Introduction
Diabetes mellitus (DM) characterized by
hyperglycemia, is the most common metabolic disease worldwide and
has become a global threat to human health (1). Patients with DM suffer from
endothelial dysfunction and vascular damage due to a variety of
harmful stimuli, including hyperglycemia, hyperlipidemia, oxidative
stress and inflammation, which eventually lead to a series of
diabetic vascular complications, including peripheral artery
disease (PAD) (2). Severe
hypoperfusion of blood flow for an extended period in patients with
PAD leads to ischemic foot ulceration and the development of
gangrene, which is one of the primary causes of foot amputation
(3), and for a number of patients,
this stage may be the only outcome. Although controlling and
managing vascular risk factors can effectively improve the
treatment of DM, the incidence, disability and mortality rates of
diabetic peripheral vascular disease remain high (4). The mechanisms underlying angiogenesis
repair following ischemia and endothelial cell function impairment
during DM are not completely understood. At present, it has been
suggested that promoting ischemic tissue angiogenesis may serve as
a therapeutic strategy for PAD (5,6).
The primary cause of delayed wound healing in
patients with diabetes is impaired angiogenesis (7). Angiogenesis refers to the migration
and proliferation of vascular endothelial cells based on original
capillaries and/or venues and the formation of new capillaries from
pre-existing vessels in the form of budding or non-budding
(8). Angiogenesis serves an
important role in promoting the recovery of diabetic vascular
complications (9) and is regulated
by angiogenic factors, involving endothelial cell proliferation,
migration and tube-like structure formation (10). Previous studies have demonstrated
that angiogenesis can be regulated by microRNAs (miRNAs) and that
vascular-specific miRNAs are key regulators of angiogenesis
(11). miRNAs are endogenous
non-coding RNAs that are ~22 nucleotides in length. miRNAs regulate
endothelial cell migration, proliferation and tube-like structure
formation, as well as the expression of angiogenesis-related
factors, ultimately affecting angiogenesis (12). miRNAs are able to promote and
inhibit angiogenesis; miRNA-126, miRNA-378 and miRNA-132 promote
angiogenesis, whereas miRNA-195, miRNA-221 and miR-192 inhibit
angiogenesis (11). Therefore,
there is a theoretical basis for the use of miRNAs as a biological
therapy for treating diabetic vascular complications. High glucose
(HG) levels can upregulate the expression levels of miRNA-328 to
induce HUVEC epithelial-mesenchymal transition, and the mechanism
of action is related to the mitogen-activated protein kinase
kinase-1/2-ERK1/2 signaling pathway (13). Previous studies have indicated that
miR-328 antagonists can improve erectile dysfunction in a rat model
of diabetes by regulating the expression levels of cGMP and
advanced glycation end products (14), and miR-328 can modulate glucose
uptake and metabolism related to diabetic retinopathy (15,16).
However, the role and mechanism of action underlying miR-328 during
HUVEC angiogenesis under hyperglycemic and ischemic conditions is
not completely understood.
In the present study, to simulate hyperglycemia
combined with ischemia-induced tissue starvation, HUVECs were
cultured in endothelial cell medium (ECM) with 25 mmol/l D-glucose
and 2% FBS for 24 h (HG + 2% FBS group). Subsequently, cell
functions related to HUVEC angiogenesis were investigated,
including migration, cytotoxicity and tube-like structure
formation. The expression levels of miRNA-328 and vascular
endothelial growth factor (VEGF), a marker of angiogenesis, were
detected. Subsequently, the expression level of miR-328 was
regulated in the HUVEC model of diabetic vascular ischemia and
subsequently, endothelial cell functions and VEGF expression levels
were observed. The mechanism of action underlying miR-328-mediated
angiogenesis regulation in the HUVEC model of diabetic vascular
ischemia was explored.
Materials and methods
Cell culture and treatment
HUVECs (ScienCell Research Laboratories, Inc.) were
grown in complete ECM (SciencCell Research Laboratories, Inc.)
containing 5% FBS (SciencCell Research Laboratories, Inc.), 1% EC
growth factors (ECGS; SciencCell Research Laboratories, Inc.) and
1% penicillin and streptomycin at 37°C in an atmosphere containing
5% CO2. Cells at passage 3–5 in the logarithmic growth
phase were used for subsequent experiments. Type 2 DM combined with
arteriosclerosis obliterans is a complex disease, and in
vitro, several stimulating factors, such as HG, low growth
factors, advanced glycation end products, oxidized LDL and serum
starvation, combined with hypoxia have been used to induce cell
models (17–19). Therefore, HUVECs were cultured for
24 h at 37°C with ECM containing 25 mmol/l D-glucose to mimic HG,
and 2% FBS and low growth factor levels to mimic ischemia-induced
tissue starvation (HG + 2% FBS group). HUVECs were exposed to
Perifosine (1 µM; cat. no. HY-50909; MedChemExpress) for 24 h at
37°C in an atmosphere containing 5% CO2, then
downregulation of miR-328 was performed and the changes in protein
and cell function were measured.
Wound healing assay
HUVECs were seeded (5×105 cells/well)
into 6-well plates. At 90% confluence, a single scratch was made in
the cell monolayer using a sterile 200 µl pipette tip.
Subsequently, the cells were cultured for 24 h at 37°C in an
atmosphere containing 5% CO2. The control group was
cultured in ECM containing 2% FBS, whereas the HG + 2% FBS group
was cultured in ECM containing HG and 2% FBS. Cells were observed
in six random fields of view at 0, 6 and 24 h using an inverted
light microscope (magnification, ×40). ImageJ version 1.49 software
(National Institutes of Health) was used for image analysis and
quantification.
Cell Counting Kit-8 (CCK-8)
assays
HUVEC cytotoxicity was determined using the CCK-8
assay, according to the manufacturer's instructions. Cells were
seeded (1×103 cells/well) into 96-well flat-bottomed
plates with 5 replicates in each group. Following treatment with 25
mmol/l D-glucose and 2% FBS for 24 h, or transfection with miR-328
inhibitor (IN328) and treatment with 25 mmol/l D-glucose and 2% FBS
for 24 h, the cells were washed twice with PBS. Subsequently, CCK-8
reagent (10 µl; Beyotime Institute of Biotechnology) was added to
each well and incubated at 37°C for 2 h in the dark. The optical
density (OD) values of each well were measured at a wavelength of
450 nm using a microplate reader (Bio-Rad Laboratories, Inc.).
Tube formation assay
To assess tube formation, pre-cooled 48-well plates
were coated with Matrigel® (30 µl) at 37°C for 30 min.
HUVECs were seeded (1.0×105 cells/ml) onto the
Matrigel®-coated plates at 37°C in an atmosphere
containing 5% CO2 for 16 h. Tube-like structure
formation was observed using an inverted light microscope
(magnification, ×100; Olympus Corporation).
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted using TRIzol®
(Invitrogen; Thermo Fisher Scientific, Inc.). Total RNA (500 ng)
was reverse transcribed to cDNA using a miRcute Plus miRNA
First-Strand cDNA kit (Tiangen Biotech Co., Ltd.), according to the
manufacturer's instructions. Subsequently, qPCR was performed using
the miRcute Plus miRNA qPCR Detection kit (Tiangen Biotech Co.,
Ltd.) and a StepOne Plus PCR system (Applied Biosystems; Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
The following thermocycling conditions were used for the qPCR:
Initial denaturation at 95°C for 15 min; followed by 40 cycles of
denaturation at 94°C for 15 sec, annealing at 55°C for 30 sec and
extension at 70°C for 30 sec. The primers for hsa-miR-328 and U6
were purchased from Guangzhou RiboBio Co., Ltd. miRNA expression
levels were quantified using the 2−ΔΔCq method and
normalized to the internal reference gene U6 (20). RT-qPCR was performed in
triplicate.
