHC
for 24 h, the percentages of the EdU-positive cells were
significantly decreased (Fig. 2A and
B). The cell proliferation percentage after 48 h of DHC
treatment exhibited a similar trend to that observed for 24 h
treatments (Fig. 2A and B). Flow
cytometric assay also reinforced the results obtained through EdU
staining assay. The number of MDA-MB-231 cells in the S-phase was
markedly decreased, and the statistical results indicated that the
percentage of cells in the S-phase was decreased from 32.87±4.63 to
21.03±3.81 (P<0.05) (Fig. 2C).
Furthermore, the effects of DHC on MDA-MB-231 cell apoptosis were
detected utilizing flow cytometry. After treatment with 50 µM DHC
for 48 h, the cell apoptosis rate was increased from 1.69±0.83 to
21.17±6.89 (P<0.05) (Fig.
2D).
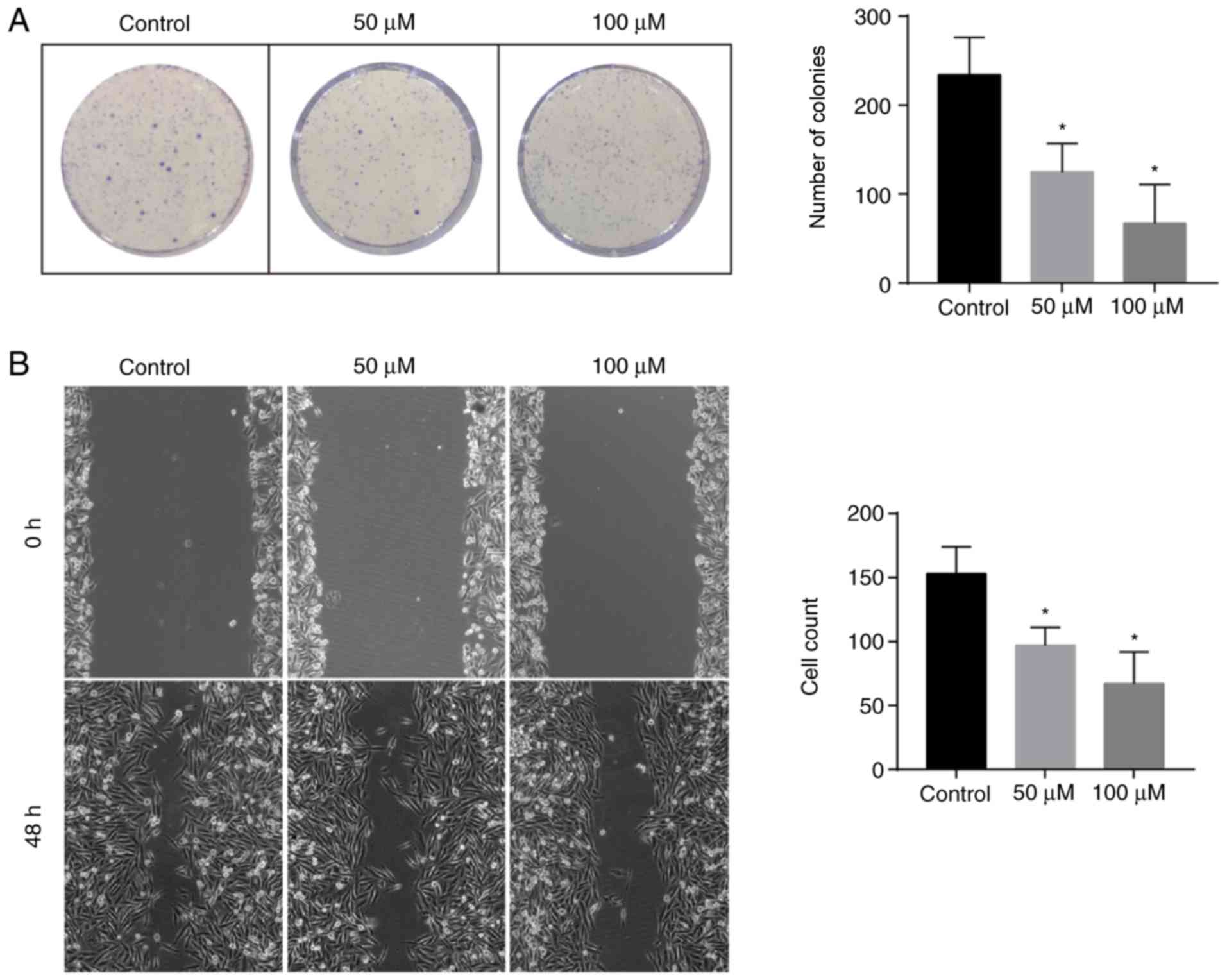
DHC inhibits the colony-forming
activity and migration of MDA-MB-231 cells
The colony-forming activity of breast cancer cells
could partially reflect the tumor malignancy. After treatment with
50 or 100 µM DHC for 12 days, the number of MDA-MB-231 colonies
decreased from 233.43±53.19 to 116.34±31.08 (P<0.05) and
69.17±55.83 (P<0.05), respectively (Fig. 3A). In addition, scratch assay
