Introduction
Lower extremity arteriosclerosis obliterans disease
(LEAOD) affects ~17% of individuals aged 55–75 years worldwide, and
is characterized by intermittent claudication, rest pain and
gangrene caused by lower extremity ischemia (1,2).
Previous studies have indicated that atherosclerosis is a
persistent chronic inflammatory disease (3,4). The
initial steps of atherosclerosis formation are endothelial damage,
followed by platelet aggregation and leukocyte infiltration under
the injured endothelial cells. Subsequently, macrophage activation,
smooth muscle cell proliferation and migration, and foam cell
formation occur, eventually leading to atherosclerosis (5,6).
Platelet-derived growth factor-BB (PDGF-BB) serves
an important role in the initiation and progression of
atherosclerosis (7). The
expression of PDGF-BB in vascular smooth muscle cells (VSMCs) is
upregulated and activated in injured vessels (7). Previous studies have reported that
PDGF-BB can induce a variety of biological effects by increasing
the expression of proinflammatory cytokines, such as interleukin
(IL)-1β, IL-17A and TNF-α, which can induce the proliferation and
migration of VSMCs (8,9). Therefore, inhibiting the
proliferation and migration of VSMCs caused by PDGF-BB may serve as
an important preventative strategy for atherosclerosis.
A number of proteins, including tumor necrosis
factor (TNF) superfamily member 11 (TRANCE) (10), IL-6 (11) and transforming growth factor
(TGF)-β (12,13), serve potentially important roles
during the development and progression of atherosclerosis. T-cell
immunoglobulin and mucin domain 3 (TIM-3) as a regulatory factor
alters the function of a number of different immune cells,
including effector/regulatory T cells, macrophages, monocytes and
NK cells (14,15). Previous studies using immune cell
experimental models have identified TIM-3 as an antiatherosclerotic
agent during atherosclerosis progression (16,17).
However, the regulatory role of TIM-3 on
PDGF-BB-induced human artery vascular smooth muscle cell (HASMC)
proliferation and migration, and how the pathway is activated
remains unclear. In the present study, the effects of TIM-3 on
HASMCs and the underlying mechanisms were investigated.
Materials and methods
Specimen collection
Serum samples and femoral arterial specimens were
collected in at The First Affiliated Hospital of Sun Yat-sen
University, China between September 2017 and July 2019. Serum
samples were collected from five patients with LEAOD with the mean
age of 63.5 years old (four male and one female; age range: 56–75
years old) and five healthy controls with the mean age of 65.5
years old (four male and one female; age range: 52–71 years old).
Serum samples of the two groups were also age, sex and
ethnicity-matched. Serum samples were stored at −80°C until further
analysis. Femoral arterial specimens were collected from five
patients with LEAOD with the mean age of 62 years old (three male
and two female; age range: 53–73 years old) who had undergone
amputations due to severe lower extremity ischemia and gangrene in
the vascular surgery department, and normal femoral arterial
specimens were collected from five cases without LEAOD with the
mean age of 60.5 years old (four male and one female; age range:
50–69 years old) who had undergone amputations due to severe lower
limb trauma caused by a traffic accident in the departments of
trauma surgery and emergency. Internal and external tissues were
removed from the blood vessels during specimen collection. After
washing with normal saline, some specimens were immediately frozen
in liquid nitrogen and some specimens were fixed in 4°C, 4%
formaldehyde until further analysis. Written informed consent was
obtained from the participants or their relatives. The present
study was approved by the Research Ethics Committee of the First
Affiliated Hospital of Sun Yat-sen University.
Protein arrays
Serum samples were analyzed using the Human Cytokine
Antibody Array 440 (cat. no. QAH-CAA-440; RayBiotech, Inc.),
according to the manufacturer's protocol.
Cell culture and treatment
HASMCs were isolated from the femoral artery
specimens and identified by anti-α-smooth muscle actin antibody
staining as previously described (18). HASMCs were cultured in SMCM medium
(Sciencell Research Laboratories, Inc.) supplemented with 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.) at 37°C in a 5%
CO2 humidified incubator. HASMCs from passages 5 to 9
were used for subsequent experiments. When the cell density reached
50–60%, HASMCs were incubated with 20 ng/ml PDGF-BB (R&D
Systems, Inc.), TIM-3 (1,000 ng/ml; R&D Systems, Inc.),
anti-TIM-3 mAb (10 µg/ml; BioLegend, Inc.) or were only incubated
with serum supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) at 37°C for 48 h. Subsequently, HASMCs were
transduced with the LV-TIM-3 lentiviral vector (Genechem Co., Ltd.)
according to the manufacturer's protocol The empty LV-vector
(Genechem Co., Ltd.) was used as the control.
EdU assay
HASMCs were treated according to the manufacturer's
protocol (Cell-Light EdU Apollo567 Kit; Guangzhou RiboBio Co.,
Ltd.). EdU-positive cells were observed using an Axio Observer Z1
fluorescence microscope (Zeiss GmbH) and analyzed using ImageJ
version 1.8.0 (National Institutes of Health).
Transwell and wound-healing
assays
For the Transwell assay, Treated HASMCs
(1×104 cells/well) resuspended in serum-free SMCM were
plated into the upper chambers of the Transwell plates (24 wells;
dr=6.5 mm; pore size: 8.0 µm; Corning, Inc.) and the lower chambers
were plated in 10% FBS SMCM. After incubation at 37°C for 24 h,
migratory cells on the lower surface of the Transwell membranes
were fixed using 4% formaldehyde at 25°C for 20 min and washed
twice with PBS. Then the membranes were stained with 0.1% crystal
violet at 25°C for 15 min.
For the wound-healing assay, at ~90% confluency, a
1,000-µl disposable pipette tip was used to form a straight scratch
wound through the treated HASMC cell monolayer. The cell layer was
washed with PBS gently and subsequently incubated with serum-free
SMCM medium. Images of the wound were captured using a light
microscope ×100 magnification (BX51W; Olympus Co., Ltd.) at 0 and
24 h. Cell migration was analyzed using ImageJ Version 1.8.0
(National Institutes of Health).
