Introduction
Alzheimer's disease (AD), a chronic but irreversible
neurodegenerative disorder with the highest incidence of
age-related dementia, is mainly caused by abnormal aggregation of
the neurotoxic amyloid-β (Aβ) peptide, a product that forms
following proteolysis of the amyloid precursor protein (APP)
(1). Pathological characteristics
of AD often include neuronal apoptosis, synaptic loss and cognitive
impairment, which are considered to be associated with abnormal
aggregation of Aβ (2,3). Furthermore, it has been revealed that
increased Aβ formation results in high levels of reactive oxygen
species and an increased activation of proinflammatory cytokines,
associated with neuroinflammation in AD (4). Because abnormal accumulation of Aβ
plays a key role in the progression of AD, understanding how to
inhibit its aggregation warrants further study.
Astrocytes can provide energy to neurons through the
astrocyte-neuron shuttle (5),
thereby, affecting their metabolism and synaptic activity, which is
known to be important for memory formation (6,7).
Notably, Aβ can promote the expression of inflammatory factors and
inhibit the activity of Aβ-cleaving serine proteases in astrocytes
(8,9), which is associated with AD severity
(10).
The α7 nicotinic acetylcholine receptors (nAChRs)
are pentameric molecules that belong to the ligand-gated ion
channel family involved in regulating the permeability of
Ca2+, associated with neuroprotective and
anti-inflammatory effects in the AD brain (11). α7 nAChRs are abundantly expressed
in the central and peripheral nervous system, and spinal cord
(12). α7 nAChRs can form a stable
complex with Aβ in neuritic plaques and neurons that are associated
with the pathogenesis of AD, which results in attenuation of Aβ
neurotoxicity (13). The
expression of α7 nAChRs is significantly increased in the cerebral
cortex and hippocampus of patients with AD, which is closely
associated with Aβ deposits (14,15).
However, it is not clear whether the physiological functions of α7
nAChRs are related to Aβ aggregation and deposition in
astrocytes.
Heat shock proteins (HSPs), a conservative protein
family induced by heat shock and heavy metals, among others, are
widely expressed in prokaryotes and eukaryotes (3). HSPs, such as HSP-70 and αB-crystallin
(Cryab), can effectively suppress the aberrant accumulation of Aβs
(16). Moreover, Cryab combines
with Aβs, significantly inhibiting their accumulation and
neurotoxicity (17,18). Heat shock factor 1 (HSF-1) is
activated through multiple steps; it then translocates to the
nucleus, where it induces HSPs in response to stress by binding to
the heat shock element (HSE). HSF-1 protects neurons from death by
regulating the expression of HSPs (19). However, it is still not known
whether this protective effect is mediated by α7 nAChRs
stimulation.
In the present study, it was reported that
PNU282987, a potent agonist of α7 nAChRs, can significantly enhance
suppression of Aβ aberrant accumulation via the upregulation of
endogenous Cryab and HSP-70 in cultured astrocytes. In fact, it was
demonstrated that HSF-1 expression was necessary for the
upregulation of Cryab, but not for that of HSP-70. Furthermore, the
neuroprotective effect of PNU282987 against Aβ aggregation was
mediated via activation of the canonical PI3K/Akt signaling pathway
after α7 nAChRs.
Materials and methods
Materials
Fetal bovine serum (FBS) and Dulbecco's modified
Eagle's medium (DMEM) were obtained from Gibco; Thermo Fisher
Scientific, Inc. Synthetic drugs included Aβ1-42,
PNU282987, dimethylsulfoxide (DMSO) and methyllycaconitine (MLA)
(Sigma-Aldrich; Merck KGaA). The following antibodies were used in
the present study: Mouse polyclonal anti-Aβ, 1–16 (cat. no. 803015;
BioLegend, Inc.), rabbit polyclonal anti-HSP-70 (cat. no. AB9920;
Sigma-Aldrich; Merck KGaA), anti-Cryab (cat. no. ab13496; Abcam),
anti-β-actin (cat. no. sc-47778; Santa Cruz Technology, Inc.),
rabbit monoclonal anti-phospho-(cat. no. 7252C) and
non-phospho-specific Akt (cat. no. 2920) and anti-HSF-1 (cat. no.
12972; Cell Signaling Technology, Inc.), and peroxidase-conjugated
secondary antibodies (cat. no. sc-2357; Santa Cruz Technology,
Inc.). Phosphate-buffered saline (PBS), trypsin and
penicillin-streptomycin were obtained from HyClone; GE Healthcare
Life Sciences, Lipofectamine® 2000 was purchased from
Invitrogen; Thermo Fisher Scientific, Inc., and the LY294002
inhibitor was obtained from Cell Signaling Technology, Inc. Culture
flasks were purchased from Corning, Inc. The enhanced
chemiluminescence system (ECL) was obtained from Amersham; GE
Healthcare, and polyvinylidene fluoride (PVDF) membranes were
purchased from EMD Millipore. Sprague-Dawley (SD) pregnant rats
were purchased from the Animal Experiment Center of Guizhou Medical
University (Guizhou, China).
