Introduction
Schizophrenia is a serious mental disorder
characterized by affective and cognitive symptoms, and affects ~1%
of the population with an estimated heritability of 64–80%
(1). Genome-wide association
studies (GWASs) have revealed a large number of risk genes in
schizophrenia (2). The potential
interaction of these genes constitutes pivotal signaling pathways
and complex networks of regulation implicated with schizophrenia
(3).
Zinc finger protein 804A (ZNF804A), located
on chromosome 2q32.1, was the first schizophrenia susceptibility
gene identified by GWASs (4).
Subsequent larger GWASs confirmed the association of ZNF804A
with schizophrenia, and extended its association to other
psychiatric diagnoses, including bipolar disorder (5,6).
Although ZNF804A regulates a number of genes and pathways
associated with schizophrenia (7),
the upstream regulation of ZNF804A, as well as its
implication in schizophrenia, have not been fully investigated.
MicroRNAs (miRNAs/miRs), a class of endogenous
non-coding single-strand RNAs that are 9–23 nucleotides in length,
are of interest as key transcriptional regulators of ~1/2 of human
genes (8). Moreover, genetic
variants in the human MIR137 gene locus increase
schizophrenia risk and have genome-wide significance (9). It has been revealed that miR-137
downregulates the presynaptic target gene synaptotagmin-1 and
impairs synaptic plasticity in the hippocampus (10). Furthermore, ephrin B2 is a
validated target gene of miR-137, and the aberrant expression of
miR-137 in the peripheral blood of patients with schizophrenia
provides further evidence for its involvement in schizophrenia
etiology and diagnosis (11).
The present study revealed significantly different
expression profiles for miR-148b-3p in peripheral blood of patients
with first-onset schizophrenia compared with healthy controls.
Thus, the present study investigated the possible upstream
regulation of ZNF804A by miR-148b-3p, as an underlying
molecular mechanism in schizophrenia. Therefore, SH-SY5Y
neuroblastoma human cells were used in the present study. SH-SY5Y
is a thrice cloned subline of the SK-N-SH cell line that was
originally established from a bone marrow biopsy from a patient
with neuroblastoma in the 1970s, and has been widely used for
examining the cellular mechanisms underlying the etiology of
neuropsychiatric disorders, including schizophrenia and bipolar
disorder (12–14).
Materials and methods
Human peripheral blood collection and
reverse transcription-quantitative PCR (RT-qPCR)
The study was approved by the Medical Ethics
Committee of Xi'an Jiaotong University Health Science Center and
was in accordance with the Declaration of Helsinki. Informed
consent was obtained from all subjects in the Affiliated Hospitals
of Xi'an Jiaotong University (Xi'an, China) from May 2014 to April
2015. A total of 88 unrelated subjects were enrolled in the present
study, including 44 patients with schizophrenia and 44 healthy
controls, the details of whom were described previously (11). Peripheral blood samples from the
participants (5 ml) were collected in EDTA-coated tubes and were
then placed on ice. Total RNA was isolated within 2 h of collection
using TRIzol® (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions and quantified
using a NanoDrop 2000 (Thermo Fisher Scientific, Inc.). The first
strand of cDNA for miRNA was synthesized using Mir-X miRNA
First-Strand Synthesis Kit (Clontech, USA), and the first strand of
cDNA for mRNA was synthesized using RevertAid First Strand cDNA
Synthesis Kit (Thermo Fisher Scientific, Inc.). RT-qPCR was
performed using FastStart Universal SYBR Green Master (Rox; Roche
Diagnostics) and an Mx3005P qPCR system (Agilent Technologies,
Inc.) according to the following procedure: 95°C for 10 min;
followed by 40 cycles of 95°C for 10 sec, 60°C for 30 sec, 72°C for
30 sec; and a final dissociation curve analysis stage. Gene
expression and miRNA expression were normalized to GAPDH and U6
small nuclear RNA expression, respectively. Each sample was tested
in triplicate, and all data were analyzed using the
2−ΔΔCq method (15).
Primer information for genes and miRNAs is summarized in Table I.
 | Table I.Primer information of miRNA and genes
for RT-qPCR. |
Table I.
Primer information of miRNA and genes
for RT-qPCR.
| Gene name | Accession no. | Primer for
RT-qPCR | Location for the
first nucleotide of the primera | Sequence
(5′→3′) | Annealing
temperature (°C) |
|---|
|
Hsa-miR-148b-3p | NR_029894 | Forward | Chr12:54337278 |
TCAGTGCATCACAGAACTTTGT | 60 |
| ZNF804A | NM_194250 | Forward | Chr2:184933670 |
GAAAACAGGAAAAGGCACTCC | 60 |
|
|
| Reverse | Chr2:184935847 |
CACAACAACTCGTTGGGAAAT |
|
| COMT | NM_000754 | Forward | Chr22:19962760 |
CATTGACACCTACTGCGAGC | 59 |
|
|
| Reverse | Chr22:19963678 |
TCTCGATGGTGATGAGCCTC |
|
| PRSS16 | NM_005865 | Forward | Chr6:27255074 |
CTTGGACATGGCACCTGAG | 58 |
|
|
| Reverse | Chr6:27255274 |
CTCTCCTTTGCCAGCTTGAG |
|
| GAPDH | NM_002046 | Forward |
Chr12:6,537,347 |
CCAAGGTCATCCATGACAACT | 60 |
|
|
| Reverse |
Chr12:6,537,683 |
CAGGGATGATGTTCTGGAGAG |
|
| U6 snRNA | NR_004394 | Forward |
Chr15:67,840,042 |
CTCGCTTCGGCAGCACA | 60 |
|
|
| Reverse |
Chr15:67,839,949 |
AACGCTTCACGAATTTGCGT |
|
Bioinformatics prediction
Human miR-148b-3p sequences were obtained from
miRBase 22.1 (mirbase.org/). ZNF804A mRNA
3′-untranslated region (3′-UTR) sequences
(Chr2:184,939,027-184,939,487, NM_194250) were obtained from the
UCSC genome browser Human GRCh38/hg38 (http://genome.ucsc.edu/cgi-bin/hgTracks). TargetScan
6.2 (http://www.targetscan.org/), DIANA TOOLS
5.0 (http://diana.imis.athena-innovation.gr/Diana
Tools/index.php) and miRanda (August 2010 Release) (http://www.microrna.org) were used to predict
miR-148b-3p and ZNF804A interaction, which was based on the
presence of binding sites in the seed region, the efficacy of
targeting and the probability of conserved targeting (16). A Venn diagram of individual
prediction among the three software programs was calculated and
drawn using Bioinformatics & Evolutionary Genomics online
software 1.0 (http://bioinformatics.psb.ugent.be/webtools/Venn/).