Western blot analysis
HASMCs were washed with precooled PBS for three
times and subsequently total protein was extracted from HASMCs
using RIPA buffer (Thermo Fisher Scientific, Inc.) on ice. Then the
protein concentration was quantified by bicinchoninic acid method.
Equal amounts of protein (20 µg/lane) were separated by 10–15%
SDS-PAGE, and then transferred to PVDF membranes (EMD Millipore).
Subsequently, the membranes were blocked with 5% bovine serum
albumin (BioFroxx, Inc.) at 22–25°C for 1 h. Then the membranes
were incubated overnight at 4°C with primary antibodies targeted
against: TIM-3 (1:500; cat. no. 1701-20; Hangzhou HuaAn
Biotechnology Co., Ltd.), PDGF-BB (1:500; cat. no. 9704; Abcam),
p65 NFκB (1:1,000; cat. no. 8242; Cell Signaling Technology, Inc.),
phosphorylated (p)-p65 NFκB (1:1,000; cat. no. 3033; Cell Signaling
Technology, Inc.), IL-6 (1:1,000; cat. no. 12153; Cell Signaling
Technology, Inc.), TNF-α (1:1,000; cat. no. 8184; Cell Signaling
Technology, Inc.) and GAPDH (1:1,000; cat. no. 5174; Cell Signaling
Technology, Inc.). Following primary incubation, the membranes were
incubated at 22–25°C for 1 h with corresponding rabbit anti-mouse
horseradish peroxidase(HRP)-conjugated secondary antibody (1:3,000;
cat. no. 58802, Cell Signaling Technology, Inc.) and mouse
anti-rabbit HRP-conjugated secondary antibody (1:5,000; cat. no.
93702; Cell Signaling Technology, Inc.). Protein bands were
visualized using the Novex ECL kit (Invitrogen; Thermo Fisher
Scientific, Inc.). Protein expression was quantified using ImageJ
Version 1.8.0 (National Institutes of Health) with GAPDH as the
loading control.
Flow cytometry
Flow cytometry was performed to detect the
quantitative proportion of TIM-3-positive HASMCs. Forward scatter
(FSC) and side scatter (SSC) were used to find viable, single cell
populations. Treated HASMCs were resuspended and labeled with the
TIM-3 (D5D5R™) Rabbit mAb (PE Conjugate) (cat. no. 67761, Cell
Signaling Technology, Inc.) Unstained HASMCs were used to set the
negative control populations. TIM-3-positive cells were then
determined from this gated population by flow cytometry using the
CyAN™ ADP flow cytometer (Beckman-Coulter, Inc.) and analyzed by
FlowJo Version 10.0 (Tree Star, Inc.).
Immunohistochemistry
Femoral arterial tissues were fixed in 4%
polyformaldehyde at 22–25°C for 24 h and then embedded in paraffin.
Following deparaffinization, rehydration with a descending alcohol
series, 3% hydrogen peroxide was used to block the endogenous
peroxidase activity, then the citric acid buffer was used for
antigen retrieval at 95°C for 10 min and PBS buffer was used for
washing. The sections (4 µm) were incubated at 4°C overnight with
anti-TIM-3 (1:100; cat. no. 1701-20; Hangzhou HuaAn Biotechnology
Co., Ltd.). Subsequently, the sections were incubated at room
temperature for 1 h with goat anti-mouse IgG-PE: (1:200; cat. no.
sc-3738; Santa Cruz Biotechnology, Inc.). The sections were
counterstained with hematoxylin at 22–25°C for 5 min and observed
using a light microscope (BX51W1; Olympus Corporation) at ×400
magnification. The images were analyzed using ImageJ Version 1.8.0
(National Institutes of Health).
Statistical analysis
Data are presented as the mean ± SEM. The Student's
t-test was used to compare data containing two groups. One-way
ANOVA followed by Tukey's post hoc test was used to compare data
containing ≥3 groups. Pearson's correlation coefficient test was
used to analyze the correlation between two indexes. Statistical
analyses were performed using SPSS software (version 17.0; SPSS,
Inc.). P<0.05 was considered to indicate a statistically
significant difference.
Results
Microarray comparison of the protein
expression profiles in the serum between patients with LEAOD and
healthy controls
To further investigate the mechanisms underlying
LEAOD, a 440-protein array was used to analyze the differential
expression of proteins in the serum of five patients with LEAOD
compared with healthy controls. A total of 22 proteins were
differentially expressed, 13 proteins were upregulated and 9
proteins were downregulated in the serum of patients with LEAOD
compared with healthy controls. The top 10 most highly expressed
proteins were TRANCE, TIM-3, CD6, TGF-β, troponin I, IL-1β,
P-cadherin, TNF-related apoptosis-inducing ligand receptor-3 (TRAIL
R3), TNF factor-like weak inducer of apoptosis receptor (TWEAK) and
vascular cell adhesion molecule-1 (VCAM-1) (Table I).
 | Table I.Top 10 most highly expressed proteins
in the serum of patients with lower extremity arteriosclerosis
obliterans disease vs. healthy control. |
Table I.
Top 10 most highly expressed proteins
in the serum of patients with lower extremity arteriosclerosis
obliterans disease vs. healthy control.
