Introduction
Autophagy is important for the removal of damaged,
degenerative, non-functional and aging proteins and organelles, as
well as for cellular homeostasis (1). Autophagy dysfunction is associated
with various types of disease, such as cardiovascular disease,
neurological disease and cancer (2). Autophagy is a continuous process
known as the autophagic flux, which includes three stages:
Autophagosome formation, autophagosome-lysosome fusion and
autolysosome degradation (3). The
association between autophagy and cancer is complex and remains
poorly understood. Autophagy has been found to lead to the
autophagic death of cancer cells, but it also can protect cancer
cells from nutritional deficiency, chemotherapy or radiotherapy
(4). A previous study also
reported that the inhibition of autophagy contributes to anticancer
therapy, in particular to the inhibition of autophagosome-lysosome
fusion or autolysosome degradation (5). Autophagic degradation inhibitors,
such as hydroxychloroquine, liensinine and andrographolide, have
also been found to enhance the anticancer activity of multiple
anticancer drugs (6–8). However, there are few studies on
whether the inhibition of autophagy degradation itself contributes
to cell death. Notably, a recent study has reported that classic
autophagy degradation inhibitors, such as ammonium chloride
chloroquine and bafilomycin, may cause autophagy-independent cell
death (9). Saikosaponin D (SSD), a
kind of triterpenoid saponin, is extracted from the Chinese
traditional herbal, Radix bupleuri (10). SSD has been found to inhibit a
variety of tumor cells, including ovarian cancer, lung cancer,
hepatoma and glioblastoma (11–14).
Previously, SSD was also discovered to induce apoptotic cell death
through the mitogen-activated protein kinase (MAPK) signaling
pathway (14). A previous study
also reported that SSD regulated autophagy; SSD induced autophagic
cell death through inhibiting sarco/endoplasmic reticulum
Ca2+ ATPase in apoptosis-defective cells (15). Additionally, SSD suppressed
enterovirus A71 infection through inhibiting autophagy (16).
The present study demonstrated that SSD induced
apoptosis by activating the p38 MAPK signaling pathway in
MDA-MB-231 cells and revealed that SSD potently blocked autophagic
degradation through inhibiting autophagosome-lysosome fusion.
Further research identified that blocking autophagy degradation was
not associated with SSD-mediated apoptosis. These findings provide
additional evidence for the association between inhibition of
autophagy degradation and cell death.
Materials and methods
Antibodies and reagents
Antibodies against phosphorylated (p)-mTOR (cat. no.
2971; dilution 1:500), m-TOR (cat. no. 2972; dilution 1:1,000),
serne/threonine protein kinase ULK1 (ULK1; cat. no. 8054S; dilution
1:1,000), p-ULK1 (cat. no. 6888; dilution 1:500), p-p38 (cat. no.
9216, dilution 1:500), p-ERK (cat. no. 4376; dilution 1:1,000), ERK
(cat. no. 9102; dilution 1:1,000), autophagy-related protein (ATG)5
(cat. no. 12994S; dilution 1:1,000), ATG7 (cat. no. 8558; dilution
1:1,000), p62 (cat. no. 5114; dilution 1:500), JNK (cat. no. 9252;
dilution 1:1,000), c-caspase-3 (cat. no. 9661S; dilution 1:200),
cathepsin D (cat. no. 2284; dilution 1:200), p-JNK (cat. no. 9251;
dilution 1:500) and p38 (cat. no. 9212; dilution 1:1,000) were
purchased from Cell Signaling Technology, Inc.; antibodies against
lysosome-associated membrane glycoprotein (LAMP)1 (cat. no.
sc-20011; dilution 1:1,000), LAMP2 (cat. no. sc-18822; dilution
1:1,000) and cathepsin B (cat. no. sc-13985; dilution 1:500) were
obtained from Santa Cruz Biotechnology, Inc.; anti-cleaved (c)-poly
(ADP-ribose) polymerase (c-PARP; cat. no. 380374; dilution 1:500)
was from Zen-Bio, Inc.; anti-GAPDH (cat. no. AG019; dilution
1:1,000) was purchased from Beyotime Institute of Biotechnology;
anti-microtubule-associated protein 1 light chain 3 beta (LC3B;
cat. no. L7543; dilution 1:1,000) and anti-Beclin 1 (cat. no.
B6186; dilution 1:500) were obtained from Sigma-Aldrich; Merck
KGaA; and horseradish peroxidase-conjugated secondary antibodies
(cat. nos. 074-1802, 074-1516 or 074-1802) were purchased from KPL,
Inc. The following reagents were used in the present study: SSD
(cat. no. A0259; Chengdu Must Bio-Technology Co. Ltd.), bafilomycin
A1 (Baf; cat. no. 11038; Cayman Chemical Company), rapamycin (Rapa;
cat. no. S1039; Selleck Chemicals), SB230580 (p38 MAPK inhibitor;
cat. no. S1076; Selleck Chemicals), LysoTracker™ Red DND-99 (cat.
no. L7528; Thermo Fisher Scientific, Inc.) DAPI (cat. no. C1006;
Beyotime Institute of Biotechnology) and DMSO (as solvent and
placebo; cat. no. D2650; Sigma-Aldrich; Merck KGaA).
Cell culture
MDA-MB-231 cells were purchased from the American
Type Culture Collection. The MDA-MB-231 cells were cultured in DMEM
(cat. no. C11995500bt; Gibco, Thermo Fisher Scientific, Inc.),
supplemented with 10% FBS (cat. no. 6140071; Gibco, Thermo Fisher
Scientific, Inc.), and maintained in a humidified atmosphere at
37°C and 5% CO2.
MTT cell viability assay
A total of 5×103 cells/well were cultured
at 37°C and 5% CO2 for 24 h in 96-well plates. Following
incubation with SSD (0, 2, 4, 6, 10, 12 or 15 µM) for 24 h, 20 µl
5% MTT solution was added to each well and incubated at 37°C and 5%
CO2 for 4 h. Subsequently, the medium was discarded and
the purple formazan was dissolved in 150 µl DMSO/well. The
absorbance was detected using a microplate reader (iMark™, Bio-Rad
Laboratories, Inc.) at 495 nm. The half maximal inhibitory
concentration (IC50) was calculated using SPSS 17.0
software (SPSS, Inc.).
Flow cytometric analysis of
apoptosis
A total of 1×105 cells/well were cultured
in 6-well plates at 37°C and 5% CO2 for 24 h. Following
incubation with SSD (8 or 10 µM) for 24 h or pretreated with 10 µM
SB203580 for 2 h and then exposed to 10 µM SSD at 37°C for 24 h,
the cells were collected by centrifuging at 600 × g for 5 min at
4°C, then the cells pellet were resuspended in 1 ml PBS and
centrifuged at 600 × g for 5 min at 4°C again. Subsequently the
cells pellet were stained with 10 µl propidium iodide (PI; cat. no.