Recombinant construction and
site-directed mutagenesis
A 461-bp-long 3′-UTR fragment of the ZNF804A
gene, which contained the putative miR-148b-3p binding site, was
synthesized by Chang Jing Bio-Tech, Ltd. The sequence was inserted
into a pmirGLO Dual-Luciferase miRNA Target Expression vector
(Promega Corporation) downstream of the firefly luciferase gene,
between the SacI and XbaI sites. The constructed
wild-type vectors were validated by restriction enzyme digestion
and Sanger sequencing. For restriction enzyme digestion, the
following reaction components were mixed and maintained at 37°C for
4 h: 10 µl 10X M buffer, 1 µl SacI, 1 µl XbaI (cat. Nos. 1078A and
1093A, respectively; both Takara Biotechnology Co., Ltd.) and 1 µg
constructed wild-type vector. The total volume was adjusted to 20
µl by adding ddH2O. The digestion products were
separated on a 1% agarose gel using DL2000 (cat. no. 3427; Takara
Biotechnology Co., Ltd.) as a DNA marker. For Sanger sequencing,
the forward primer (5′-GACGAGGTGCCTAAAGGACT-3′), constructed
vectors, and BigDye™ terminator v3.1 (cat. no. 4336913; Applied
Biosystems; Thermo Fisher Scientific, Inc.) were used for
sequencing reaction to generate fluorescently labeled DNA fragments
with different sizes. Subsequently, the DNA fragments were
separated by capillary electrophoresis, during which the
fluorescent labels were excited by laser and transformed to
chromatogram. The Sanger sequencing results were identified to be
consistent with the GRCh38/hg38 reference sequence from the UCSC
genome browser (genome.ucsc.edu/cgi-bin/hgTracks) and the NCBI
database (ncbi.nlm.nih.gov/gene/91752). The mutated
ZNF804A 3′-UTR sequence, containing all six nucleotides in
the putative seed-pair region, was synthesized according to A-T,
C-G substitution criteria (17).
The mutated product was constructed by inserting the mutant
sequence into the pmirGLO Dual-Luciferase miRNA Target Expression
vector, and it was validated by restriction enzyme digestion and
Sanger sequencing.
Cell culture
The SH-SY5Y cell line was purchased from American
Type Culture Collection (cat. no. CRL-2266), and has been
authenticated by short tandem repeat profiling. Another
neuroblastoma cell line, SK-N-BE(2), and 293T cells were obtained
from the Cell Bank of the Chinese Academy of Sciences, and were
authenticated. All cells were maintained in a humidified incubator
at 37°C with 5% CO2. 293T cells were cultured in DMEM
with high glucose (HyClone; GE Healthcare Life Sciences)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.),
100 U/ml penicillin and 100 µg/ml streptomycin
(Penicillin-Streptomycin solution; HyClone; GE Healthcare Life
Sciences). SH-SY5Y and SK-N-BE(2) cells were cultured in a 1:1
mixture of DMEM:nutrient mixture and F-12 medium (DMEM/F-12)
containing 1.5 mM L-glutamine (HyClone; GE Healthcare Life
Sciences); the mix was supplemented with 10% FBS, 100 U/ml
penicillin and 100 µg/ml streptomycin.
Co-transfection and dual luciferase
reporter assays
293T cells were seeded on 48-well plates at a
density of 5×104/well and cultured at 37°C for 24 h
before transfection. A miR-148b-3p mimic
(5′-UCAGUGCAUCACAGAACUUUGU-3′), a mimic negative control (NC;
5′-UUCUCCGAACGUGUCACGUTT-3′), a miR-148b-3p inhibitor
(5′-ACAAAGUUCUGUGAUGCACUGA-3′) and an inhibitor NC
(5′-CAGUACUUUUGUGUAGUACAA-3′) were obtained from Chang Jing
Bio-Tech, Ltd. To evaluate the interaction between miR-148b-3p and
the ZNF804A 3′-UTR, combinations of 50 nM mimic or mimic NC
with 1 µg wild-type and mutant UTR vectors were co-transfected by
using Lipofectamine™ 2000 (Invitrogen; Thermo Fisher Scientific,
Inc.). At 24 and 48 h after transfection, cells were harvested. The
activity of the luciferase reporter gene was detected using the
Dual-Luciferase® Reporter Assay system (Promega
Corporation). Each sample was tested in triplicate, and the
relative luciferase activity was calculated as the ratio of firefly
to Renilla luciferase activity, which was normalized against
a blank control.
Cell transfection and RT-qPCR
SH-SY5Y cells were seeded on 12-well plates at a
density of 2.5×105/well and cultured at 37°C for 24 h,
followed by transfection with 50 nM miR-148b-3p mimic, 50 nM mimic
NC, 50 nM miR-148b-3p inhibitor or 50 nM inhibitor NC, by
Lipofectamine™ 2000. For comparison, blank and mock controls using
the sample volume of media (1 ml) and transfection reagent (2 µl)
were prepared. Each experimental group was transfected in
triplicate. After a 24-h transfection period, total RNA was
isolated using TRIzol® (Invitrogen; Thermo Fisher
Scientific, Inc.) and quantified with NanoDrop 2000 (Thermo Fisher
Scientific, Inc.). The same experimental procedure was performed in
SK-N-BE(2) to verify the results in SH-SY5Y. RT-qPCR for mRNA
expression was performed as aforementioned.
Western blotting
Following 48 h of transfection, proteins were
extracted from SH-SY5Y cells using RIPA lysis buffer (Beyotime
Institute of Biotechnology) containing the protease inhibitor
phenylmethanesulfonyl fluoride (Beyotime Institute of
Biotechnology). Total protein was quantified using a bicinchoninic
acid assay kit (Pierce; Thermo Fisher Scientific, Inc.). A total of
20 µg protein samples was loaded onto each lane of 8% SDS-PAGE gel,
followed by electrophoresis and transfer to PVDF membranes (EMD
Millipore). Membranes were probed with the following primary
antibodies overnight at 4°C: Rabbit polyclonal anti-ZNF804A
(1:1,000; cat. no. ab229346; Abcam), mouse monoclonal anti-GAPDH
(1:1,000, cat. no. TA-08; OriGene Technologies, Inc.) or rabbit
polyclonal anti-β-actin (1:1,000; cat. no. ab0035; Abways
Technology). Subsequently, membranes were incubated with
horseradish peroxidase-conjugated goat anti-mouse IgG (1:5,000;
cat. no. 31430; Thermo Fisher Scientific, Inc.) or goat anti-rabbit
IgG (1:5,000; cat. no. 31460; Thermo Fisher Scientific, Inc.)
secondary antibody at room temperature for 2 h. The target proteins
were detected by an enhanced chemiluminescence system (EMD
Millipore) and scanned using the GeneGnome XRQ system (Syngene)
with GeneTools analysis software 4.0; Syngene, and the densitometry
of specific western blotting bands was analyzed with GeneTools
analysis software. Protein expression was determined by comparing
the band intensity of ZNF804A to that of the internal control,
GAPDH. The same experimental procedure was performed in SK-N-BE(2)
to verify the results in SH-SY5Y. Each experiment was performed in
triplicate.
Ad-ZNF804A construction and cell
infection
The human ZNF804A gene fragment was amplified
by PCR and cloned into the adenoviral vector pHBAd-EF1-MCS-GFP
(Hanbio Biotechnology Co., Ltd.). The forward primer,
5′-GTGACCGGCGCCTACGAATTCGCCACCATGGAGTGTTACTACATTGT-3′ and reverse
primer, 5′-TCGGGATCCCGCCCGGGGGAAGAGAGGTTGCAAAGGGA-3′ as well as
human genome DNA template were used to amplify the ZNF804A
gene fragment using KOD Plus (Toyobo Life Science) according to the
following procedure: Pre-denatured at 95°C for 5 min; then 27
cycles of 94°C for 20 sec, 55°C for 20 sec and 72°C for 90 sec; and
final extension at 72°C for 10 min. Subsequently, the recombinant
plasmid and pHBAd-BHG plasmid (Hanbio Biotechnology Co., Ltd.) were
co-transfected into 293T cells using LipoFiter™ (Hanbio
Biotechnology Co., Ltd.) to generate recombinant ZNF804A
adenovirus, which was named ‘Ad-ZNF804A’. Ad-green
fluorescent protein was used as a NC. SH-SY5Y cells were
pre-transfected with miR-148b-3p mimics (50 nM), and then infected
with Ad-ZNF804A and Ad-NC for 24 h. The multiplicity of
infection (MOI) was 50, and the blank group was used as a control.