| Rank | Protein | Fold-change | P-value |
|---|
| 1 | TRANCE | 3.12 | 0.012a |
| 2 | TIM-3 | 2.38 | 0.027a |
| 3 | CD6 | 2.16 | 0.011a |
| 4 | TGF-β | 1.66 | 0.044a |
| 5 | Troponin I | 1.59 | 0.022a |
| 6 | IL-1β | 1.17 | 0.010a |
| 7 | P-cadherin | 1.13 | 0.011a |
| 8 | TRAIL R3 | 1.06 | 0.027a |
| 9 | TWEAK | 0.95 | 0.018a |
| 10 | VCAM-1 | 0.72 | 0.015a |
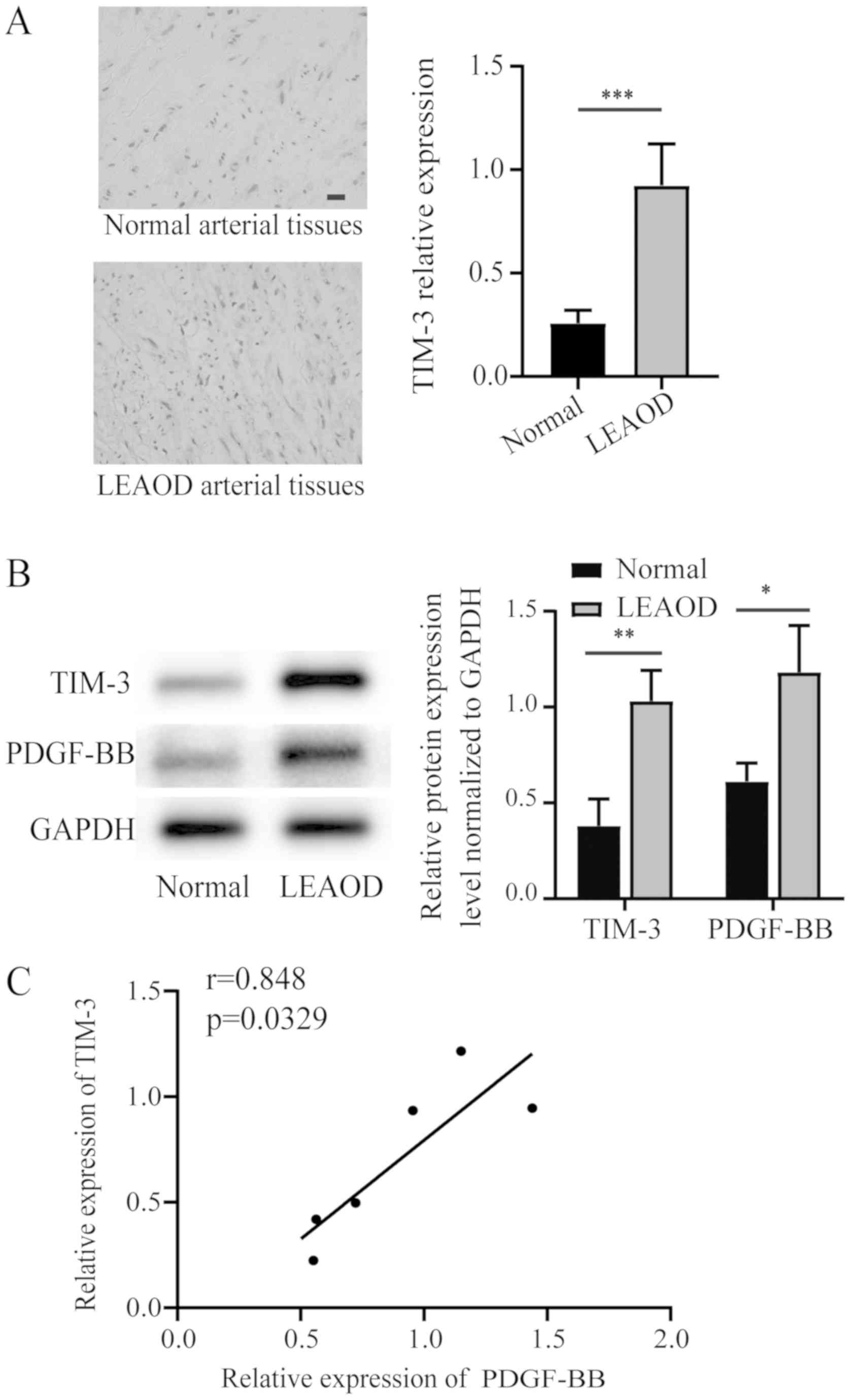
Increased expression of TIM-3 in LEAOD
arterial tissues
According to the immunohistochemistry results, the
tunica media of LEAOD arterial tissues exhibited high levels of
TIM-3 positive staining, whereas normal arterial tissues exhibited
little TIM-3 positive staining (Fig.
1A). The protein array data for TIM-3 were further investigated
using western blotting. TIM-3 and PDGF-BB expression levels were
significantly higher in LEAOD arterial tissues compared to normal
arterial tissues (Fig. 1B). A
positive correlation between the relative expression PDGF-BB and
TIM-3 was also identified (Fig.
1C).
TIM-3 inhibits proliferation and
migration in PDGF-BB-stimulated HASMCs
Previous studies have reported that the expression
of TIM-3 is upregulated in atherosclerotic patients or animal
models (16,19,20).
In the present study, it was demonstrated that the expression of
TIM-3 was upregulated in LEAOD arterial tissues (Fig. 1B). It is likely that the expression
of TIM-3 is upregulated in atherosclerotic VSMCs, due to there are
high numbers of VSMCs are in the media of arterial tissues
(21). PDGF-BB is a critical
factor that promotes the migration and proliferation of VSMCs,
which causes atherosclerosis (7).
