Introduction
Artemisinin is a sesquiterpene lactone that is
isolated from the Chinese plant Artemisia annua (Qinghao) (1). Artemisinin has been reported to
possess antimalarial properties and it has also been discovered to
serve roles in several types of disease, including different types
of cancer and inflammatory disorders (1–3).
Dihydroartemisinin (DHA) is a semisynthetic derivative of
artemisinin that has exhibited a broad range of biological
activities, excluding antimalarial effects (2). For example, Chen et al
(4) demonstrated that DHA
ameliorated liver damage, steatosis and hepatocyte lipoapoptosis of
alcoholic fatty liver model mice by regulating lipin-1 signaling.
Li et al (5) revealed that
DHA also inhibited the proliferation and migration of breast cancer
cell lines. DHA was also reported to serve important roles in
downregulating the prostaglandin E2 synthesis signaling cascade and
the inflammation of endothelial cells in the cardiovascular system
(6). Furthermore, Zhu and Ji
(7) suggested that DHA may have
the potential to be used as a treatment option for asthma by
regulating the microRNA-183C and interleukin-6/STAT3 signaling
pathway.
In more recent years, gut microbiota has become a
research hotspot due to its involvement in several types of
disease, including obesity, diabetes, stroke, allergic asthma,
nonalcoholic fatty liver disease/nonalcoholic steatohepatitis,
cancer, inflammation, and behavioral and physiological
abnormalities (8–10). DHA has demonstrated the potential
to be used as a treatment option for fatty liver disease,
inflammatory disorders and different types of cancer (4–6),
except for malaria; however, its underlying molecular mechanism of
action remains unknown. Chen et al (11), reported that artesunate (AS)
exhibited the potential to regulate the gut microbiota, ameliorate
inflammation and improve the conditions of the liver and intestine
following the damage caused by carbon tetrachloride, alcohol and a
high fat diet in Sprague Dawley rats, suggesting that DHA may have
similar effects on the gut microbiota. Thus, the present study
aimed to investigate the effects of DHA in the liver, kidney,
intestine and gut microbiota to identify the underlying molecular
mechanisms of DHA and its pharmacological activities.
Materials and methods
Animal studies and tissue
collection
The experimental procedures in the present study
were approved by the Committee on Laboratory Animal Care and Use of
Guangdong Pharmaceutical University (Guangzhou, China). A total of
18 male C57BL/6 mice (weight, 22.53±1.23 g; age, 7 weeks old;
provided by Hunan Lex Jingda Laboratory Animal Co., Ltd.) were
housed in a temperature-controlled environment at 25°C and 50±10%
humidity, with a 12-h light/dark cycle and free access to a
standard diet and water. The mice were randomly divided into two
groups (n=9/group): Control and DHA group. According to the results
obtained in a pre-experiment and previous studies (12–14),
the experimental group was treated with DHA (150 mg/kg; Shanghai
Aladdin Biochemical Technology Co., Ltd.) diluted in 0.5%
carboxymethyl cellulose-Na (CMC-Na; Tianjin Zhiyuan Chemical
Reagent Co., Ltd.), once daily for 7 days by oral gavage. The
control group was treated with 0.5% CMC-Na once daily for 7 days by
oral gavage. After 7 days, the whole blood was collected from the
orbit; the mice were adequately anesthetized with ether for blood
collection and the pinch reflex was monitored to ensure full
anesthesia. After 1 ml blood collection, the mice were sacrificed
by cervical dislocation. The liver, kidney and intestine tissues
were collected, and fixed in 4% paraformaldehyde overnight at 4°C
for hematoxylin and eosin (H&E) staining. The experimental
procedures were performed as previously described (15).
Serum lipid profile assays. Serum was obtained from
the blood by centrifugation (1,160 × g) at 4°C for 15 min. The
concentrations of serum triglyceride (TG), total cholesterol (TC),
high density lipoprotein-cholesterol (HDL-C), low density
lipoprotein-cholesterol (LDL-C), alanine aminotransferase (ALT) and
aspartate aminotransferase (AST) were analyzed using their
respective kits (Nanjing Jiancheng Bioengineering Institute; cat.
no. A110-1-1 for TG; cat. no. A111-1-1 for TC; cat. no. A112-1-1
for HDL-C; cat. no. A113-1-1 for LDL-C; cat. no. C009-2-1 for ALT;
cat. no. C010-2-1 for AST), according to the manufacturers
protocols. The serum glucose levels were detected using a detection
kit purchased from Shanghai Rongsheng Biopharmaceutical Co., Ltd.,
according to the manufacturers protocols, and the serum
liposaccharide (LPS) concentration was determined using a detection
kit purchased from Jiangsu Enzyme Immunity Co., Ltd., according to
the manufacturers protocols.
16S ribosomal (r)DNA gene analysis. Fecal bacterial
DNA extraction, 16S rDNA gene amplification and subsequent 16S rDNA
gene analysis were performed by Gene Denovo Biotechnology Company
(Guangzhou, China). The experimental procedures were performed as
previously described (15,16). All α diversity indexes were
calculated in QIIME (17) (version
1.9.1; http://qiime.org). Shannon rarefaction curves,
Operational taxonomic units (OTU) rarefaction curve and rank
abundance curves were plotted in R project ggplot2 package
(18) (v2.2.1).
The abundance of bacteria was calculated using the
method of Ace and Chao (19), and
the diversity of bacteria was estimated using the method of Shannon
and Simpson (20). Principal
coordinates analysis (Vegan; v2.5–4; http://cran.r-project.org/web/packages/vegan) was
performed in order to present the differences between the gut
microbial communities of different groups. The Kyoto Encyclopedia
of Genes and Genomes (KEGG) pathway analysis was performed at
http://www.kegg.jp/kegg/kegg1.html.