Primary astrocyte cultures
All animal experiments in the present study were
approved by the Animal Care Committee of Guizhou Medical University
and were implemented in strict accordance with the relevant
guidelines (no. 1503008). A total of 20 SD rats (8 weeks old,
weight 240–280 g) were randomly divided into two groups, 10 (5
males and 5 females) in each group. Rats had access to food and
water ad libitum and were maintained at 25°C (relative
humidity 65%) with a 12-h light/dark cycle. Astrocytes were
separated from the cerebral cortex of newborn SD rats according to
a previously described method (20). Briefly, the cerebral cortex was
separated and cut, a 10X volume of trypsin digestion buffer was
added for 15 min at 37°C. Samples were centrifuged at 1,006.2 × g,
25°C for 5 min in complete medium (DMEM with 10% FBS and 1%
penicillin-streptomycin). Cells were then resuspended with complete
medium and transferred to 25-cm2 culture flasks for ~9
days. Cells were then purified by centrifugal methods (83.8 × g,
37°C, 6 h). Purified cells were sub-cultured in 25 cm2
culture flasks (37°C, 5% CO2). In this way, ~95% of the
astrocytes could be purified, and they were identified using
immunostaining with rabbit anti-GFAP antibody (1:200) and
anti-rabbit IgG Cy3 (1:200). Rats were placed into enclosed flow
cages for 5 min, and then 100% CO2 was infused at a 30%
volume per minute displacement, in order to euthanize the animals.
Subsequently, rats were rendered unconscious by CO2
inhalation (confirmed by slow deep breathing and absence of
response to toe-pinch).
Preparation of Aβ oligomers
Aβ oligomers were prepared according to a previous
method (21). Briefly, Aβ peptides
were dissolved in hexafluoroisopropanol (HFIP) at a concentration
of 1 mM. After evaporating HFIP, the peptide membrane was stored at
−80°C. Aβ peptides were dissolved with DMSO and diluted to 1 µm
with DMEM under sterile conditions. The quality of Aβ oligomers was
identified by 12 alkyl sulfate polyacrylamide gel electrophoresis
(SDS-PAGE), as previously described (22).
Treatment of astrocytes
To evaluate the role of PNU282987 in HSP regulation,
cultured astrocytes were treated with PNU282987 (5 µM) at different
time-points (6, 12, 18, 24 h) in an incubator at 37°C. The
regulatory effect of PNU282987 was partly inhibited by MLA (100
nM,), an α7 nAChR antagonist, at 37°C for 2 h. The concentration of
MLA used was based on previous studies (23,24).
To explore whether the regulatory effect of PNU282987 was mediated
by the PI3K signaling pathway, astrocytes were pretreated with
LY294002 (10 µM) for 2 h at 37°C before PNU282987 treatment. The
concentration of LY294002 was based on previous studies (19,20).
The medium was replaced by DMEM containing PNU282987 (5 µM), and
the astrocytic cultures were incubated for 18 h at 37°C. After
removing the medium, astrocytes were further exposed to DMEM
containing Aβ oligomers (1 µM) for 24 h at 37°C.
Western blot analysis
Cells were washed three times with pre-chilled PBS
in a six-well plate; then, cells were incubated on ice for 3 h with
60 µl/well of RIPA buffer containing protease and phosphatase
inhibitors (cat. no. 9806S; Cell Signaling Technology., Inc.). The
supernatant was collected after centrifugation at 4°C for 15 min at
4,024.8 × g, and the concentration of the supernatant protein was
assessed with a BCA assay (Thermo Fisher Scientific, Inc.),
according to the manufacturer's instructions. Proteins were loaded
(10 µg/lane) and resolved in 12% SDS-PAGE gels. Proteins were
transferred to nitrocellulose membranes, and then blocked with 5%
non-fat milk at room temperature (RT) for 2 h. Membranes were
washed three times with TBS buffer with 0.1% Tween-20 (TBST);
subsequently, the proteins were incubated with various primary
antibodies overnight at 4°C. Antibodies included: Anti-phospho-Akt
(Ser 473; 1:1,000); anti-HSF-1 (1:1,000); anti-Cryab (1:1,000);
anti-β-Amyloid 1–16 (1:1,000); anti-HSP-70 (1:1,000); or
anti-non-phospho-Akt (1:1,000). Following the overnight incubation,
the membranes were rinsed three times with TBST (5 min each) and
proteins were incubated with the appropriate secondary antibody for
1 h at RT. GAPDH (1:1,000; cat. no. MA5-157381; Thermo Fisher
Scientific, Inc.) or β-actin (1:1,000; cat. no. sc-47778; Santa
Cruz Biotechnology, Inc.) were used as internal controls. Protein
bands were detected with an ECL kit, according to the
manufacturer's instructions. Quantification was performed using
ImageJ software (version 1.46; National Institutes of Health).
Transfections
Cell transfections were carried out according to
previously described protocols (25,26).
Briefly, cells were cultured in six-well plates for 24 h at a
density of 5×105 cells and allowed to adhere for 24 h.
The target sequence of the rat HSF-1 shRNA plasmid (Qiagen,
Inc.) was 5′-TGTCAACAAGCTCATCCAATT-3′. Scrambled shRNA (Qiagen,
Inc.) or HSF-1-specific shRNA (2 µg), plus reagents (2 µl),
and Lipofectamine 2000 (6 µl) were mixed with Opti-MEM medium (500
µl; cat. no. 31985062; Thermo Fisher Scientific, Inc.) for 20 min.
Cells were incubated at 37°C for 48 h in this medium containing
shRNA. Subsequently, the cells were washed 3 times with PBS and
fresh medium was added. The transfected cells were seeded in the
selection medium (containing Hygromycin 340 µg/ml) for 12 days.
Well-separated antibiotic resistant clone cells were identified and
expanded for subsequent experiments. The identified cells were
treated with PNU (5 µM) for 18 h at 37°C. The time interval between
transfection and subsequent experimentation was 15 days.
Statistical analysis
To compare differences among groups, one-way ANOVA
was used with Tukey's post hoc test for multiple comparisons. Data
results were analyzed with SPSS 22 software (IBM Corp.). P<0.05
was considered to indicate a statistically significant
difference.