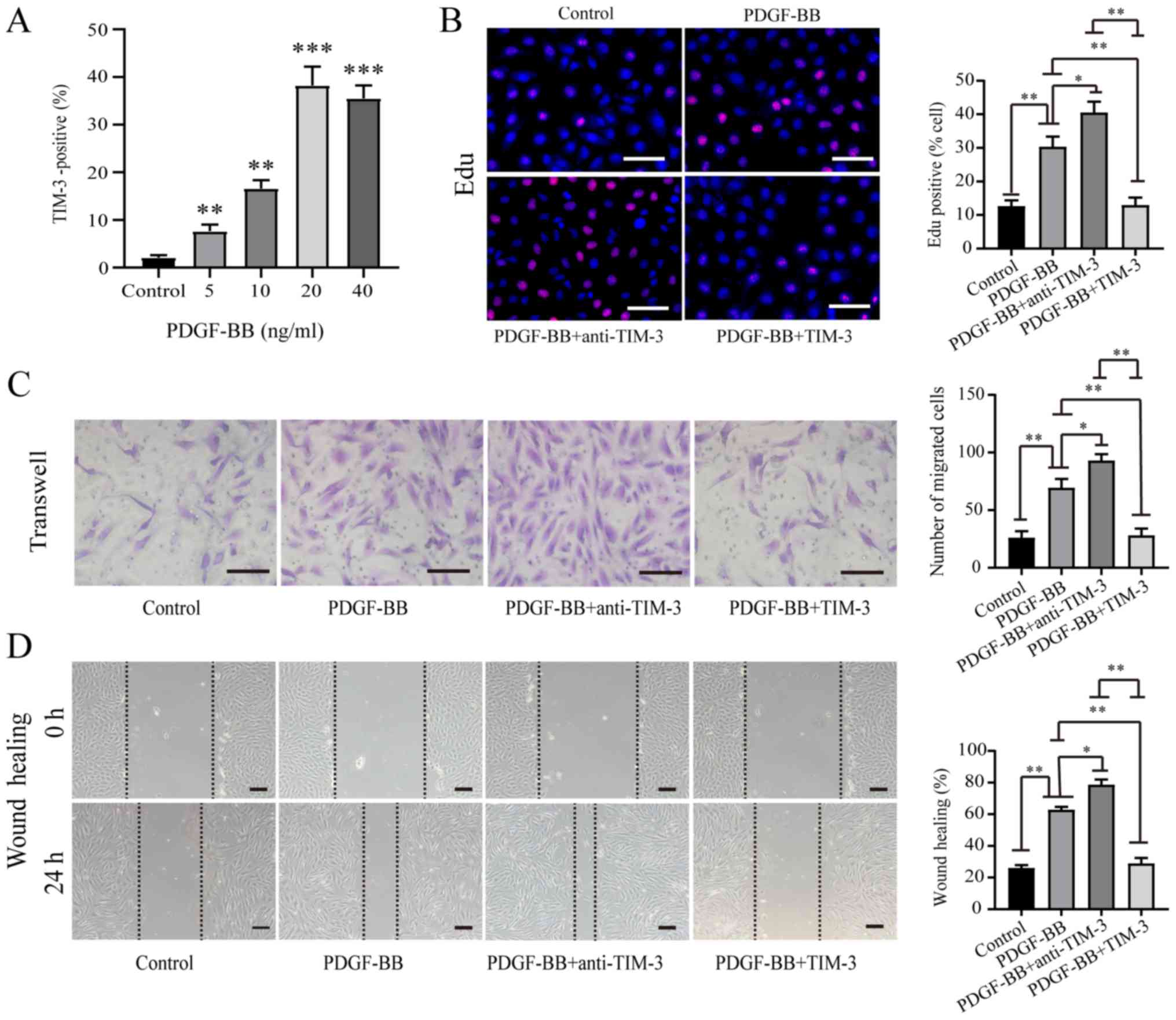
HASMCs were stimulated with various concentrations of PDGF-BB. A
dose-dependent relationship was identified between PDGF-BB
concentration and TIM-3 expression by HASMCs (Fig. 2A). EdU (Fig. 2B), Transwell (Fig. 2C) and wound healing (Fig. 2D) assays were performed to assess
HASMC proliferation and migration following stimulation with
PDGF-BB (20 ng/ml) in the absence or presence of TIM-3 (1,000
ng/ml) and anti-TIM-3 mAb (10 µg/ml). The proportion of
EdU-positive cells was significantly higher in the PDGF-BB group
(30.36±3.02%) compared to the control group (12.64±1.71%).
Furthermore, the proportion of EdU-positive cells was significantly
higher in the PDGF-BB + anti-TIM-3 group (40.54±3.24%) compared to
the PDGF-BB group (30.36±3.02%). However, the proportion of
EdU-positive cells was significantly reduced in the PDGF-BB + TIM-3
group compared to the PDGF-BB and PDGF-BB + anti-TIM-3 groups, but
was not significantly different to the control group (Fig. 2B). Treatment with PDGF-BB
significantly increased the mean number of migratory cells compared
to the control group (69.6±7.4 vs. 26.2±5.6). The PDGF-BB +
anti-TIM-3 group exhibited a significantly increased mean number of
migratory cells compared to the PDGF-BB group (93.0±5.5 vs.
69.6±7.4). However, the PDGF-BB + TIM-3 group exhibited
significantly decreased migratory ability compared to the PDGF-BB
and PDGF-BB + anti-TIM-3 groups, and a similar level compared to
the control group (Fig. 2C). The
wound healing assay results were similar to the Transwell assay
results (Fig. 2D). The results
indicated that PDGF-BB promoted the proliferation and migration of
HASMCs, TIM-3 inhibited the effect of PDGF-BB on HASMCs, and
anti-TIM-3 blocked the effect of TIM-3 to promote the proliferation
and migration of HASMCs.
