Introduction
Tyrosine phosphorylation is a post-translational
protein modification, which serves as a rapid on/off switch for
cellular machinery under regulated conditions. It is used in cell
signaling pathways to regulate cell growth, migration, adhesion,
differentiation and cell survival (1). This protein modification is regulated
by tyrosine kinases; c-Abl tyrosine kinase is a crucial
non-receptor tyrosine kinase responsible for the tyrosine
phosphorylation of several proteins, mostly members of the DNA
repair machinery (1). c-Abl has
been associated with different aspects of the DNA damage response
(DDR) and it is considered a major player in this area (1). c-Abl binds with various nuclear
proteins implicated in different aspects of DNA repair, which
suggests that c-Abl may be associated with the regulation of
double-strand break repair (2).
The role of c-Abl in DNA repair mechanisms first came into the
spotlight in female germ cells under conditions of genotoxic stress
(2). c-Abl was also discovered to
be activated in cells upon DNA damage, whereby it successively
activated signal transduction networks required to reinstate
cellular homeostasis (3–7). It has since been found that DDR
proteins are targets of c-Abl, similar to other non-receptor
tyrosine kinases (8–11). c-Abl was also observed to
translocate between the cytosol and nucleus, which revealed its
important intermediary role in cellular signaling, capable of
transducing signals from activated tyrosine kinases directly to the
nuclear compartment to activate particular DDR pathways (8–11).
The present review aimed to summarize the different roles of c-Abl
substrates belonging to the DDR machinery. These substrates are
phosphorylated by c-Abl tyrosine kinase and are involved in
different cellular functions, such as DNA damage repair, cell
survival, tumor suppression, apoptosis, cell cycle arrest, DNA
damage regulation and immune responses (Table I). In addition, the current review
discussed the tyrosine phosphorylation events of DDR proteins
mediated by c-Abl tyrosine kinase and summarized their implications
in important cellular functions.
 | Table I.c-ABL substrates belonging to the DNA
damage response machinery: Phosphorylation sites and functional
roles. |
Table I.
c-ABL substrates belonging to the DNA
damage response machinery: Phosphorylation sites and functional
roles.
| Authors (year) | c-ABL
substrate | Phosphotyrosine
site | Function | (Refs.) |
|---|
| Chen et al
(1999) |
|
|
| (41) |
| Shimizu et
al (2009) | RAD51 | 315 | DNA recombination,
repair and survival | (42) |
| Kitao and Yuan
(2002) | RAD52 | 104 | DNA recombination,
repair and survival | (43) |
| Goldberg et
al (2002) |
|
|
| (46) |
| Carr et al
(2016) | Mouse double minute
2 | 394 | Tumor suppression
and apoptosis | (47) |
| Dias et al
(2006) | HDM2 (human homolog
of MDM2) | 276 | Tumor suppression
and apoptosis | (48) |
| Chen et al
(2016) |
|
|
| (49) |
| Xiong et al
(2006) | MDMX (MDM4) | 99 | Tumor suppression
and apoptosis | (50) |
| Reuven et al
(2015) |
Homeodomain-interacting protein kinase
2 | 360 | Tumor suppression
and apoptosis | (4) |
| Gonfloni et
al (2009) | p63 (TP63) | 149 | Tumor suppression
and apoptosis | (53) |
| Yuan et al
(1999) | P73 | 99 | Tumor suppression
and apoptosis | (54) |
| Keshet et al
(2015) | Yes-associated
protein 1 | 357 | Tumor suppression
and apoptosis | (55) |
| Yamaguchi et
al (2015) |
|
|
| (56) |
| Karimian et
al (2016) | Jun B | 173, 182 &
188 | Cell cycle
arrest | (57) |
| Jin et al
(1997) |
|
|
| (58) |
| Hartley et
al (1995) |
|
|
| (59) |
| Keith and Schreiber
(1995) | DNA-dependent
protein kinase, catalytic subunit | pYC-terminal
domain | DNA damage
regulation | (60) |
| Yi et al
(2006) | Mut S protein
homology 5 | NH2 terminus ′
1–109 | DNA damage
regulation | (61) |
| Foray et al
(2002) | Breast cancer type
1 susceptibility protein | C-terminal region
1, 314–1, 863 | c-ABL kinase
activation | (63) |
| Bohio et al
(2019) | Poly (ADP-ribose)
polymerase 1 | 829 | Immune
response | (69) |
Tyrosine phosphorylation
Historical overview
Post-translational protein modifications (for
example, tyrosine phosphorylation) were first identified in 1979;
however, it is now known that post-translational modifications have
occurred for over a billion years throughout the evolution of
unicellular eukaryotic organisms, the first multiple-celled
eukaryotic animals (12). As
regulated cell signal transduction is essential for multicellular
organisms, the development of cell surface receptor systems that
use tyrosine phosphorylation for transmembrane signal transduction
and intracellular signaling seems likely to have been an important
event in the evolution of multicellular organisms (12). Since the discovery of these
post-translational modifications, rapid progress has been made in
understanding the function of tyrosine phosphorylation. For
example, the early observation that ligand binding induced a rapid
increase in the autophosphorylation of epithelial growth factor and
platelet-derived growth factor receptor tyrosine kinases suggested
an important role for tyrosine phosphorylation in growth factor
signaling and proliferation; by extension, this function was also
suggested to be exploited in oncogenesis, through the hijacking of
the growth factor tyrosine phosphorylation signaling pathways
(13).
Significance of tyrosine
phosphorylation and signaling mechanisms
Tyrosine phosphorylation is a post-translational
protein modification, which is considered to be a major signaling
transduction and regulatory mechanism in all eukaryotic cells.
Tyrosine phosphorylation governs a number of cellular processes,
such as cell differentiation, cell proliferation, transcriptional
activation, neural transmission, cell cycle progression, aging,
metabolic homeostasis and development (14). In fact, abnormal tyrosine
phosphorylation, which is responsible for a number of human
diseases, including cancer, has prompted the development of
tyrosine kinase inhibitors (14).