Total RNA was harvested using TRIzol® (Invitrogen;
Thermo Fisher Scientific, Inc.) after 48 h, and RT-qPCR for gene
expression was performed as aforementioned.
Statistical analysis
Data are presented as the mean ± SEM. Statistically
significant differences among groups were detected with one-way
ANOVA. Fisher's Least Significant Difference test or Tukey's test
were carried out for post hoc analyses. Student's t-tests were used
for comparisons between two groups that fitted normal distribution
and had a homogeneity of variance. For data that did not fit normal
distribution and had a homogeneity of variance, a non-parametric
test (Mann Whitney test) was performed. Correlation analysis was
performed using Pearson's correlation. Data analyses were performed
using GraphPad Prism 5 software (GraphPad Prism Software, Inc.) or
SPSS software version 18.0 (IBM Corp.). P<0.05 was considered to
indicate a statistically significant difference.
Results
Bioinformatic prediction of ZNF804A as
a target of miR-148b-3p
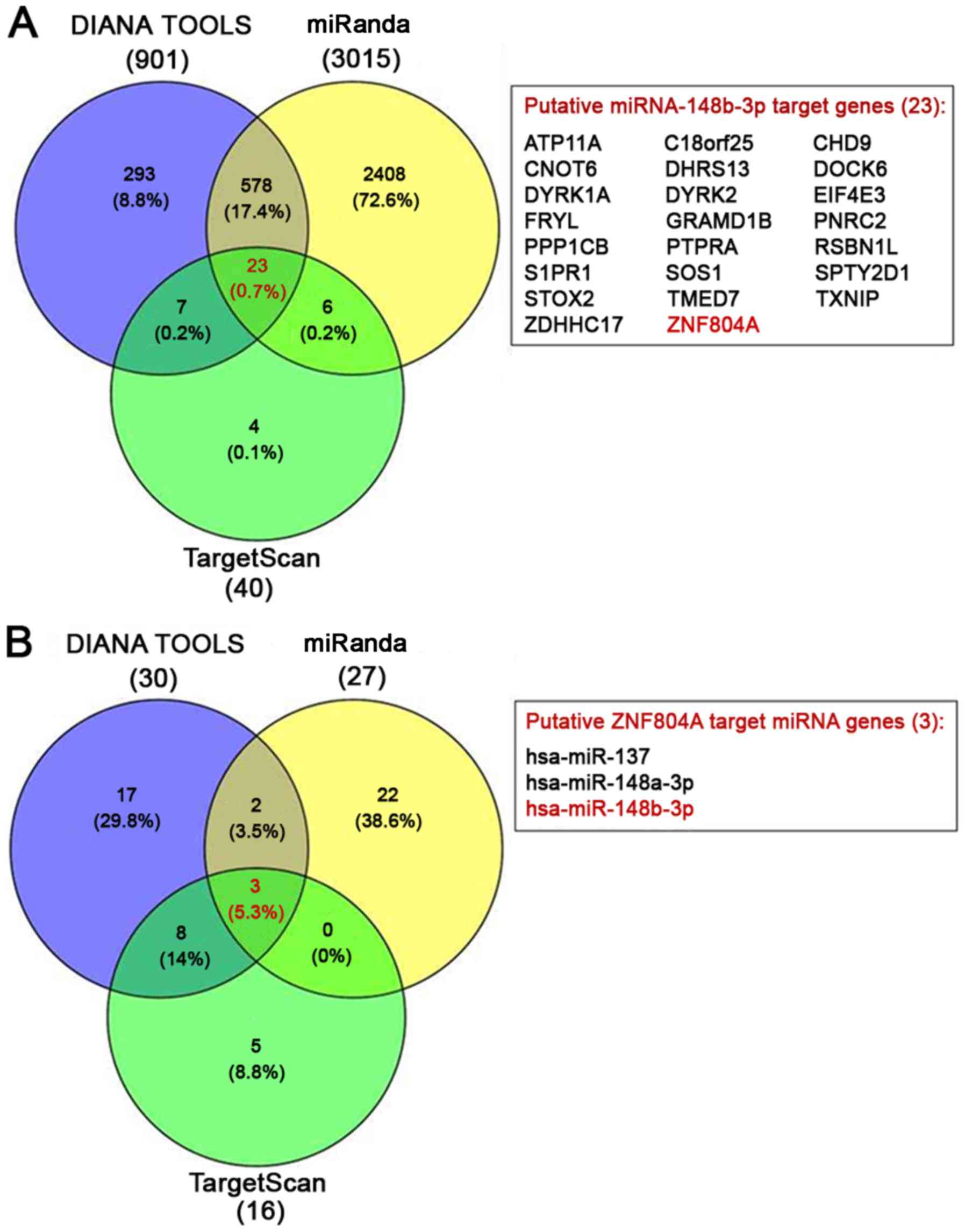
TargetScan, DIANA TOOLS and miRanda predicted 40,
901 and 3,015 putative target genes of miR-148b-3p, respectively.
ZNF804A was one of 23 shared target genes identified by all
prediction tools (Fig. 1A).
Furthermore, 16, 30 and 27 miRNAs that target the ZNF804A
gene were predicted by TargetScan, DIANA TOOLS and miRanda,
respectively. Moreover, miR-148b-3p was one of three shared miRNAs
identified by all prediction tools (Fig. 1B). It was demonstrated that all
three databases predicted that miR-148b-3p interacted with the
ZNF804A 3′-UTR from nucleotides 198–204. The binding site
sequence of the ZNF804A 3′-UTR is broadly conserved among
vertebrates, such as humans, chimpanzees, rhesus monkeys, rodents,
rabbits, pigs, cats, elephants and chickens, suggesting that it has
important roles in the regulation of gene function (genome.ucsc.edu/cgi-bin/hgTracks?db=hg38&lastVirtModeType=default&lastVirtModeExtraState=&virtModeType=default&virtMode=0&nonVirtPosition=&position=chr2%3A184939027%2D184939487&hgsid=820743199_5AAECt1MXtmem7EgvQiIixlwuapI).