Transfections
IN328 (100 nmol/l; Guangzhou RiboBio Co., Ltd.) or
its negative control (NC; 100 nmol/l; Guangzhou RiboBio Co., Ltd.)
were used in the present study. Following transfection at 37°C in
an atmosphere containing 5% CO2 for 12 h, the culture
medium was replaced with fresh medium. After 24 h, HUVECs were
exposed to HG and 2% FBS at 37°C in an atmosphere containing 5%
CO2 and subsequently, wound healing, CCK-8, tube
formation and western blot assays were performed.
Western blotting
Cells were lysed using RIPA buffer (Beyotime
Institute of Biotechnology). Total protein was quantified using a
bicinchoninic acid assay kit (Thermo Fisher Scientific, Inc.).
Subsequently, proteins (30 µg) were separated via 10% SDS-PAGE and
transferred onto PVDF membranes. Following blocking with non-fat
milk for 2 h at room temperature (Beyotime Institute of
Biotechnology), the membranes were incubated at 4°C overnight with
the following primary antibodies: Anti-VEGF (1:1,000; cat. no.
AV202; Beyotime Institute of Biotechnology), anti-phosphorylated
(p)-AKT (Ser-473; 1:1,000; cat. no. ab81823; Abcam), anti-AKT
(1:1,000; cat. no. ab18785; Abcam), anti-p-ERK (1:1,000; cat. no.
ab47339; Abcam), anti-ERK (1:1,000; cat. no. ab17942; Abcam),
anti-p-JNK (1:1,000; cat. no. ab124956; Abcam), anti-JNK (1:1,000;
cat. no. ab208035; Abcam), anti-mTOR (1:1,000, cat. no. AM832;
Beyotime Institute of Biotechnology), anti-p-mTOR (1:1,000, cat.
no. 5536; Cell Signaling Technologies), anti-PIM1 (1:1,000; cat.
no. 54523; Cell Signaling Technologies), anti-GAPDH (1:1,000; cat.
no.97166; Cell Signaling Technologies), anti-actin (1:1,000; cat.
no. 3700; Cell Signaling Technologies) and anti-β-tubulin (1:1,000;
cat. no.2146; Cell Signaling Technologies). Following primary
incubation, the membranes were incubated with horseradish
peroxidase-conjugated secondary antibodies for 1 h at room
temperature: Anti rabbit IgG (1:2,000; cat. no. A0208) and anti
mouse IgG (1:2,000; cat. no. A0216) were purchased from Beyotime
Institute of Biotechnology. Protein bands were visualized by
enhanced chemiluminescence detection reagent (Bio Rad Laboratories,
Inc.), and blots were semi quantified by densitometric analysis
using Quantity One v4.6.2 software (Bio Rad Laboratories, Inc.).
GAPDH, actin and β-tubulin were used as the loading controls.
Predict target gene
Firstly, the target gene of miR-328 was predicted in
the miRWalk2.0 database (http://zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk/),
and the intersection of target genes in TargetScanHuman 7.1
(http://www.targetscan.org/vert_71/),
miRDB (http://mirdb.org/) and miRWalk2.0 database was
selected as the predicted target gene. Then, the target genes and
signal pathways related to angiogenesis were found in the Database
for Annotation, Visualization, and Integrated Discovery (DAVID;
http://david.ncifcrf.gov/). Then, the
target genes for promoting angiogenesis were found by reference
(21).
Statistical analysis
All experiments were performed in at least
triplicate. Data are presented as the mean ± standard deviation
(SD). One-way ANOVA followed by the Student-Newman-Keuls post hoc
test was used to analyze differences among multiple groups. An
unpaired Student's t-test was used to analyze differences between
two groups. Statistical analyses were performed using SPSS software
(v19.0; IBM Corp.). P<0.05 was considered to indicate a
statistically significant difference.
Results
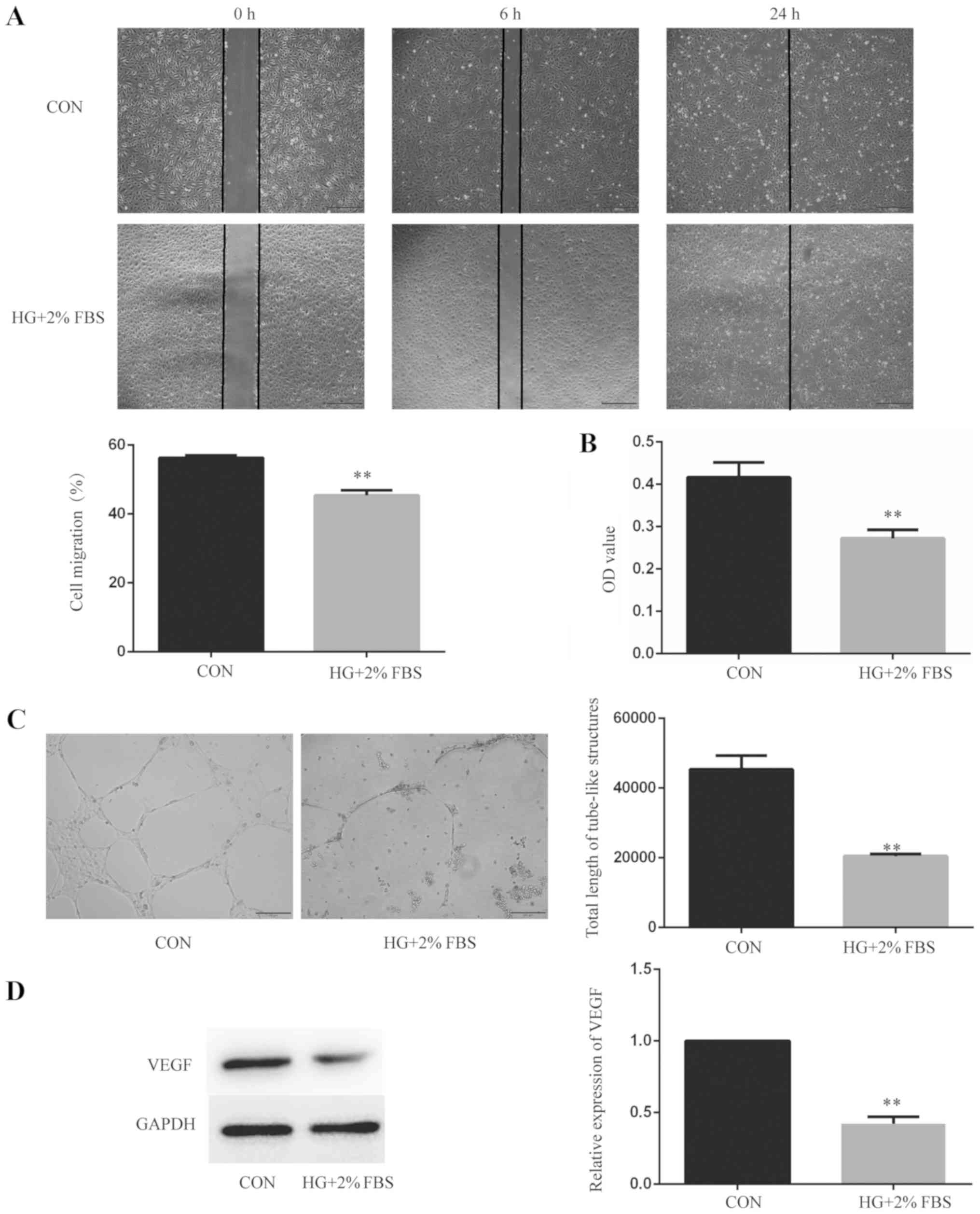
HUVEC angiogenesis is inhibited under
HG and low serum conditions
Alterations to HUVEC angiogenesis, cell function and
VEGF protein expression levels were observed following culture with
HG and low serum for 24 h. In terms of cell function, cell
migration, cytotoxicity and tube-like structure formation were
evaluated. Cells were divided into two groups: i) Control group;
and ii) HG + 2% FBS group. Wound healing assays were performed to
measure HUVEC migration (Fig. 1A).