Tyrosine kinases serve a prominent role in cellular differentiation
and development in metazoans; in evolutionary terms, tyrosine
kinases are present in all metazoans, where they have been
discovered to serve a major role in cell-cell signaling and
transmembrane signal transduction, a crucial function for the
establishment of a multicellular organism (14). The regulation of proteins by
tyrosine phosphorylation occurs via several mechanisms, such as
promoting electrostatic repulsion and inducing allosteric
transitions; however, the most important role of tyrosine
phosphorylation is to function as a docking site that supports
specific interactions between a tyrosine phosphorylated protein and
the protein containing a phosphotyrosine-binding (PTB) domain, such
as the Src homology 2 (SH2) domain (14).
Docking interactions are crucial for the signal
transduction downstream from receptor tyrosine kinases on the cell
surface, which are activated upon the binding of a related
extracellular ligand and in response, elicit specific cellular
outcomes (12). Thousands of
phosphotyrosine sites have been discovered; however, the function
of tyrosine phosphorylation events occurring at these sites have
not been investigated to date, to the best of our knowledge
(12). Therefore, it is possible
that a large number of these sites do not have a functional impact
and may be referred as “noise” tyrosine phosphorylation events
(12). In conclusion, this protein
modification has been revealed to regulate a number of cellular
functions and is often involved in oncogenesis (15).
c-Abl tyrosine kinase
Structural characteristics of
c-Abl
c-Abl is a non-receptor tyrosine kinase that belongs
to the ABL family of tyrosine kinases, which are located in both
the cytoplasm and nucleus of the cell (16). c-Abl tyrosine kinase has been
discovered to serve distinct roles in the cytoplasm and nucleus
(17). The c-Abl protein possesses
several defined domains, including the SH2 domain, which is located
at the N-terminus of c-Abl and has a high level attraction for
phosphorylated tyrosine residues, and the Src homology 3 (SH3)
domain, which preferentially binds to proline-rich domains
containing a PxxP motif. Interestingly, the identification of c-Abl
SH3 binding proteins has proved useful for defining its close
downstream proteins targets. c-Abl is also characterized by its
long C-terminal tail, which contains both nuclear localization and
nuclear export signals that regulate the c-Abl subcellular
localization. The C-terminal tail also contains a DNA-binding
domain, as well as F-actin and G-actin binding domains. Thus,
through direct protein-protein interactions, c-Abl (with its
structural domains) is likely to be simultaneously involved in a
number of processes (1,17,18).
Mechanisms of c-Abl activation
c-Abl is a tyrosine kinase and it is activated in
response to several extracellular and intracellular stimuli
(1). Generally, c-Abl exists in an
inactive form in the cell, and the activity of c-Abl is strongly
regulated by intramolecular bonds and by binding to other proteins,
as well as by linkage to the membranes via an amino terminal
myristoyl group. DNA damage, autophosphorylation and the actions of
other proteins have all been reported to stimulate the activation
of c-Abl (16,19). The production of reactive oxidative
species increases following DNA damage during cell stress, which
may be responsible for cell degeneration (17). In addition, multiple cell processes
are influenced by c-Abl through its interaction with other protein
kinases and it has been suggested that the prime protein targets of
c-Abl may differ in various cell types (16). The activity of c-Abl has also been
discovered to be strongly regulated and activated by ionizing
radiation, as well as different types of genotoxic offence.
According to previous studies, c-Abl has also been found to be
activated by an unclear, slow and gradual mechanism (1,20,21).
Downstream effects of c-Abl
activation
The consensus sequence for the target
phosphorylation of c-Abl is non-promiscuous. Therefore, novel
protein targets of c-Abl should be experimentally verified under
actual physiological conditions. Previously, c-Abl was investigated
as a regulator of different types of human cancer and mutations in
c-ABL were found to accelerate protein kinase activity, which was
responsible for the high levels of cell proliferation (16). A number of c-Abl protein binding
partners have been investigated; however, the functional outcomes
of these compounds remain to be determined. To investigate c-Abl
function, it is often beneficial to detect its immediate downstream
partners. For instance, this method has led to the identification
of a number of proteins, including TP53 and RAD51, which are
members of the DNA repair machinery (17); c-Abl is capable of binding to
several of these proteins (22).
c-Abl has been found to be part of an intricate network of protein
interactions, as well as phosphorylation events, in the cell and it
is expressed in all types of cells. Therefore, it was hypothesized
that c-Abl is involved in a variety of physiological functions,
such as cell growth, cell survival, autophagy, DNA repair,
motility, cytoskeleton dynamics and receptor endocytosis (16). c-Abl has also been discovered to
enhance the activity of a number of substrates to regulate DNA
double strand break repair pathways (17,23).
In addition, c-Abl has been implicated in various cell signaling
pathways, including those originating from cell adhesion, DNA
damage, growth factor stimulation and oxidative stress (24–27).
c-Abl has been found to support the DNA damage repair mechanism;
however, it has been discovered to promote apoptosis in
circumstances when DNA damage repair is not possible (28).
DNA damage response
DNA damage and repair mechanism
DNA damage occurs following the exposure to
environmental (UV light and ionizing radiation) or intracellular
(reactive oxygen species as byproducts of routine metabolic
processes) genotoxic agents (29).
Fortunately, cells are equipped with several DNA repair mechanisms,
including base excision repair (BER) that removes damaged bases,
mismatch repair that recognizes base incorporation errors and base
damage, nucleotide excision repair that removes bulky DNA adducts
and cross-link repair that removes interstrand cross-links. Breaks
in the DNA backbone are repaired by double-strand break repair
signaling pathways, including homologous recombination and
non-homologous end joining (30).
Several of these mechanisms can operate independently to repair
simple lesions, for instance, BER removes base lesions including,
and oxidation (31). DNA
glycosylases scan the DNA in order to recognize and remove the
damage, along with other proteins, such as AP endonuclease 1, DNA
polymerase β and DNA ligase in a ‘hand-off’ model, in order to
restore the DNA duplex (31).