miR-148b-3p and ZNF804A have
aberrantly reversed expression in the peripheral blood of patients
with schizophrenia
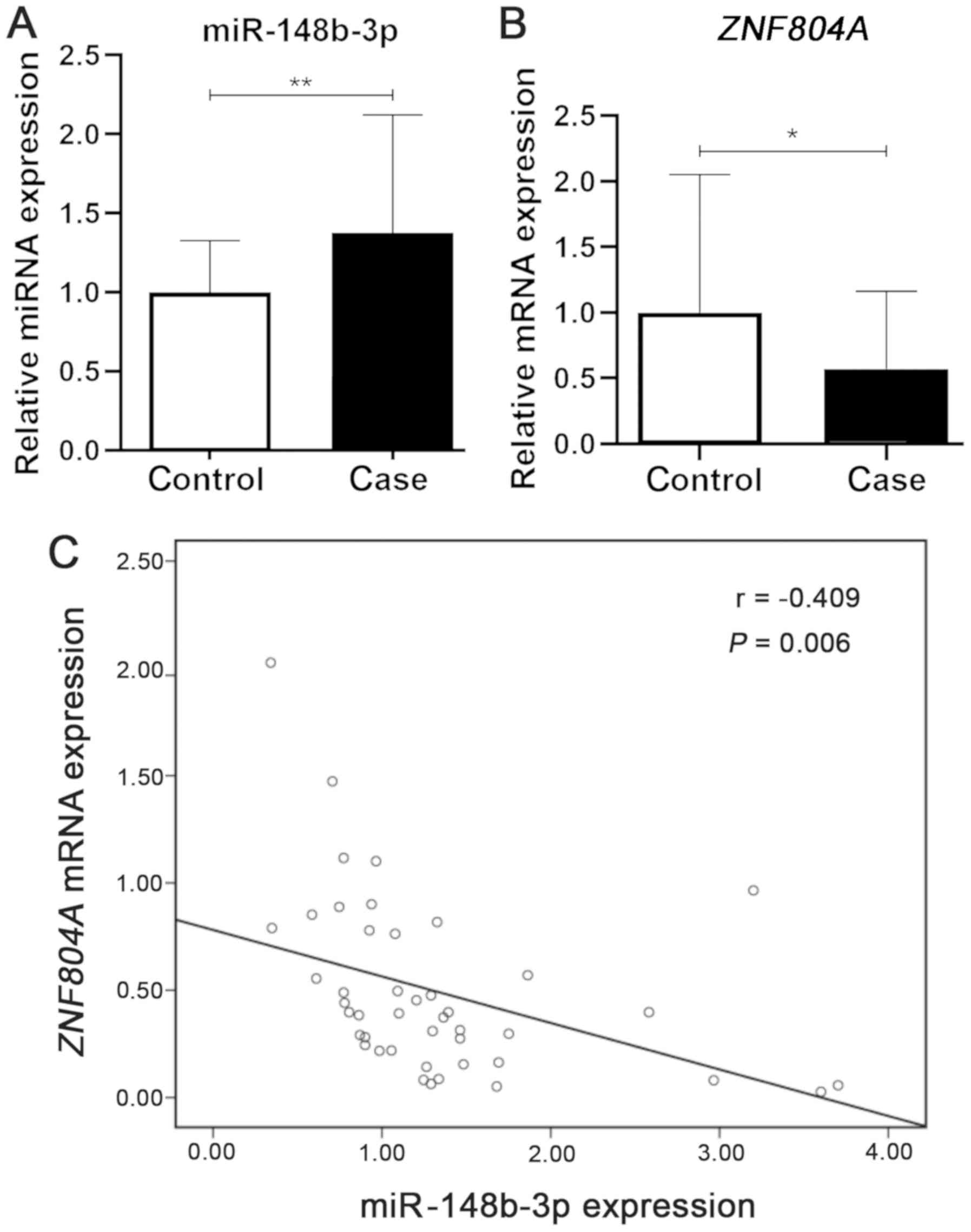
It was determined that the expression of miR-148b-3p
was 1.35-fold higher in the peripheral blood of patients with
schizophrenia compared with healthy controls (P=0.0051; Fig. 2A). Furthermore, the expression of
ZNF804A was significantly decreased in patients with
schizophrenia compared with healthy controls (P=0.0218; Fig. 2B). In addition, a correlation
analysis between miR-148b-3p and ZNF804A was performed and a
moderate negative correlation was identified (r=−0.409; P=0.006;
Fig. 2C).
Biological verification of miR-148b-3p
directly targeting ZNF804A
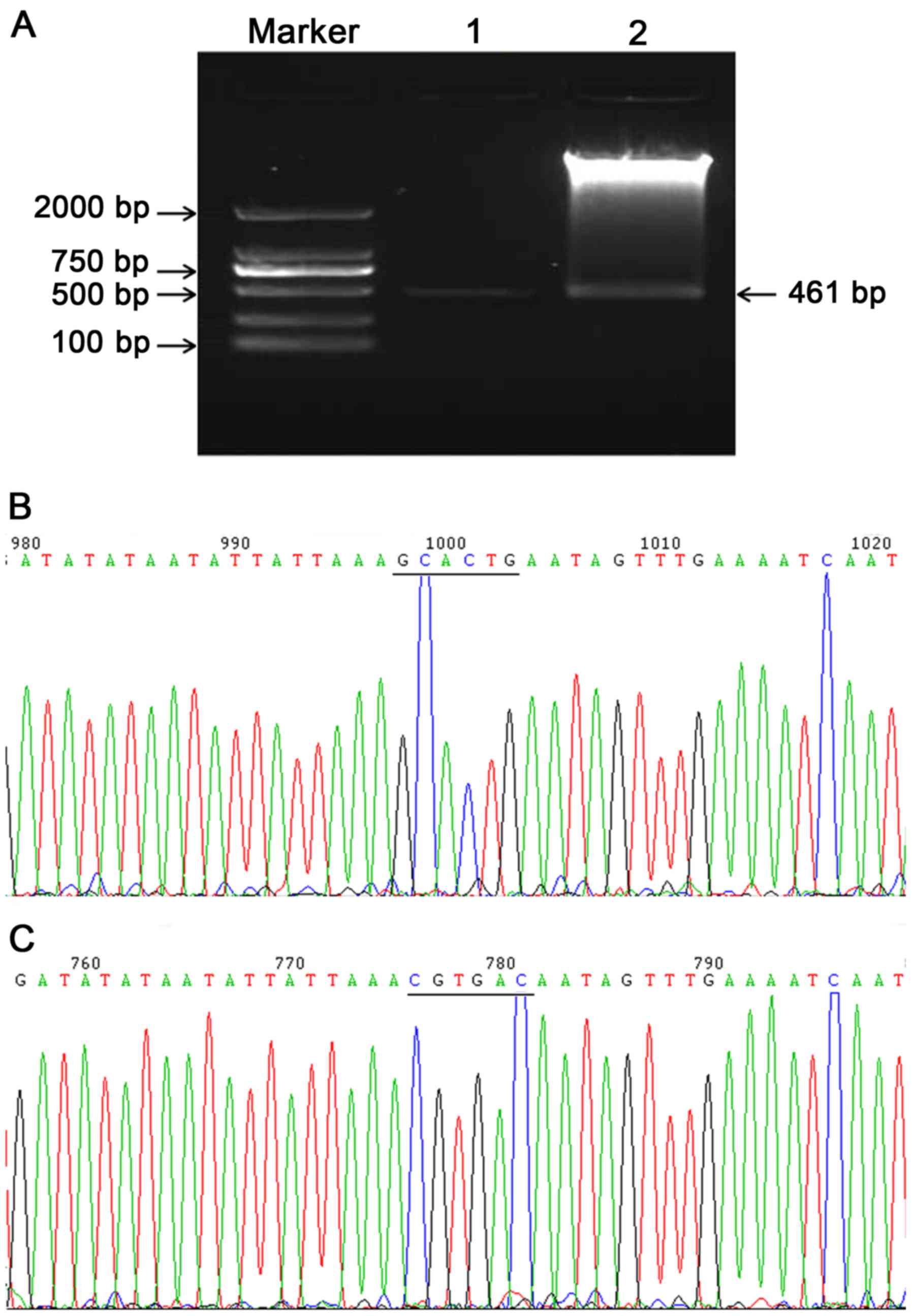
PmirGLO reconstructed vectors were generated and
were then assessed by restriction enzyme digestion and Sanger
sequencing. For this, one product of restriction enzyme digestion
was the pmirGLO vector, which was 7,350 bp, and the other product
was 461 bp, which was the same size as the PCR product before
cloning into the vector (Fig. 3A).