Canonical Correspondence Analysis (CCA) was performed using Vegan
(21), and the correlation network
analysis was performed using Psych (v1.8.4; http://cran.r-project.org/web/packages/psych/index.html).
H&E staining. Mouse tissues were fixed in 4%
paraformaldehyde overnight at 4°C, dehydrated with ascending series
of alcohol (70% alcohol for 45 min, 75% alcohol for 45 min, 80%
alcohol for 45 min, 85% alcohol for 45 min, 95% alcohol for 45 min,
pure alcohol twice for 1 h each) and embedded in paraffin.
Paraffin-embedded tissues were sliced into 4-µm sections and
stained with hematoxylin for 3 min and eosin for 20 sec, both at
room temperature (Sigma-Aldrich; Merck KGaA). The stained sections
were visualized using an automated quantitative pathology system
(Vutomated Quantitative Pathology Imaging System; PerkinElmer,
Inc.; the magnification of kidney and liver was ×40, and the
magnification of intestinal tissue was ×20).
Statistical analysis. Statistical analysis was
performed using SPSS software (v23.0; National Institutes of
Health) and the data are presented as the mean ± standard deviation
(SD). Statistical differences among the two groups were determined
using an unpaired Students t-test for equal variances or a U-Mann
Whitney test for unequal variances. Each experiment was repeated ≥3
times. P<0.05 was considered to indicate a statistically
significant difference.
Results
Effects of DHA on the serum lipid profile and
important organs. DHA was demonstrated to significantly decrease
the serum TG levels compared with the control group (Fig. 1B); however, no significant
differences were observed in the serum glucose, TC, LPS, HDL-C,
LDL-C, ALT and AST levels, or the body weight, between the DHA and
control groups (Fig. 1A, C-I).
H&E staining revealed that the liver and kidney tissues of the
DHA group were similar in pathology compared with the control group
(Fig. 1J). Furthermore, no
significant differences were observed in the pathology of the