P4170; Sigma-Aldrich; Merck KGaA; 5.0 µg/ml) and 5 µl Annexin
V-FITC (cat. no. 556419; BD Pharmingen). Apoptotic cells were
subsequently detected by flow cytometry (Accuri C6; BD Diagnostics;
Becton, Dickinson & Company) and analyzed by FlowJo 7.6.1
software (FlowJo LLC).
Plasmids and establishment of stable
cell lines
A total of 5×103 cells/well were cultured
in 24-well plates at 37°C and 5% CO2 for 24 h, then
transfected with 500 ng plasmids for 24 h before the subsequent
experimentation using Lipofectamine® 3000 reagent (cat.
no. L3000015; Invitrogen; Thermo Fisher Scientific, Inc.) according
to the manufacturer's protocol. The plasmids include LAMP1-mGFP
plasmid (cat. no. 34831; Addgene, Inc.), tfLC3 plasmid (a tandem
reporter construct carrying both EGFP-LC3 and mRFP-LC3; cat. no.
21074; Addgene, Inc. and mRFP-LC3 plasmid (cat. no. 21075; Addgene,
Inc.). The short hairpin (sh)RNA plasmids of target ATG5 (shATG5;
5′-TTTCATTCAGAAGCTGTTT-3′) and non-specific control (shCon;
5′-TTCTCCGAACGTGTCACGT-3′) were purchased from Gene Chem Co. Ltd.
(Shanghai, China). A total of 1×106 293FT cells cultured
in 60 mm plate were co-transfected with lentiviral packaging
vectors (cat. no. A43237, Invitrogen; Thermo Fisher Scientific,
Inc.) along with 1.5 µg shATG5 plasmin or 1.5 µg shCon plasmid
using Lipofectamine® 3000 (cat. no. L3000015;
Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocols. Supernatant containing the lentivirus was
harvested after 48 h. A total of 1.5×105 MDA-MB-231
cells cultured in 6-well were infected with 1 ml lentivirus for 24
h, subsequently knockdown stable cell lines were selected using 4
µg/ml puromycin (cat. no. P9620; Sigma-Aldrich; Merck KGaA).
Confocal fluorescence microscopy
detection
Cells (5×103 cells/well) transfected with
LAMP1-mGFP, tfLC3 or mRFP-LC3 were treated with 8 µM SSD, 20 nM Baf
or 0.25 µM Rapa for 24 h. Lysosomes were stained with 75 nM
LysoTracker™ Red DND-99 at 37°C for 2 h. The fluorescence in the
cells was detected using a Carl Zeiss LSM 780NLO confocal
microscopy with a 63× oil objective.
Western blotting
Total protein was extracted from cells using RIPA
lysis buffer (cat. no. P0013; Beyotime Institute of Biotechnology).
The concentration of protein was determined by the BCA method and
15–60 µg proteins were separated by 10–12% SDS-PAGE. The separated
proteins were subsequently transferred onto PVDF membranes and
blocked with 5% milk at room temperature for 30 min. The membranes
were incubated with the aforementioned primary antibodies at 4°C
overnight. Following the primary antibody incubation, the membranes
were incubated with the secondary antibodies (1:10,000) at room
temperature for 2 h. The membrane was washed three times for 30 min
each and total protein was visualized using an ECL substrate (cat.
no. 170–5060; Bio-Rad Laboratories, Inc.). The densitometric
analysis was performed by ImageJ 1.37C software (National
Institutes of Health).
Statistical analysis
Data were expressed as the means ± standard
deviation from three independent experiments. Statistical analysis
was performed using SPSS 17.0 software (SPSS, Inc.). Statistical
differences among multiple groups were compared using a one-way
ANOVA and Scheffe post hoc test. The statistical differences among
two groups were determined using a Student's t-test if the data
followed a normal distribution or with a U-Mann Whitney test
otherwise. P<0.05 was considered to indicate a statistically
significant difference.
Results
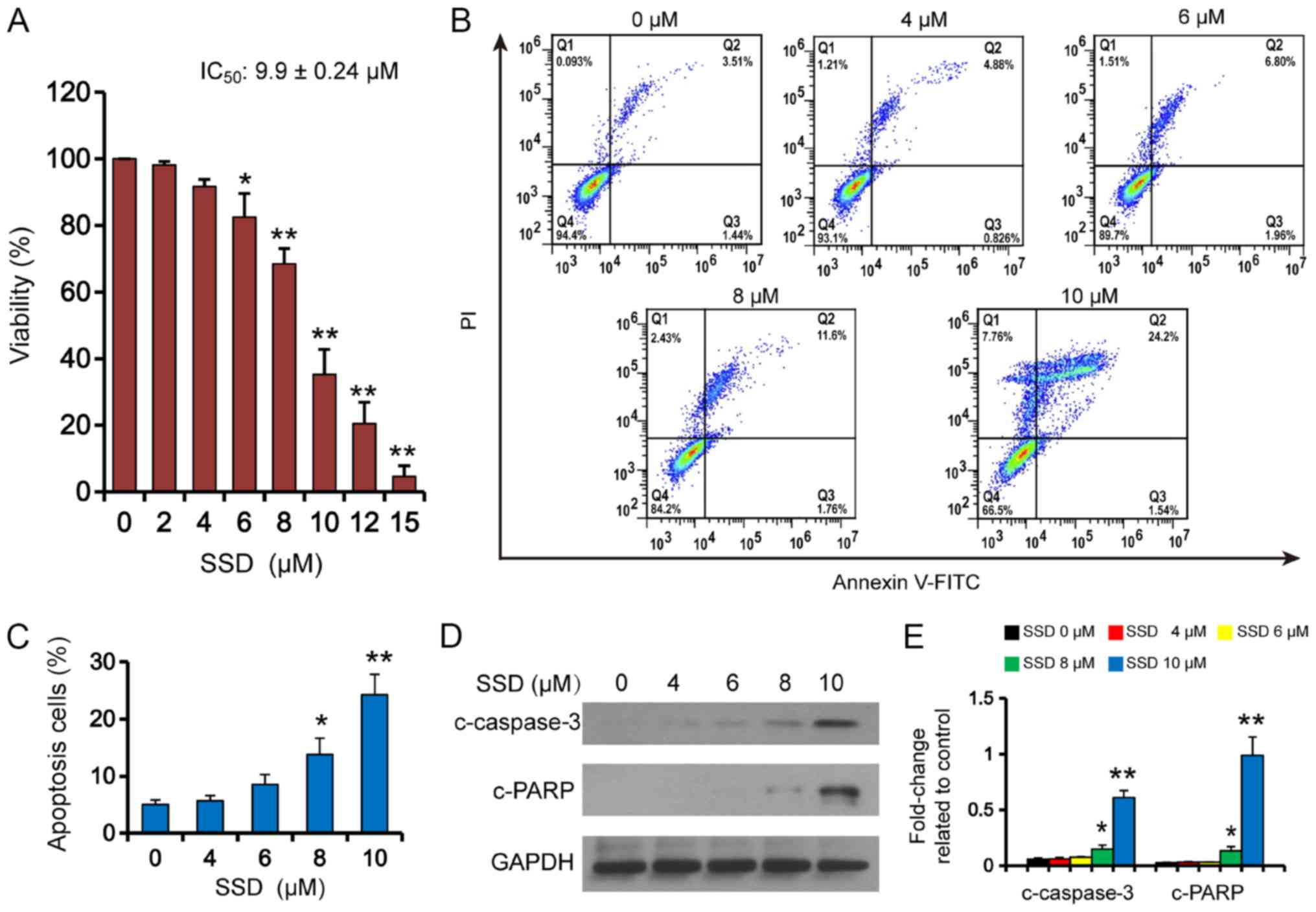
SSD induces apoptosis in human breast
cancer MDA-MB-231 cells
The cytotoxic effects of SSD were tested in
MDA-MB-231 cells using an MTT assay. 6–15 µM SSD significantly
inhibited the viability of MDA-MB-231 cells in a dose dependent
manner (Fig. 1A); IC50
was found to be 9.9±0.24 µM. Flow cytometric analysis of apoptosis
also demonstrated that 8–10 µM SSD significantly increased the rate
of apoptosis in MDA-MB-231 cells (Fig.