The sequencing results of the amplified fragments in the wild-type
construct were consistent with the GRCh38/hg38 reference sequences
from the UCSC genome browser and the NCBI database (Fig. 3B). Furthermore, the sequence in the
binding sites was mutated according to the A-T, C-G substitution
criterion (Fig. 3C).
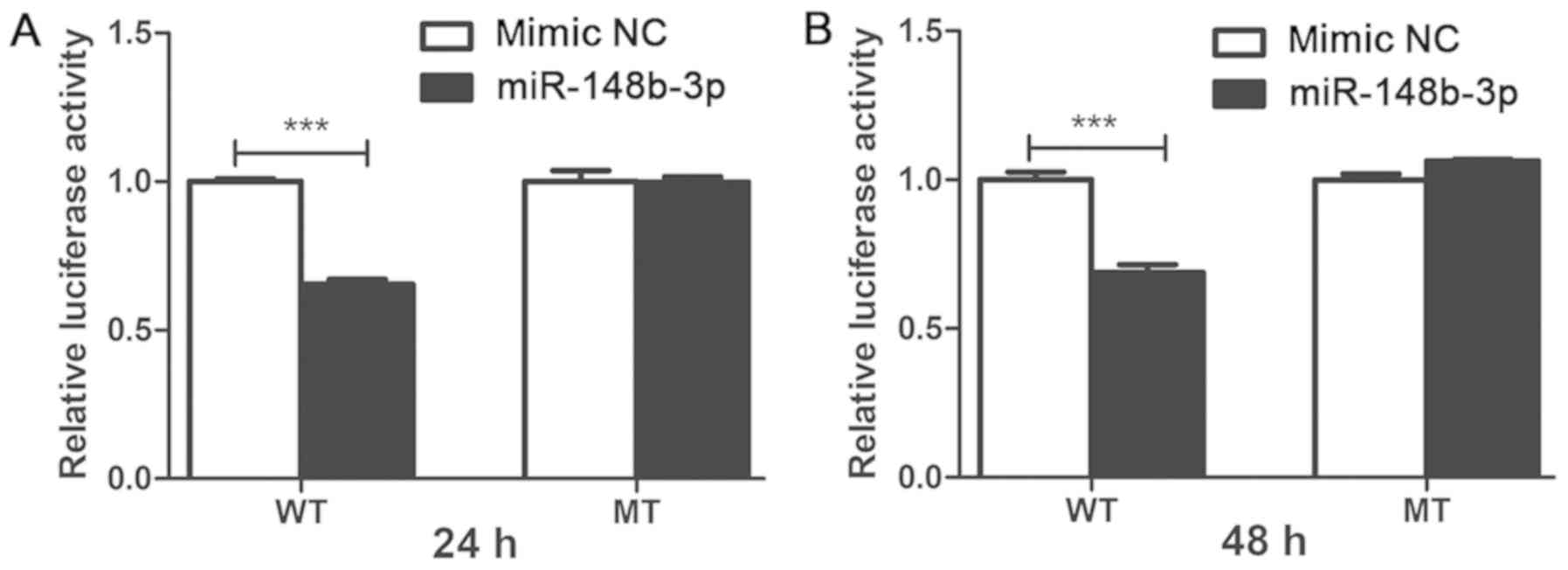
The present results indicated that the luciferase
activity in 293T cells co-transfected with the miR-148b-3p mimic
was significantly decreased after 24 h (P<0.001; Fig. 4A) and 48 h (P<0.001; Fig. 4B) compared with the NC group.
However, the luciferase activity did not reveal a significant
difference for the mutant group with miR-148b-3p mimic compared
with the NC group, thus suggesting that miR-148b-3p directly binds
to the ZNF804A 3′-UTR and inhibits its expression.
miR-148b-3p inhibits the endogenous
expression of ZNF804A
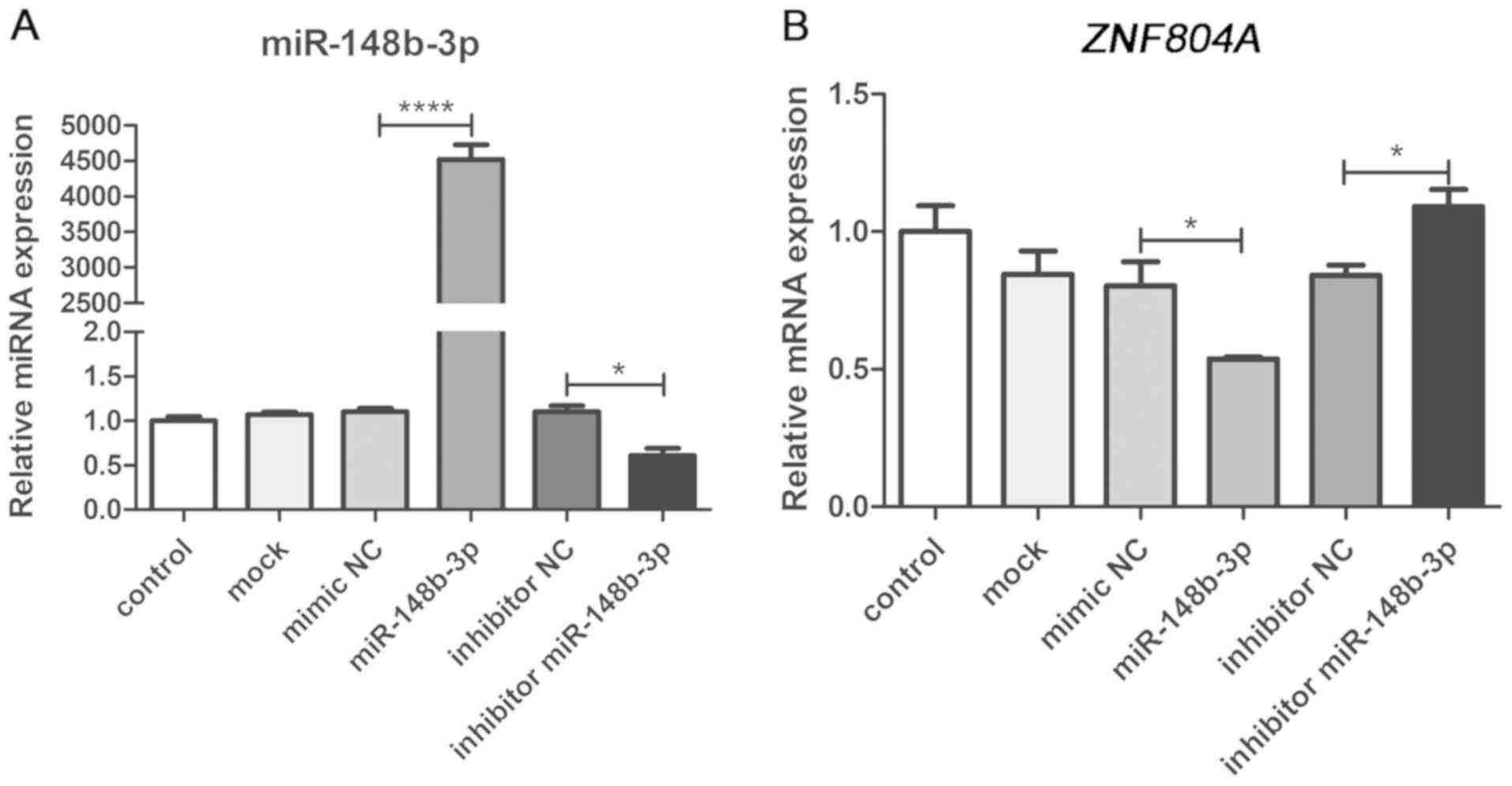
The results of RT-qPCR experiments revealed that the
miR-148b-3p mimic significantly increased the expression of
miR-148b-3p compared with the NC, mock or control groups, while the
miR-148b-3p inhibitor significantly decreased miR-148b-3p
expression. These results demonstrated the transfection efficiency
with the successful gain or loss of miR-148b-3p function in SH-SY5Y
cells (Fig. 5A). Moreover,
ZNF804A expression was significantly decreased in the
miR-148b-3p mimic group compared with the NC group (P=0.048). In
addition, ZNF804A expression was increased significantly in
the group transfected with the miR-148b-3p inhibitor compared with
the inhibitor NC group (P=0.044; Fig.