However, the DDRs, such as DSB response, regulate the repair of
more complex lesions, which involves multiple DNA processing steps.
DDR is considered to be essential for the successful repair of the
more difficult lesions (30). In
stable or normal cells, genomic integrity is maintained by a strong
and effective DDR signaling network, which includes DNA repair
signaling pathways and cell cycle checkpoints (29). The repair of DNA damage is
necessary for RNA and DNA polymerases to accurately read and
transcribe the information in the genome. During double-strand
breaks, multiple enzymes and mediators translocate to the sites of
the lesion to release a network of DNA repair processes known as
the DDR (2). The functions of the
DDR and TP53 were found to be necessary for the determination of
cell fate in gastric cancer (32).
In addition, tumor necrosis factor receptor associated factor 1
interacting protein was discovered to be a novel regulator of
histone H2B monoubiquitination in the DDR and during cancer
development in lung adenocarcinoma (33).
Role of protein kinases in the
DDR
DNA damage arising from both internal and
environmental sources challenges the integrity of the genome. DDR
protein kinases have been discovered to regulate these repair
mechanisms (30). These kinases
phosphorylate DNA repair proteins, and they have been found to
increase the efficiency of DNA repair by initiating a series of
multiple changes in the local chromatin structure near the damage
site and by changing the overall environment of the cell to make it
more efficient for repair (30).
The DNA damage response kinases were indicated to share multiple
common regulatory mechanisms of activation (34), for example, they sense DNA damage
through protein-protein interactions that serve to recruit kinases
to the sites of damage. Once recruited, protein-protein
interactions and post-translational modifications fully activate
the kinases to begin a cascade of phosphorylation events (35,36).
Role of c-Abl in the DDR
The c-Abl tyrosine kinase is activated by DNA
damaging agents (ionizing radiation), alkylating agents (mitomycin
C) and DNA cross-linking agents (cisplatin); these triggering
factors initiate a strong c-Abl response to DNA damage signaling
(1). The
ataxia-telangiectasia-mutated (ATM) protein is an upstream effector
of c-Abl and binds with the c-Abl SH3 domain (1). c-Abl is phosphorylated by ATM at the
serine site of 465 following ionizing radiation in cells, which
results in c-Abl activation. Therefore, in response to DNA damage,
ATM serves a crucial role in c-Abl activation (1,18).
DNA transaction processes (DNA replication, recombination and
repair) serve a basic role in the stability of the genome and
diversity. It has been discovered that c-Abl is involved in these
processes through its DNA binding domain (17). A number of auxiliary proteins,
enzymes and effectors participating in DNA transactions have been
observed to bind with, and be modified, by the c-Abl tyrosine
kinase (17). In fact, c-Abl has
been discovered to bind with and phosphorylate several DDR proteins
(1,37,38);
therefore, it is suggested that the DDR is regulated by c-Abl. In
addition, the tyrosine kinase c-Abl may act as a decision maker in
the cellular environment due to its role in the activation of the
proapoptotic signaling pathway in circumstances when DNA damage
repair is not possible (1,37,38).
Cellular functions regulated by the
c-Abl-mediated tyrosine phosphorylation events of DNA damage
response proteins
c-Abl tyrosine kinase is an unique kinase activated
by genotoxic stress in cells, which directly phosphorylates various
major cellular proteins, including DDR proteins to regulate
important cellular functions in the cell (Fig. 1) (39,40).
Here in, these DDR proteins are categorized according to their
functional roles.
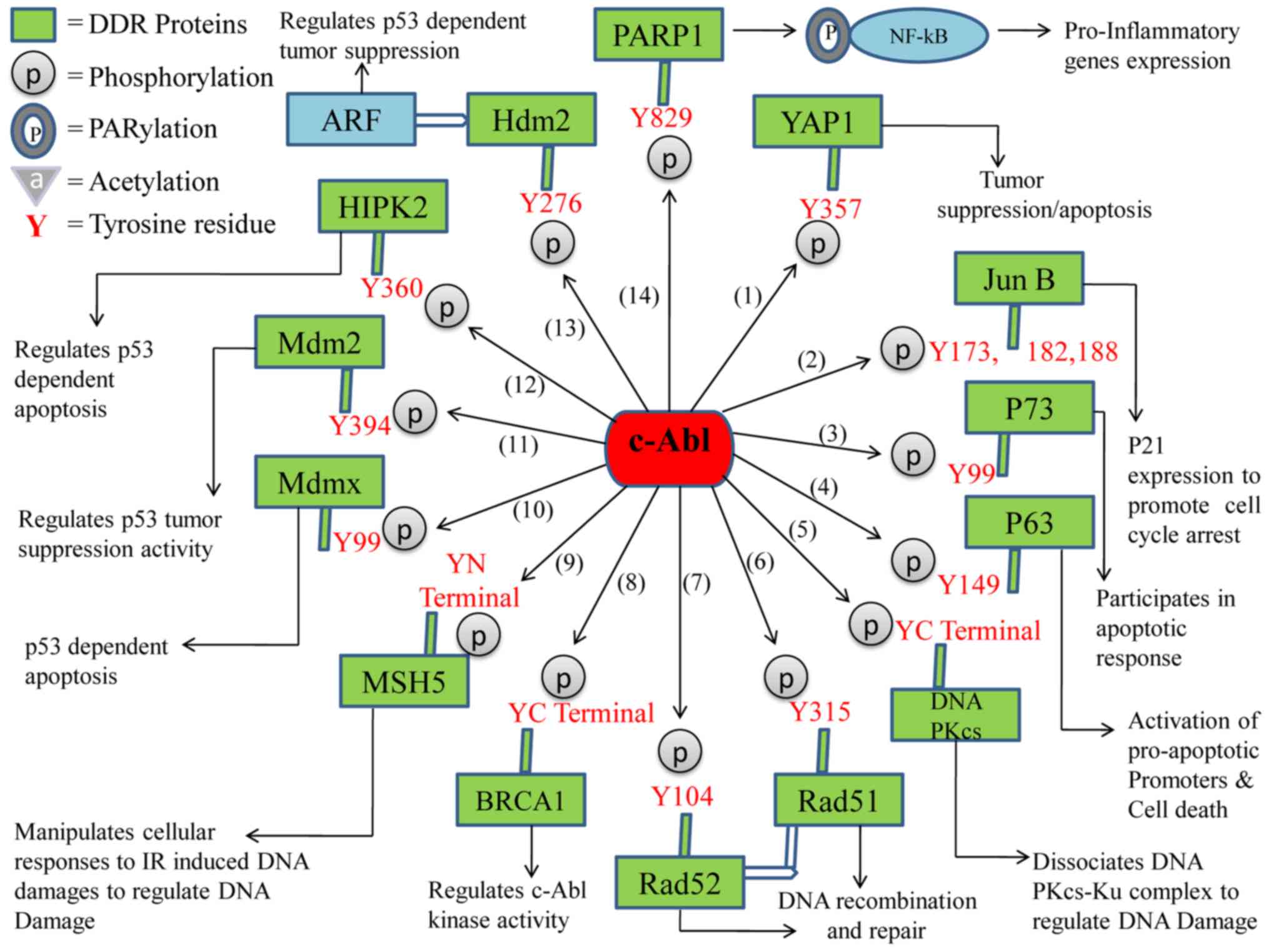 | Figure 1.Schematic diagram of the
c-ABL-mediated phosphotyrosine events of DDR proteins and the
functional outcomes. (1) The
phosphorylation of YAP1 on Y357 inactivates the oncogenic function
of YAP1, and regulates tumor suppression and apoptosis. (2) Phosphorylated Jun B at Y173, 182,188
initiates the expression of p21 to promote cell cycle arrest.