results indicated that treatment with both 50 and 100 µM DHC
decreased the migratory capacity of MDA-MB-231 cells, as the number
of migratory cells was reduced from 152.42±26.34 to 97.84±13.02
(P<0.05) and 66.34±28.37 (P<0.05), respectively (Fig. 3B).
DHC regulates the expression level of
cell proliferation-, migration- and apoptosis-associated genes
The expression levels of genes involved in cell
proliferation, migration and apoptosis was quantified after the
treatment of MDA-MB-231 cells with DHC. Both CDK1 and
CCND1 were critical proliferation-relevant genes, and both
were significantly downregulated after treatment with 20 and 50 µM
DHC for 48 h (Fig. 4A). Moreover,
MMP2 and MMP9 play a key role in tumor metastasis
through the degradation of the extra-cellular matrix. In this
study, DHC also inhibited the expression of MMP2 and
MMP9, both at the mRNA and protein level (Fig. 4A, C and E). Furthermore, the mRNA
levels of caspases 3/8/9, and the production of cleaved
caspases 3/9 were analyzed. As revealed in Fig. 4B, 20 and 50 µM DHC treatment
significantly increased the mRNA levels of caspases 3/8/9 by
48.93 (P<0.05) and 103.87 (P<0.05), 97.83 (P<0.05) and