Compared with the control group, the 6 h wound healing rate of the
HG + 2% FBS group was decreased (P<0.01), which suggested that
HG and low serum conditions inhibited the migration of endothelial
cells. The CCK-8 assay was used to measure HUVEC cytotoxicity
(Fig. 1B). Compared with the
control group, the OD value of the HG + 2% FBS group was decreased
(P<0.01), which indicated that HG and low serum conditions
increased the cytotoxicity of endothelial cells. Tube formation
assays were performed to detect the tube-like structure formation
of HUVECs (Fig. 1C). Compared with
the control group, the total length of the tube-like structures was
decreased in the HG + 2% FBS group (P<0.01), which indicated
that HG and low serum conditions inhibited the tube-like structure
formation of endothelial cells. Furthermore, western blotting was
performed to detect VEGF protein expression levels (Fig. 1D). Compared with the control group,
the relative expression levels of VEGF in the HG + 2% FBS group
were decreased (P<0.01), which indicated that HG and low serum
conditions inhibited the angiogenesis of endothelial cells at the
protein level. Collectively, the results suggested that endothelial
cell angiogenesis was inhibited under HG and low serum
conditions.
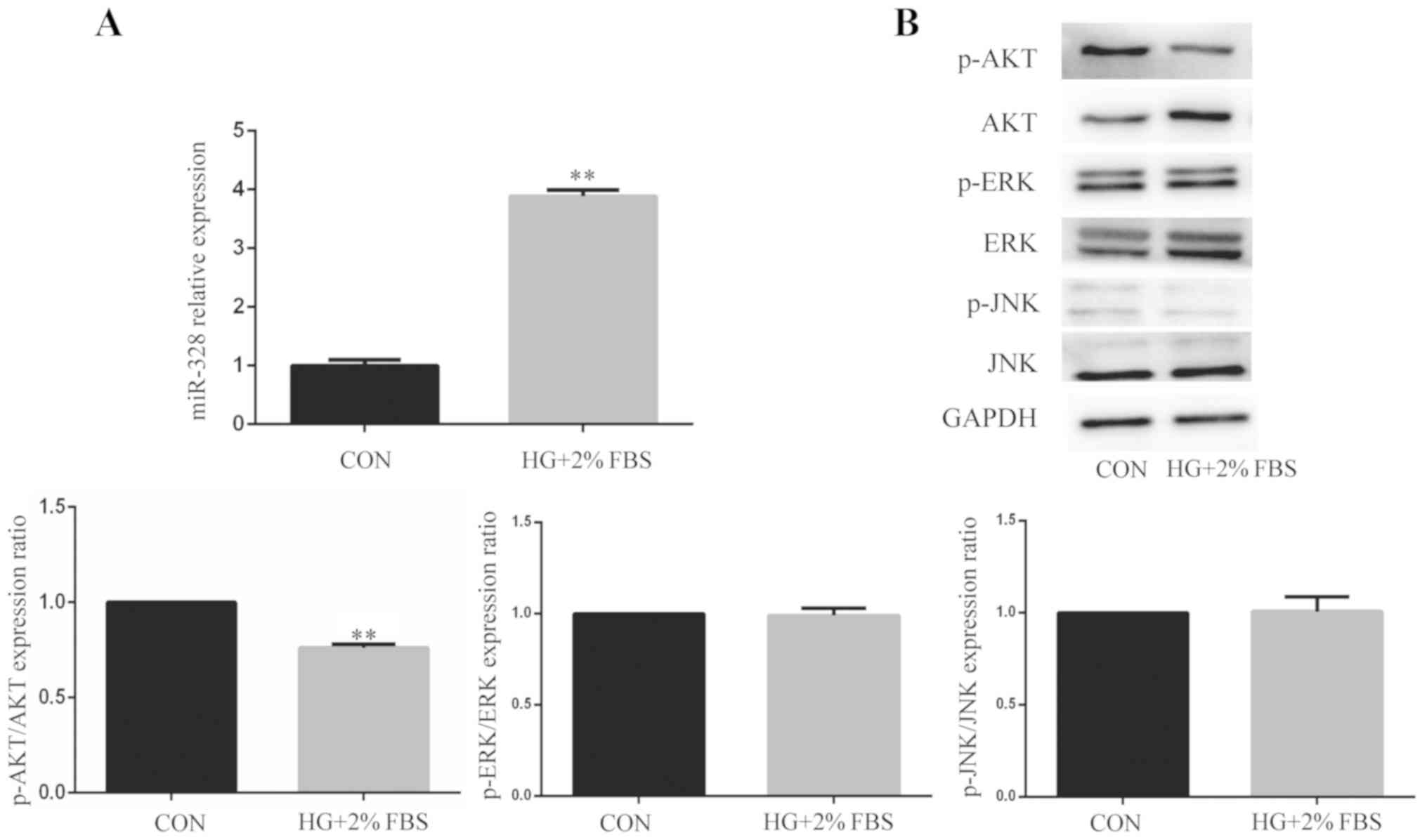
miR-328 expression is increased and
the AKT signaling pathway is suppressed in HUVECs under HG and low
serum conditions
To clarify the effect of HG and low serum
conditions, miR-328 expression levels and the ratios of p-AKT/AKT,
p-ERK/ERK and p-JNK/JNK were determined in HUVECs following
treatment with HG and low serum for 24 h. miR-328 expression levels
were significantly increased in the HG + 2% FBS group compared with
the control group (Fig. 2A).
Moreover, the ratio of p-AKT/AKT was significantly decreased in the
HG + 2% FBS group compared with the control group. However, the
ratios of p-ERK/ERK and p-JNK/JNK were not significantly different
between the control group and the HG + 2% FBS group (Fig. 2B). The results indicated that the
AKT signaling pathway was associated with miR-328-mediated
regulation of cell function, but the ERK and JNK signaling pathways
are not associated with miR-328-mediated effects. Based on the
finding that only the AKT signaling pathway was altered in HUVECs
following treatment with HG and 2% FBS, only the AKT signaling
pathway was investigated in subsequent experiments. The results
suggested that the inhibition of endothelial cell angiogenesis by
HG and low serum conditions may be related to miR-328 and the AKT
signaling pathway.
Downregulation of miR-328 promotes
HUVEC angiogenesis under HG and low serum conditions
To investigate the angiogenesis function of miR-328
under HG and low serum conditions, HUVECs were divided into the NC
and IN328 groups. Transfection efficiency of IN328 was detected
using RT-qPCR. Compared with the NC group, the expression of
miR-328 in the IN328 group was significantly lower, which indicated
that the transfection was successful (Fig. S1). Subsequently, angiogenesis and
VEGF protein expression levels were investigated. The wound healing
assay was performed to detect the migratory ability of HUVECs.