(3) P73 phosphorylation on Y99
increases apoptosis. (4) p63
phosphorylation on Y149 activates proapoptotic promoters and
regulates cell death. (5) Tyrosine
phosphorylation of DNA-PKcs at the C-terminal region results in the
dissociation of the DNA-PKcs-Ku complex and the regulation of DNA
damage. (6) Phosphorylated RAD51
on Y315 promotes the formation of the complex between RAD51 and
RAD52 protein, which cooperates with RAD51 in DNA recombination and
repair. (7) Phosphorylated RAD52
on Y104 regulates DNA recombination and repair. (8) BRCA1 tyrosine phosphorylation at the
C-terminal region regulates c-ABL kinase activity. (9) Phosphorylated MSH5 at the N-terminal
region manipulates cellular responses to IR-induced DNA damages to
regulate DNA damage. (10) MDMX
phosphorylation at Y99 regulates p53 dependent apoptosis. (11) MDM2 phosphorylation at Y394
regulates p53 tumor suppressor activity. (12) HIPK2 phosphorylation at Y360
regulates p53 dependent apoptosis. (13) Phosphorylation of HDM2 at Y276
regulates the interaction with ARF that contributes functionally to
the induction of p53 and tumor suppression. (14) Phosphorylated PARP1 at the Y829 site
results in PARP1/NF-κB signaling pathway dependent proinflammatory
gene expression and inflammation. DDR, DNA damage response; YAP1,
yes-associated protein 1; DNA-PKcs, DNA-dependent protein kinase,
catalytic subunit; BRCA1, breast cancer type 1 susceptibility
protein; MSH5, mut S protein homology 5; MDM2, mouse double minute
2; HIPK2, homeodomain-interacting protein kinase 2; HDM2, human
homolog of MDM2; alternate reading frame protein product of the
INK4a/Arf locus, p14ARF; PARP1, poly (ADP-ribose)
polymerase 1; IR, ionizing radiation. |
c-Abl targets proteins involved in DNA
recombination, repair and survival
RAD51
RAD51 is a DNA recombination and repair protein,
which is a homologue of the bacterial RecA protein. The
c-Abl-dependent tyrosine phosphorylation of RAD51 is dependent on
ATM, which is activated following ionizing radiation. c-Abl, ATM
and RAD51 can be co-immunoprecipitated from cell extracts,
suggesting the ATM/c-Abl signaling pathway is responsible for the
correct post-translational modification of RAD51, which is
necessary for the assembly of the RAD51 repair protein complex
following ionizing radiation. Specifically, DNA damage-activated
c-Abl tyrosine kinase phosphorylates RAD51 on tyrosine site 315 and
this phosphorylation induces the formation of the RAD51/RAD52
protein complex, which assists RAD51 in DNA recombination and
repair (41,42).
RAD52
The RAD52 protein plays an important role in the
homologous recombination repair of double-strand breaks in
collaboration with RAD51 and RAD54, which are all known as the
epistasis group of proteins. A well-known feature linked with the
ability of these proteins to repair double-strand breaks is the
formation of inducible nuclear foci at damaged sites. c-Abl
tyrosine kinase has been discovered to bind with, and phosphorylate
RAD52 on tyrosine site 104 in cells following ionizing radiation
(43).
c-Abl targets proteins involved in tumor
suppression and apoptosis
Mouse double minute 2 (MDM2)
MDM2 has been established to be a major negative
regulator of the tumor suppressor protein p53, which promotes tumor
formation (43,44). In response to DNA damage, c-Abl
interacts with and phosphorylates MDM2 at the tyrosine site Y394;
the c-Abl-dependent tyrosine phosphorylation of MDM2 neutralizes
the MDM2-induced inhibitory effects over p53. In fact, the
phosphorylation of MDM2 by c-Abl has been found to regulate the p53
tumor suppressive activity both in vitro and in vivo
(45,46).
Human homologue of MDM2 (HDM2)
HDM2 is a major negative regulator of the tumor
suppressor protein, p53, which can promote tumor formation
(47). In response to DNA damage,
c-Abl interacts with HDM2 and induces its phosphorylation at
tyrosine site Y276. The c-Abl-dependent tyrosine phosphorylation of
HDM2 subsequently regulates its interaction with an alternate
reading frame protein product of the INK4a/Arf locus, known as
p14ARF, which results in an increased level of nucleolar
HDM2 and a low turnover of p53. Thus, indicating that the
c-Abl-induced phosphorylation of HDM2 tyrosine site Y276 is
involved in the induction of p53 and regulation of p53-dependent
tumor suppression (48).