Results
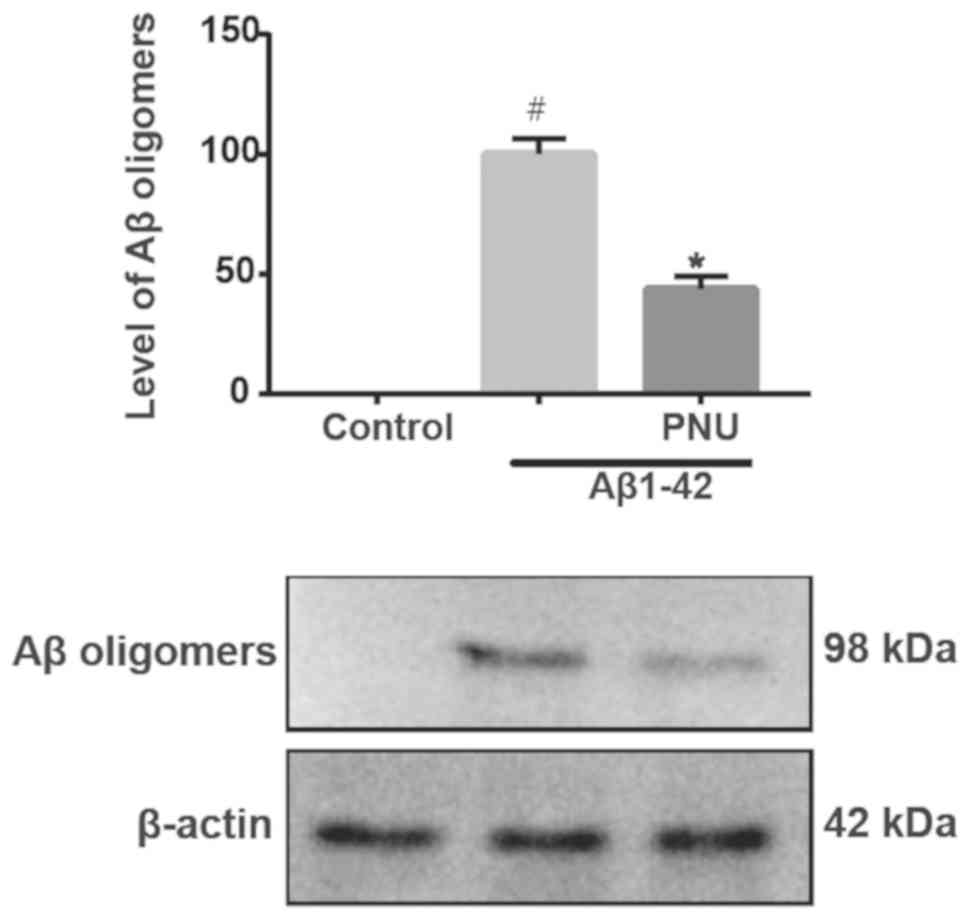
PNU282987 can enhance astrocytes to
inhibit Aβ aggregation
Pretreatment with 5 µM of the potent agonist of α7
nAChRs PNU282987 for 18 h followed by treatment with l µM
Aβ1-42 for 24 h significantly inhibited Aβ accumulation,
indicating that PNU282987 enhanced astrocytes to inhibit Aβ
aggregation (Fig. 1). The control
was used as the healthy control (without any treatment.), and the
Aβ1-42 treatment group was employed as the
PNU282987-treated control.
PNU282987 upregulates HSF-1, Cryab and
HSP-70 expression in astrocytes
To evaluate the effect of PNU282987 on the HSP
response, astrocytes were treated with 5 µM PNU282987 at various
time-points (0, 6, 12, 18 and 24 h). This resulted in the
upregulation of HSF-1, Cryab and HSP-70 (Fig. 2A and B). The upregulatory effect of
PNU282987 was time-dependent and peaked at 18 h. Therefore, this
time-point was selected for subsequent experiments.
PNU282987-mediated upregulation of endogenous HSF-1, Cryab and
HSP-70 was significantly antagonized by the α7 nAChR antagonist MLA
(0.1 µM), indicating that α7 nAChR was involved in the upregulatory
effect of PNU282987 (Fig. 2C and
D). Western blot analysis revealed that the HSF-1 knockdown
(KD) effectively decreased Cryab expression at 18 h after PNU282987
treatment. However, HSP-70 expression did not exhibit a significant
change in HSF-1-KD cells compared with negative control (NC;
scrambled shRNA) cells (Fig. 2E and
F).