intestine, including the duodenum, jejunum, ileum and colon,
between the two groups (Fig.
S1).
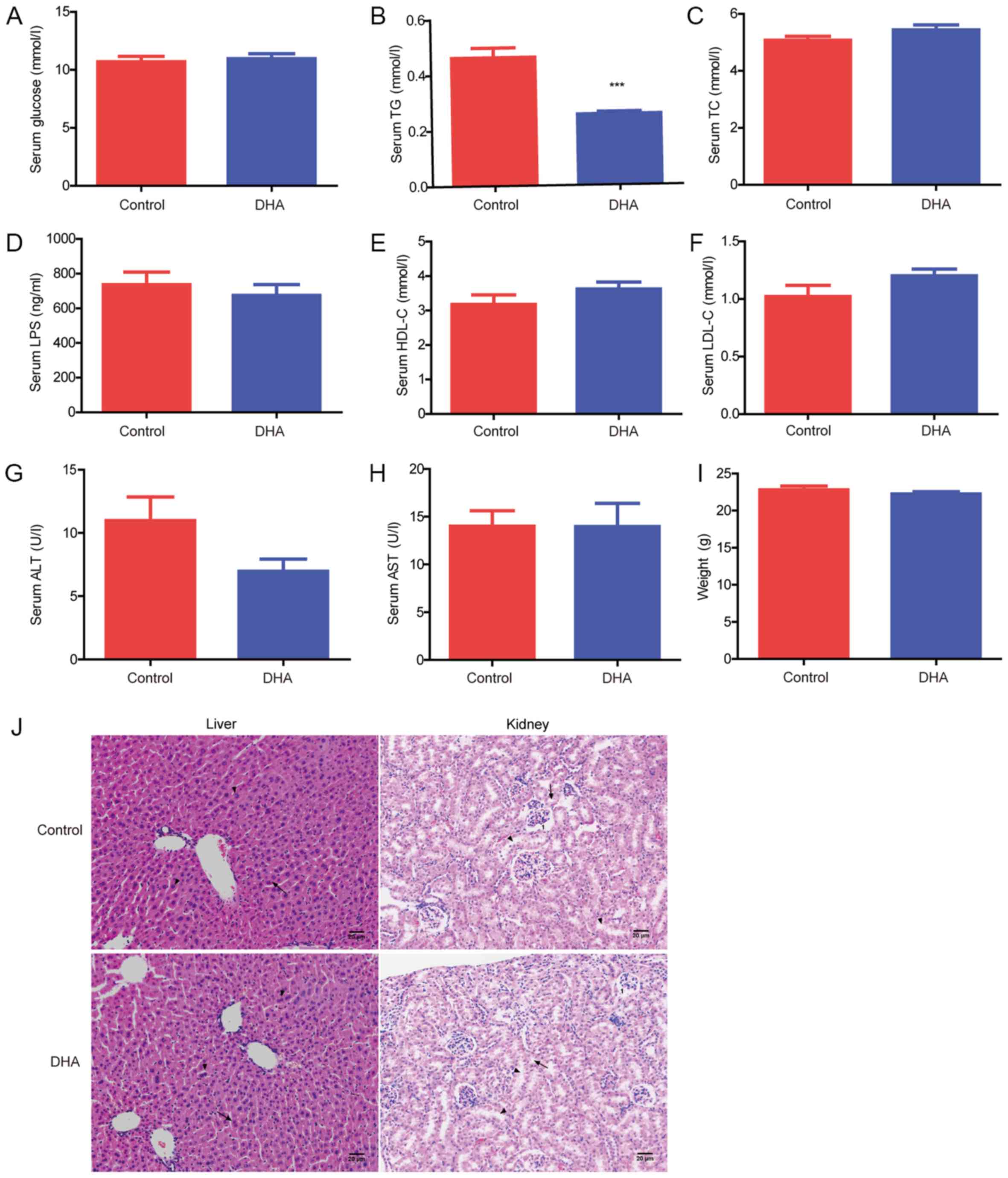 | Figure 1.Effects of DHA on the serum lipid
profile and important organs. Serum (A) glucose, (B) TG, (C) TC,
(D) LPS, (E) HDL-C, (F) LDL-C, (G) ALT and (H) AST levels were
analyzed in the DHA and control groups. (I) Body weight of the mice
in the DHA and control groups. (J) Hematoxylin and eosin staining
of the liver and kidney tissues obtained from the DHA and control
groups. Scale bar, 20 µm. In the liver: Arrows, hepatocytes;
triangles, hepatic cords. In the kidney: 1, spherical renal
corpuscles; arrows, proximal convoluted tubule; triangles, distal
convoluted tubule. ***P<0.001 vs. control. TG, triglyceride; TC,
total cholesterol; LPS, lipopolysaccharide; HDL-C, high density
lipoprotein-cholesterol; LDL-C, low density
lipoprotein-cholesterol; ALT, alanine aminotransferase; AST,
aspartate aminotransferase; DHA, dihydroartemisinin. |
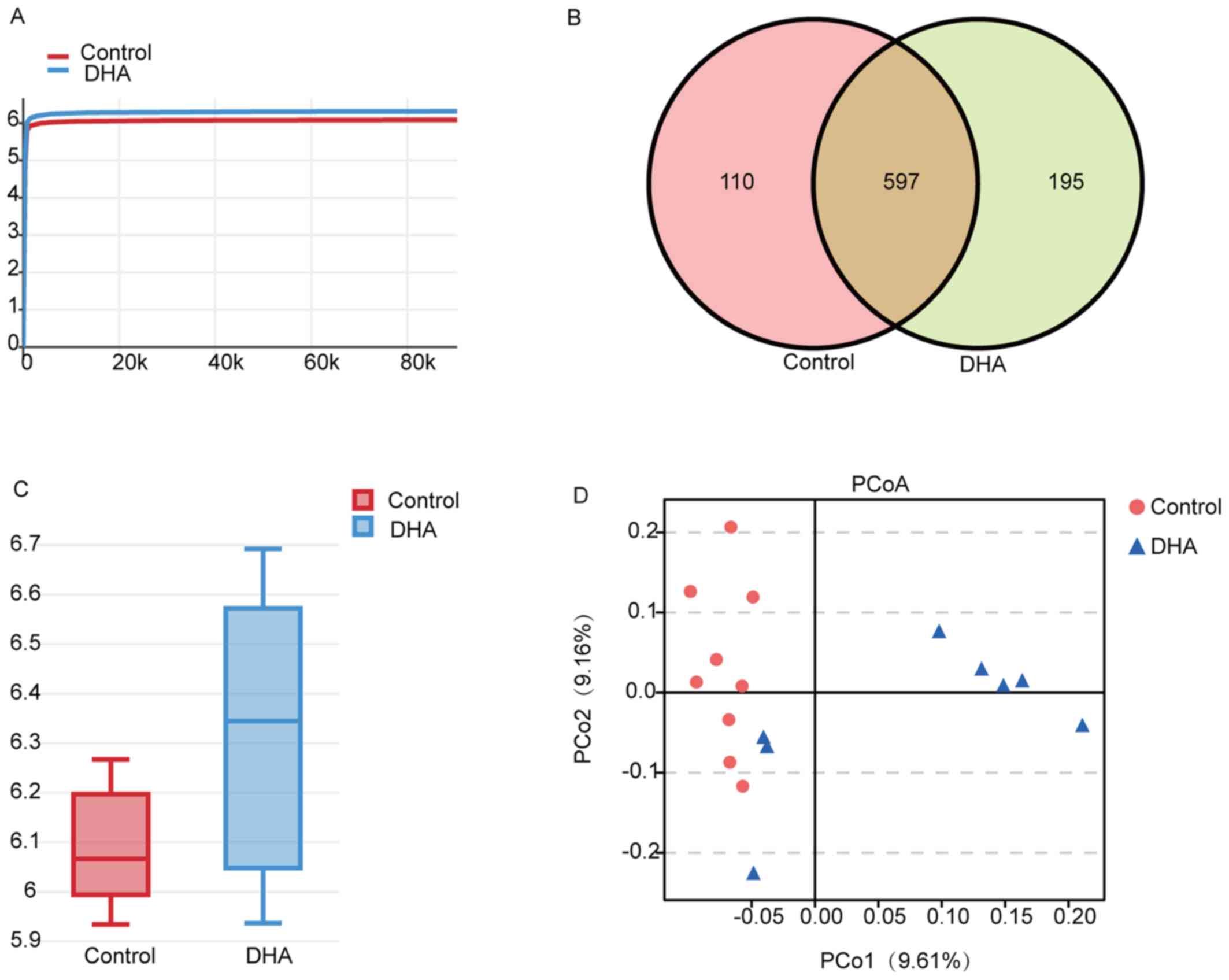
Overview of the 16S rDNA gene sequencing. In order
to determine the effect of DHA on the gut microbiota, mice fecal
samples were collected and the 16S rDNA genes were sequenced. The
indices of Shannon and Simpson (calculated for the diversity of
bacteria), Chao and Ace (calculated for the abundance of bacteria),
together with the total tags and OTUs of each group are presented
in Table I. The Shannon
rarefaction curves for each group were observed to reach the
saturation plateau, suggesting the sufficient sequence coverage of
the samples to describe the composition of the gut microbiota
(Fig. 2A). The Venn diagram
demonstrated that there were 597 common OTUs identified between the
DHA and control groups, and there were 195 and 110 OTUs specific to
the DHA-treated mice and the control group, respectively (Fig. 2B).
 | Table I.Diversity estimation of the 16S
ribosomal RNA gene library of the DHA-treated mice and the control
group. |
Table I.