1B and C). Consistent with these findings, 8–10 µM SSD was also
revealed to promote the cleavage of caspase-3 and PARP (Fig. 1D and E).
SSD activates the p38 MAPK signaling
pathway
The MAPK cascade is an important signaling pathway
involved in the apoptotic process (17). JNK, ERK and p38 MAPK all belong to
MAPK signaling pathways (18). To
understand whether these MAPKs were involved in SSD-mediated
apoptosis, the protein expression levels of p-JNK, JNK, p-ERK1/2,
ERK1/2, p-p38 and p38 were detected using western blotting. SSD
promoted a significant increase in p-p38 expression levels;
however, it did not affect the expression levels of p-JNK or p-ERK
(Fig. 2A and B). These results
indicated that p38 MAPK may be involved in SSD-mediated
apoptosis.
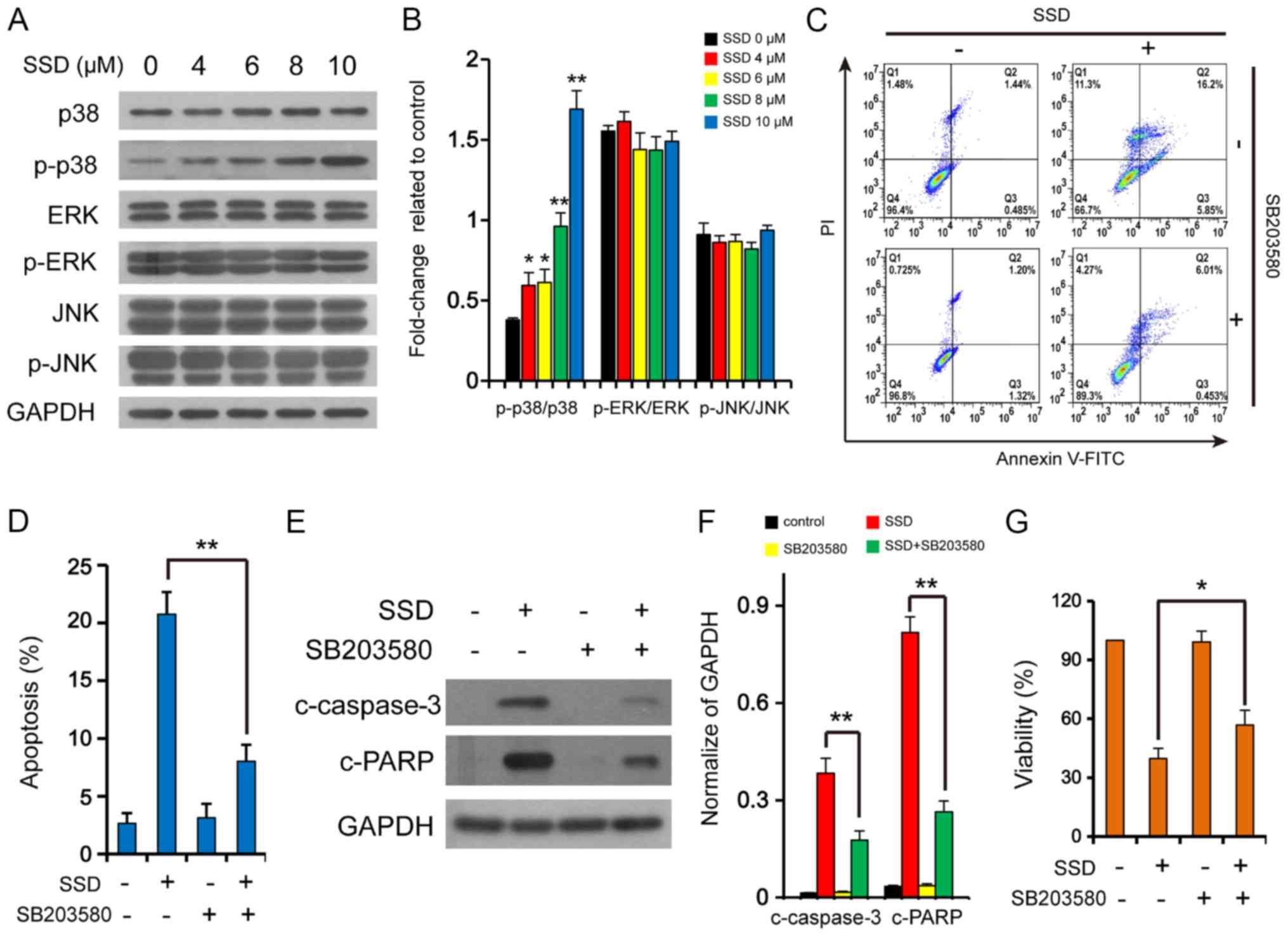 | Figure 2.SSD activates the p38
mitogen-activated protein kinase signaling pathway. (A) MDA-MB-231
cells were exposed to SSD for 24 h and the expression levels of
JNK, p-JNK, ERK, p-ERK, p38 and p-p38 were detected by western
blotting (n=3). (B) Semi-quantitative analysis of the expression
levels from part A. *P<0.05, **P<0.01 vs. 0 µM. MDA-MB-231
cells were pretreated with 10 µM SB203580 for 2 h and then exposed
to 10 µM SSD for 24 h. (C) Apoptotic cells were determined using
flow cytometry (n=3). (D) Quantitative analysis of the apoptotic
rate. **P<0.01. (E) Expression levels of c-caspase-3 and c-PARP
were detected using western blotting. (F) Semi-quantitative
analysis of the expression levels in part E. **P<0.01 vs.
control. (G) Cell viability was determined using MTT assays (n=3).