5B). Therefore, the present results indicated that endogenous
expression of ZNF804A may be regulated by miR-148b-3p at the
mRNA level.
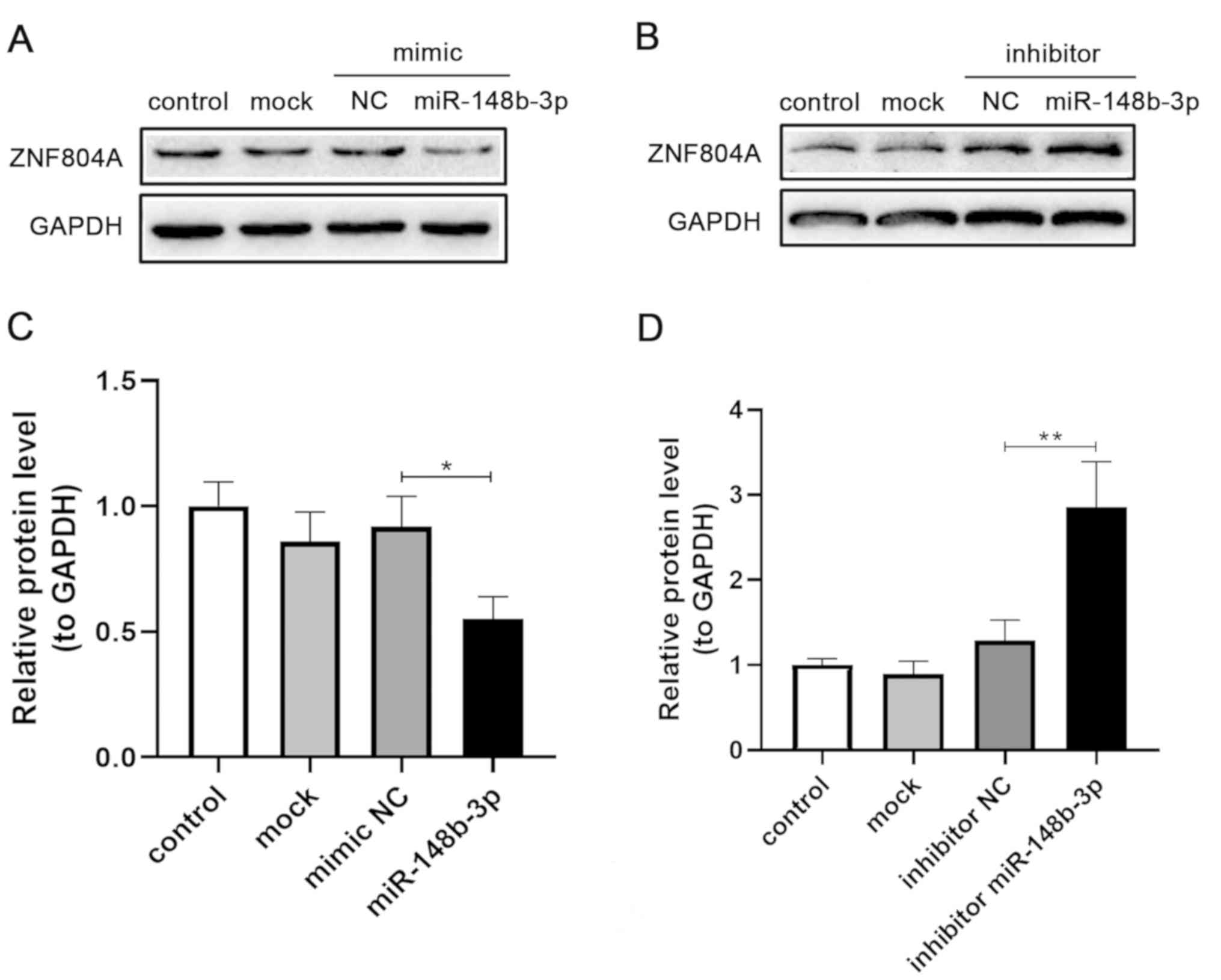
Western blotting results indicated that the
miR-148b-3p mimic significantly decreased ZNF804A protein
expression compared with the NC (P=0.013), mock or control groups
(Fig. 6A and C). However,
treatment with the miR-148b-3p inhibitor significantly increased
ZNF804A protein expression (P=0.001; Fig. 6B and D). Collectively, the present
results indicated that the endogenous expression of ZNF804A
was downregulated by miR-148b-3p at the protein level.