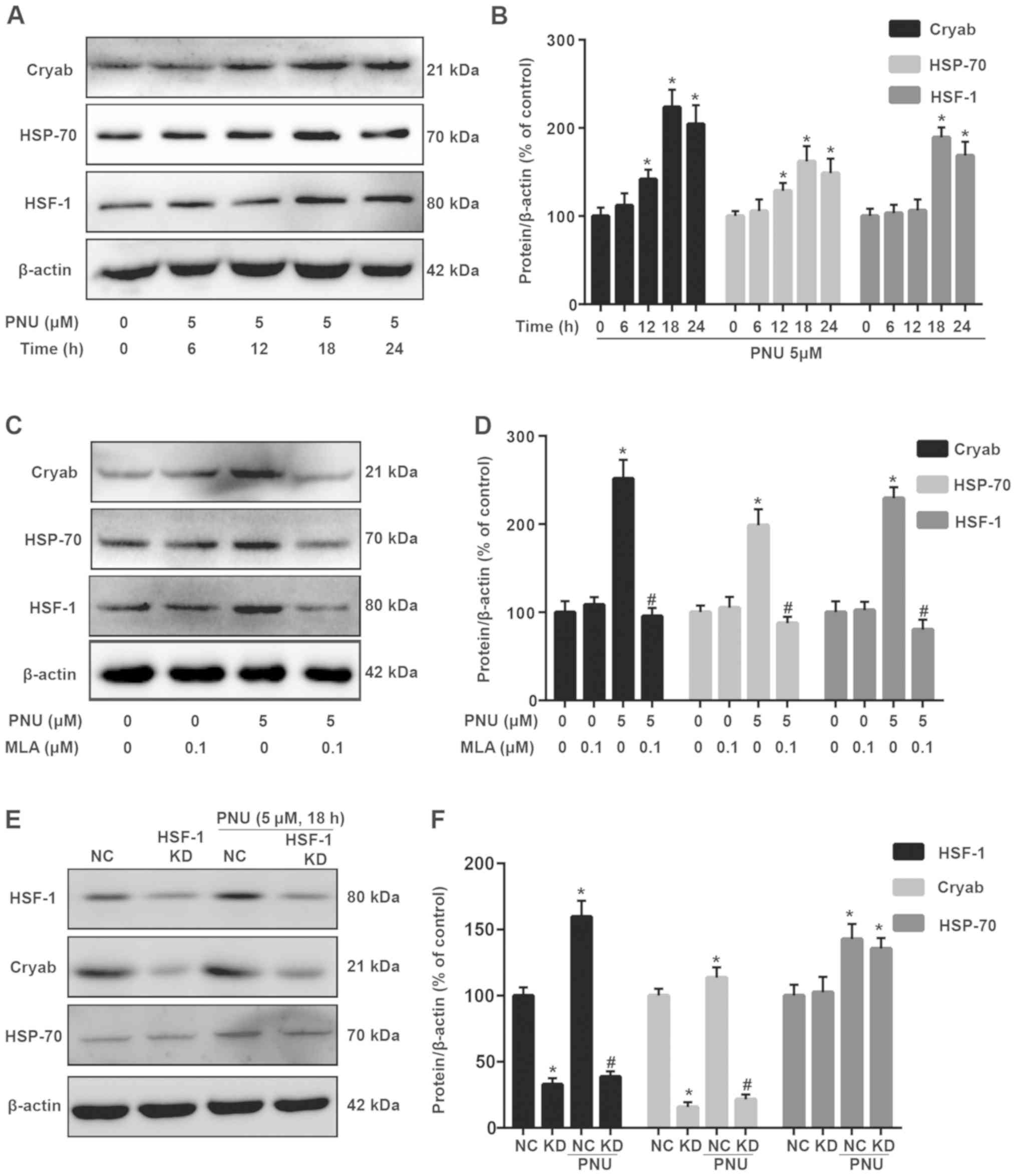 | Figure 2.HSP reaction is activated in
PNU-treated astrocytes. (A) Detection of Cryab, HSP-70 and HSF-1
expression levels in cells (treated with PNU at different
time-points) using western blot analysis. (B) Statistical analysis
of A. (C) Detection of Cryab, HSP-70 and HSF-1 expression in cells
(treated with PNU + MLA) using western blot analysis. (D)
Statistical analysis of C. (E) Detection of Cryab, HSP-70 and HSF-1
expression in cells (treated with PNU + HSF-1 shRNA) using western
blot analysis. NC indicates scramble shRNA. (F) Statistical
analysis of E. β-actin was used as an internal loading control.
SPSS was used for statistical analysis. Data are presented as the
mean ± SD; each experiment was repeated independently three times.
*P<0.05 vs. the control group; #P<0.05 vs. the PNU
pre-treatment group. HSP, heat shock protein; PNU, PNU282987;
Cryab, αb-crystallin; HSP-70, heat shock protein 70; HSF-1, heat
shock factor 1; MLA, methyllycaconitine; shRNA, short hairpin RNA;
NC, negative control. |
PNU282987 upregulates HSF-1, Cryab and
HSP-70 via the PI3K/Akt signaling pathway
PNU282987 upregulated p-Akt levels at various
time-points (0, 5, 10, 20 and 30 min) (Fig. 3A). Moreover, pre-treatment with the
PI3K inhibitor LY294002 (10 µM) or the α7 nAChRs inhibitor MLA (0.1
µM) for 2 h, followed by co-incubation with PNU282987 (5 µM),
indicated that MLA and LY294002 could significantly inhibit p-Akt
upregulation during PNU282987 treatment of astrocytes (Fig. 3B and C). Moreover, pre-treatment
with LY294002 for 2 h significantly inhibited the upregulation of
HSF-1, Cryab and HSP-70 during PNU282987 treatment of astrocytes
(Fig. 3D and E), indicating the
involvement of the PI3K pathway in the upregulatory effect of
PNU282987 (PNU282987 upregulated the expression of HSF-1, Cryab and
HSP-70).