*P<0.05. Data are presented as the mean ± SD. SSD, saikosaponin
D; c-, cleaved; p-, phosphorylated; PI, propidium iodide; PARP,
poly (ADP-ribose) polymerase. |
The co-administration of SB203580, a p38-MAPK
inhibitor, was found to significantly attenuate the apoptotic rate
induced by SSD (Fig. 2C and D),
reduce the upregulation of c-caspase-3 and c-PARP expression levels
mediated by SSD (Fig. 2E and F),
and weaken the inhibitory effect on cell viability mediated by SSD
in MDA-MB-231 cells (Fig. 2G).
These results further suggested that SSD may induce apoptosis
through the p38 MAPK signaling pathway.
SSD inhibits autophagic degradation in
MDA-MB-231 cells
ATGs are important for autophagy (19). The treatment of cells with SSD
promoted the accumulation of LC3-II and p62; however, it did not
affect the expression levels of Beclin 1, ATG5, ATG7, p-ULK1, ULK1,
p-mTOR or mTOR (Fig. 3A and B).
These findings indicated that SSD may not promote autophagosome
formation but inhibit autophagic degradation (20). To further confirm these
observations, tfLC3 plasmid (a tandem reporter construct carrying
both EGFP-LC3 and mRFP-LC3) was applied (21). Cells transfected with tfLC3 were
treated with Baf, Rapa and SSD and the fluorescence of EGFP-LC3 and
mRFP-LC3 puncta were detected using confocal laser microscopy. Both
Baf and SSD treatment induced the formation of a large number of
both red and green puncta in cells, which displayed a yellow
overlay (Fig. 3C). However, only
red puncta were detected in cells treated with Rapa. These findings
indicated that SSD blocked the process of autophagic
degradation.
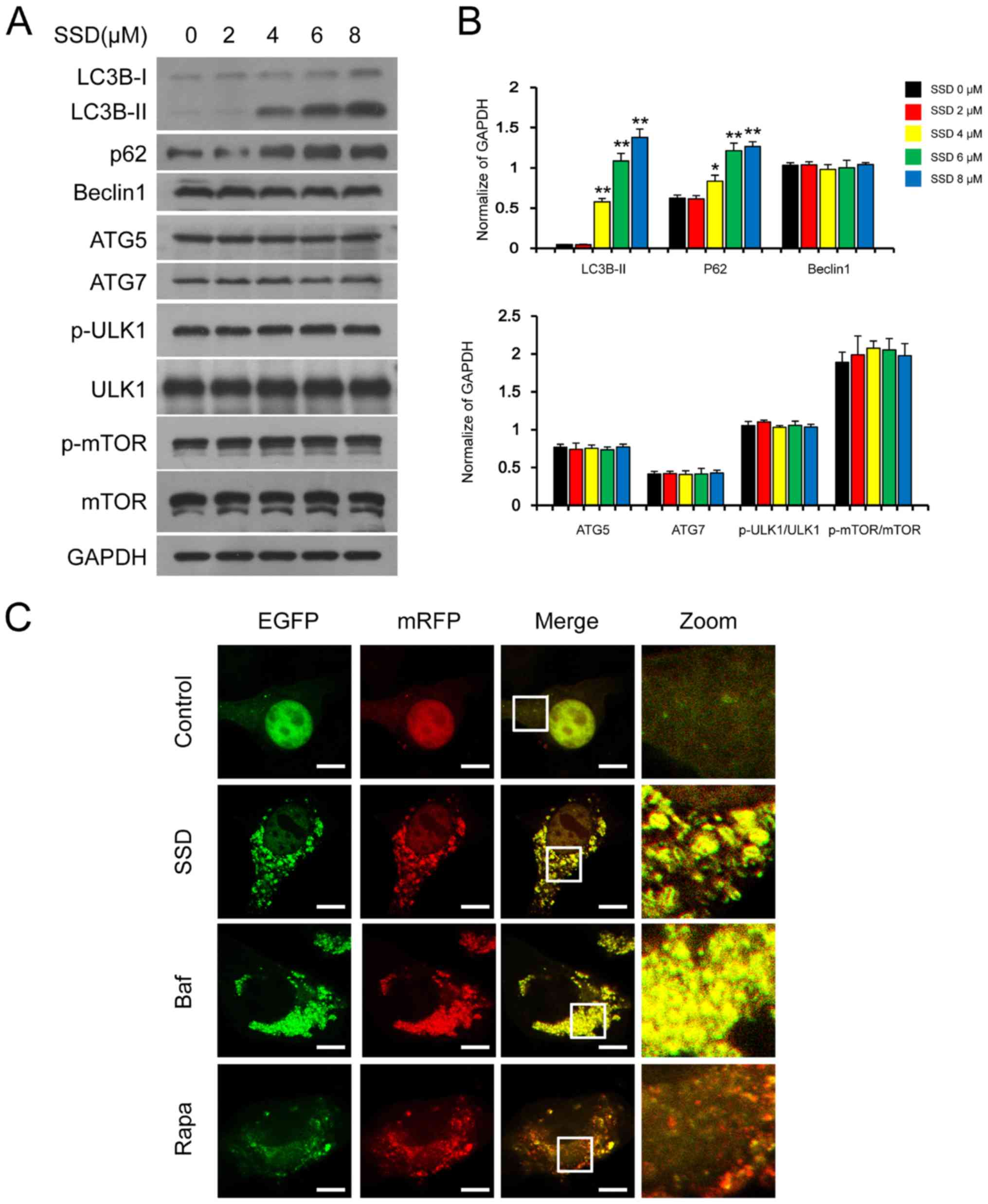 | Figure 3.SSD inhibits autophagic degradation.
(A) MDA-MB-231 cells were exposed to SSD for 24 h and the
expression levels of LC3, p62, Beclin 1, p-mTOR, mTOR, ATG7, ATG5,
p-ULK1 and ULK1 were analyzed using western blotting. (B)
Semi-quantitative analysis of the expression levels of part A. n=3;
*P<0.05, **P<0.01 vs. 0 µM. (C) Cells were transfected with
tfLC3 and exposed to 8 µM SSD, 0.25 µM Rapa or 20 nM Baf for 24 h.