Compared with the NC group, the 6 h wound healing rate of the IN328
group was increased (P<0.01), which indicated that
downregulation of miR-328 promoted endothelial cell migration
(Fig. 3A). Compared with the NC
group, the OD value of the IN328 group was increased (P<0.01),
which demonstrated that downregulation of miR-328 reduced cell
cytotoxicity (Fig. 3B). Tube
formation assays were performed to detect tube-like structure
formation in HUVECs. Compared with the NC group, the total length
of tube-like structures in the IN328 group was increased
(P<0.01), which suggested that downregulation of miR-328
promoted tube-like structure formation in endothelial cells
(Fig. 3C). Additionally, western
blotting was performed to detect VEGF protein expression levels in
HUVECs. Compared with the NC group, the relative expression levels
of VEGF in the IN328 group were increased (P<0.01), which
indicated that downregulation of miR-328 promoted endothelial cell
angiogenesis at the protein level (Fig. 3D). The results suggested that
endothelial cell angiogenesis was enhanced by downregulation of
miR-328 under HG and low serum conditions.
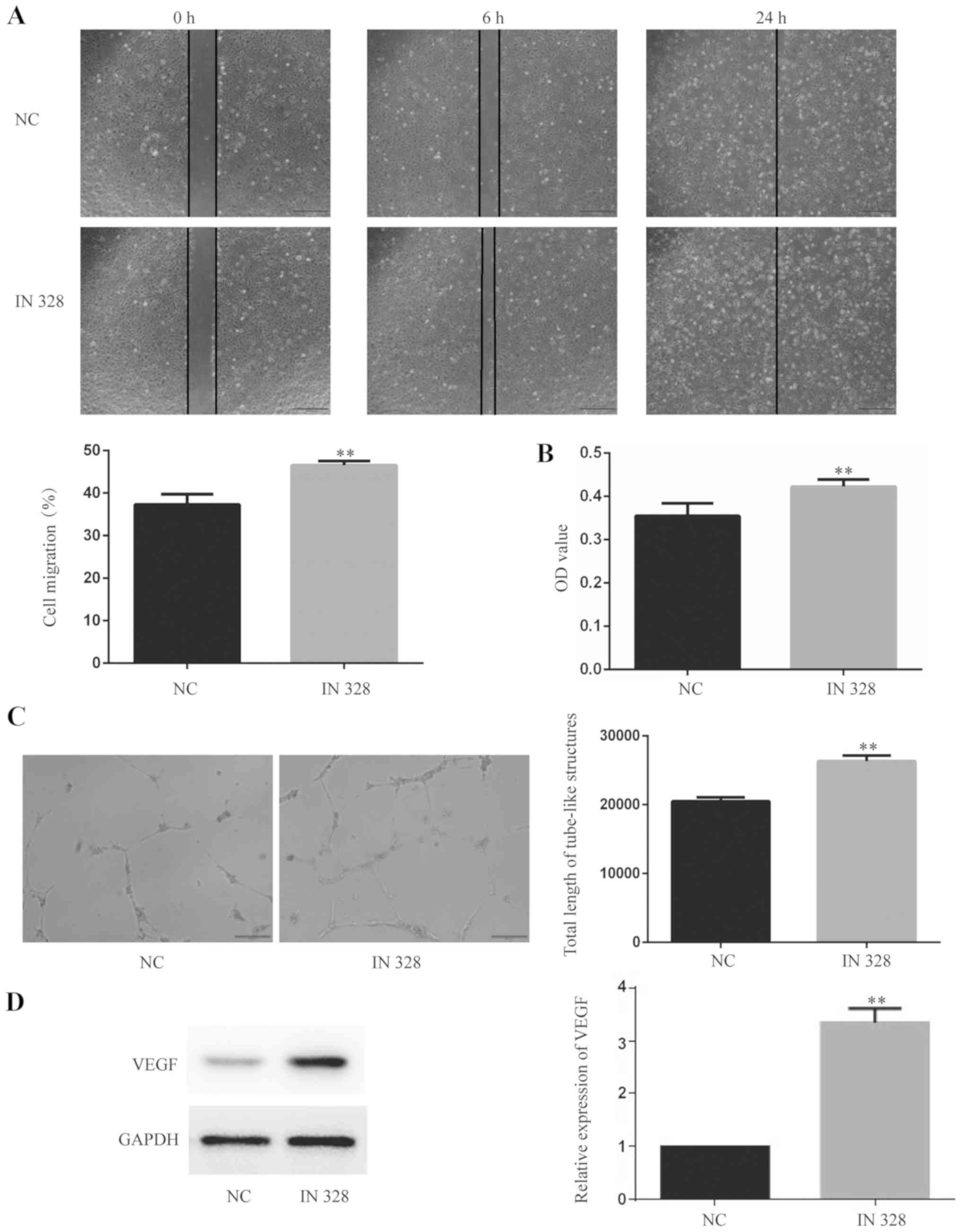 | Figure 3.Downregulation of miR-328 promotes
HUVECs angiogenesis under HG and low serum condition. (A) HUVEC
migration was determined using wound healing assays (magnification,
×40). The data presented in the graph are from the 6 h time point.
Scale bar, 500 µm. (B) HUVEC cytotoxicity was determined using the
Cell Counting Kit-8 assay. (C) The tube-like structure formation of
HUVECs was detected using tube formation assays (magnification,
×100). Scale bar, 200 µm. (D) VEGF protein expression levels in
HUVECs were determined by western blotting. **P<0.01 vs. CON.
miR, microRNA; HG, high glucose; VEGF, vascular endothelial growth