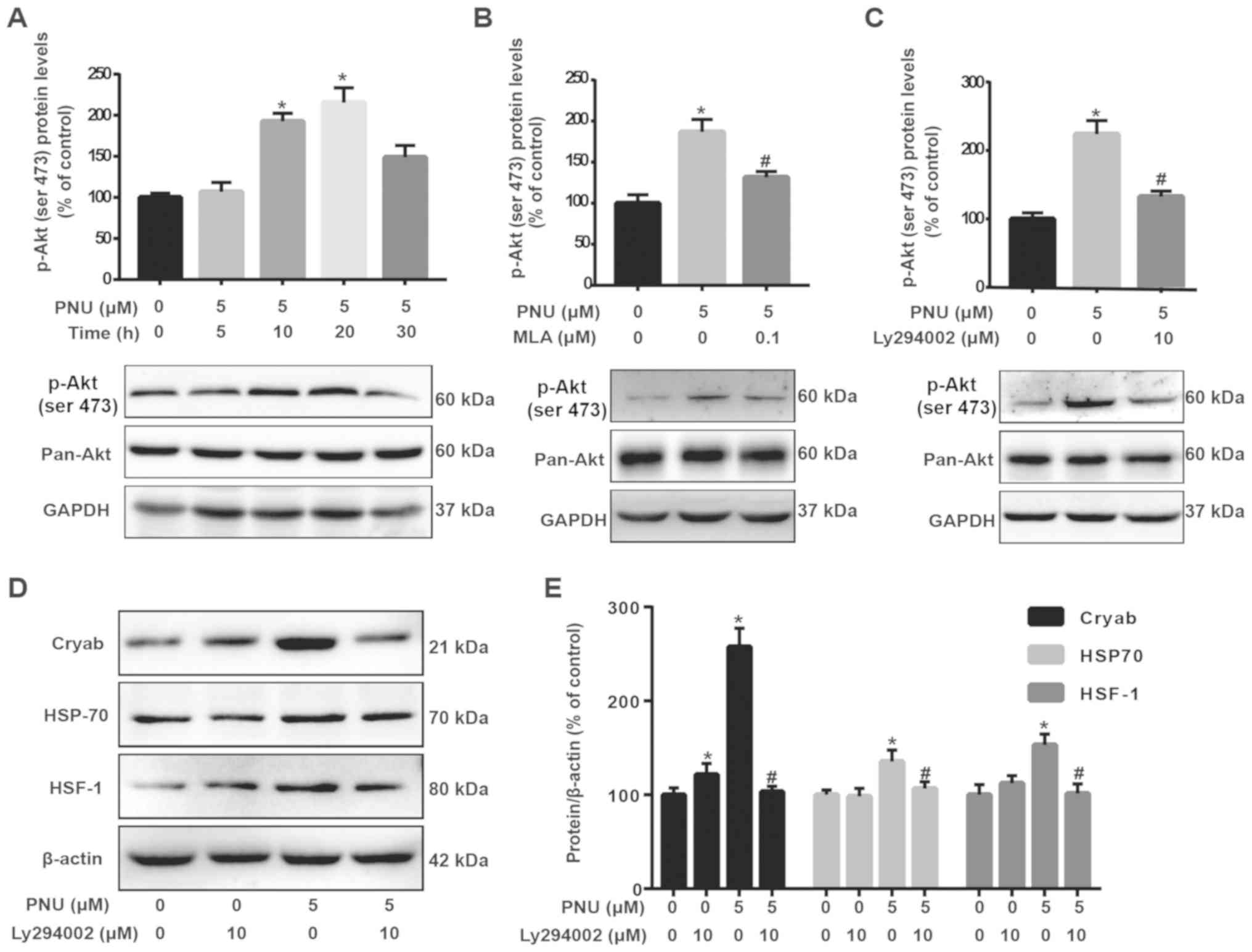 | Figure 3.PNU activates the PI3K/Akt pathway
via α7 nAChRs. (A) Cells treated with 5 µM PNU for 0, 5, 10, 20 and
30 min. (B) Cells were pre-treated with MLA (0.1 µM) for 2 h, and
then co-incubated with PNU (5 µM) for another 20 min. (C) Cells
were pre-treated with LY294002 (10 µM) for 2 h, and then
co-incubated with PNU (5 µM) for another 20 min. Cell lysates were
collected for immunoblot analysis of p-Akt and pan-Akt; β-actin was
used as an internal loading control. ImageJ was used to analyze the
protein expression of p-Akt and pan-Akt, and SPSS for statistical
analysis. (D and E) Pre-treatment with LY294002 for 2 h
significantly inhibited the upregulation of HSF-1, Cryab and HSP-70
during PNU treatment of astrocytes. Data are presented as the mean
± SD; each experiment was repeated independently three times.
*P<0.05 vs. the control group; #P<0.01 vs. the
PNU282987 only treatment group. PNU, PNU282987; Cryab,
αb-crystallin; HSP-70, heat shock protein 70; HSF-1, heat shock
factor 1; p-, phosphorylated; pan-, non-phosphorylated. |
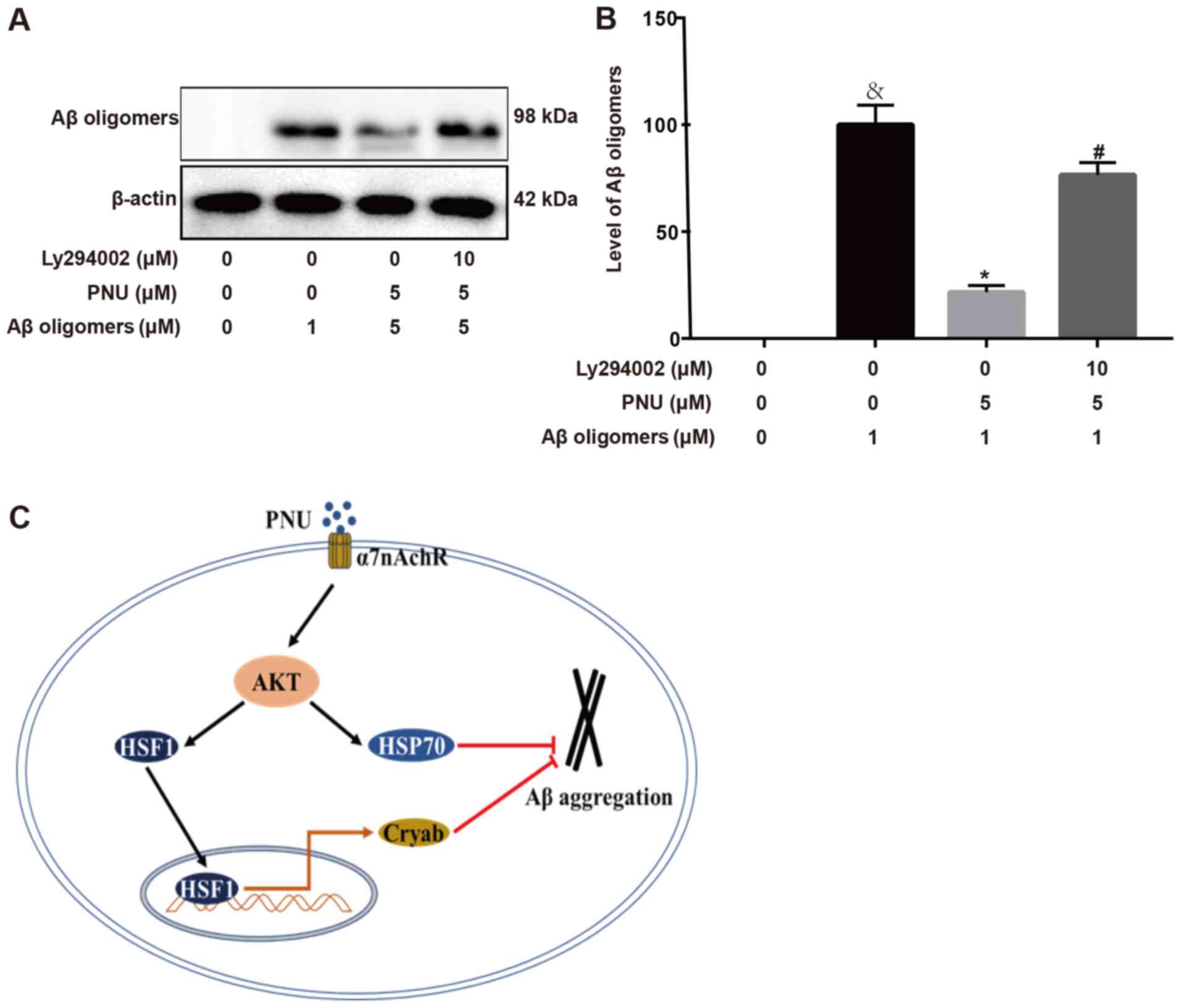
PNU282987 inhibits Aβ aggregation via
the PI3K/Akt signaling pathway
Next, it was explored whether the PI3K/Akt pathway
was involved in the inhibition of Aβ accumulation during
PNU282987-treatment of astrocytes. The results revealed that
PNU282987 treatment led to the inhibition of Aβ aggregation in
astrocytes; however, pre-treatment with LY294002 for 2 h
significantly attenuated this effect (Fig. 4A and B). The proposed pathway by
which PNU282987 regulates Aβ accumulation in astrocytes is
presented in Fig. 4C.
Discussion
PNU282987 can inhibit Aβ aggregation in astrocytes;
however, the specific mechanism underlying this effect remains to
be clarified. The present results indicated that the
PNU282987-mediated neuroprotection against Aβ accumulation was
associated with upregulation of endogenous Cryab and HSP-70 in
astrocyte cultures. Moreover, upregulation of astrocytic Cryab
depended on HSF-1, and HSF-1 KD effectively decreased Cryab
expression. These effects were regulated by α7 nAChRs and the
PI3K/Akt signal transduction pathway in astrocyte cultures.
α7 nAChRs are ion channel receptors associated with
memory and cognitive function in AD and Parkinson's disease (PD)