Diversity estimation of the 16S
ribosomal RNA gene library of the DHA-treated mice and the control
group.
| Group | Total tags | Operational
taxonomic units | Shannon | Simpson | Chao | Ace |
|---|
| Control |
119009.11±17477.13 |
953.22±84.17 | 6.09±0.12 | 0.97±0.01 | 1661.94±104.26 | 1717.02±110.90 |
| DHA |
132965.38±10623.35 | 1010.13±51.92 | 6.32±0.27 | 0.97±0.01 | 1763.40±149.16 | 1728.54±120.93 |
Effect of DHA on the relative abundance and
composition of the gut microbiota of mice. The α diversity analysis
of the two groups demonstrated that the bacterial diversity of the
DHA group was richer compared with the control group (Fig. 2C). Furthermore, principal
coordinates analysis revealed that the DHA and the control groups
could be distinguished from each other (Fig. 2D), suggesting that DHA may alter
the gut bacteria of the mice.
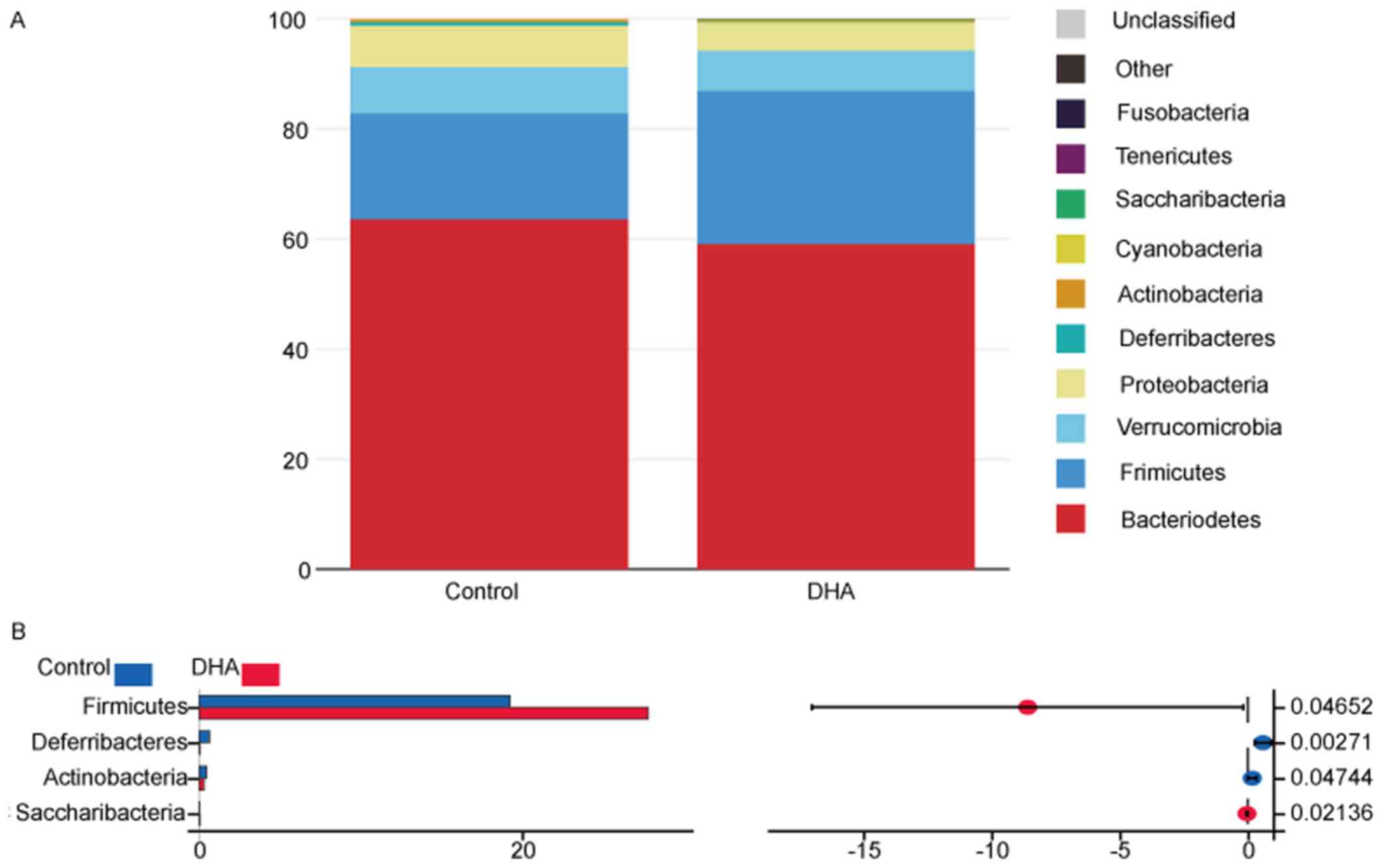
The taxonomic compositions of the bacterial phylum
of the two groups are presented in Fig. 3A and B. The majority of the samples
exhibited high percentages of Bacteroidetes, Firmicutes,
Verrucomicrobia and Proteobacteria on the phylum level,
and the DHA group had increased Firmicutes and
Saccharibacteria species, and decreased
Deferribacteres and Actinobacteria species compared
with the control group (Fig. 3A and
B; Table II). The taxonomic
compositions of the two groups on the class and family level are
presented in Fig. S2, Table SI and Table SII.
| Table II.Taxonomic composition of the gut
bacteria on the phylum level of the DHA and control groups. |
Table II.