factor; CON, control; IN328, miR-328 inhibitor; OD, optical
density. |
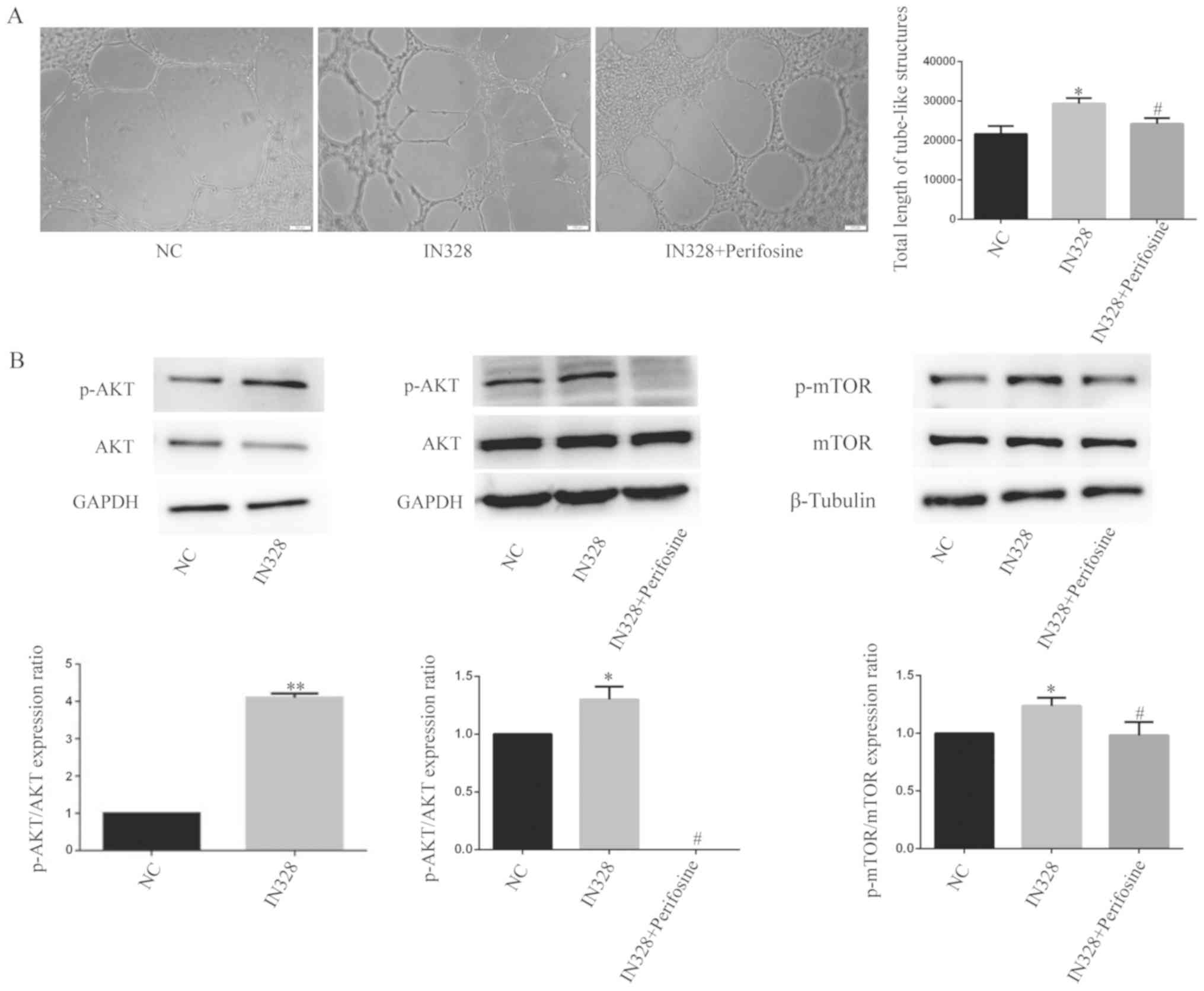
Downregulation of miR-328 activates
the AKT/mTOR signaling pathway in HUVECs under HG and low serum
conditions
The aforementioned results indicated that HG and low
serum conditions inhibited activation of the AKT signaling pathway
in HUVECs. Moreover, it has been reported that the AKT/mTOR
signaling pathway can regulate angiogenesis; therefore, alterations
in the AKT/mTOR signaling pathway were investigated following
downregulation of miR-328. Following miR-328 downregulation in
HUVECs under HG and low serum conditions, inhibiting AKT
phosphorylation with perifosine partially reversed IN328-mediated
tube-like structure formation, which indicated that perifosine at
least partially blocked IN328-mediated stimulation of HUVEC
angiogenesis (Fig. 4A). Following
downregulation of miR-328 in HUVECs under HG and low serum
conditions, the ratios of p-AKT/AKT and p-mTOR/mTOR were
significantly increased compared with the NC group, indicating that
the AKT/mTOR signaling pathway was activated (Fig. 4B). By inhibiting AKT
phosphorylation with perifosine in the IN328 group, the ratios of
p-AKT/AKT and p-mTOR/mTOR were decreased, which indicated that
perifosine blocked the AKT/mTOR signaling pathway (Fig. 4B). The results demonstrated that
downregulation of miR-328 promoted HUVECs angiogenesis by
activating the AKT/mTOR signaling pathway under HG and low serum
conditions.
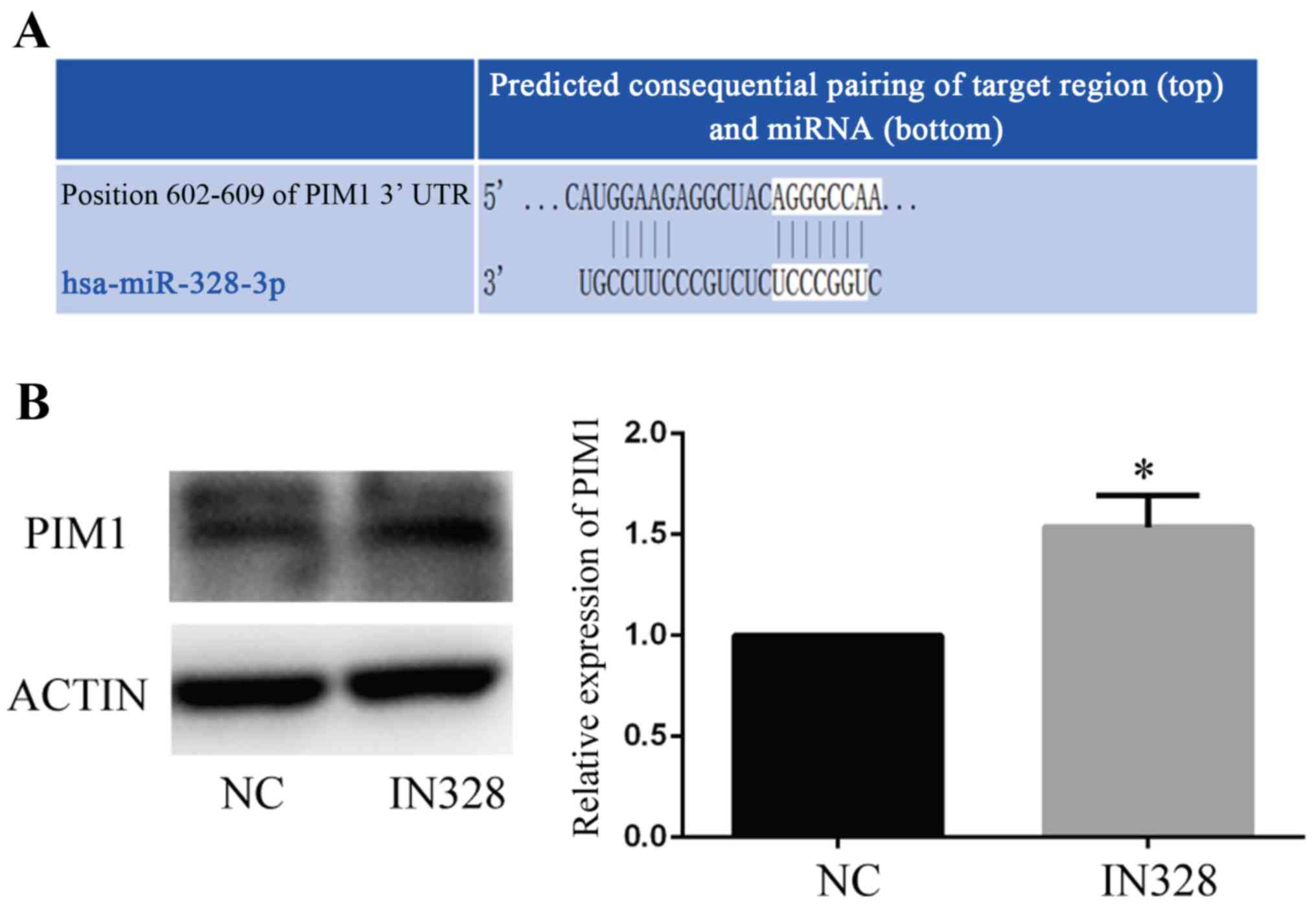
miR-328 may serve a regulatory role
via PIM1
Based on the sequence of miR-328, possible binding
targets of miR-328 were predicted by miRWalk2.0 and DAVID database.
Subsequently, proangiogenesis-associated predicted targets were
selected. PIM1, a gene that is associated with cell survival, was
one of the predicted targets (Fig.