(27,28). These receptors are widely expressed
in the human cerebral cortex, especially in astrocytes and
microglia. α7 nAChRs are activated in neurons with simvastatin via
the calmodulin-kinase II signaling pathway (29). Activation of α7 nAChRs has been
revealed to protect neurons from the neurotoxic anticancer drug
oxaliplatin (30). Hence,
PNU282987, an α7 nAChR agonist, was employed to explore its
neuroprotective effect in Aβ-treated astrocytes. PNU282987 enhanced
astrocytes to inhibit Aβ aggregation, a process closely associated
to the upregulation of HSP.
Aβ aggregates and deposits are associated with
cognitive dysfunction, neuronal death and synaptic impairment in
patients with AD (3,31,32).
Previous studies have revealed a significant upregulation of α7
nAChR in astrocytes of AD brains that is closely associated with Aβ
accumulation (14,15). Moreover, α7 nAChRs have been
revealed to interact with Aβ (33). However, the relationship between α7
nAChRs and Aβ aggregation is still not clear in astrocytes. The
present results indicated the involvement of α7 nAChRs and the
PI3K/Akt signaling pathway in PNU282987-mediated protection against
Aβ neurotoxicity, and the upregulation of endogenous Cryab and
HSP-70 to inhibit Aβ aggregation in astrocytes. Moreover, HSF-1 KD
effectively decreased Cryab expression. Therefore, HSF-1 may
specifically activate the expression of Cryab. Cryab can
effectively prevent the accumulation and cytotoxicity of Aβs by
directly combining with them; in addition HSP-70 can inhibit the
production of Aβs by combining directly with APP (17,34).
Subsequently, HSF-1 becomes activated in in vitro models of
PNU282987 neuroprotective activity, causing the upregulation of
Cryab. Furthermore, in the present study, HSF-1 KD was associated
with a notable decrease in Cryab in both control and
PNU282987-treated astrocytes but did not affect HSP-70 expression.
Collectively, the present results indicated that the increase of
Cryab and HSP-70 may play a key role in the PNU282987-induced
neuroprotection against Aβ aggregation in in vitro models,
and the upregulation of Cryab seems to be mediated by HSF-1. The
present results indicated that Cryab could directly combine with
Aβs, preventing their aggregation and cytotoxicity, which is
consistent with past research (17).