TIM-3 overexpression inhibits
proliferation and migration in PDGF-BB-stimulated HASMCs
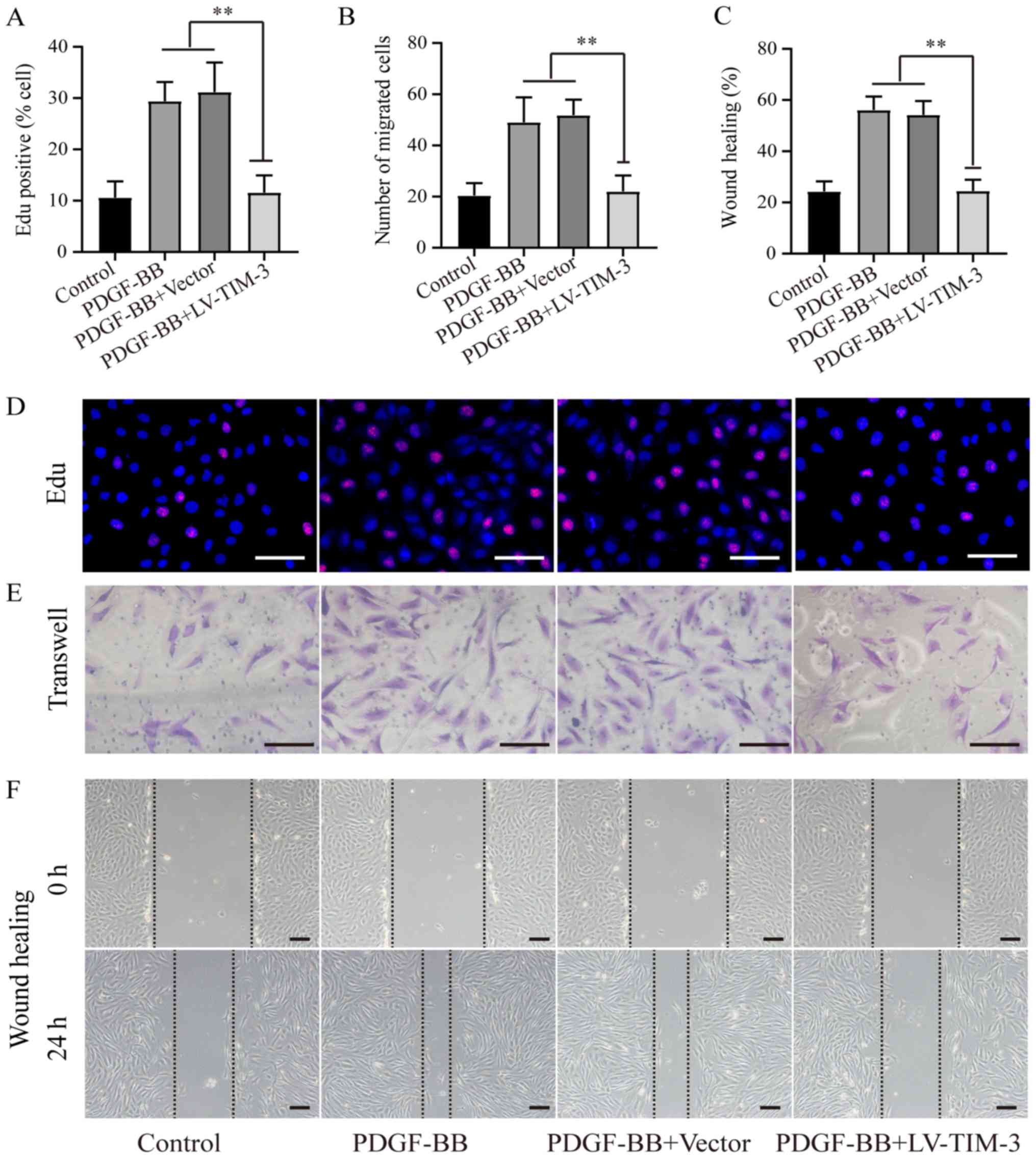
To further investigate the functional role of TIM-3,
a lentiviral vector (LV-TIM-3) was used to overexpress TIM-3 in
HASMCs (Fig. S1). To determine
the effect of TIM-3 on the proliferation and migration of HASMCs,
EdU (Fig. 3A and D), Transwell
(Fig. 3B and E) and wound healing
(Fig. 3C and F) assays were
performed. The proportion of EdU-positive cells was significantly
lower in the PDGF-BB + LV-TIM-3 group (11.58±3.36%) compared to the
PDGF-BB (29.42±3.72%) and the PDGF-BB + Vector (31.18±7.78%)
groups, but was not significantly different compared to the control
group (10.06±3.18%; Fig. 3A). The
mean number of migratory cells was significantly reduced in the
PDGF-BB + LV-TIM-3 group (22.0±6.2) compared to the PDGF-BB
(49.0±9.8) and PDGF-BB + Vector (51.8±6.1) groups, but was also not
significantly different compared to the control group (22.4±4.9;
Fig. 3B). The wound healing assay
results were similar to the Transwell assay results (Fig. 3C). The results demonstrated that
LV-TIM-3-mediated TIM-3 overexpression in HASMCs inhibited
proliferation and migration in PDGF-BB-stimulated HASMCs.
TIM-3 decreases IL-6 and TNF-α
expression
TIM-3 is an important inflammatory response
regulator (16). To further
investigate the role of TIM-3 in atherosclerosis, the expression of
proinflammatory cytokines in PDGF-BB-stimulated HASMCs treated with
or without LV-TIM-3 was investigated. Western blot analysis
indicated that the PDGF-BB group exhibited higher expression levels
of IL-6 and TNF-α compared with the control group. TIM-3
overexpression in HASMCs resulted in a significant decrease in IL-6
and TNF-α expression compared to the PDGF-BB group (Fig. 4A and B). The results indicated that
TIM-3 suppressed PDGF-BB-induced inflammatory responses by
decreasing the expression levels of proinflammatory factors.
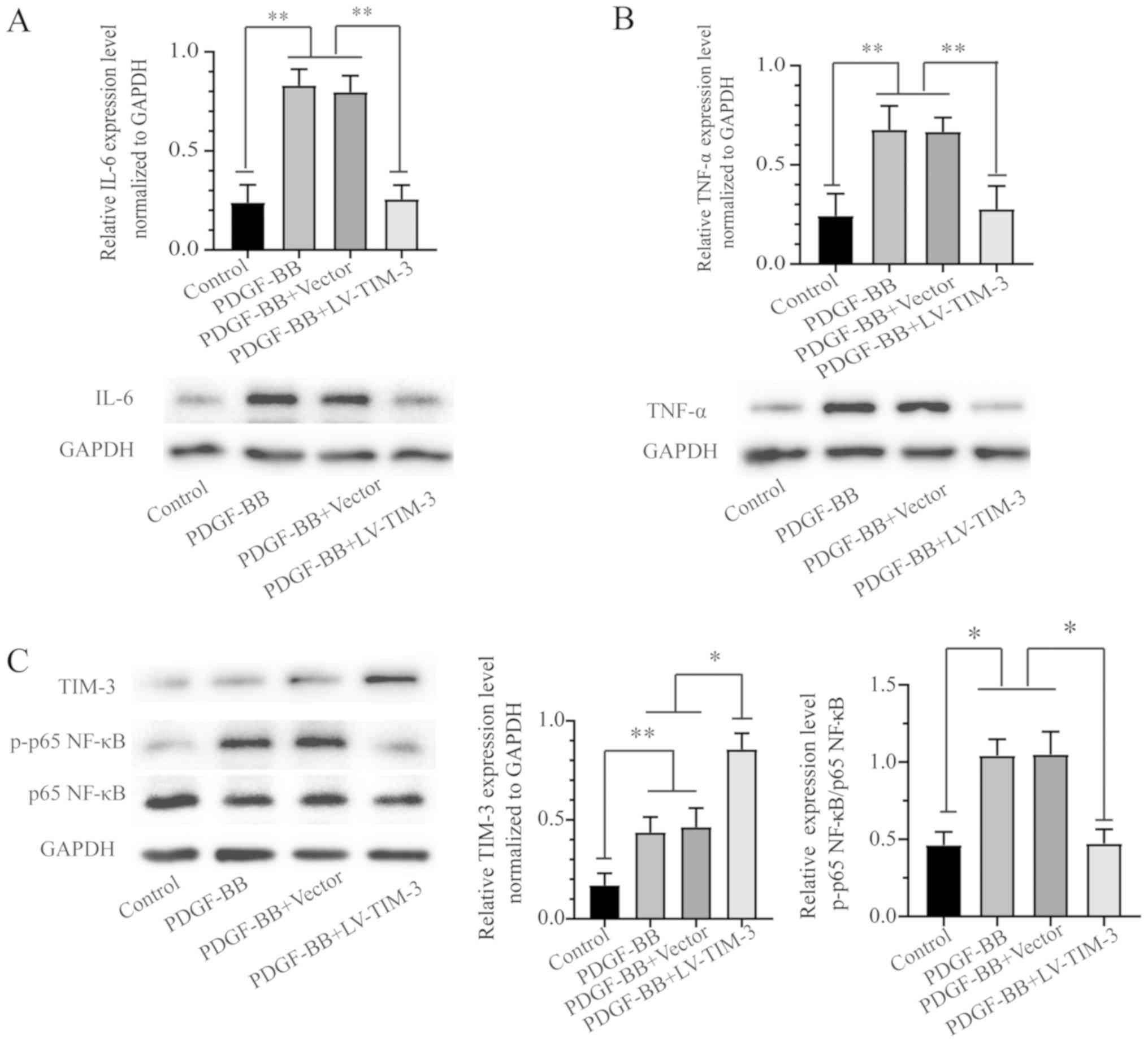 | Figure 4.IL-6 and TNF-α expression in the four
treatment groups and the effect of TIM-3 overexpression on the
NF-κB signaling pathway in PDGF-BB-stimulated HASMCs. Protein
expression levels of (A) IL-6, (B) TNF-α, (C) TIM-3, p-p65 NF-κB
and p65 NF-κB were detected by western blotting. *P<0.05,
**P<0.01. IL-6, interleukin-6; TNF-α, tumor necrosis factor-α;
TIM-3, T-cell immunoglobulin and mucin domain 3; PDGF-BB,
platelet-derived growth factor-BB; HASMCs, human artery vascular
smooth muscle cells; p, phosphorylated; LV, lentivirus. |
TIM-3 protects HASMCs from the
PDGF-BB-induced proinflammatory response by inhibiting NF-κB
signaling
The NF-κB signaling pathway is important for
PDGF-BB-induced VSMC inflammatory factor secretion, which promotes
proliferation and migration in VSMCs (22). To further investigate whether the
NF-κB signaling pathway was related to TIM-3, the protein
expression of members of the signaling pathway were assessed by
western blotting in the control, PDGF-BB, PDGF-BB + Vector and
PDGF-BB + LV-TIM-3 groups. The expression of TIM-3 and p-p65 NF-κB