miR-148b-3p regulates the
schizophrenia susceptibility genes catechol-O-methyltransferase
(COMT) and serine protease 16 (PRSS16) by targeting ZNF804A
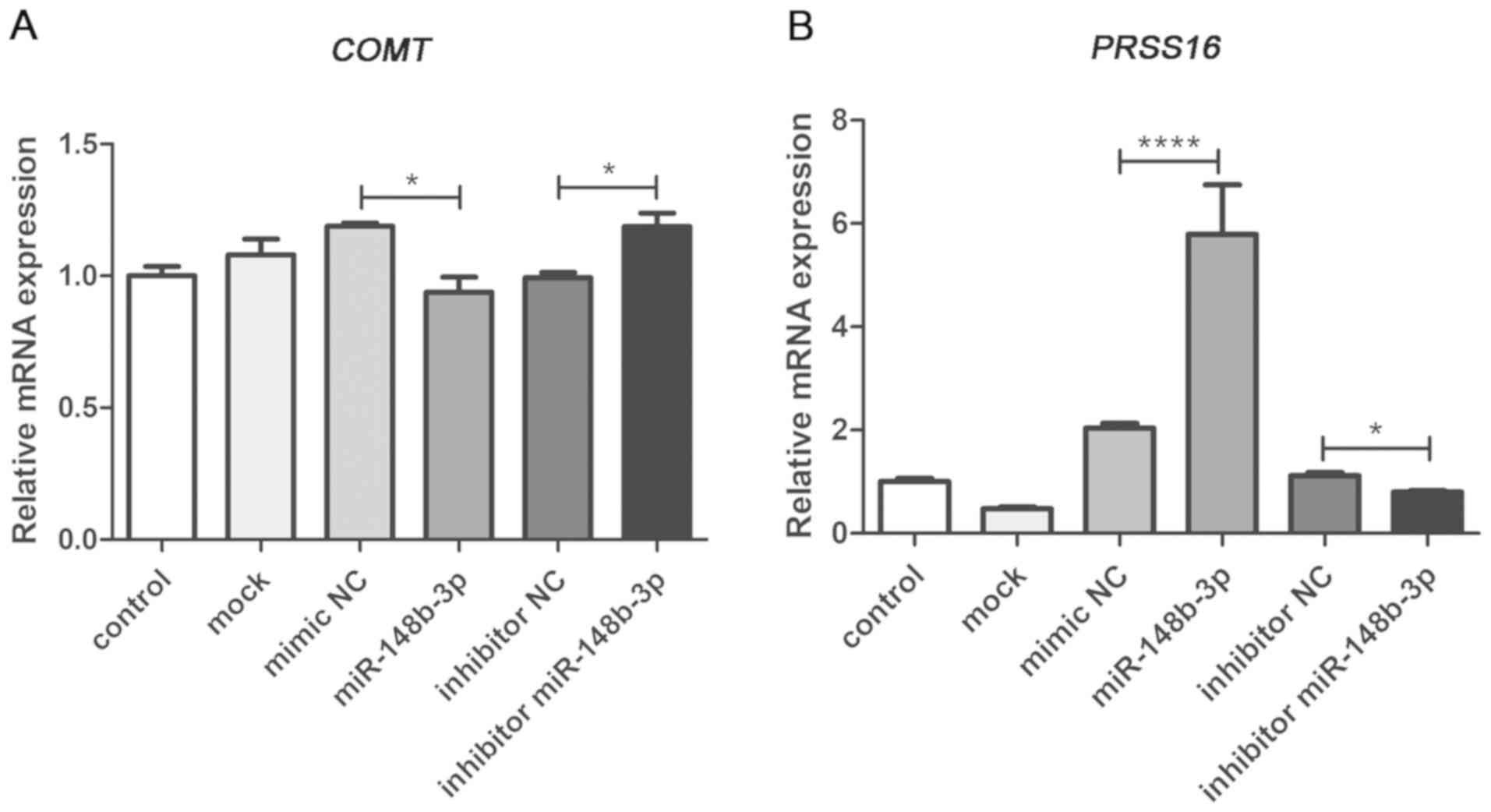
SH-SY5Y cells were transfected with the miR-148b-3p
mimic, mimic NC, miR-148b-3p inhibitor and inhibitor NC, to
identify whether COMT and PRSS16 are regulated by
miR-148b-3p. It was demonstrated that transfection with the
miR-148b-3p mimic induced a significant decrease in COMT
compared with the NC (P=0.025), mock or control levels, and
COMT expression was increased significantly in the
miR-148b-3p inhibitor group compared with the inhibitor NC group
(P=0.040; Fig. 7A). Furthermore,
transfection with the miR-148b-3p mimic significantly increased the
mRNA expression of PRSS16 compared with the NC
(P<0.0001), mock or control levels, and PRSS16 expression
was significantly decreased in the miR-148b-3p inhibitor group
compared with the inhibitor NC group (P=0.047; Fig. 7B). Moreover, western blotting
results of the quantification of the protein expression of COMT
were consistent with the RT-qPCR data (Fig. S1).
Similar experiments were performed in another
neuroblastoma cell line, SK-N-BE(2). It was revealed that the
results obtained in the SK-N-BE(2) cell line (Fig. S2) were consistent with those
generated in the SH-SY5Y cell line.
It has been previously reported that COMT and
PRSS16 are regulated by ZNF804A (7). To assess whether miR-148b-3p
regulates COMT and PRSS16 by targeting the
ZNF804A gene, ZNF804A expression was rescued in
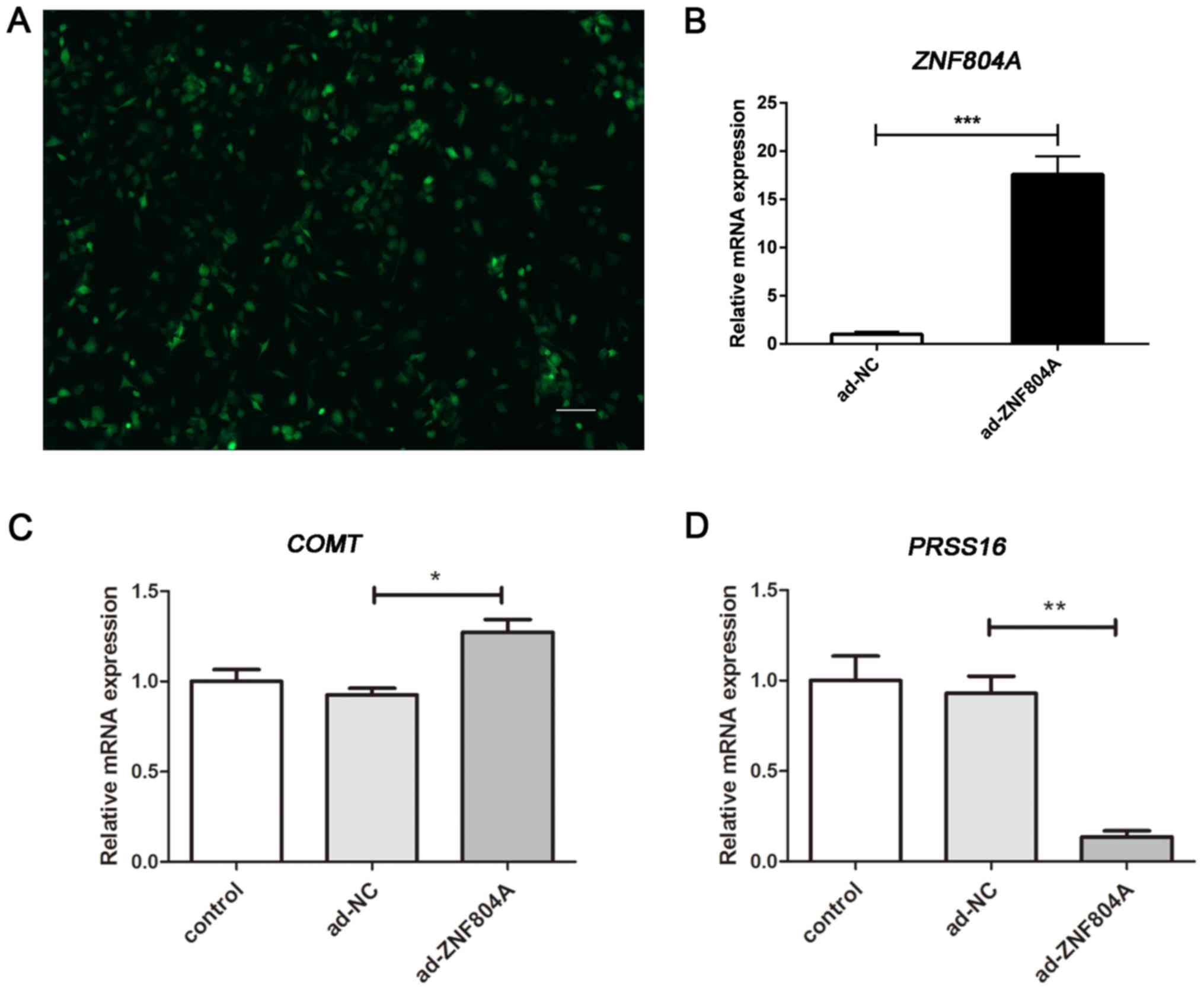
miR-148b-3p-expressing SH-SY5Y cells using an Ad-ZNF804A
vector. It was revealed that the infection efficiency was as high
as 90% when the MOI of adenovirus was 50 (Fig. 8A). Moreover, the mRNA expression
level of ZNF804A revealed that the transfections were
successful (P<0.001; Fig. 8B).