147.64 (P<0.05), 26.13 (no significant difference) and 97.47
(P<0.05), respectively. Concerning the protein production
levels, the levels of activated caspase-3 and caspase-9 were both
decreased (P<0.05; Fig. 4D and
F).
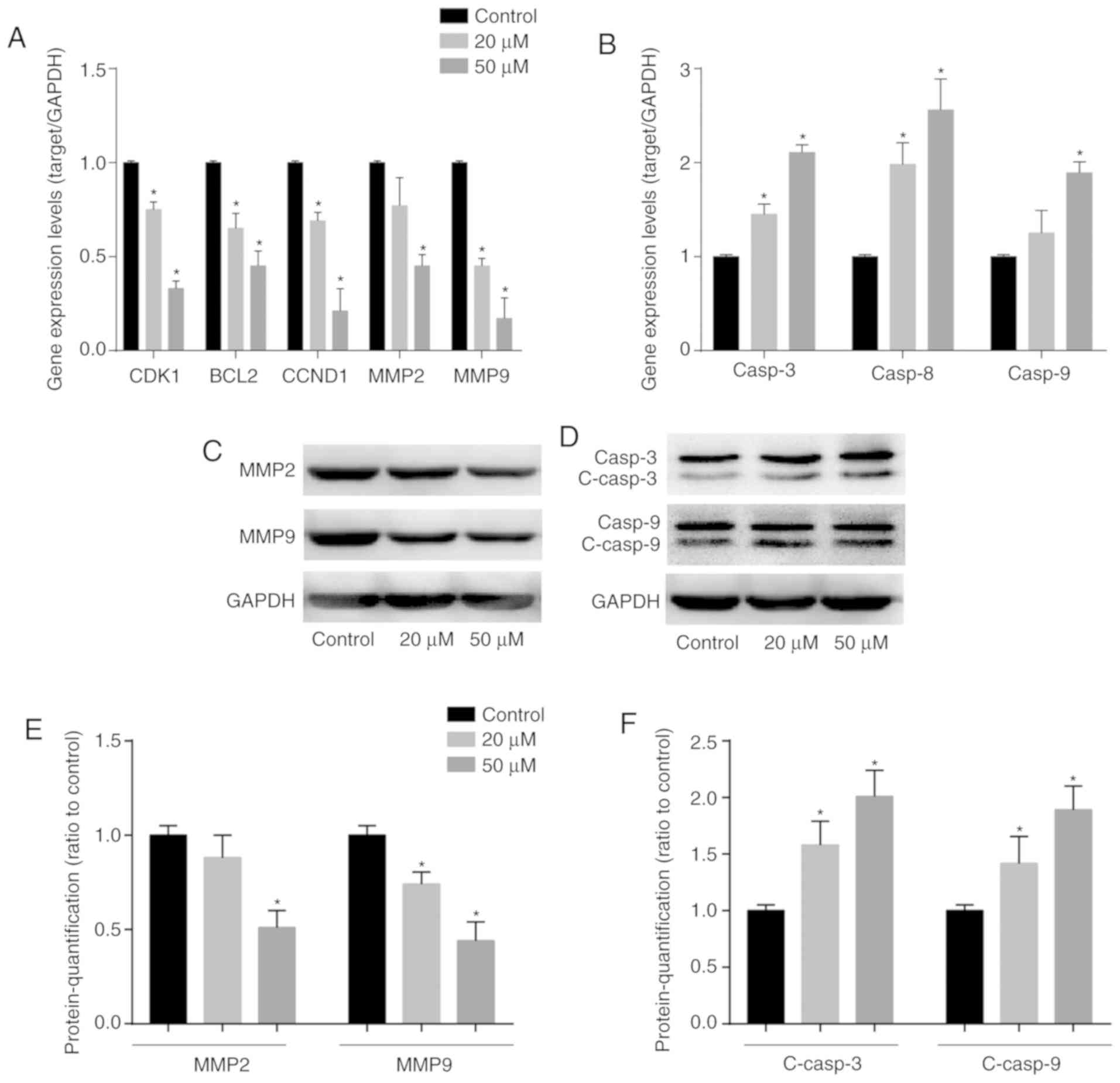 | Figure 4.DHC affects the expression levels of
oncogenes, apoptotic markers and MMPs. Reverse
transcription-quantitative PCR was used to analyze the expression
levels of (A) CDK1, BCL2, CCND1, MMP2, MMP9 and (B) caspase 3/8/9
of MDA-MB-231 cells treated with DHC (20 and 50 µM) for 48 h.
Western blotting was used to analyze the protein expression levels
of (C) MMP2, MMP9 and (D) caspase 3, caspase 9, cleaved caspase 3
and cleaved caspase 9 in MDA-MB-231 cells treated with DHC (20 and
50 µM) for 48 h. Quantification of the western blot analysis for
the protein expression of (E) MMP2 and MMP9 and (F) cleaved caspase
3 and cleaved caspase 9. *P<0.05 vs. the control group. DHC,
dehydrocorydaline. CDK1, cyclin-dependent kinases 1; CCND1, cyclin
D1; MMP, matrix metalloproteinase; casp, caspase; c, cleaved. |
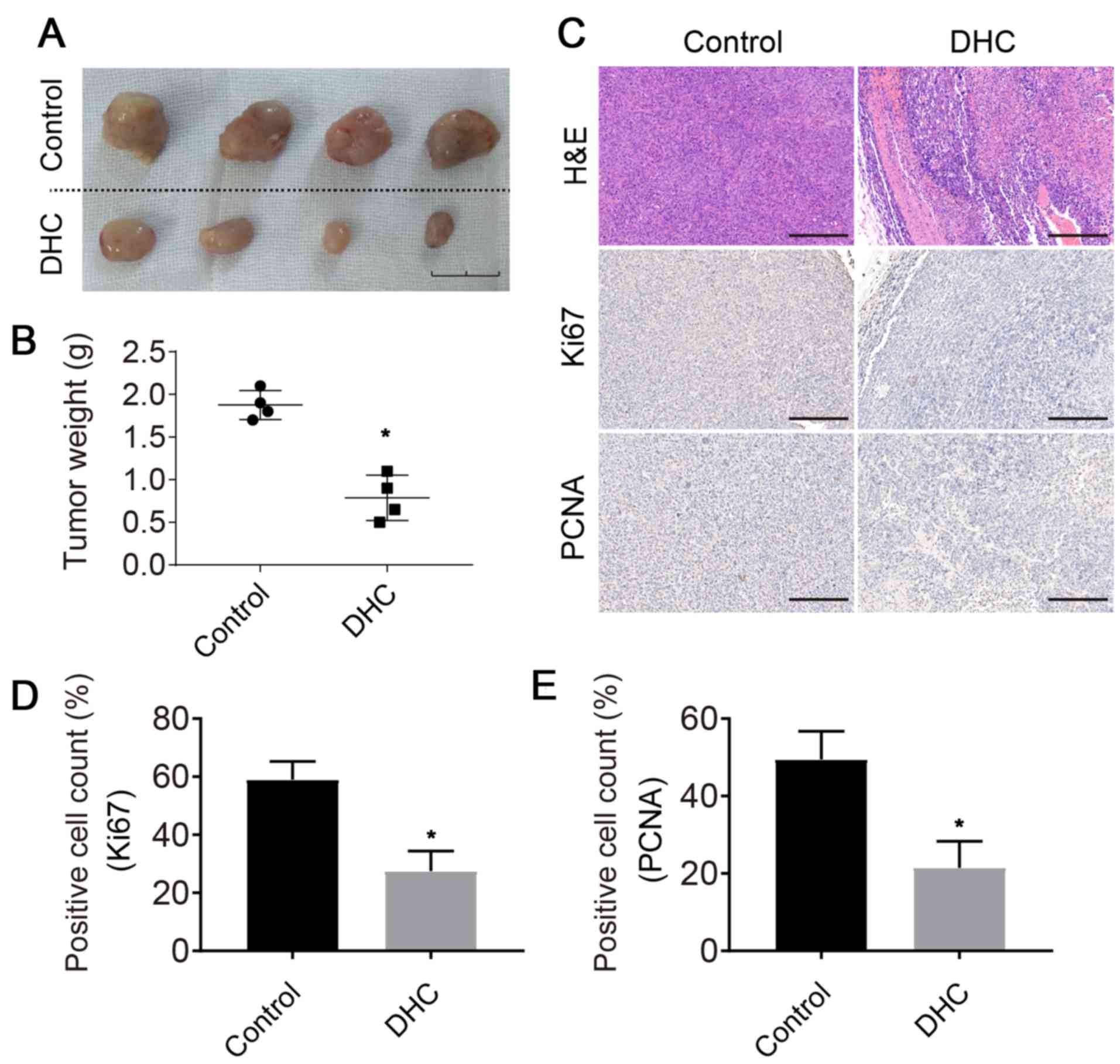
DHC inhibits the growth of MDA-MB-231
tumor xenografts in SCID mice
The results obtained in this study revealed that DHC
could reduce the tumor growth activity in vivo. After
subcutaneous injection for 3 weeks, the newly formed tumor tissues
were removed, photographed and weighed. The tumor volume was
significantly reduced after DHC treatment (Fig. 5A). In addition, the tumor weight
also decreased from 1.78±2.31 to 0.68±0.28 g (P<0.05; Fig. 5B).
DHC inhibits cell proliferation in
newly-formed tumor in vivo
The impact of DHC on the proliferation of MDA-MB-231
cells was detected through hematoxylin-eosin (H&E) staining,
and Ki67 and PCNA IHC. H&E staining revealed that the cell
density was markedly reduced in DHC groups (Fig. 5C). Furthermore, the expression
levels of Ki67 and PCNA were employed to evaluate cell
proliferation alterations. In this study, continuous treatment with
DHC limited cell proliferation in the newly formed tumors (Fig. 5C), therefore effectively hindering
tumor progression. Ki67- and PCNA-positive cells were counted. The
results revealed that Ki67- and PCNA-positive cell ratios were
significantly decreased from 59.12±5.32 to 26.47±5.92 (P<0.05)
and from 48.36±9.76 to 21.78±9.83 (P<0.05), respectively
(Fig. 5D and E).