was significantly increased in the PDGF-BB group compared to the
control group, but the total p65 NF-κB levels were not
significantly altered (Fig. 4C).
The results indicated that PDGF-BB treatment increased the
expression of p-p65-NF-κB but not total-NF-κB, and activated the
NF-κB signaling pathway in HASMCs. Furthermore TIM-3 overexpression
reversed the PDGF-BB-induced effects on p65 NF-κB.
Discussion
TIM-3 plays an important role in autoimmune and
alloimmune diseases, immune tolerance and tumors (23,24).
Previous studies have reported that TIM-3 displays an
antiatherosclerotic role during the generation and progression of
atherosclerosis in traditional immune cells, including mononuclear
macrophages, T cells and NK cells (16,19);
however, few studies have investigated the function of TIM-3 in
non-classical immune cells. In the present study, TIM-3 expression
was significantly increased in LEAOD arterial tissues compared to
normal arterial tissues. The present study revealed that TIM-3 was
expressed in HASMCs, which are non-classical immune cells, and was
upregulated following PDGF-BB stimulation.
PDGF-BB initiates a number of biological processes
by increasing the levels of various proinflammatory factors,
including high motility group box 1 protein, IL-1, TNF-α and IL-6,
and plays a key role in VSMC proliferation and migration (25). Inhibiting VSMC proliferation and
migration is an important therapeutic strategy for atherosclerosis
(26–28). TIM-3 displays antiatherosclerotic
effects during the development of atherosclerosis in traditional
immune cells, but its effect on VSMC proliferation and migration
has not been reported (16). In
the present study, a positive correlation between the relative
expression of PDGF-BB and TIM-3 in tissues was identified. The
relationship may be due to the increased expression of PDGF-BB
during atherogenesis, followed by the upregulation of inflammatory
factors in VSMCs, such as IL-6 and TNF-α. In the present study,
VSMCs displayed an adaptive response against inflammation, and the
expression of TIM-3, an immune tolerance factor, increased
correspondingly.
In the present study, the expression of TIM-3
increased following PDGF-BB stimulation of HASMCs, and TIM-3
reduced PDGF-BB-induced migration and proliferation in HASMCs.
Treatment of HASMCs with anti-TIM-3 reversed the effects of TIM-3.
Therefore, it was hypothesized that TIM-3 displays increased
adaptability to PDGF-BB, negatively regulates the PDGF-BB-induced
inflammatory response, and inhibits migration and proliferation in
HASMCs.
Subsequently, TIM-3 was overexpressed in HASMCs
using LV-TIM-3 to investigate whether TIM-3 was a functional
effector in HASMCs. TIM-3 overexpression inhibited proliferation
and migration in PDGF-BB-stimulated HASMCs. Qiu et al
(29) reported that TIM-3
inhibited ox-low density lipoprotein-induced atherogenic responses
in HUVECs, which was consistent with the results of the present
study, indicating that TIM-3 displays antiatherosclerotic
effects.