Previous studies have suggested an increased
activation of the PI3K/Akt pathway during Tau hyperphosphorylation
in AD mice (35). However, it is
unclear whether the PI3K/Akt pathway is activated as a result of a
neuroprotective heat shock response elicited by PNU282987. The
present results revealed an increase in Akt phosphorylation in
astrocytes during PNU282987 treatment. However, pre-treatment with
the PI3K inhibitor LY294002 weakened the upregulatory effect of
PNU282987. Moreover, pre-treatment with the PI3K inhibitor for 2 h
significantly inhibited the PNU282987-mediated upregulation of
endogenous HSF-1, Cryab and HSP-70 in astrocytes. In addition,
HSF-1 KD was associated with a marked decrease in Cryab in both the
negative control and HSF-1-KD cells, which supports a role of Cryab
in regulating the protective effect of HSF-1 during the treatment
of cells with PNU282987.
In conclusion, in response to stress or injury
induced by Aβ, astrocytes may activate heat shock response for
adaptation and cell survival. The present study provided evidence
that PNU282987 can significantly enhance astrocytes to inhibit Aβ
accumulation by the activation of α7 nAChRs and the PI3K/Akt
signaling pathway. It was further verified that upregulation of
endogenous HSF-1, Cryab and HSP-70 is one of the heat shock
response mechanisms for astrocyte protection or adaption during
PNU282987 treatment. Collectively, these results demonstrated the
neuroprotective actions of PNU282987 in vitro, which will
guide future studies exploring this mechanism in AD animal
models.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (Project no.
81360199), a Special Grant of the Central Government Supporting
Local Science and Technology Development, the Science and
Technology Department of Guizhou Province [Guizhou specific grant
(2019) 4008], the Science and Technology Fund Project of Guizhou
Health and Health Commission (grant no. gzwjkj2019-1-039), and the
Science and Technology Fund Project of Southwest Guizhou Autonomous
Prefecture (2019-1-10).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
The first two authors contributed equally to this
work. WY and ZR designed the study, ZR wrote and revised the
manuscript. ZD analyzed the data and revised the manuscript. JL, PX
and YH performed the experiments. CZ and ZG helped perform the
analysis of the data with constructive discussions. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments in the present study were
approved by the Animal Care Committee of Guizhou Medical University
and were implemented in strict accordance with the relevant
guidelines (no. 1503008).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
α7 nAChR
|
α7 nicotinic acetylcholine
receptor
|
|
Aβ
|
amyloid-β
|
|
AD
|
Alzheimer's disease
|
|
APP
|
amyloid precursor protein
|
|
Cryab
|
αB-crystallin
|
|
HSF-1
|
heat shock factor 1
|
|
HSE
|
heat shock element
|
|
HSP
|
heat shock protein
|
|
KD
|
knock-down
|
|
MLA
|
methyllycaconitine
|
References
|
1
|
Schilling LP, Pascoal TA, Zimmer ER,
Mathotaarachchi S, Shin M, de Mello Rieder CR, Gauthier S, Palmini
A and Rosa-Neto P; Alzheimer's Disease Neuroimaging Initiative, :
Regional Amyloid-β load and white matter abnormalities contribute
to hypometabolism in Alzheimer's dementia. Mol Neurobiol.
56:4916–4924. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Karran E, Mercken M and De Strooper B: The
amyloid cascade hypothesis for Alzheimer's disease: An appraisal
for the development of therapeutics. Nat Rev Drug Discov.
10:698–712. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Glabe CG and Kayed R: Common structure and
toxic function of amyloid oligomers implies a common mechanism of
pathogenesis. Neurology. 66 (2 Suppl 1):S74–S78. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Saito T, Hisahara S, Iwahara N, Emoto MC,
Yokokawa K, Suzuki H, Manabe T, Matsumura A, Suzuki S, Matsushita
T, et al: Early administration of galantamine from preplaque phase
suppresses oxidative stress and improves cognitive behavior in
APPswe/PS1dE9 mouse model of Alzheimer's disease. Free Radic Biol
Med. 145:20–32. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sidoryk-Wegrzynowicz M, Wegrzynowicz M,
Lee E, Bowman AB and Aschner M: Role of astrocytes in brain
function and disease. Toxicol Pathol. 39:115–123. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Alberini CM, Cruz E, Descalzi G, Bessieres
B and Gao V: Astrocyte glycogen and lactate: New insights into
learning and memory mechanisms. Glia. 66:1244–1262. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Steinman MQ, Gao V and Alberini CM: The
role of lactate-mediated metabolic coupling between astrocytes and
neurons in long-term memory formation. Front Integr Neurosci.
10:102016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nielsen HM, Veerhuis R, Holmqvist B and
Janciauskiene S: Binding and uptake of A beta1-42 by primary human
astrocytes in vitro. Glia. 57:978–988. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pihlaja R, Koistinaho J, Malm T, Sikkilä
H, Vainio S and Koistinaho M: Transplanted astrocytes internalize
deposited beta-amyloid peptides in a transgenic mouse model of
Alzheimer's disease. Glia. 56:154–163. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jabbari Azad F, Talaei A, Rafatpanah H,
Yousefzadeh H, Jafari R, Talaei A and Farid Hosseini R: Association
between Cytokine production and disease severity in Alzheimer's
disease. Iran J Allergy Asthma Immunol. 13:433–439. 2014.PubMed/NCBI
|
|
11
|
Hoskin JL, Al-Hasan Y and Sabbagh MN:
Nicotinic acetylcholine receptor agonists for the treatment of
Alzheimer's dementia: An update. Nicotine Tob Res. 21:370–376.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Broide RS, Winzer-Serhan UH, Chen Y and
Leslie FM: Distribution of α7 nicotinic acetylcholine receptor
subunit mRNA in the developing mouse. Front Neuroanat. 13:762019.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Parri HR, Hernandez CM and Dineley KT:
Research update: Alpha7 nicotinic acetylcholine receptor mechanisms
in Alzheimer's disease. Biochem Pharmacol. 82:931–942. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu WF, Guan ZZ, Bogdanovic N and Nordberg
A: High selective expression of alpha7 nicotinic receptors on
astrocytes in the brains of patients with sporadic Alzheimer's
disease and patients carrying Swedish APP 670/671 mutation: A
possible association with neuritic plaques. Exp Neurol.