5A). To verify the prediction, IN328 and a NC were transfected
into HUVECs under HG and low serum conditions. The western blotting
results indicated that IN328 significantly increased PIM1 protein
expression levels compared with the NC group (Fig. 5B). The results indicated that
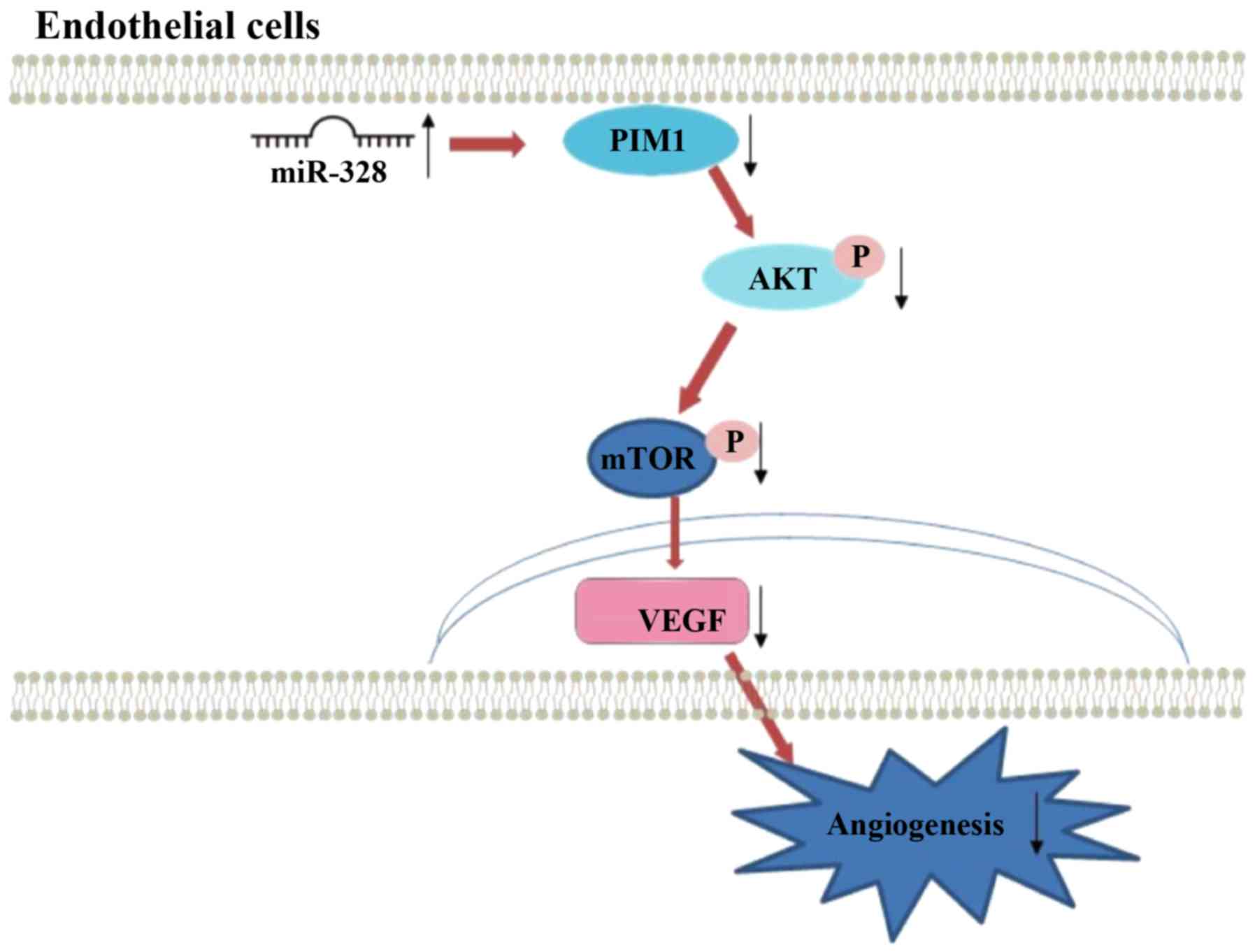
miR-328 regulated endothelial cell angiogenesis via PIM1 (Fig. 6).
Discussion
Increasing evidence suggests that endothelial cell
dysfunction contributes to the onset and progression of
diabetes-related vascular complications (22); therefore, the role of miRNAs as
regulatory factors during such processes have gained increasing
attention. Although the effects of miR-328 have been studied
extensively, the mechanism underlying miR-328 during HUVEC
angiogenesis is not completely understood. In the present study,
whether miR-328 expression levels were altered in response to HG
and low serum conditions, as well as whether the alterations
contributed to HUVEC angiogenesis damage under HG and low serum
conditions were investigated. To the best of our knowledge, the
present study was the first to investigate the role of miR-328 in
HUVECs under HG and low serum conditions. The aim of the present
study was to preliminary explore the role of miR-328 in
diabetes-related vascular complications; therefore, high glucose
combined with low serum conditions were used to establish a cell
model that directly simulated the state of vascular endothelial
cells during diabetic limb ischemia (17). The results indicated that
endothelial cell angiogenesis was inhibited under HG and low serum
condition, which was consistent with the results reported by
Caporali et al (1). It was
also speculated whether either condition was sufficient to give
rise to the effects observed in the present study. High-glucose can
significantly decreased cell features such as angiogenic capability
(23), however, further
investigation is required. In the present study, miR-328 expression
levels were significantly upregulated in HUVECs under HG and low
serum conditions compared with control HUVECs, which indicated that
downregulation of miR-328 promoted HUVEC angiogenesis under HG and
low serum conditions. Further experiments indicated that miR-328
mediated endothelial cell angiogenesis, at least in part, by
regulating PIM1 and the AKT/mTOR signaling pathway.
Angiogenesis is primarily associated with
endothelial cell migration, proliferation and tube-like structure
formation, which can be regulated by miRNAs (24). In the wound healing assay,
serum-free conditions alter cell proliferation and migration;
therefore, to observe the effects of HG + 2% FBS on cell migration,
the control group was cultured with 2% FBS; the use of 2% FBS
during the wound healing assay was a limitation of the study. It
has been reported that miR-328 is related to DM and can regulate
the proliferation and migration of tumor cells (14,25).
The majority of studies on miR-328 have focused on tumors and
cardiovascular diseases. It has been reported that the expression
of miR-328 is decreased in esophageal, liver and colon cancer,
where it can inhibit the proliferation, migration and survival of
cancer cells (26–28). In cardiovascular diseases, miR-328
is associated with endothelial cell mesenchymal transformation,
atrial fibrillation and myocardial fibrosis (13,29–31);
therefore, it was hypothesized that miR-328 may regulate
endothelial cell function and thus, affect angiogenesis (13,26–28).
Firstly, the effect of HG and low serum conditions on endothelial
cell function was investigated, which indicated that the conditions
inhibited cell migration and tube-like structure formation, and
promoted cytotoxicity. Subsequently, similar to its function in
tumor cells and pulmonary arterial smooth muscle cells (25), the results of the present study
further indicated that miR-328 expression levels were upregulated
under HG and low serum conditions compared with the control group.
Among the endothelial cell survival-related signaling pathways,
ERK, JNK and AKT were investigated, and the results suggested that
only AKT protein expression levels were decreased under HG and low
serum conditions compared with the control group, which suggested
that AKT signaling was involved.
Subsequently, the specific mechanism of action
underlying miR-328-mediated regulation of endothelial cells and the
signaling pathways involved were investigated. Following
downregulation of miR-328, HG and low serum condition-mediated
inhibitory effects on cell survival, including migration and
tube-like structure formation, were reversed, which indicated that
miR-328 regulated cell angiogenesis. In addition, downregulation of
miR-328 promoted AKT activation, which suggested that miR-328
mediated AKT-regulated angiogenesis in DM-associated endothelial
dysfunction. Furthermore, inhibition of AKT using the AKT specific
inhibitor periosine partially reduced IN328-mediated stimulation of
angiogenesis.
AKT, a protein kinase B, is a
serine/threonine-specific protein kinase, which serves a key role
in cell proliferation-related processes, such as glucose
metabolism, cell cycle regulation, angiogenesis and cell invasion
(32). Previous studies have
demonstrated that AKT is an effective angiogenesis factor (33–35).