The ‘inflammation theory’ and ‘injury-response
theory’ have become mainstream theories for the pathogenesis of
atherosclerosis (30,31). NF-κB is a transcription factor that
is abundantly expressed in human atherosclerotic lesion VSMCs,
macrophages and endothelial cells, and is associated with various
signaling pathways involved in the inflammatory response, which
induce atherosclerosis development (32,33).
The p50/p65 heterodimer is the most common NF-κB/Rel proto-oncogene
complex, which is present in the majority of cells in vivo.
Increased p65 NF-κB phosphorylation often indicates activation of
the NF-κB signaling pathway (34,35).
In the present study, the proinflammatory factors IL-6 and TNF-α
were expressed at high levels in PDGF-BB-stimulated HASMCs
alongside increased levels of p65 NF-κB phosphorylation. The
results indicated that PDGF-BB stimulation activated the NF-κB
signaling pathway and the expression of associated proinflammatory
factors in VSMCs. In addition, based on the results of the present
study, TIM-3 upregulation may serve as a self-regulatory mechanism
of VSMCs against inflammation, but this induction may not be
sufficient to counteract the proinflammatory effects of PDGF-BB.
TIM-3 overexpression resulted in a significant decrease in the
expression levels of proinflammatory factors and p65 NF-κB
phosphorylation in HASMCs, which suggested that TIM-3
overexpression inhibited the activation of the NF-κB signaling
pathway and exerted an anti-inflammatory effect.
In previous studies, it has been reported that TIM-3
can inhibit the development of atherosclerosis (16,17,29),
which were similar to the results of the present study. Although
the antiatherosclerotic effect of TIM-3 in non-classical immune
cells was indicated in the present study, the underlying mechanisms
were not identified. Therefore, further investigation is required
to identify the mediators and factors underlying the results
obtained in the present study, and to identify other signaling
pathways that may mediate the effects of TIM-3 on HASMC
proliferation and migration.
In the present study, protein arrays indicated that
TIM-3 was upregulated in the serum of patients with LEAOD.
Immunohistochemistry and western blotting of arterial tissue
further revealed that TIM-3 expression was increased in LEAOD
artery tissue compared with normal artery tissue. Furthermore, to
the best of our knowledge, the present study revealed for the first
time that TIM-3 inhibited proliferation and migration in
PDGF-BB-induced HASMCs by inhibiting HASMC inflammatory
responses.
In conclusion, TIM-3 decreased the proliferation and
migration of PDGF-BB-induced HASMCs and downregulated the
expression of proinflammatory factors by inhibiting the NF-κB
signaling pathway. The results suggested that TIM-3 may serve as a
protective factor against inflammation and atherogenic responses in
HASMCs. Furthermore, TIM-3 may serve as a potential target for the
prevention and treatment of atherosclerosis.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Dr Lei Zhao and Dr
Jin Cui (The First Affiliated Hospital of Sun Yat-sen University)
for their editorial support.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81873813).
Availability of data and materials
The datasets used and during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
CL conceived the study, performed the experiments
and wrote the manuscript. ZCW, JCQ and BHJ drafted the work, and
collected and analyzed the data. JBL and RZH contributed to the
interpretation of the data. RML and WL designed and drafted the
work. SMW and JSW conceived the experiments and revised the
manuscript for important intellectual content. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the First Affiliated Hospital of Sun Yat-sen
University (approval no. 151507). Written informed consent was
obtained from the participants or their relatives.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Criqui MH, Vargas V, Denenberg JO, Ho E,
Allison M, Langer RD, Gamst A, Bundens WP and Fronek A: Ethnicity
and peripheral arterial disease: The San Diego Population Study.
Circulation. 112:2703–2707. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fowkes FG, Rudan D, Rudan I, Aboyans V,
Denenberg JO, McDermott MM, Norman PE, Sampson UK, Williams LJ,
Mensah GA and Criqui MH: Comparison of global estimates of
prevalence and risk factors for peripheral artery disease in 2000
and 2010: A systematic review and analysis. Lancet. 382:1329–1340.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Crotty S: T Follicular Helper cell
biology: A decade of discovery and diseases. Immunity.
50:1132–1148. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yu E, Hsu HY, Huang CY and Hwang LC:
Inflammatory biomarkers and risk of atherosclerotic cardiovascular
disease. Open Med (Wars). 13:208–213. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim J, Zhang L, Peppel K, Wu JH, Zidar DA,
Brian L, DeWire SM, Exum ST, Lefkowitz RJ and Freedman NJ:
Beta-arrestins regulate atherosclerosis and neointimal hyperplasia
by controlling smooth muscle cell proliferation and migration. Circ
Res. 103:70–79. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee J and Kang H: Hypoxia Promotes
vascular smooth muscle cell proliferation through microRNA-mediated
suppression of Cyclin-dependent kinase inhibitors. Cells. 8(pii):
E8022019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Heldin CH and Westermark B: Mechanism of
action and in vivo role of platelet-derived growth factor. Physiol
Rev. 79:1283–1316. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu K, Liu C and Zhang Z: lncRNA GAS5 acts
as a ceRNA for miR-21 in suppressing PDGF-bb-induced proliferation
and migration in vascular smooth muscle cells. J Cell Biochem.
120:15233–15240. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hu W and Huang Y: Targeting the
platelet-derived growth factor signalling in cardiovascular
disease. Clin Exp Pharmacol Physiol. 42:1221–1224. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zofková I: Osteoporosis and
aterosclerosis-is there any pathogenetic association? Cas Lek Cesk.
146:246–250. 2007.(In Czech). PubMed/NCBI
|
|
11
|
Hartman J and Frishman WH: Inflammation
and atherosclerosis: A review of the role of interleukin-6 in the
development of atherosclerosis and the potential for targeted drug
therapy. Cardiol Rev. 22:147–151. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Song C, Wang Y, Cui L, Yan F and Shen S:
Triptolide attenuates lipopolysaccharide-induced inflammatory
responses in human endothelial cells: Involvement of NF-κB pathway.
BMC Complement Altern Med. 19:1982019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ponte E and Ursu HI: Overt and subclinical
hypothyroidism and atherosclerotic arteriopathy of the lower limbs
(clinical and subclinical). Rom J Endocrinol. 31:71–79.