The EGFP and mRFP puncta were observed using confocal laser
microscopy (scale bar=10 µm). SSD, saikosaponin D; p-,
phosphorylated; ATG, autophagy-related protein; Baf, Bafilomycin
A1; Rapa, rapamycin; LC3B, microtubule-associated protein 1 light
chain 3 β; ULK1, serine/threonine-protein kinase ULK1. |
SSD blocks autophagic degradation by
inhibiting autophagosome-lysosome fusion
LAMP1 and LAMP2, major lysosomal membrane proteins,
are important for maintaining lysosomal function and assisting in
autophagosome-lysosome fusion (22). Cathepsin B and D, types of
lysosomal cathepsins, are also important for maintaining lysosomal
function and the loss of lysosomal function is often accompanied by
a decrease in cathepsin expression (23). The expression levels of LAMP1 and
LAMP2 were significantly upregulated by SSD treatment compared with
untreated cells; however, the expression levels of cathepsin B and
D were not affected, suggesting that SSD may not impair lysosomal
function (Fig. 4A and B).
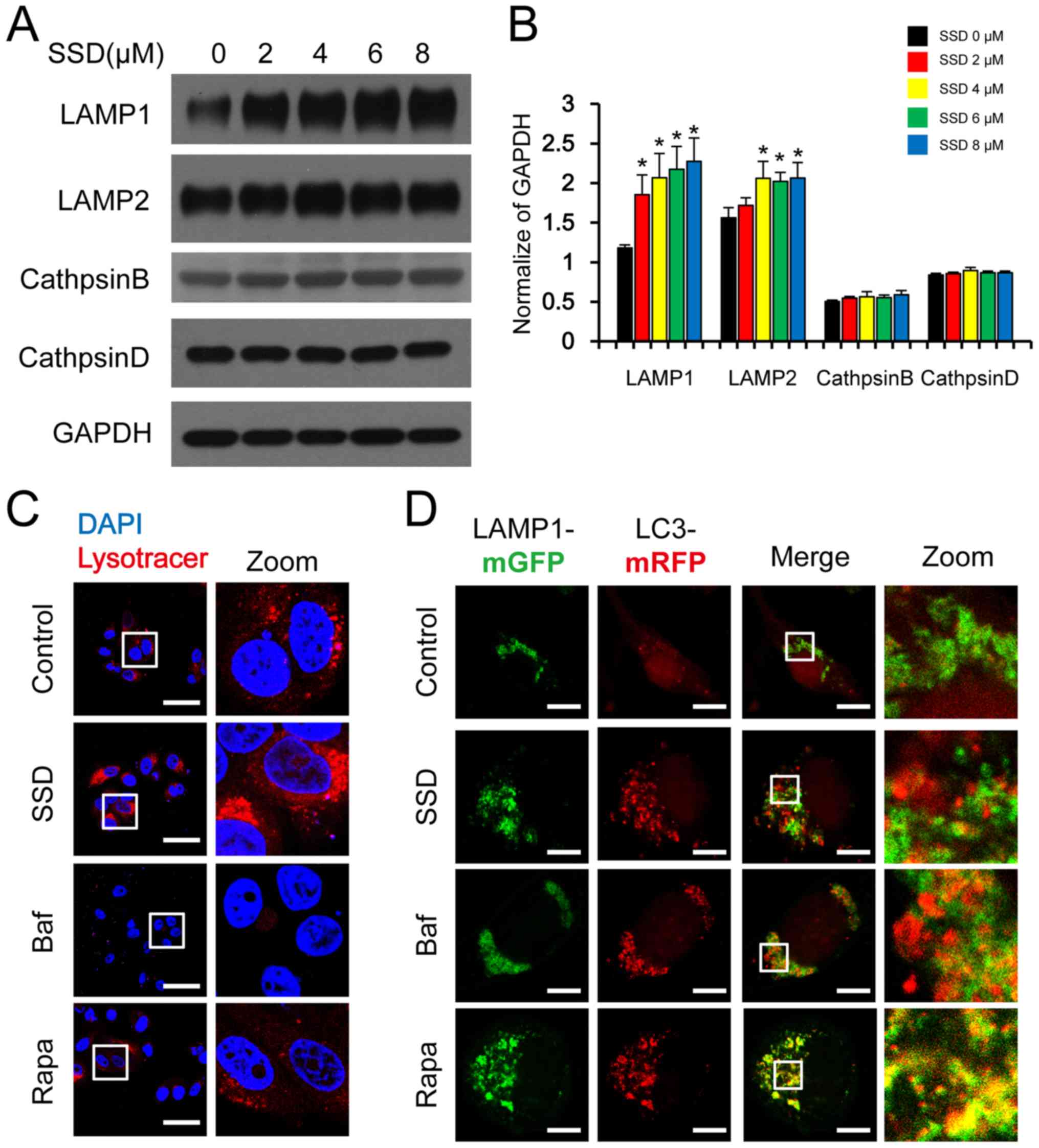 | Figure 4.SSD blocks autophagosome-lysosome
fusion. (A) MDA-MB-231 cells were exposed to SSD for 24 h. The
expression levels of LAMP1, LAMP2, cathepsin B and cathepsin D were
analyzed using western blotting. (B) Semi-quantitative analysis of
part A. Data are presented as the mean ± SD; n=3; *P<0.05 vs.
SSD 0 µM. (C) MDA-MB-231 cells were exposed to 8 µM SSD, 20 nM Baf
or 0.25 µM Rapa for 24 h and then stained with LysoTracker™ Red.
Fluorescence was observed using a confocal laser microscope (scale
bar=50 µm) (D) MDA-MB-231 cells were co-transfected with mRFP-LC3
and LAMP1-mGFP plasmids and then exposed to 8 µM SSD, 20 nM Baf or
0.25 µM Rapa for 24 h. The fluorescence of mRFP-LC3 and LAMP1-mGFP
puncta was observed by confocal laser microscopy (scale bar=10 µm).
SSD, saikosaponin D; Baf, bafilomycin A1; Rapa, rapamycin; LAMP,
lysosome-associated membrane glycoprotein; GFP, green fluorescent
protein; RFP, red fluorescent protein. |
The intralysosomal acidic environment is another
important factor for lysosomal functions (24). The intralysosomal acidic
environment can be detected using LysoTracker™ Red DND-99, an
acidic organelle tracer that can emit deep red fluorescence
(25). The red fluorescence was
observed in cells treated with SSD, Rapa or the placebo, but not in
cells treated with Baf. These results indicated that SSD may not
affect the intralysosomal acidic environment (Fig. 4C).
Detecting the colocalization of LAMP1 and LC3 can be
used to confirm autophagosome-lysosome fusion (26). There was no overlap between the
green fluorescence of LAMP1-mGFP and the red fluorescence of
mRFP-LC3 in cells treated with SSD or Baf; however, a yellow
overlap could be observed in cells treated with Rapa (Fig. 4D), suggesting that SSD may impair
the autophagosome-lysosome fusion, resulting in the inhibition of
autolysosome formation, which eventually leads to the blockage of
autophagic degradation.