Discussion
DHC exhibits antitumor effects and plays an active
role in inhibiting the proliferation and migration of cancer cells.
For example, Huo et al (8)
and other groups reported the effects of DHC on spinal bone cancer
pain and microglia polarization, since it had been revealed that
DHC could attenuate bone cancer pain. Lee et al (9) reported that DHC could decrease the
migration capacity of non-small cell lung carcinoma cells. However,
there are very few studies on the effects of DHC treatment in
breast cancer. Considering that DHC certainly has cytotoxic
activity, unraveling the mechanism of action through which DHC
inhibits the proliferation and migration of breast cancer cells is
of great interest for the clinical treatment of breast cancer.
A previous study (9) revealed that DHC impairs the migration
of non-small cell lung carcinoma cells by downregulating MMP
protein production, which is consistent with our findings. Our
experimental results confirmed that DHC inhibited the expression of
MMP2 and MMP9, thereby inhibiting the metastasis of
MDA-MB-231 cells. MMP2 and MMP9 are capable of degrading
extracellular matrices, and play a key role during tumor
metastasis. MMPs are a class of extracellular matrix-degrading
enzymes that are considered to be important enzymes for the
degradation of the extracellular matrix and basement membrane
(10). For instance, MMP2 plays an
important role in cell migration and tumor invasion (11), MMP7 inhibits cell adhesion
(12), and MMP9 is a prerequisite
for cell migration (13).
DHC-induced apoptosis may be the key activity
granting its anticancer efficacy. The present results revealed that
DHC treatment resulted in a significant decrease in the production
of the anti-apoptotic protein BCL2, and an increase in the
expression of the pro-apoptotic caspases 3/8/9. Flow cytometric
results revealed that the percentage of apoptotic breast cancer
cells increased after 48 h of treatment with 50 µM DHC.
Apoptosis-related proteins are classified into two types: An
anti-apoptotic class (that prevents apoptosis) and a pro-apoptotic
class (that induces apoptosis). BCL2 family proteins are critical
apoptosis-related proteins. Some authors have reported that BCL2
overexpression blocks the effects of DHC on cell proliferation and
migration, suggesting that BCL2-mediated anti-apoptotic activity
could underlie the DHC anticancer activity. Inhibition of BCL2
activates downstream caspase proteins and triggers cancer cell
apoptosis (14). The present
experimental results revealed that the effect of DHC treatment on
proliferation-related proteins CDK1 and CCDN1 was consistent with
the one exerted on BCL2.
Other studies have revealed that DHC inhibits the
proliferation of MCF-7 cells by modulating the Bax/Bcl-2-mediated
apoptosis and activating proteins belonging to the caspase family
(15). This indicates that
BCL2-mediated apoptosis plays an important role in the antitumor
effect of DHC. Caspase family proteins play an important role in
the induction, transduction, and amplification of intracellular
apoptotic signaling (16).
Caspase-8 and −10 are generally considered apoptotic activators in
the exogenous pathway, while caspase-9 and −2 play important roles
in the endogenous pathway, and Caspase-3 and −7 are the main
effectors of apoptosis (10).
Although DHC has the effect of inhibiting triple negative breast
cancer cells MDA-MB-231, it is unknown whether DHC exerts
inhibition on other types of human breast cancer cell lines such as
SK-BR-3, BT-20 and MCF7. Therefore, in a follow-up study,
comprehensive examination of the inhibitory effect of DHC on
different types of breast cancer will be performed.
In conclusion, DHC was revealed to inhibit the
migration and proliferation of MDA-MB-231 cells. In addition, DHC
also promoted the apoptosis of breast cancer cells by inhibiting
the production of BCL2 and upregulating the expression of caspases.