1993.PubMed/NCBI
|
|
14
|
Hou N, Zhao D, Liu Y, Gao L, Liang X, Liu
X, Gai X, Zhang X, Zhu F, Ni M, et al: Increased expression of T
cell immunoglobulin- and mucin domain-containing molecule-3 on
natural killer cells in atherogenesis. Atherosclerosis. 222:67–73.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liang X, Xu Z, Yuan M, Zhang Y, Zhao B,
Wang J, Zhang A and Li G: MicroRNA-16 suppresses the activation of
inflammatory macrophages in atherosclerosis by targeting PDCD4. Int
J Mol Med. 37:967–975. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Foks AC, Ran IA, Wasserman L, Frodermann
V, Ter Borg MN, de Jager SC, van Santbrink PJ, Yagita H, Akiba H,
Bot I, et al: T-cell immunoglobulin and mucin domain 3 acts as a
negative regulator of atherosclerosis. Arterioscler Thromb Vasc
Biol. 33:2558–2565. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Qiu MK, Wang SC, Dai YX, Wang SQ, Ou JM
and Quan ZW: PD-1 and Tim-3 pathways regulate CD8+ T cells function
in atherosclerosis. PLoS One. 10:e01285232015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hu W, Wang M, Yin H, Yao C, He Q, Yin L,
Zhang C, Li W, Chang G and Wang S: MicroRNA-1298 is regulated by
DNA methylation and affects vascular smooth muscle cell function by
targeting connexin 43. Cardiovasc Res. 107:534–545. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang N, Zhang M, Liu RT, Zhang P, Yang
CL, Yue LT, Li H, Li YK and Duan RS: Statins reduce the expressions
of Tim-3 on NK cells and NKT cells in atherosclerosis. Eur J
Pharmacol. 821:49–56. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang F, Zhao J, Sun D and Wei N: miR-155
inhibits transformation of macrophages into foam cells via
regulating CEH expression. Biomed Pharmacother. 104:645–651. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bennett MR, Sinha S and Owens GK: Vascular
smooth muscle cells in atherosclerosis. Circ Res. 118:692–702.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Maracle CX, Agca R, Helder B, Meeuwsen
JAL, Niessen HWM, Biessen EAL, de Winther MPJ, de Jager SCA,
Nurmohamed MT and Tas SW: Noncanonical NF-κB signaling in
microvessels of atherosclerotic lesions is associated with
inflammation, atheromatous plaque morphology and myocardial
infarction. Atherosclerosis. 270:33–41. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Das M, Zhu C and Kuchroo VK: Tim-3 and its
role in regulating anti-tumor immunity. Immunol Rev. 276:97–111.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sánchez-Fueyo A, Tian J, Picarella D,
Domenig C, Zheng XX, Sabatos CA, Manlongat N, Bender O, Kamradt T,
Kuchroo VK, et al: Tim-3 inhibits T helper type 1-mediated auto-
and alloimmune responses and promotes immunological tolerance. Nat
Immunol. 4:1093–1101. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang SC, Wang M, Wu WB, Wang R, Cui J, Li
W, Li ZL, Li W and Wang SM: miR-22-3p inhibits arterial smooth
muscle cell proliferation and migration and neointimal hyperplasia
by targeting HMGB1 in arteriosclerosis obliterans. Cell Physiol
Biochem. 42:2492–2506. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Andrés V: Control of vascular cell
proliferation and migration by cyclin-dependent kinase signalling:
New perspectives and therapeutic potential. Cardiovasc Res.
63:11–21. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang Y, Qian X, Sun X, Lin C, Jing Y, Yao
Y, Ma Z, Kuai M, Lu Y, Kong X, et al: Liuwei Dihuang, a traditional
Chinese medicinal formula, inhibits proliferation and migration of
vascular smooth muscle cells via modulation of estrogen receptors.
Int J Mol Med. 42:31–40. 2018.PubMed/NCBI
|
|
28
|
Hao B, Xiao Y, Song F, Long X, Huang J,
Tian M, Deng S and Wu Q: Metformin-induced activation of AMPK
inhibits the proliferation and migration of human aortic smooth
muscle cells through upregulation of p53 and IFI16. Int J Mol Med.
41:1365–1376. 2018.PubMed/NCBI
|
|
29
|
Qiu MK, Wang SC, Tang Y, Pan C, Wang Y,
Wang SQ, Quan ZW and Ou JM: Tim-3 inhibits low-density
lipoprotein-induced atherogenic responses in human umbilical vein
endothelial cells. Oncotarget. 8:61001–61010. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ohira H, Tsutsui W and Fujioka Y: Are
Short Chain Fatty Acids in Gut Microbiota defensive players for
inflammation and atherosclerosis. J Atheroscler Thromb. 24:660–672.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pant S, Deshmukh A, Gurumurthy GS,
Pothineni NV, Watts TE, Romeo F and Mehta JL: Inflammation and
atherosclerosis-revisited. J Cardiovasc Pharmacol Ther. 19:170–178.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gliozzi M, Scicchitano M, Bosco F,
Musolino V, Carresi C, Scarano F, Maiuolo J, Nucera S, Maretta A,
Paone S, et al: Modulation of nitric oxide synthases by oxidized
LDLs: Role in vascular inflammation and atherosclerosis
development. Int J Mol Sci. 20(pii): E32942019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pan JX: LncRNA H19 promotes
atherosclerosis by regulating MAPK and NF-κB signaling pathway. Eur
Rev Med Pharmacol Sci. 21:322–328. 2017.PubMed/NCBI
|
|
34
|
Ge H, Tang H, Liang Y, Wu J, Yang Q, Zeng
L and Ma Z: Rhein attenuates inflammation through inhibition of
NF-κB and NALP3 inflammasome in vivo and in vitro. Drug Des Devel
Ther. 11:1663–1671. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lai JL, Liu YH, Liu C, Qi MP, Liu RN, Zhu
XF, Zhou QG, Chen YY, Guo AZ and Hu CM: Indirubin inhibits
LPS-induced inflammation via TLR4 abrogation mediated by the NF-κB
and MAPK signaling pathways. Inflammation. 40:1–12. 2017.
View Article : Google Scholar : PubMed/NCBI
|