Excessive accumulation of
autophagosomes does not contribute to SSD-mediated apoptosis
To further understand whether the excessive
accumulation of autophagosomes contributed to the SSD-mediated
apoptosis, the expression levels of LC3B-II were reduced following
the stable knockdown of ATG5 expression levels and the treatment
with SSD (Fig. 5A and B). Knocking
down ATG5 expression levels did not affect the apoptotic rate of
SDD-treated cells (Fig. 5C and D)
or the inhibitory effect of SDD on cell proliferation (Fig. 5E) in MDA-MB-231 cells. In addition,
the knockdown of ATG5 did not alter the SSD-mediated activation of
c-caspase-3, c-PARP or p-p38 (Fig. 5A
and B). These results indicated that the excessive accumulation
of autophagosomes may not contribute to the SSD-mediated apoptosis
in MDA-MB-231 cells.
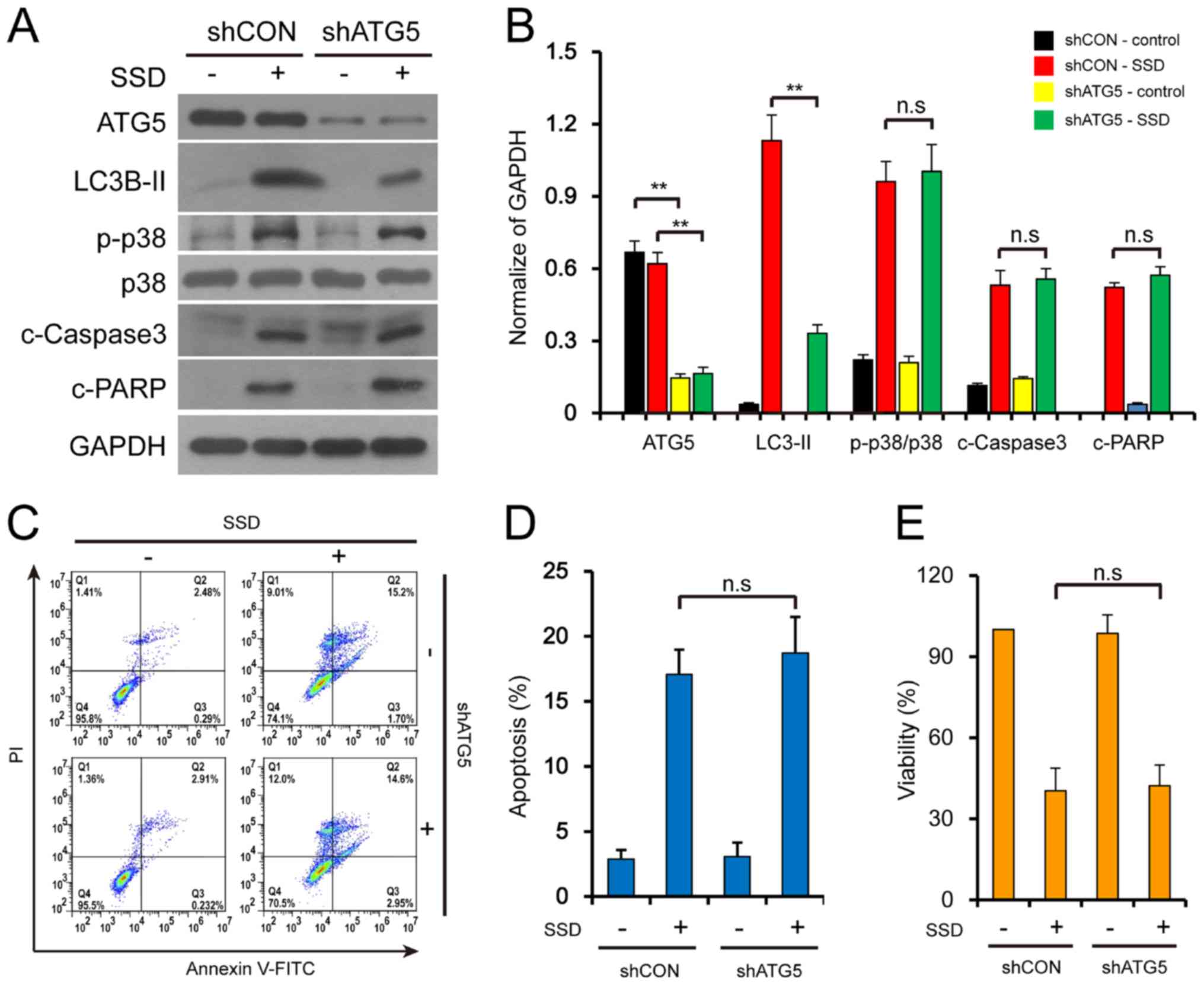 | Figure 5.Excessive accumulation of
autophagosomes does not contribute to SSD-mediated apoptosis.
Stable knockdown MDA-MB-231 cells (shCON or shATG5) were exposed to
10 µM SSD for 24 h. (A) Following treatment, the expression levels
of ATG5, LC3B-II, p-p38, p38, c-caspase-3 and c-PARP were detected
using western blotting. (B) Semi-quantitative analysis of the
expression levels in part A, n=3; **P<0.01. (C) Apoptotic cells
were determined using flow cytometry. (D) Quantitative analysis of
the apoptotic rate form part C. n=3. (E) Cell viability was
determined using MTT assays. n=3. All data are presented as the
mean ± SD. sh, small hairpin RNA; CON, control; ATG,
autophagy-related protein; c-, cleaved; n.s., not significant; p-,
phosphorylated; LC3B, microtubule-associated protein 1 light chain
3 β; PARP, poly (ADP-ribose) polymerase; SSD, saikosaponin D; PI,
propidium iodide. |
Discussion
The present study provided evidence to suggest that
SSD may induce apoptosis in MDA-MB-231 cells through the activation
of p38 MAPK signaling. In addition, the current results revealed
that SSD blocked autophagic degradation through the inhibition of
autophagosome-lysosome fusion and that the blockade of autophagic
degradation was not involved in SSD-mediated apoptosis.
SSD, a kind of triterpenoid saponins extracted from
Radix bupleuri, has been demonstrated to have an inhibitory
effect on tumor cells. For example, SSD suppressed liver cancer
proliferation by inhibiting cyclooxygenase 2 expression (27), as well as inhibiting the
proliferation, invasion and migration of osteosarcoma cells
(28). Previous studies also
reported that SSD inhibited the proliferative activity of breast
cancer cells, such as HCC1937 and MCF-7 (29,30).
The results of the present study revealed that SSD induced
apoptosis and inhibited the proliferative activity of MDA-MB-231
breast cancer cells.
p38 MAPK, one of the MAPK family members, serves an
important role in differentiation, growth inhibition, cell cycle
arrest and apoptosis (31). Thus,
p38 demonstrates potential to be developed into a cancer inhibitor,
as the phosphorylation of p38 is important for the induction of
apoptosis (32,33). The present results confirmed that
SSD induced apoptosis through the phosphorylation/activation of the
p38 MAPK signaling pathway. First, SSD induced the
phosphorylation/activation of p38 MAPK. Second, the p38 MAPK
inhibitor SB203580 reversed the SSD-mediated increase of apoptosis,
the inhibitory effect over cell viability, and the activation of
caspase-3 and PARP.