192:215–225. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu W, Mechawar N, Krantic S, Chabot JG and
Quirion R: Upregulation of astrocytic α7 nicotinic receptors in
Alzheimer's disease brain-possible relevant to amyloid pathology.
Mol Neurodegener. 7 (Suppl 1):O72012. View Article : Google Scholar
|
|
16
|
Shammas SL, Waudby CA, Wang S, Buell AK,
Knowles TP, Ecroyd H, Welland ME, Carver JA, Dobson CM and Meehan
S: Binding of the molecular chaperone αB-crystallin to Aβ amyloid
fibrils inhibits fibril elongation. Biophys J. 101:1681–1689. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wilhelmus MM, Boelens WC, Otte-Höller I,
Kamps B, de Waal RM and Verbeek MM: Small heat shock proteins
inhibit amyloid-beta protein aggregation and cerebrovascular
amyloid-beta protein toxicity. Brain Res. 1089:67–78. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Raman B, Ban T, Sakai M, Pasta SY,
Ramakrishna T, Naiki H, Goto Y and Rao ChM: AlphaB-crystallin, a
small heat-shock protein, prevents the amyloid fibril growth of an
amyloid beta-peptide and beta2-microglobulin. Biochem J.
392:573–581. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Volovik Y, Moll L, Marques FC, Maman M,
Bejerano-Sagie M and Cohen E: Differential regulation of the heat
shock factor 1 and DAF-16 by neuronal nhl-1 in the nematode C.
elegans. Cell Rep. 9:2192–2205. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
McCarthy KD and de Vellis J: Preparation
of separate astroglial and oligodendroglial cell cultures from rat
cerebral tissue. J Cell Biol. 85:890–902. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Klein WL: Abeta toxicity in Alzheimer's
disease: Globular oligomers (ADDLs) as new vaccine and drug
targets. Neurochem Int. 41:345–352. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rönicke R, Mikhaylova M, Rönicke S,
Meinhardt J, Schröder UH, Fändrich M, Reiser G, Kreutz MR and
Reymann KG: Early neuronal dysfunction by amyloid β oligomers
depends on activation of NR2B-containing NMDA receptors. Neurobiol
Aging. 32:2219–2228. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kihara T, Shimohama S, Urushitani M,
Sawada H, Kimura J, Kume T, Maeda T and Akaike A: Stimulation of
alpha4beta2 nicotinic acetylcholine receptors inhibits beta-amyloid
toxicity. Brain Res. 792:331–334. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Steiner RC, Heath CJ and Picciotto MR:
Nicotine-induced phosphorylation of ERK in mouse primary cortical
neurons: Evidence for involvement of glutamatergic signaling and
CaMKII. J Neurochem. 103:666–678. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Valle-Casuso JC, Gonzalez-Sanchez A,
Medina JM and Tabernero A: HIF-1 and c-Src mediate increased
glucose uptake induced by endothelin-1 and connexin43 in
astrocytes. PLoS One. 7:e324482012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Herrero-González S, Valle-Casuso JC,
Sánchez-Alvarez R, Giaume C, Medina JM and Tabernero A: Connexin43
is involved in the effect of endothelin-1 on astrocyte
proliferation and glucose uptake. Glia. 57:222–233. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang T, Xiao T, Sun Q and Wang K: The
current agonists and positive allosteric modulators of α7 nAChR for
CNS indications in clinical trials. Acta Pharm Sin B. 7:611–622.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kalkman HO and Feuerbach D: Modulatory
effects of α7 nAChRs on the immune system and its relevance for CNS
disorders. Cell Mol Life Sci. 73:2511–2530. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen T, Wang Y, Zhang T, Zhang B and Chen
L, Zhao L and Chen L: Simvastatin Enhances activity and trafficking
of α7 nicotinic acetylcholine receptor in hippocampal neurons
through PKC and CaMKII signaling pathways. Front Pharmacol.
9:3622018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mannelli LDC, Tenci B, Zanardelli M,
Failli P and Ghelardini C: α7 nicotinic receptor promotes the
neuroprotective functions of astrocytes against oxaliplatin
neurotoxicity. Neural Plasticity. 2015:1–10. 2015. View Article : Google Scholar
|
|
31
|
Hardy JA and Higgins GA: Alzheimer's
disease: The amyloid cascade hypothesis. Science. 256:184–185.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gendron R, Plamondon P and Grenier D:
Binding of pro-matrix metalloproteinase 9 by Fusobacterium
nucleatum subsp. nucleatum as a mechanism to promote the invasion
of a reconstituted basement membrane. Infect Immun. 72:6160–6163.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gustavo D, Glogowski CM, Eliezer M and
Heinemann SF: Deletion of the α7 nicotinic acetylcholine receptor
gene improves cognitive deficits and synaptic pathology in a mouse
model of Alzheimer's disease. J Neurosci. 29:8805–8815. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Maiti P, Manna J, Veleri S and Frautschy
S: Molecular chaperone dysfunction in neurodegenerative diseases
and effects of curcumin. Biomed Res Int. 2014:4950912014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qi Y, Dou DQ, Jiang H, Zhang BB, Qin WY,
Kang K, Zhang N and Jia D: Arctigenin attenuates learning and
memory deficits through PI3k/Akt/GSK-3β pathway reducing tau
hyperphosphorylation in Aβ-induced AD mice. Planta Med. 83:51–56.
2017.PubMed/NCBI
|