MDMX (MDM4)
MDMX is a homologue of the E3 ubiquitin ligase MDM2
protein. MDMX has been discovered to negatively regulate the tumor
suppressor protein p53 in normal cells, through both MDM2-dependent
and -independent mechanisms. c-Abl tyrosine kinase phosphorylates
MDMX at tyrosine site 99 following DNA damage, which subsequently
disrupts the binding of MDMX to p53 and as a result, p53 escapes
MDMX- and MDM2-dependent functional inhibition and degradation.
Therefore, the phosphorylation of MDMX by c-Abl protects damaged
cells from proliferation through the induction of cell cycle
arrest, DNA repair, senescence or programmed cell death (49,50).
Homeodomain-interacting protein kinase
2 (HIPK2)
HIPK2 is an important regulator of p53-dependent
programmed cell death. Several sites of HIPK2 have been discovered
to be phosphorylated by c-Abl, which protects HIPK2 from ubiquitin
E3 ligase Siah-1-mediated degradation. The tyrosine phosphorylation
of HIPK2 at tyrosine residue 360 (Y360) by c-Abl occurs following
DNA damage, which regulates p53-dependent programmed cell death
(4).
Tumor protein 63 (TP63)/p63
p63 or TP63 belongs to the p53 family of
tumor-suppressor proteins (51,52).
c-Abl has been discovered to phosphorylate p63 on the Y149 tyrosine
residue following cisplatin treatment in vitro. The
subsequent modification has been revealed to affect p63 stability
and induce the p63-dependent activation of proapoptotic promoters.
In fact, the activation of the c-Abl/p63 signaling pathway by
chemotherapeutic DNA-damaging drugs has been observed in model
human cell lines and mouse oocytes, where it plays an important
role in apoptosis (53).
p73
The p73 protein belongs to the p53 family of
tumor-suppressor proteins and it can also induce programmed cell
death. c-Abl was revealed to interact with the carboxy-terminal
homo-oligomerization domain of the p73 protein through the SH3
domain. Following ionizing radiation, p73 is phosphorylated by the
c-Abl tyrosine kinase on the tyrosine site of 99 both in vitro
and in vivo. Importantly, the p73-mediated transactivation and
apoptosis have both been found to be regulated by c-Abl. In
response to DNA damage, the c-Abl-mediated regulation of p73 has
also been observed with the failure of ionizing radiation-induced
apoptosis following the interruption of the c-Abl/p73 interaction.
Thus, a c-Abl-dependent mechanism regulating both p53 and p73 is
suggested to be involved in the apoptotic response to DNA damage
(54).
YES-associated protein 1 (YAP1)
YAP1 has been discovered to function as an oncogene
in a number of experimental systems, as well as functioning as a
central transcription co-activator. The transcriptional enhancer
activator domain (TEAD) family of transcription factors are known
to be co-activated by YAP. The tyrosine kinase c-Abl and DNA damage
has been discovered to serve as negative regulators of TEAD
co-activation through YAP; c-Abl tyrosine kinase phosphorylates
YAP1 at the Y357 residue under conditions of DNA damage. This
phosphorylation event results in the activation of proapoptotic
genes in relation to p73 and inactivates the oncogenic function of
YAP1 (55).
c-Abl targets proteins involved in cell
cycle arrest
Jun B
Jun B belongs to the activator protein 1 (AP1)
transcription factor family and it has since been discovered to be
associated with the DDR. c-Abl was found to phosphorylate Jun B at
the tyrosine sites of 173, 182 and 188, which inhibits its
suppressive role in p21 promoter activity, most likely through
blocking the formation of the Jun B/c-Fos AP1 complex. c-Abl
targets Jun B, which is required for Adriamycin-induced expression
of p21 (56). In response to a
variety of stimuli, p21 has been found to promote cell cycle arrest
(57).
c-Abl targets proteins involved in DNA
damage regulation
DNA-dependent protein kinase, catalytic subunit
(DNA-PKcs)
DNA-PKcs is the 470 kDa catalytic subunit of
DNA-dependent protein kinase (DNA-PK). In mammalian cells, the
double-stranded DNA break repair mechanism is controlled by
DNA-PKcs. The structure of DNA-PKcs is similar to multiple other
proteins involved in radiation-induced checkpoint responses.
DNA-PKcs target DNA breaks together with the DNA binding
heterodimer, Ku (58). It was
reported that the C-terminal fragments of DNA-PKcs were
phosphorylated by c-Abl, specifically the amino acids 3,414 to
3,850. The c-Abl-mediated phosphorylation of DNA-PKcs was
discovered to subsequently disrupt the DNA-PKcs and Ku complex.
Therefore, c-Abl and Ku may perform opposing functions with
relation to regulating DNA-PK activity (58–60).
MutS protein homolog 5 (MSH5)
MSH5 belongs to the DNA mismatch repair family of
proteins. Human MSH5 and c-Abl are associated by the direct
physical interaction between the NH2 terminus (residues 1–109) of
human MSH5 (hMSH5) and the c-Abl SH3 domain. The activation of
c-Abl tyrosine kinase and the phosphorylation of hMSH5 in response
to ionizing radiation were found to be dependent upon this physical
interaction. This interaction of hMSH5 with c-Abl also implied that
the crosstalk between hMSH5 and c-Abl may manipulate the cellular
responses to ionizing radiation-induced DNA damage (61).
c-Abl kinase activation
Breast cancer type 1 susceptibility
protein (BRCA1)
BRCA1 is expressed in breast cells, as well as in
other tissues (62). BRCA1
supports DNA repair; however, it also destroys cells in
circumstances when DNA repair is not possible (62). c-Abl was discovered to bind to the
C-terminal region of BRCA1 (residues 1,314 to 1,863) and
phosphorylate the C-terminus in vitro. In addition, BRCA1
was also found to be phosphorylated at tyrosine residues in an ATM-
and radiation-dependent manner. The tyrosine phosphorylation of
BRCA1 is not required to disrupt the complex between BRCA1/c-Abl;
the complex co-binds essentially; however, the exposure to ionizing
radiation triggers an ATM-dependent disruption of the BRCA1/c-Abl
complex, which coincides with the activation of c-Abl kinase
activity. In fact, in one previous study, c-Abl kinase activity was
found to be constitutively elevated following the loss of BRCA1
(63).
c-Abl targets proteins involved in the
immune response
Poly (ADP-ribose) polymerase 1
(PARP1)
PARP1 is a crucial DDR protein (64,65).