Taxonomic composition of the gut
bacteria on the phylum level of the DHA and control groups.
| Phylum | Control (%) | DHA (%) | DHA/control | P-value |
|---|
|
Bacteroidetes | 63.61675556 | 59.1229500 | 0.92936129 | 0.262040377 |
|
Firmicutes | 19.26385556 | 27.8502625 | 1.44572629 | 0.046519675 |
|
Verrucomicrobia | 8.32234444 | 7.2688500 | 0.87341374 | 0.723052299 |
|
Proteobacteria | 7.55047778 | 5.1879000 | 0.68709559 | 0.119181387 |
|
Deferribacteres | 0.67160000 | 0.0777500 | 0.11576831 | 0.002711787 |
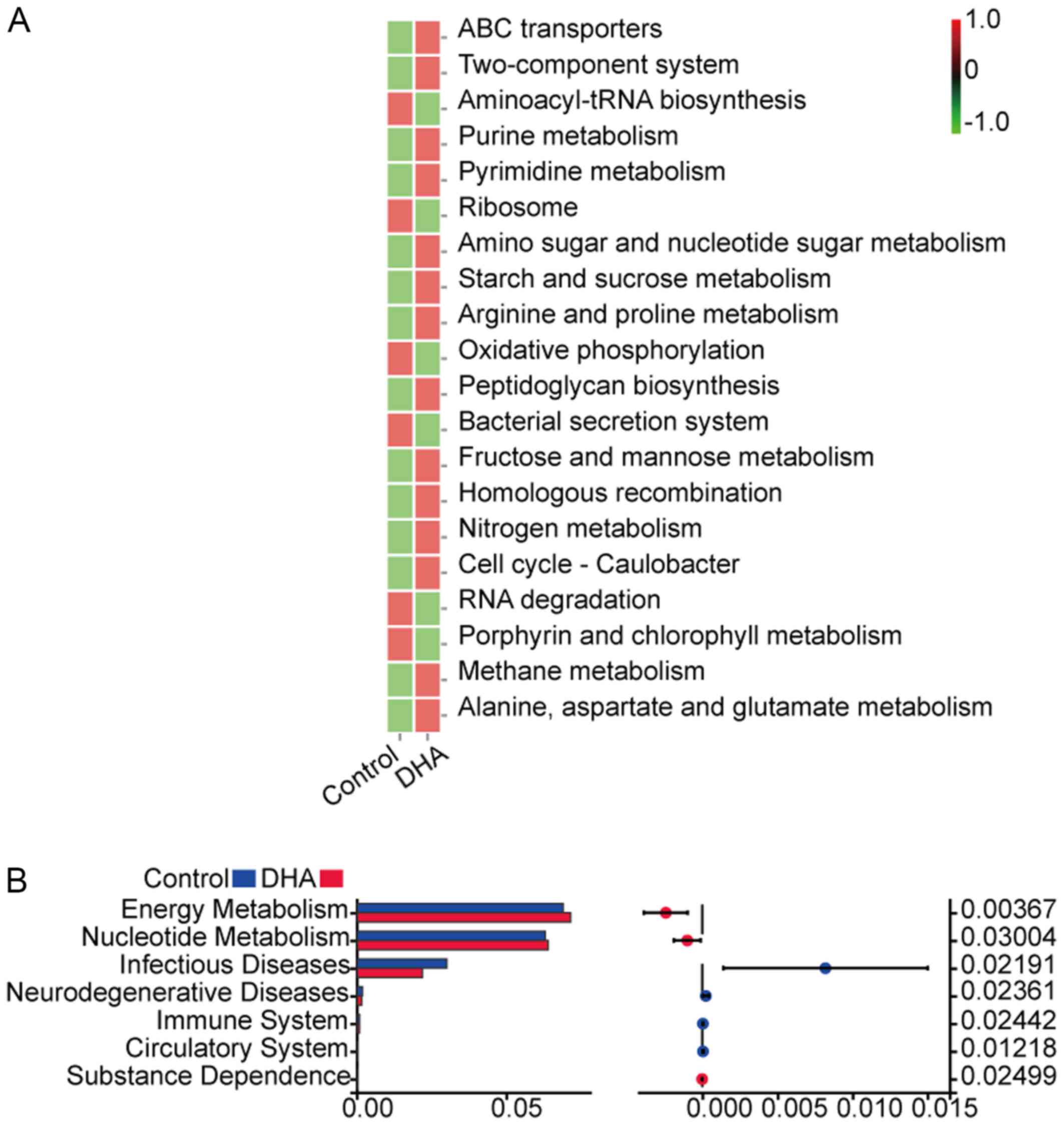
The heatmap of KEGG signaling pathway enrichment
analysis demonstrated that the following signaling pathways were
upregulated in the DHA group: ‘ABC transporters’, ‘Two-component
system’, ‘Purine metabolism’, ‘Pyrimidine metabolism’, ‘Amino sugar
and nucleotide sugar metabolism’, ‘Starch and sucrose metabolism’,
‘Arginine and proline metabolism’, ‘Peptidoglycan biosynthesis’,
‘Fructose and mannose metabolism’, ‘Homologous recombination’,
‘Nitrogen metabolism’, ‘Cell cycle-Caulobacter’, ‘Methane
metabolism’ and ‘Alanine, aspartate and glutamate metabolism’
(Fig. 4A). Conversely, the
following signaling pathways were downregulated in the DHA group
compared with the control group: ‘Aminoacyl-tRNA biosynthesis’,
‘Ribosome’, ‘Oxidative phosphorylation’, ‘Bacterial secretion
system’, ‘RNA degradation’ and ‘Porphyrin and chlorophyll
metabolism’ (Fig. 4A). The
signaling pathways associated with ‘Energy Metabolism’ and
‘Nucleotide Metabolism’ were demonstrated to be significantly
upregulated in the DHA group compared with the control group
(Fig. 4B), while the signaling
pathways associated with ‘Infectious Diseases’ and
‘Neurodegenerative Diseases’ were significantly downregulated in
the DHA group compared with the control group (Fig. 4B). The associations between HDL-C,
LDL-C, TG, TC and LPS with the bacterial taxa are presented in
Fig. S3. The result of CCA
demonstrated a correlation of the bacteria with different
environmental factors, and that LDL-C, LPS and TC influenced the
structure and composition of gut microbiota most (Fig. S3A). The result of the correlation
network analysis indicated that LDL-C had a positive correlation
with Verrucomicrobia, but had a negative correlation with
Bacteroidetes. HDL had a positive correlation with
Saccharibacteria, and TC had a positive correlation with
Fimicutes, but had a negative correlation with
Proteobacteria. Furthermore, TG had a positive correlation
with Deferribacteres, but had a negative correlation with
Saccharibacteria (Fig.