Hu et al (36) and Fathi
et al (37), reported that
PIM1 inhibition inhibited AKT phosphorylation, indicating that AKT
was downstream of PIM1. To further study the downstream effects of
the AKT signaling pathway, the angiogenesis-related mTOR signaling
pathway was screened for validation. The results further indicated
that mTOR was a downstream signal of AKT. Similar to the results
reported by Chen et al (38), it has also been hypothesized that
the AKT/mTOR signaling pathway regulates cell proliferation and
angiogenesis by inducing the expression of hypoxia-inducible factor
1α and VEGF (39). Therefore,
miR-328-mediated AKT/mTOR signaling may be associated with HUVEC
angiogenesis.
miRNAs can guide ribosome binding to the
3′-untranslated region of target genes and post-transcriptionally
inhibit target gene translation (40); therefore, the function of miR-328
depends on the target molecule. Bioinformatics analysis predicted
that PIM1 may be a target gene of miR-328. The results indicated
that PIM1 protein expression levels were regulated by miR-328 and
that PIM1 may be a target of miR-328. However, further
investigation is required to confirm that PIM1 is a target gene of
miR-328. PIM1 is a serine/threonine kinase that has diverse
biological roles in cell survival, proliferation and
differentiation (41). The PIM
family of proteins, a family of proto-oncogenes, display increased
expression levels in a variety of malignancies. PIM kinases enhance
cell survival and suppress apoptosis in multiple types of tumor
cells (36,37,42).
Previous studies have also indicated that PIM1 can promote
endothelial migration and angiogenesis, and inhibit cytotoxicity
(21,43,44).
The results of the present study demonstrated that miR-328 altered
endothelial cell angiogenesis, at least in part, by interacting
with PIM1. Further studies are required to investigate the direct
or indirect relationship between miR-328 and PIM1. Moreover,
alterations to endothelial cell angiogenesis, including cell
migration, cytotoxicity and tube formation, and the AKT signaling
pathway following PIM1 gene regulation need to be investigated in
future studies. Based on the results of the present study and the
aforementioned previous studies, it could be suggested that the
mechanism underlying endothelial cell angiogenesis inhibition under
HG and low serum conditions involves increasing the expression of
miR-328, which negatively regulates PIM1 and subsequently, inhibits
the AKT/mTOR signaling pathway to inhibit angiogenesis.
In conclusion, the present study demonstrated that
HG and low serum conditions inhibited endothelial cell angiogenesis
and increased the expression of miR-328 in HUVECs. Moreover,
downregulation of miR-328 in HUVECs under HG and low serum
condition promoted endothelial cell angiogenesis. The mechanism of
action underlying miR-328-mediated regulation of endothelial cell
angiogenesis under HG and low serum conditions may be via the PIM1
and AKT/mTOR signaling pathway. The results of the present study
may provide novel therapeutic targets for diabetic vascular
complications.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Education
Department of Sichuan Province (grant nos. 16ZA0179 and 14TD0018)
and the Joint Project of Luzhou City and Southwest Medical
University (grant no. 2017LZXNYD-J18).
Availability of data and materials
All datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ and XZ conceived and designed the study. YZ, FW,
QL and XD performed the cell culture, RT-qPCR and cell function
experiments, and interpreted the data. RH and XH performed the
western blotting experiments and interpreted the data. YZ wrote the
manuscript. All authors read and approved the final manuscript and
agree to be accountable for all aspects of the research in ensuring
that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Caporali A, Meloni M, Nailor A, Mitić T,
Shantikumar S, Riu F, Sala-Newby GB, Rose L, Besnier M, Katare R,
et al: p75(NTR)-dependent activation of NF-κB regulates
microRNA-503 transcription and pericyte-endothelial crosstalk in
diabetes after limb ischaemia. Nat Commun. 6:80242015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lunder M, Janić M and Šabovič M:
Prevention of vascular complications in diabetes mellitus patients:
Focus on the arterial wall. Curr Vasc Pharmacol. 17:6–15. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Barshes NR and Grant CL: Advances in the
management of peripheral artery disease. Curr Diab Rep. 19:362019.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dal Canto E, Ceriello A, Rydén L, Ferrini
M, Hansen TB, Schnell O, Standl E and Beulens JW: Diabetes as a
cardiovascular risk factor: An overview of global trends of macro
and micro vascular complications. Eur J Prev Cardiol. 26
(2_suppl):25–32. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
He S, Zhao T, Guo H, Meng Y, Qin G,
Goukassian DA, Han J, Gao X and Zhu Y: Coordinated activation of
VEGF/VEGFR-2 and PPARδ pathways by a multi-component Chinese
medicine DHI accelerated recovery from peripheral arterial disease
in type 2 diabetic mice. PLoS One. 11:e01673052016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nazer B, Ghahghaie F, Kashima R, Khokhlova
T, Perez C, Crum L, Matula T and Hata A: Therapeutic ultrasound
promotes reperfusion and angiogenesis in a rat model of peripheral
arterial disease. Circ J. 79:2043–2049. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhu Y, Wang Y, Jia Y, Xu J and Chai Y:
Roxadustat promotes angiogenesis through HIF-1α/VEGF/VEGFR2
signaling and accelerates cutaneous wound healing in diabetic rats.
Wound Repair Regen. 27:324–334. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lazarovici P, Lahiani A, Gincberg G, Haham
D, Fluksman A, Benny O, Marcinkiewicz C and Lelkes PI: Nerve growth
factor-induced angiogenesis: 1. Endothelial cell tube formation
assay. Methods Mol Biol. 1727:239–250. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Can L, Junxiong Z, Bao H, Wen Z, Hong W,
Huijie L, Yingsheng X and Chunli S: Single intraosseous injection
of simvastatin promotes endothelial progenitor cell mobilization,
neovascularization, and wound healing in diabetic rats. Plast
Reconstr Surg. 145:433–443. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Odent Grigorescu G, Rosca AM, Preda MB,
Tutuianu R, Simionescu M and Burlacu A: Synergic effects of VEGF-A
and SDF-1 on the angiogenic properties of endothelial progenitor
cells. J Tissue Eng Regen Med. 11:3241–3252. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tiwari A, Mukherjee B and Dixit M:
MicroRNA key to angiogenesis regulation: MiRNA biology and therapy.
Curr Cancer Drug Targets. 18:266–277. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gong M, Yu B, Wang J, Wang Y, Liu M, Paul
C, Millard RW, Xiao DS, Ashraf M and Xu M: Mesenchymal stem cells
release exosomes that transfer miRNAs to endothelial cells and
promote angiogenesis. Oncotarget. 8:45200–45212. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen Y, Yang Q, Zhan Y, Ke J, Lv P and
Huang J: The role of miR-328 in high glucose-induced
endothelial-to-mesenchymal transition in human umbilical vein
endothelial cells. Life Sci. 207:110–116. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li DS, Feng L, Luo LH, Duan ZF, Li XL, Yin
CH and Sun X: The Effect of microRNA-328 antagomir on erectile
dysfunction in streptozotocin-induced diabetic rats. Biomed
Pharmacother. 92:888–895. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yi W, Tu MJ, Liu Z, Zhang C, Batra N, Yu
AX and Yu AM: Bioengineered miR-328-3p modulates GLUT1-mediated
glucose uptake and metabolism to exert synergistic
antiproliferative effects with chemotherapeutics. Acta Pharm Sin B.