In vivo experiments confirmed that DHC could further slow
tumor progression by restricting angiogenesis at this level. The
present results indicated that DHC is a promising drug candidate
for breast cancer treatment.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Research
project of Wuhan health and family planning commission, 2014
(project leader, YH; grant no. WX14D04).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YH and GZ conceived and designed the study. YH and
HH analyzed the data, and wrote and revised the manuscript. SW and
FC analyzed and interpreted the data. GZ provided financial
support, and wrote and revised the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Fifth Hospital
of Wuhan.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CDK1
|
cyclin-dependent kinases 1
|
|
CCND1
|
cyclin D1
|
|
MMP
|
matrix metalloproteinase
|
|
PCNA
|
proliferating cell nuclear antigen
|
References
|
1
|
Park Y, Shackney S and Schwartz R:
Network-based inference of cancer progression from microarray data.
IEEE/ACM Trans Comput Biol Bioinforma. 6:200–212. 2009. View Article : Google Scholar
|
|
2
|
Ghoncheh M, Pournamdar Z and Salehiniya H:
Incidence and mortality and epidemiology of breast cancer in the
world. Asian Pac J Cancer Prev. 17:43–46. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mukherjee AK, Basu S, Sarkar N and Ghosh
AC: Advances in cancer therapy with plant based natural products.
Curr Med Chem. 8:1467–1486. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sun Y, Xun K, Wang Y and Chen X: A
systematic review of the anticancer properties of berberine, a
natural product from Chinese herbs. Anticancer Drugs. 20:757–769.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xu Z, Chen X, Fu S, Bao J, Dang Y, Huang
M, Chen L and Wang Y: Dehydrocorydaline inhibits breast cancer
cells proliferation by inducing apoptosis in MCF-7 cells. Am J Chin
Med. 40:177–185. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Masood A, Azmi AS and Mohammad RM: Small
molecule inhibitors of Bcl-2 family proteins for pancreatic cancer
therapy. Cancers (Basel). 3:1527–1549. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huo W, Zhang Y, Liu Y, Lei Y, Sun R, Zhang
W, Huang Y, Mao Y, Wang C, Ma Z and Gu X: Dehydrocorydaline
attenuates bone cancer pain by shifting microglial M1/M2
polarization toward the M2 phenotype. Mol Pain.
14:17448069187817332018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee J, Sohn EJ, Yoon SW, Kim CG, Lee S,
Kim JY, Baek N and Kim SH: Anti-metastatic effect of
dehydrocorydaline on H1299 non-small cell lung carcinoma cells via
inhibition of matrix metalloproteinases and B cell lymphoma 2.
Phyther Res. 31:441–448. 2017. View
Article : Google Scholar
|
|
10
|
Lu P, Takai K, Weaver VM and Werb Z:
Extracellular Matrix degradation and remodeling in development and
disease. Cold Spring Harb Perspect Biol. 3(pii):
a0050582011.PubMed/NCBI
|
|
11
|
Fromigué O, Hamidouche Z and Marie PJ:
Blockade of the RhoA-JNK-c-Jun-MMP2 cascade by atorvastatin reduces
osteosarcoma cell invasion. J Biol Chem. 283:30549–30556. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Basu S, Thorat R and Dalal SN: MMP7 is
required to mediate cell invasion and tumor formation upon
plakophilin3 loss. PLoS One. 10:e01239792015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dufour A, Zucker S, Sampson NS, Kuscu C
and Cao J: Role of matrix metalloproteinase-9 dimers in cell
migration: Design of inhibitory peptides. J Biol Chem.
285:35944–35956. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li WH, Wu HJ, Li YX, Pan HG, Meng T and
Wang X: MicroRNA-143 promotes apoptosis of osteosarcoma cells by
caspase-3 activation via targeting Bcl-2. Biomed Pharmacother.
80:8–15. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dolka I, Król M and Sapierzyński R:
Evaluation of apoptosis-associated protein (Bcl-2, Bax, cleaved
caspase-3 and p53) expression in canine mammary tumors: An
immunohistochemical and prognostic study. Res Vet Sci. 105:124–133.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Galluzzi L, Bravo-San Pedro JM and Kroemer
G: Ferroptosis in p53-dependent oncosuppression and organismal
homeostasis. Cell Death Differ. 22:1237–1238. 2015. View Article : Google Scholar : PubMed/NCBI
|