Numerous studies have reported that autophagy can be
blocked at different stages of the autophagic flux (34–37).
For example, 3-MA inhibited the formation of autophagosomes by
inhibiting phosphatidylinositol 3-kinase catalytic subunit type 3
(38). In addition, Baf was found
to disrupt the autophagic flux by changing the lysosomal acidity
through inhibiting H+ V-ATPase (39). It is also important to maintain the
normal function of lysosomes during autophagy, since lysosome
dysfunction resulted in the blockade of autophagic degradation
(40). LAMP1 and LAMP2, two major
lysosomal membrane proteins, are important for maintaining
lysosomal function and assisting autophagosome-lysosome fusion, and
the disruption of both LAMP1 and LAMP2 was found to result in the
increased accumulation of autophagosomes (22). Lysosomal cathepsins (cathepsin B
and cathepsin D) are also essential for maintaining normal lysosome
function, and the inhibition of lysosomal cathepsins has been
observed to result in the blockage of autophagic degradation
(41). In addition, the blockade
of autophagosome-lysosome fusion inhibited the formation of
autolysosomes, which subsequently resulted in the inhibition of the
autophagic flux (42).
The results of the present study confirmed that SSD
was unable to impair the lysosomal function; however, it inhibited
the fusion of autophagosomes and lysosomes, resulting in the
inhibition of autolysosome formation. First, SSD treatment did not
affect the acidic environment of the lysosomes; second, SSD
treatment did not reduce the expression levels of LAMP1 or LAMP2;
third, SSD treatment did not affect the expression levels of the
lysosomal cathepsins; and finally, SSD treatment disrupted the
co-localization of LC3 and LAMP1.
Cell death may be caused by either the induction or
inhibition of autophagy (43,44).
However, previous research has reported that classical autophagy
degradation inhibitors, such as ammonium chloride chloroquine and
Baf can cause autophagy-independent cell death (9). Consistent with these findings,
results of the current study also showed that the accumulation of
autophagosomes was not involved in SSD-mediated apoptosis. The
genetic knockdown of ATG5 expression levels significantly decreased
the SSD-mediated accumulation of LC3B-II; however, it did not
affect the SSD-mediated apoptotic rate, or the activation of
caspase-3, PARP or p38 MAPK. These results indicated that the
blockade of autophagic degradation and apoptosis, both of which are
mediated by SSD, may be two separate events. Thus, these findings
provided additional evidence to support the association between the
inhibition of autophagy degradation and cell death.
Acknowledgements
The authors would like to thank Professor Tamotsu
Yoshimori (Department of Genetics, Osaka University, Osaka, Japan)
for providing the mRFP-LC3 and tfLC3 plasmids, and Professor
Esteban C. Dell'Angelica (Department of Human Genetics, University
of California, Los Angeles, USA) for providing the LAMP1-mGFP
plasmid.
Funding
The present study was funded by the project of
Science and Health Combined Traditional Chinese Medicine of
Chongqing (grant no. ZY201801004) and the project of Science and
Health Combined Medical of Chongqing (grant no. 2018MSXM002).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
RF and JC conceived and designed the study; RF, LZ
and YL performed the experiments; RF, LZ and HX designed the
experiments; YM, ZL and BL analyzed the data; BL and JC reviewed
and edited the manuscript; RF and HX wrote the original manuscript;
and JC and BL acquired the funding. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yin Z, Pascual C and Klionsky DJ:
Autophagy: Machinery and regulation. Microb Cell. 3:588–596. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bostancıklıoğlu M: An update on the
interactions between Alzheimer's disease, autophagy and
inflammation. Gene. 705:157–166. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jiang X, Overholtzer M and Thompson CB:
Autophagy in cellular metabolism and cancer. J Clin Invest.
125:47–54. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
White E: The role for autophagy in cancer.
J Clin Invest. 125:42–46. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Levy JMM, Towers CG and Thorburn A:
Targeting autophagy in cancer. Nat Rev Cancer. 17:528–542. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chude CI and Amaravadi RK: Targeting
autophagy in cancer: Update on clinical trials and novel
inhibitors. Int J Mol Sci. 18:12792017. View Article : Google Scholar
|
|
7
|
Zhou J, Li G, Zheng Y, Shen HM, Hu X, Ming
QL, Huang C, Li P and Gao N: A novel autophagy/mitophagy inhibitor
liensinine sensitizes breast cancer cells to chemotherapy through
DNM1L-mediated mitochondrial fission. Autophagy. 1:1259–1279. 2015.
View Article : Google Scholar
|
|
8
|
Zhou J, Hu SE, Tan SH, Cao R, Chen Y, Xia
D, Zhu X, Yang XF, Ong CN and Shen HM: Andrographolide sensitizes
cisplatin-inducedapoptosis via suppression of autophagosome
lysosome fusion in human cancer cells. Autophagy. 8:338–349. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Stamenkovic M, Janjetovic K, Paunovic V,
Ciric D, Kravic-Stevovic T and Trajkovic V: Comparative analysis of
cell death mechanisms induced by lysosomal autophagy inhibitors.
Eur J Pharmacol. 859:1725402019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang YY, Tang YZ, Fan CL, Luo HT, Guo PR
and Chen JX: Identification and determination of the saikosaponins
in Radix bupleuri by accelerated solvent extraction combined
with rapid-resolution LC-MS. J Sep Sci. 33:1933–1945. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tsuyoshi H, Wong VKW, Han Y, Orisaka M,
Yoshida Y and Tsang BK: Saikosaponin-d, a calcium mobilizing agent,
sensitizes chemoresistant ovarian cancer cells to cisplatin-induced
apoptosis by facilitating mitochondrial fission and G2/M arrest.
Oncotarget. 8:99825–99840. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen X, Liu C, Zhao R, Zhao P, Wu J, Zhou
N and Ying M: Synergetic and antagonistic molecular effects
mediated by the feedback loop of p53 and JNK between Saikosaponin D
and SP600125 on lung cancer A549 cells. Mol Pharm. 15:4974–4984.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chun YZ, Zhong MJ, Xiao FM, Yue L, Xiao
ZL, Li LL, Wen HW and Tao W: Saikosaponin-d inhibits the hepatoma
cells and enhances chemosensitivity through SENP5-dependent
inhibition of Gli1 SUMOylation under hypoxia. Front Pharmacol.