S3B).
Discussion
Artemisinin is well-known for its significant role
in the treatment of malaria (22)
and it was previously demonstrated that artemisinin had the
potential to manage ‘artemisinin-resistance’ (23). In addition to its antimalarial
properties, artemisinin and its derivatives were discovered to
exhibit a range of pharmaceutical activities, such as anti-viruses,
anti-inflammation and anti-tumors (1). DHA, an important derivative of
artemisinin, was revealed to be associated with several biological
activities, including improving liver damage, inhibiting cancer and
inflammation (4–7); however, its effect on the gut
microbiota remains poorly understood. In the present study, C57BL/6
mice were treated with DHA to determine its effects on the gut
microbiota, lipid metabolism and important organs, including the
liver, kidney and intestine. The H&E staining results of the
liver, kidney and intestinal tissues demonstrated that there were
no significant differences in the pathology between the DHA group
and the control group, indicating that DHA did not exert toxic
effects on these organs. TG serum levels were significantly
decreased in the DHA group compared with the control group;
however, there was no significant differences observed in the serum
levels of glucose, TC, LPS, HDL-C, LDL-C, ALT and AST between the
two groups. It was previously reported that AS significantly
decreased the TG plasma levels, thus these findings suggested the
potential of DHA to be used as a treatment for hyperlipidemia when
combined with ursolic acid (24,25).
The results of the present study indicated that DHA may exert
similar biological functions as AS.
The effects of DHA on the gut microbiota were
further analyzed via 16S rDNA gene analysis; the α diversity
accounts for the abundance of species residing in the host, whereby
a high α diversity indicates a healthy host pattern. For example,
Wolff et al (26) reported
that the healthy oral microbiome had a significantly higher
diversity compared with the advanced caries microbiome, whereas
Clarke et al (27) reported
that the gut microbiota of athletes was more diverse compared with
the controls. The results of the present study demonstrated that
the bacterial diversity of the DHA group was richer compared with
the control group, suggesting that DHA may have the potential to
increase the diversity of the gut bacteria.
Obese individuals have exhibited a tendency to have
an increased ratio of Firmicutes to Bacteroidetes in
the gut microbiota compared with lean humans and mice (28–30).
In the present study, DHA was demonstrated to increase the relative
abundance of the Firmicutes phylum from 19.26 to 27.85%,
while the Bacteroidetes phylum decreased from 63.62 to
59.12% compared with the control group, respectively. Although DHA
increased the ratio of Firmicutes to Bacteroidetes,
the mice treated with DHA failed to exhibit an obese phenotype as
there was no difference in body weight between the control and DHA
groups. Furthermore, there were no significant differences observed
in the body weights of the mice between the DHA and control groups.
According to Mazloom et al (31), the importance of the
Firmicutes to Bacteroidetes ratio in obesity remains
controversial as several studies have reported no significant
differences between the two phyla in obesity. The results of the
present study suggested that the effect of DHA on the
Firmicutes to Bacteroidetes ratio was not associated
with obesity. The present study also revealed that DHA
significantly decreased TG serum levels, whilst KEGG signaling
pathway enrichment analysis indicated that the signaling pathways
associated with ‘Energy Metabolism’ and ‘Nucleotide Metabolism’,
such as ‘Purine metabolism’, ‘Pyrimidine metabolism’, ‘Amino sugar
and nucleotide sugar metabolism’, ‘Starch and sucrose metabolism’,
‘Arginine and proline metabolism’, ‘Peptidoglycan biosynthesis’,
‘Fructose and mannose metabolism’, ‘Nitrogen metabolism’, ‘Methane
metabolism’ and ‘Alanine, aspartate and glutamate metabolism’ were
upregulated in the DHA group compared with the control group.
Overall, these results suggest that DHA may affect lipid metabolism
by regulating the gut microbiota. The effects of DHA on the gut
microbiota were evaluated in the present study. A high fat diet was
previously discovered to affect the relative abundance and
composition of the gut microbiota (15). To ensure that the alterations of
the gut microbiota were due to the effects of DHA, mice were fed a
normal diet in the present study. Since the serum TG levels were
observed to be significantly decreased in the DHA group, the effect
of DHA on the lipid metabolism will be further evaluated using mice
fed with a high fat diet in our future study.