PARP1 has been observed to be activated by DNA damage, including
strand breaks and apurinic/apyrimidinic sites, and is thus
physiologically associated with DNA damage detection, DNA repair
and cell death (66–68). According to a recent study by our
group, upon immunological challenges, c-Abl was found to undergo
nuclear translocation, and bind with and phosphorylate PARP1 at the
conserved tyrosine site, Y829. The c-ABL-mediated tyrosine
phosphorylation of PARP1 resulted in the activation of a
PARP1/NF-κB signaling pathway-dependent immune response by inducing
proinflammatory gene expression (69).
Conclusion
In conclusion, c-Abl has been discovered to interact
with DDR proteins to dictate cell fate/DNA repair, transcriptional
regulation, tumor suppression, cell death or cell cycle arrest
following DNA damage. The present review presented an overview of
all DDR proteins phosphorylated by c-Abl tyrosine kinase and the
functional outcomes of these phosphotyrosine events. c-Abl tyrosine
kinase was found to phosphorylate DDR proteins on different
tyrosine sites and these phosphotyrosine events of DDR proteins
were reported to be crucial for the activation of other cell
signaling pathways to regulate important cellular functions.
Altogether, this review provided evidence to suggest that the
pharmacological inhibition of c-Abl may be of clinical use in
certain physiopathological contexts.
Acknowledgements
The authors would like to thank Dr. Istvan Boldogh
(Department of Microbiology and Immunology, University of Texas
Medical Branch at Galveston) for his intellectual input in this
review.
Funding
The present work was supported by grants from the
National Natural Science Foundation of China (grant nos. 31970686
and 31571339 to XB), the Program for Introducing Talent to
Universities (grant no. B07017 to XB) and the Natural Science
Foundation of Jilin, China (grant no. 20180101236JC to XB).
Availability of data and materials
Not applicable.
Authors' contributions
XB and XZ decided on the topics discussed in the
review; RW and AAB obtained the relevant literature; and AAB wrote
the article, which was reviewed by XB. All authors read and
approved the final manuscript and agree to be accountable for the
accuracy and referencing of the information included in the
review.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Shaul Y and Ben-Yehoyada M: Role of c-Abl
in the DNA damage stress response. Cell Res. 15:33–35. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maiani E, Diederich M and Gonfloni S: DNA
damage response: The emerging role of c-Abl as a regulatory switch?
Biochem Pharmacol. 82:1269–1276. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tu CC, Zhong Y, Nguyen L, Tsai A, Sridevi
P, Tarn WY and Wang JY: The kinase ABL phosphorylates the
microprocessor subunit DGCR8 to stimulate primary microRNA
processing in response to DNA damage. Sci Signal. 8:ra642015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Reuven N, Adler J, Porat Z, Polonio-Vallon
T, Hofmann TG and Shaul Y: The tyrosine kinase c-Abl promotes
homeodomain-interacting protein kinase 2 (HIPK2) accumulation and
activation in response to DNA damage. J Biol Chem. 290:16478–16488.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Staquicini FI, Qian MD, Salameh A, Dobroff
AS, Edwards JK, Cimino DF, Moeller BJ, Kelly P, Nunez MI, Tang X,
et al: Receptor tyrosine kinase EphA5 is a functional molecular
target in human lung cancer. J Biol Chem. 290:7345–7359. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen E, Ahn JS, Massie CE, Clynes D,
Godfrey AL, Li J, Park HJ, Nangalia J, Silber Y, Mullally A, et al:
JAK2V617F promotes replication fork stalling with
disease-restricted impairment of the intra-S checkpoint response.
Proc Natl Acad Sci USA. 111:15190–15195. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Reuven N, Adler J, Meltser V and Shaul Y:
The Hippo pathway kinase Lats2 prevents DNA damage-induced
apoptosis through inhibition of the tyrosine kinase c-Abl. Cell
Death Differ. 20:1330–1340. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fukumoto Y, Morii M, Miura T, Kubota S,
Ishibashi K, Honda T, Okamoto A and Yamaguchi N, Iwama A, Nakayama
Y and Yamaguchi N: Src family kinases promote silencing of
ATR-Chk1signaling in termination of DNA damage checkpoint. J Bol
Chem. 289:12313–12329. 2014. View Article : Google Scholar
|
|
9
|
Mahajan K, Coppola D, Rawal B, Chen YA,
Lawrence HR, Engelman RW, Lawrence NJ and Mahajan NP: Ack1-mediated
androgen receptor phosphorylation modulates radiation resistance in
castration-resistant prostate cancer. J Biol Chem. 287:22112–22122.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kharbanda S, Saleem A, Yuan ZM, Kraeft S,
Weichselbaum R, Chen LB and Kufe D: Nuclear signaling induced by
ionizing radiation involves colocalization of the activated
p56/p53lyn tyrosine kinase with p34cdc2. Cancer Res. 56:3617–3621.
1996.PubMed/NCBI
|
|
11
|
Srinivasan D and Plattner R: Activation of
Abl tyrosine kinases promotes invasion of aggressive breast cancer
cells. Cancer Res. 66:5648–5655. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hunter T: The genesis of tyrosine
phosphorylation. Cold Spring Harb Perspect Biol. 6:a0206442014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pawson T: Specificity in signal
transduction: From phosphotyrosine-SH2 domain interactions to
complex cellular systems. Cell. 116:191–203. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hunter T: Tyrosine phosphorylation: Thirty
years and counting. Curr Opin Cell Biol. 21:140–146. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lind SB, Artemenko KA and Pettersson U: A
strategy for identification of protein tyrosine phosphorylation.