10:1039–1052. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li Y, Cai T, Zhang W, Zhu W and Lv S:
Effects of Saikosaponin D on apoptosis in human U87 glioblastoma
cells. Mol Med Rep. 16:1459–1464. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wong VK, Li T, Law BY, Ma ED, Yip NC,
Michelangeli F, Law CK, Zhang MM, Lam KY, Chan PL and Liu L:
Saikosaponin-d, a novel SERCA inhibitor, induces autophagic cell
death in apoptosis-defective cells. Cell Death Dis. 4:e7202013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li C, Huang L, Sun W, Chen Y, He ML, Yue J
and Ballard H: Saikosaponin D suppresses enterovirus A71 infection
by inhibiting autophagy. Signal Transduct Target Ther. 4:42019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sui X, Kong N, Ye L, Han W, Zhou J, Zhang
Q, He C and Pan H: p38 and JNK MAPK pathways control the balance of
apoptosis and autophagy in response to chemotherapeutic agents.
Cancer Lett. 344:174–179. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu RL, Yun JK, Myung HK and June MK: MAPK
cascades in guard cell signal transduction. Front Plant Sci.
7:80–87. 2016.PubMed/NCBI
|
|
19
|
Mizushima N, Yoshimori T and Ohsumi Y: The
role of Atg proteins in autophagosome formation. Annu Rev Cell Dev
Biol. 27:107–132. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Katsuragi Y, Ichimura Y and Komatsu M:
p62/SQSTM1 functions as a signaling hub and an autophagy adaptor.
FEBS J. 282:4672–4678. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kimura S, Noda T and Yoshimori T:
Dissection of the autophagosome maturation process by a novel
reporter protein, tandem fluorescent-tagged LC3. Autophagy.
3:452–460. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Eskelinen EL: Roles of LAMP-1 and LAMP-2
in lysosome biogenesis and autophagy. Mol Aspects Med. 27:495–502.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Siong Tan HW, Anjum B, Shen HM, Ghosh S,
Yen PM and Sinha RA: Lysosomal inhibition attenuates peroxisomal
gene transcription via suppression of PPARA and PPARGC1A levels.
Autophagy. 15:1455–1459. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Colacurcio DJ and Nixon RA: Disorders of
lysosomal acidification-The emerging role of v-ATPase in aging and
neurodegenerative disease. Ageing Res Rev. 32:75–88. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Woolbright BL, Ramachandran A, McGill MR,
Yan HM, Bajt ML, Sharpe MR, Lemasters JJ and Jaeschke H: Lysosomal
instability and cathepsin B release during acetaminophen
hepatotoxicity. Basic Clin Pharmacol Toxicol. 111:417–425. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Miyagawa K, Oe S, Honma Y, Izumi H, Baba R
and Harada M: Lipid-induced endoplasmic reticulum stress impairs
selective autophagy at the step of autophagosome-lysosome fusion in
hepatocytes. Am J Pathol. 186:1861–1873. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ren M, McGowan E, Li Y, Zhu X, Lu X, Zhu
Z, Lin Y and He S: Saikosaponin-d Suppresses COX2 through
p-STAT3/C/EBPβ signaling pathway in liver cancer: A novel mechanism
of action. Front Pharmacol. 10:6232019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gao T, Zhao P, Yu X, Cao S, Zhang B and
Dai M: Use of Saikosaponin D and JNK inhibitor SP600125, alone or
in combination, inhibits malignant properties of human osteosarcoma
U2 cells. Am J Transl Res. 11:2070–2080. 2019.PubMed/NCBI
|
|
29
|
Li C, Guan X, Xue H, Wang P, Wang M and
Gai X: Reversal of P-glycoprotein-mediated multidrug resistance is
induced by saikosaponin D in breast cancer MCF-7/adriamycin cells.
Pathol Res Pract. 213:848–853. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang J, Qi H, Zhang X, Si W, Xu F, Hou T,
Zhou H, Wang A, Li G, Liu Y, et al: Saikosaponin D from Radix
bupleuri suppresses triple-negative breast cancer cell growth
by targeting β-catenin signaling. Biomed Pharmacother. 108:724–733.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cuadrado A and Nebreda AR: Mechanisms and
functions of p38 MAPK signalling. Biochem J. 429:403–417. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen W, Tan Y and Zhang Y: p38 MAPK
signaling pathway activation by phenyl benzoxime in SNU-306 cells
causes induction of apoptosis. Microb Pathog. 126:74–78. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou Q, Wu X, Wen C, Wang H, Wang H, Liu H
and Peng J: Toosendanin induces caspase-dependent apoptosis through
the p38 MAPK pathway in human gastric cancer cells. Biochem Biophys
Res Commun. 505:261–266. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gewirtz DA: The challenge of developing
autophagy inhibition as a therapeutic strategy. Cancer Res.
76:5610–5614. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Palmeira dos Santos C, Pereira GJ, Barbosa
CM, Jurkiewicz A, Smaili SS and Bincoletto C: Comparative study of
autophagy inhibition by 3MA and CQ on cytarabine-induced death of
leukaemia cells. J Cancer Res Clin Oncol. 140:909–920. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Somya V and Ravi M: A reversible autophagy
inhibitor blocks autophagosome-lysosome fusion by preventing Stx17
loading onto autophagosomes. Mol Biol Cell. 30:2283–2295. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yu L, Wu WK, Gu C, Zhong D, Zhao X, Kong
Y, Lin Q, Chan MT, Zhou Z and Liu S: Obatoclax impairs lysosomal
function to block autophagy in cisplatin-sensitive and -resistant
esophageal cancer cells. Oncotarget. 7:14693–14707. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dong LL, Yang Y, Quan L and Jian JW:
Inhibition of autophagy by 3-MA potentiates cisplatin-induced
apoptosis in esophageal squamous cell carcinoma cells. Med Oncol.
28:105–111. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mauvezin C and Neufeld TP: Bafilomycin A1
disrupts autophagic flux by inhibiting both V-ATPase-dependent
acidification and Ca-P60A/SERCA-dependent autophagosome-lysosome
fusion. Autophagy. 11:1437–1438. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Malicdan MC and Nishino I: Autophagy in
lysosomal myopathies. Brain Pathol. 22:82–88. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Man SM and Kanneganti TD: Regulation of
lysosomal dynamics and autophagy by CTSB/cathepsin B. Autophagy.
12:2504–2505. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Mauthe M, Orhon I, Rocchi C, Zhou X, Luhr
M, Hijlkema KJ, Coppes RP, Engedal N, Mari M and Reggiori F:
Chloroquine inhibits autophagic flux by decreasing
autophagosome-lysosome fusion. Autophagy. 14:1435–1455. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shimizu S, Yoshida T, Tsujioka M and
Arakawa S: Autophagic cell death and cancer. Int J Mol Sci.
15:3145–3153. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhang H, Ge S, He K, Zhao X, Wu Y, Shao Y
and Wu X: FoxO1 inhibits autophagosome-lysosome fusion leading to
endothelial-apoptosis in diabetes. Cardiovasc Res. 115:2008–2020.
2019. View Article : Google Scholar : PubMed/NCBI
|