In the present study, KEGG signaling pathway
enrichment analysis revealed that the signaling pathways involved
in ‘Infectious Diseases’ and ‘Neurodegenerative Diseases’ were
significantly downregulated in the DHA group compared with the
control group. Notably, Liu et al (2) reported that DHA had the potential to
suppress the NF-κB signaling pathway and inhibit the inflammatory
response in lipopolysaccharide-induced septic acute kidney injury
within a mouse model. In addition, Kim et al (32) reported that artemisinin inhibited
the interaction between nod-like receptor protein 3 (NLRP3) and
never in mitosis geneA-related kinase 7 (Nek7) in NLRP3
inflammasome activation. Furthermore, it was indicated that
artemisinin may inhibit AMP kinase/NF-κB/NLRP3 inflammasome
signaling in macrophages in order to protect the aorta from
atherosclerotic lesions (33).
Neurodegenerative diseases are characterized by the deterioration
of neuronal structures and functions (34), and it has been suggested that
artemisinin may be used as a treatment for neurodegenerative
diseases by inhibiting oxidation, inflammation and decreasing the
level of amyloid β protein (34).
The results of the present study indicated that DHA may suppress
the signaling pathways involved in ‘Infectious Diseases’ and
‘Neurodegenerative Diseases’ by regulating the gut microbiota.
In conclusion, the findings of the present study
revealed that DHA decreased the TG serum levels in mice and 16S
rDNA gene analysis suggested that DHA may have the potential to
increase the diversity of the gut bacteria. Mice treated with DHA
exhibited increased numbers of Firmicutes and
Saccharibacteria, and decreased Deferribacteres and
Actinobacteria on the phylum level compared with the control
group. KEGG signaling pathway enrichment analysis subsequently
demonstrated that DHA upregulated the signaling pathways involved
in ‘Energy Metabolism’ and ‘Nucleotide Metabolism’, while
downregulating the signaling pathways involved in ‘Infectious
Diseases’ and ‘Neurodegenerative Diseases’. Altogether, these
results suggested that DHA may exert its functions, including
decreasing the serum TG levels and inducing anti-inflammatory and
anti-neurodegenerative responses through regulating the composition
of the gut microbiota.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81803912 and
31671520), the Scientific Research Project of the Administration of
Traditional Chinese Medicine of Guangdong Province (grant no.
20182079), the Characteristic Innovation Project (Natural Science)
of the Education Department of Guangdong Province, the ‘Innovation
Strong School Project’ of Guangdong Pharmaceutical University
(grant no. 2017KTSCX102) and the Science and Technology Project of
Yue-Xiu District of Guangzhou (grant no. 2018-WS-011).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors contributions
ZL and JY designed and conceived the study; YYL
established the model mice and performed the serum analysis; YY
designed the study, analyzed and interpreted the results, wrote the
first draft of the manuscript and critically revised the
manuscript; and YTL, LY and XZ performed the H&E staining. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The experimental procedures in the present study
were approved by the Committee on Laboratory Animal Care and Use of
Guangdong Pharmaceutical University (Guangzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ho WE, Peh HY, Chan TK and Wong WS:
Artemisinins: Pharmacological actions beyond anti-malarial.
Pharmacol Ther. 142:126–139. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu X, Lu J, Liao Y, Liu S, Chen Y, He R,
Men L, Lu C, Chen Z, Li S, et al: Dihydroartemisinin attenuates
lipopolysaccharide-induced acute kidney injury by inhibiting
inflammation and oxidative stress. Biomed Pharmacother.
117:1090702019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lu F, He XL, Richard C and Cao J: A brief
history of artemisinin: Modes of action and mechanisms of
resistance. Chin J Nat Med. 17:331–336. 2019.PubMed/NCBI
|
|
4
|
Chen X, Bian M, Jin H, Lian N, Shao J,
Zhang F and Zheng S: Dihydroartemisinin attenuates alcoholic fatty
liver through regulation of lipin-1 signaling. IUBMB Life.
71:1740–1750. 2019. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li Y, Zhou X, Liu J, Gao N, Yang R, Wang
Q, Ji J, Ma L and He Q: Dihydroartemisinin inhibits the
tumorigenesis and metastasis of breast cancer via downregulating
CIZ1 expression associated with TGF-β1 signaling. Life Sci.