Methods. 56:275–283. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hantschel O and Superti-Furga G:
Regulation of the c-Abl and Bcr-Abl tyrosine kinases. Nat Rev Mol
Cell Biol. 5:33–44. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shaul Y: c-Abl: Activation and nuclear
targets. Cell Death Differ. 7:10–16. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shiloh Y: ATM and related protein kinases:
Safeguarding genome integrity. Nat Rev Cancer. 3:155–168. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lindholm D, Pham DD, Cascone A, Eriksson
O, Wennerberg K and Saarma M: c-Abl inhibitors enable insights into
the pathophysiology and neuroprotection in parkinson's disease.
Front Aging Neurosci. 8:2542016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu ZG, Baskaran R, Lea-Chou ET, Wood LD,
Chen Y, Karin M and Wang JY: Three distinct signalling responses by
murine fibroblasts to genotoxic stress. Nature. 384:273–276. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kharbanda S, Ren R, Pandey P, Shafman TD,
Feller SM, Weichselbaum RR and Kufe DW: Activation of the c-Abl
tyrosine kinase in the stress response to DNA-damaging agents.
Nature. 376:785–788. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Colicelli J: ABL tyrosine kinases:
Evolution of function, regulation, and specificity. Sci Signal.
3:re62010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zuckerman V, Lenos K, Popowicz GM,
Silberman I, Grossman T, Marine JC, Holak TA, Jochemsen AG and
Haupt Y: c-Abl phosphorylates Hdmx and regulates its interaction
with p53. J Biol Chem. 284:4031–4039. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gu JJ, Ryu JR and Pendergast AM: Abl
tyrosine kinases in T-cell signaling. Immunol Rev. 228:170–183.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sirvent A, Benistant C and Roche S:
Cytoplasmic signalling by the c-Abl tyrosine kinase in normal and
cancer cells. Biol Cell. 100:617–631. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhu J and Wang JY: Death by Abl: A matter
of location. Curr Top Dev Biol. 59:165–192. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pendergast AM: The Abl family kinases:
Mechanisms of regulation and signaling. Adv Cancer Res. 85:51–100.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gonfloni S: DNA damage stress response in
germ cells: Role of c-Abl and clinical implications. Oncogene.
29:6193–6202. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Curtin NJ: DNA repair dysregulation from
cancer driver to therapeutic target. Nat Rev Cancer. 12:801–817.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sirbu BM and Cortez D: DNA damage
response: Three levels of DNA repair regulation. Cold Spring Harb
Perspect Biol. 5:a0127242013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ba X and Boldogh I: 8-Oxoguanine DNA
glycosylase 1: Beyond repair of the oxidatively modified base
lesions. Redox Biol. 14:669–678. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Arai H, Wada R, Ishino K, Kudo M, Uchida E
and Naito Z: Expression of DNA damage response proteins in gastric
cancer: Comprehensive protein profiling and histological analysis.
Int J Oncol. 52:978–988. 2018.PubMed/NCBI
|
|
33
|
Han YG, Yun M, Choi M, Lee SG and Kim H:
TRAIP regulates Histone H2B monoubiquitination in DNA damage
response pathways. Oncol Rep. 41:3305–3312. 2019.PubMed/NCBI
|
|
34
|
Lovejoy CA and Cortez D: Common mechanisms
of PIKK regulation. DNA Repair (Amst). 8:1004–1008. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Weterings E and Chen DJ: DNA-dependent
protein kinase in nonhomologous end joining: A lock with multiple
keys? J Cell Biol. 179:183–186. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dobbs TA, Tainer JA and Lees-Miller SP: A
structural model for regulation of NHEJ by DNA-PKcs
autophosphorylation. DNA Repair (Amst). 9:1307–1314. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kharbanda S, Yuan ZM, Weichselbaum R and
Kufe D: Functional role for the c-Abl protein tyrosine kinase in
the cellular response to genotoxic stress. Biochim Biophys Acta.
1333:O1–O7. 1997.PubMed/NCBI
|
|
38
|
Wang JY: Controlling Abl: Auto-inhibition
and co-inhibition? Nat Cell Biol. 6:3–7. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tang J, Wang JY and Parker LL: Detection
of early Abl kinase activation after ionizing radiation by using a
peptide biosensor. Chembiochem. 13:665–673. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Meltser V, Ben-Yehoyada M and Shaul Y:
c-Abl tyrosine kinase in the DNA damage response: Cell death and
more. Cell Death Differ. 18:2–4. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen G, Yuan SS, Liu W, Xu Y, Trujillo K,
Song B, Cong F, Goff SP, Wu Y, Arlinghaus R, et al:
Radiation-induced assembly of Rad51 and Rad52 recombination complex
requires ATM and c-Abl. J Biol Chem. 274:12748–12752. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Shimizu H, Popova M, Fleury F, Kobayashi
M, Hayashi N, Sakane I, Kurumizaka H, Venkitaraman AR, Takahashi M
and Yamamoto K: c-ABL tyrosine kinase stabilizes RAD51 chromatin
association. Biochem Biophys Res Commun. 382:286–291. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kitao H and Yuan ZM: Regulation of
ionizing radiation-induced Rad52 nuclear foci formation by
c-Abl-mediated phosphorylation. J Biol Chem. 277:48944–48948. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Oliner JD, Kinzler KW, Meltzer PS, George
DL and Vogelstein B: Amplification of a gene encoding a
p53-associated protein in human sarcomas. Nature. 358:80–83. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Goldberg Z, Vogt Sionov R, Berger M, Zwang
Y, Perets R, Van Etten RA, Oren M, Taya Y and Haupt Y: Tyrosine
phosphorylation of Mdm2 by c-Abl: Implications for p53 regulation.