10:159–170. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Prado MSG, de Goes TC, de Jesus ML,
Mendonça LSO, Nascimento JS and Kaneto CM: Identification of
miR-328-3p as an endogenous reference gene for the normalization of
miRNA expression data from patients with Diabetic Retinopathy. Sci
Rep. 9:196772019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Caporali A, Meloni M, Völlenkle C, Bonci
D, Sala-Newby GB, Addis R, Spinetti G, Losa S, Masson R, Baker AH,
et al: Deregulation of microRNA-503 contributes to diabetes
mellitus-induced impairment of endothelial function and reparative
angiogenesis after limb ischemia. Circulation. 123:282–291. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ying C, Sui-Xin L, Kang-Ling X, Wen-Liang
Z, Lei D, Yuan L, Fan Z and Chen Z: MicroRNA-492 reverses high
glucose-induced insulin resistance in HUVEC cells through targeting
resistin. Mol Cell Biochem. 391:117–125. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang M, Li W, Chang GQ, Ye CS, Ou JS, Li
XX, Liu Y, Cheang TY, Huang XL and Wang SM: MicroRNA-21 regulates
vascular smooth muscle cell function via targeting tropomyosin 1 in
arteriosclerosis obliterans of lower extremities. Arterioscler
Thromb Vasc Biol. 31:2044–2053. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−ΔΔC(T)) Method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen M, Yi B, Zhu N, Wei X, Zhang GX,
Huang S and Sun J: Pim1 kinase promotes angiogenesis through
phosphorylation of endothelial nitric oxide synthase at Ser-633.
Cardiovasc Res. 109:141–150. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Durak-Kozica M, Paszek E and Stępień EŁ:
Role of the Wnt signalling pathway in the development of
endothelial disorders in response to hyperglycaemia. Expert Rev Mol
Med. 21:e72019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schiano C, Grimaldi V, Franzese M, Fiorito
C, De Nigris F, Donatelli F, Soricelli A, Salvatore M and Napoli C:
Non-nutritional sweeteners effects on endothelial vascular
function. Toxicol In Vitro. 62:1046942020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang J, Yang L, Liang F, Chen Y and Yang
G: Integrin alpha × stimulates cancer angiogenesis through PI3K/Akt
signaling-mediated VEGFR2/VEGF-A overexpression in blood vessel
endothelial cells. J Cell Biochem. 120:1807–1818. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qian Z, Zhang L, Chen J, Li Y, Kang K, Qu
J, Wang Z, Zhai Y, Li L and Gou D: MiR-328 targeting PIM-1 inhibits
proliferation and migration of pulmonary arterial smooth muscle
cells in PDGFBB signaling pathway. Oncotarget. 7:54998–55011. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Han N, Zhao W, Zhang Z and Zheng P:
MiR-328 suppresses the survival of esophageal cancer cells by
targeting PLCE1. Biochem Biophys Res Commun. 470:175–180. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang X, Xin G and Sun D: Serum exosomal
miR-328, miR-575, miR-134 and miR-671-5p as potential biomarkers
for the diagnosis of Kawasaki disease and the prediction of
therapeutic outcomes of intravenous immunoglobulin therapy. Exp
Ther Med. 16:2420–2432. 2018.PubMed/NCBI
|
|
28
|
Li Y, Duo Y, Zhai P, He L, Zhong K, Zhang
Y, Huang K, Luo J, Zhang H and Yu X: Dual targeting delivery of
miR-328 by functionalized mesoporous silica nanoparticles for
colorectal cancer therapy. Nanomedicine (Lond). 13:1743–5889. 2018.
View Article : Google Scholar
|
|
29
|
Zhao D, Li C, Yan H, Li T, Qian M, Zheng
N, Jiang H, Liu L, Xu B, Wu Q, et al: Cardiomyocyte derived mir-328
promotes cardiac fibrosis by paracrinely regulating adjacent
fibroblasts. Cell Physiol Biochem. 46:1555–1565. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Soeki T, Matsuura T, Bando S, Tobiume T,
Uematsu E, Ise T, Kusunose K, Yamaguchi K, Yagi S, Fukuda D, et al:
Relationship between local production of microRNA-328 and atrial
substrate remodeling in atrial fibrillation. J Cardiol. 68:472–477.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
He F, Lv P, Zhao X, Wang X, Ma X, Meng W,
Meng X and Dong S: Predictive value of circulating miR-328 and
miR-134 for acute myocardial infarction. Mol Cell Biochem.
394:137–144. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yan X, Hui Y, Hua Y, Huang L, Wang L, Peng
F, Tang C, Liu D, Song J and Wang F: EG-VEGF silencing inhibits
cell proliferation and promotes cell apoptosis in pancreatic
carcinoma via PI3K/AKT/mTOR signaling pathway. Biomed Pharmacother.
109:762–769. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Di Y and Chen XL: Inhibition of LY294002
in retinal neovascularization via down-regulation the PI3K/AKT-VEGF
pathway in vivo and in vitro. Int J Ophthalmol. 11:1284–1289.
2018.PubMed/NCBI
|
|
34
|
Chen J, Li F, Xu Y, Zhang W, Hu Y, Fu Y,
Xu W, Ge S, Fan X and Lu L: Cholesterol modification of
SDF-1-specific siRNA enables therapeutic targeting of angiogenesis
through Akt pathway inhibition. Exp Eye Res. 184:64–71. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wu J, Zhao X, Sun Q, Jiang Y, Zhang W, Luo
J and Li Y: Synergic effect of PD-1 blockade and endostar on the
PI3K/AKT/mTOR-mediated autophagy and angiogenesis in Lewis lung
carcinoma mouse model. Biomed Pharmacother. 125:1097462020.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hu XF, Li J, Vandervalk S, Wang Z,
Magnuson NS and Xing PX: PIM-1-specific mAb suppresses human and
mouse tumor growth by decreasing PIM-1 levels, reducing Akt
phosphorylation, and activating apoptosis. J Clin Invest.
119:362–375. 2009.PubMed/NCBI
|
|
37
|
Fathi AT, Arowojolu O, Swinnen I, Sato T,
Rajkhowa T, Small D, Marmsater F, Robinson JE, Gross SD, Martinson
M, et al: A potential therapeutic target for FLT3-ITD AML: PIM1
kinase. Leuk Res. 36:224–231. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen QY, Jiao DM, Wu YQ, Chen J, Wang J,
Tang XL, Mou H, Hu HZ, Song J, Yan J, et al: MiR-206 inhibits
HGF-induced epithelial-mesenchymal transition and angiogenesis in
non-small cell lung cancer via c-Met /PI3k/Akt/mTOR pathway.
Oncotarget. 7:18247–18261. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang Y, Cheng H, Li W, Wu H and Yang Y:
Highly-expressed P2X7 receptor promotes growth and metastasis of
human HOS/MNNG osteosarcoma cells via PI3K/Akt/GSK3β/β-catenin and
mTOR/HIF1α/VEGF signaling. Int J Cancer. 145:1068–1082. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Baigude H and Rana TM: Strategies to
antagonize miRNA functions in vitro and in vivo. Nanomediwcine
(Lond). 9:2545–2555. 2014. View Article : Google Scholar
|
|
41
|
Zhang F, Beharry ZM, Harris TE, Lilly MB,
Smith CD, Mahajan S and Kraft AS: PIM1 protein kinase regulates
PRAS40 phosphorylation and mTOR activity in FDCP1 cells. Cancer
Biol Ther. 8:846–853. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Leung CO, Wong CC, Fan DN, Kai AK, Tung
EK, Xu IM, Ng IO and Lo RC: PIM1 regulates glycolysis and promotes
tumor progression in hepatocellular carcinoma. Oncotarget.
6:10880–10892. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Walpen T, Kalus I, Schwaller J, Peier MA,
Battegay EJ and Humar R: Nuclear PIM1 confers resistance to
rapamycin-impaired endothelial proliferation. Biochem Biophys Res
Commun. 429:24–30. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Walpen T, Peier M, Haas E, Kalus I,
Schwaller J, Battegay E and Humar R: Loss of pim1 imposes a
hyperadhesive phenotype on endothelial cells. Cell Physiol Biochem.
30:1083–1096. 2012. View Article : Google Scholar : PubMed/NCBI
|