248:1174542020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yin J, Xia W, Zhang Y, Ding G, Chen L,
Yang G, Huang S, Jia Z and Zhang A: Role of dihydroartemisinin in
regulating prostaglandin E2 synthesis cascade and inflammation in
endothelial cells. Heart Vessels. 33:1411–1422. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhu H and Ji W: Dihydroartemisinin
ameliorated ovalbumin-induced asthma in mice via regulation of
miR-183C. Med Sci Monit. 25:3804–3814. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schroeder BO and Bäckhed F: Signals from
the gut microbiota to distant organs in physiology and disease. Nat
Med. 22:1079–1089. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kong CY, Li ZM, Han B, Zhang ZY, Chen HL,
Zhang SL, Xu JQ, Mao YQ, Zhao YP and Wang LS: Diet consisting of
balanced yogurt, fruit and vegetables modifies the gut microbiota
and protects mice against nonalcoholic fatty liver disease. Mol
Nutr Food Res. 63:e19002492019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kareva I: Metabolism and gut microbiota in
cancer immunoediting, CD8/Treg ratios, immune cell homeostasis and
implications for cancer (immuno) therapy: Concise Review. Stem
Cells. 37:1273–1280. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen YX, Lai LN, Zhang HY, Bi YH, Meng L,
Li XJ, Tian XX, Wang LM, Fan YM, Zhao ZF, et al: Effect of
artesunate supplementation on bacterial translocation and dysbiosis
of gut microbiota in rats with liver cirrhosis. World J
Gastroenterol. 22:2949–2959. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Efferth T: From ancient herb to modern
drug: Artemisia annua and artemisinin for cancer therapy. Semin
Cancer Biol. 46:65–83. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Olliaro PL, Nair NK, Sathasivam K, Mansor
SM and Navaratnam V: Pharmacokinetics of artesunate after single
oral administration to rats. BMC Pharmacol. 1:122001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dai T, Jiang W, Guo Z, Xie Y and Dai R:
Comparison of in vitro/in vivo blood distribution and
pharmacokinetics of artemisinin, artemether and dihydroartemisinin
in rats. J Pharm Biomed Anal. 162:140–148. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang Y, Yang F, Huang M, Wu H, Yang C,
Zhang X, Yang L, Chen G, Li S, Wang Q, et al: Fatty liver and
alteration of the gut microbiome induced by diallyl disulfide. Int
J Mol Med. 44:1908–1920. 2019.PubMed/NCBI
|
|
16
|
Yang Y, Lei Z, Huang L, Yang F, Zhang N,
Yuan J, Li K, Chen J and Zhang J: Antitumor ability of berberine
accompanied by modulation of gut microbiome in sarcoma-180
tumor-bearing mice. Int J Pharmacol. 14:460–470. 2018. View Article : Google Scholar
|
|
17
|
Caporaso JG, Kuczynski J, Stombaugh J,
Bittinger K, Bushman FD, Costello EK, Fierer N, Peña AG, Goodrich
JK, Gordon JI, et al: QIIME allows analysis of high-throughput
community sequencing data. Nat Methods. 7:335–336. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Valero-Mora PM: ggplot2: Elegant Graphics
for Data Analysis. Journal of Statistical Software. 35:2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kemp PF and Aller JY: Bacterial diversity
in aquatic and other environments: What 16S rDNA libraries can tell
us. FEMS Microbiol Ecol. 47:161–177. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Frosini BV: Descriptive measures of
ecological diversity. Environmetrics, in Encyclopedia of Life
Support Systems (EOLSS). Jureckova J and El-Shaarawi AH: Eolss
Publishers; Oxoford, UK: 2004
|
|
21
|
Oksanen J, Blanchet FG, Kindt R, Legendre
P, OHara RB, Simpson GL, Solymos P, Stevens MHH and Wagner H:
Vegan: Community Ecology Package. R package. 1:17–12. 2011.
|
|
22
|
Tu Y: Artemisinin-a gift from traditional
Chinese medicine to the world (Nobel Lecture). Angew Chem Int Ed
Engl. 55:10210–10226. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang J, Xu C, Liao FL, Jiang T, Krishna S
and Tu Y: A Temporizing Solution to “Artemisinin Resistance”. N
Engl J Med. 380:2087–2089. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yuliang W, Zejian W, Hanlin S, Ming Y and
Kexuan T: The hypolipidemic effect of artesunate and ursolic acid
in rats. Pak J Pharm Sci. 28:871–874. 2015.PubMed/NCBI
|
|
25
|
Wang YL, Wang ZJ, Shen HL, Yin M and Tang
KX: Effects of artesunate and ursolic acid on hyperlipidemia and
its complications in rabbit. Eur J Pharm Sci. 50:366–371. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wolff D, Frese C, Schoilew K, Dalpke A,
Wolff B and Boutin S: Amplicon-based microbiome study highlights
the loss of diversity and the establishment of a set of species in
patients with dentin caries. PLoS One. 14:e02197142019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Clarke SF, Murphy EF, OSullivan O, Lucey
AJ, Humphreys M, Hogan A, Hayes P, OReilly M, Jeffery IB,
Wood-Martin R, et al: Exercise and associated dietary extremes
impact on gut microbial diversity. Gut. 63:1913–1920. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hills RD Jr, Pontefract BA, Mishcon HR,
Black CA, Sutton SC and Theberge CR: Gut microbiome: Profound
implications for diet and disease. Nutrients. 11:E16132019.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu R, Zhao D, An R, Wang Z, Li Y, Shi B
and Ni Q: Linggui Zhugan Formula improves glucose and lipid levels
and alters gut microbiota in high-fat diet-induced diabetic mice.
Front Physiol. 10:9182019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Song B, Zhong YZ, Zheng CB, Li FN, Duan YH
and Deng JP: Propionate alleviates high-fat diet-induced lipid
dysmetabolism by modulating gut microbiota in mice. J Appl
Microbiol. 127:1546–1555. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mazloom K, Siddiqi I and Covasa M:
Probiotics: How effective are they in the fight against obesity?
Nutrients. 11:E2582019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim SK, Choe JY and Park KY:
Anti-inflammatory effect of artemisinin on uric acid-induced NLRP3
inflammasome activation through blocking interaction between NLRP3
and NEK7. Biochem Biophys Res Commun. 517:338–345. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jiang Y, Du H, Liu X, Fu X, Li X and Cao
Q: Artemisinin alleviates atherosclerotic lesion by reducing
macrophage inflammation via regulation of AMPK/NF-κB/NLRP3
inflammasomes pathway. J Drug Target. 16:1–10. 2019.
|
|
34
|
Lu BW, Baum L, So KF, Chiu K and Xie LK:
More than anti-malarial agents: Therapeutic potential of
artemisinins in neurodegeneration. Neural Regen Res. 14:1494–1498.
2019. View Article : Google Scholar : PubMed/NCBI
|