EMBO J. 21:3715–3727. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Carr MI, Roderick JE, Zhang H, Woda BA,
Kelliher MA and Jones SN: Phosphorylation of the Mdm2 oncoprotein
by the c-Abl tyrosine kinase regulates p53 tumor suppression and
the radiosensitivity of mice. Proc Natl Acad Sci USA.
113:15024–15029. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wade M, Wong ET, Tang M, Stommel JM and
Wahl GM: Hdmx modulates the outcome of p53 activation in human
tumor cells. J Biol Chem. 281:33036–33044. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Dias SS, Milne DM and Meek DW: c-Abl
phosphorylates Hdm2 at tyrosine 276 in response to DNA damage and
regulates interaction with ARF. Oncogene. 25:6666–6671. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chen X, Gohain N, Zhan C, Lu WY, Pazgier M
and Lu W: Structural basis of how stress-induced MDMX
phosphorylation activates p53. Oncogene. 35:1919–1925. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Xiong S, Van Pelt CS, Elizondo-Fraire AC,
Liu G and Lozano G: Synergistic roles of Mdm2 and Mdm4 for p53
inhibition in central nervous system development. Proc Natl Acad
Sci USA. 103:3226–3231. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yang A, Kaghad M, Wang Y, Gillett E,
Fleming MD, Dötsch V, Andrews NC, Caput D and McKeon F: p63, a p53
homolog at 3q27-29, encodes multiple products with transactivating,
death-inducing, and dominant-negative activities. Mol Cell.
2:305–316. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wu G, Nomoto S, Hoque MO, Dracheva T,
Osada M, Lee CC, Dong SM, Guo Z, Benoit N, Cohen Y, et al:
DeltaNp63alpha and TAp63alpha regulate transcription of genes with
distinct biological functions in cancer and development. Cancer
Res. 63:2351–2357. 2003.PubMed/NCBI
|
|
53
|
Gonfloni S, Di Tella L, Caldarola S,
Cannata SM, Klinger FG, Di Bartolomeo C, Mattei M, Candi E, De
Felici M, Melino G and Cesareni G: Inhibition of the c-Abl-TAp63
pathway protects mouse oocytes from chemotherapy-induced death. Nat
Med. 15:1179–1185. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yuan ZM, Shioya H, Ishiko T, Sun X, Gu J,
Huang YY, Lu H, Kharbanda S, Weichselbaum R and Kufe D: p73 is
regulated by tyrosine kinase c-Abl in the apoptotic response to DNA
damage. Nature. 399:814–817. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
55
|
Keshet R, Adler J, Ricardo Lax I, Shanzer
M, Porat Z, Reuven N and Shaul Y: c-Abl antagonizes the YAP
oncogenic function. Cell Death Differ. 22:935–945. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yamaguchi N, Yuki R, Kubota S, Aoyama K,
Kuga T, Hashimoto Y, Tomonaga T and Yamaguchi N: c-Abl-mediated
tyrosine phosphorylation of JunB is required for adriamycin-induced
expression of p21. Biochem J. 471:67–77. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Karimian A, Ahmadi Y and Yousefi B:
Multiple functions of p21 in cell cycle, apoptosis and
transcriptional regulation after DNA damage. DNA Repair (Amst).
42:63–71. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Jin S, Kharbanda S, Mayer B, Kufe D and
Weaver DT: Binding of Ku and c-Abl at the kinase homology region of
DNA-dependent protein kinase catalytic subunit. J Biol Chem.
272:24763–24766. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Hartley KO, Gell D, Smith GC, Zhang H,
Divecha N, Connelly MA, Admon A, Lees-Miller SP, Anderson CW and
Jackson SP: DNA-dependent protein kinase catalytic subunit: A
relative of phosphatidylinositol 3-kinase and the ataxia
telangiectasia gene product. Cell. 82:849–856. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Keith CT and Schreiber SL: PIK-related
kinases: DNA repair, recombination, and cell cycle checkpoints.
Science. 270:50–51. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Yi W, Lee TH, Tompkins JD, Zhu F, Wu X and
Her C: Physical and functional interaction between hMSH5 and c-Abl.
Cancer Res. 66:151–158. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Friedenson B: The BRCA1/2 pathway prevents
hematologic cancers in addition to breast and ovarian cancers. BMC
Cancer. 7:1522007. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Foray N, Marot D, Randrianarison V,
Venezia ND, Picard D, Perricaudet M, Favaudon V and Jeggo P:
Constitutive association of BRCA1 and c-Abl and its ATM-dependent
disruption after irradiation. Mol Cell Biol. 22:4020–4032. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Schreiber V, Dantzer F, Ame JC and de
Murcia G: Poly(ADP-ribose): Novel functions for an old molecule.
Nat Rev Mol Cell Biol. 7:517–528. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Erdélyi K, Bakondi E, Gergely P, Szabó C
and Virág L: Pathophysiologic role of oxidative stress-induced
poly(ADP-ribose) polymerase-1 activation: Focus on cell death and
transcriptional regulation. Cell Mol Life Sci. 62:751–759. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Malanga M and Althaus FR: The role of
poly(ADP-ribose) in the DNA damage signaling network. Biochem Cell
Biol. 83:354–364. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Haince JF, McDonald D, Rodrigue A, Déry U,
Masson JY, Hendzel MJ and Poirier GG: PARP1-dependent kinetics of
recruitment of MRE11 and NBS1 proteins to multiple DNA damage
sites. J Biol Chem. 283:1197–1208. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Bonicalzi ME, Haince JF, Droit A and
Poirier GG: Regulation of poly(ADP-ribose) metabolism by
poly(ADP-ribose) glycohydrolase: Where and when? Cell Mol Life Sci.
62:739–750. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Bohio AA, Sattout A, Wang R, Wang K, Sah
RK, Guo X, Zeng X, Ke Y, Boldogh I and Ba X: c-Abl-mediated
tyrosine phosphorylation of PARP1 is crucial for expression of
proinflammatory genes. J Immunol. 203:1521–1531. 2019. View Article : Google Scholar : PubMed/NCBI
|