The results of the gene expression experiments demonstrated that
the transfection of the miR-148b-3p mimic, in addition to the
infection of the ZNF804A expression vector, rescued the
expression levels of COMT and PRSS16, which indicated
that gene regulation was mediated by miR-148b-3p (Fig. 8C and D). Therefore, the present
results suggested that miR-148b-3p regulated COMT and
PRSS16 by inhibiting ZNF804A.
Discussion
The present study revealed that the expression of
miR-148b-3p was significantly increased in the peripheral blood of
patients with first-onset schizophrenia compared with healthy
controls. Bioinformatics analyses and dual luciferase reporter
assays indicated that ZNF804A may be directly targeted by
miR-148b-3p. In human neuroblastoma cells transfected with a
miR-148b-3p mimic or a mimic NC, expression of ZNF804A at
both the mRNA and protein levels was influenced by miR-148b-3p
expression. Furthermore, two susceptibility genes, COMT and
PRSS16, were determined to be indirectly regulated by
miR-148b-3p via ZNF804A in SH-SY5Y cells. It has been
previously shown that dopamine (DA) dysfunction influences both
cortical and subcortical circuitry, thus facilitating different
symptomatology of schizophrenia (18). SH-SY5Y cells possess numerous
characteristics of DAergic neurons, such as the ability to
synthesize DA and express DA transporters (19,20).
Furthermore, under certain conditions SH-SY5Y cells can
differentiate into a more pronounced DAergic neuronal phenotype,
which present several of the characteristics of primary cultures of
neurons (20,21). Therefore, this cell line has been
widely used for schizophrenia research (19–21).
miR-148b-3p belongs to the miR-148/152 family, which
regulates the growth and development of healthy tissues (22). Differential expression between
members of the miR-148/152 family in patients with tumor or type 1
diabetes compared with healthy controls indicates their important
roles in the initiation and process of different diseases (22). In our previous study, it was
revealed that miR-148b-3p expression was upregulated in the
post-mortem dorsolateral prefrontal cortex of the schizophrenia
group compared with healthy controls (23). Moreover, miR-148b has been revealed
to affect neural stem cell proliferation, and their differentiation
into new neurons and astrocytes (24).
While the precise function of ZNF804A is
unknown in psychiatric disorders, the biological function and
molecular mechanisms of ZNF804A have been revealed by
previous studies (25,26). Furthermore, the neuronal function
and molecular mechanisms of ZNF804A have been revealed by
two landmark studies (25,26). Using neurons derived from human
neural progenitor cells (NPCs), human induced pluripotent stem
cells or primary rat cortical neurons, Deans et al (25) revealed that ZNF804A localizes to
synapses, and regulates neurite formation and dendritic spine
structure. In addition, Zhou et al (26) revealed that the rodent homologue of
ZNF804A (Zfp804a) is required for NPC proliferation
and neuronal migration, and also identified a novel role of
ZNF804A in modulating protein translational efficiency
during neurodevelopment.
COMT is a downstream gene of ZNF804A
that codes for catechol-O-methyltransferase, which degrades
extracellular catecholamines (27). DA is a catecholamine and is also a
principal neurotransmitter in the central nervous system (28). In pharmacology, COMT is a
biological target for the treatment of various central and
peripheral nervous system disorders, including Parkinson's disease,
depression and other DA-related diseases (29). The DAergic system has been
implicated in neuropsychological processes that are impaired in
patients with schizophrenia, and these impairments are correlated
with structural abnormalities observed in the brains of these
patients (30,31). In addition, DA is a powerful
activator of resting effector T cells by acting on its receptors on
immune effector cells (28). Both
altered immune functions and abnormal DAergic systems have been
observed in patients with schizophrenia (32). COMT, as a susceptibility
gene for schizophrenia, also represents an important target for the
treatment of the disease (33).
PRSS16 is located at 6p22-p21.3 on the chromosome near the
major histocompatibility complex class I region, and encodes a
serine protease that is abundantly expressed in the thymus; this
serine protease plays an important role in immune function
(34). Moreover, a GWAS indicated
that the single-nucleotide polymorphisms rs6932590 and rs13219354
in the PRSS16 loci are associated with schizophrenia risk
(35).
Thus, it is speculated that miR-148b-3p may affect
neurotransmitter synthesis and immune function by regulating the
expression levels of COMT and PRSS16, which are
downstream of ZNF804A, in schizophrenia. In the present
study, it was revealed that expression of miR-148b-3p was
aberrantly higher in the peripheral blood of patients with
schizophrenia compared with healthy controls. Moreover, this
expression pattern may reflect pathological processes in the brain,
as significant gene expression overlap is found between whole blood
and brain tissue (36).
Brain-derived exosomes and their miRNA cargo may be key players in
the cross-talk between the brain and the immune system in
psychiatric disorders (37).
Furthermore, the secretion of brain or immune cell exosomes is
partially regulated by neurotransmitters (38,39).
Exosomes or liposomes packaged with a specifically selected miRNA
cargo may readily cross the blood-brain barrier, and therefore
represent ideal carriers for nucleic acid-based drugs (40,41).
The present results indicated that the
miR-148b-3p/ZNF804A/COMT/PRSS16 pathway may play an
important role in the pathophysiology of schizophrenia, and that
miR-148b-3p may be a novel drug target in schizophrenia.
In conclusion, the present results indicated that
miR-148b-3p directly regulated the expression of ZNF804A,
and as a result influenced the expression levels of COMT and
PRSS16, which are downstream genes of ZNF804A.
Therefore, this regulatory pathway may affect the immune function
of patients with schizophrenia. It was also speculated that the
miR-148b-3p/ZNF804A/COMT/PRSS16 pathway may play an
important role in the pathophysiology of schizophrenia. In
addition, the present results may facilitate the development of
drug discovery and gene therapy for schizophrenia.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Dr Huan Zhang and Dr
Jin Wang (both Department of Psychiatry, the Affiliated Hospital of
Xi'an Jiaotong University, Xi'an, China) for help with the present
research.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 31371298 and
81660210), the Natural Science Basic Research Plan in Shaanxi
Province of China (program no. 2019JM-098) and the Opening Project
of the Key Laboratory of Shaanxi Province for Craniofacial
Precision Medicine Research, College of Stomatology, Xi'an Jiaotong
University (grant no. 2017LHM-KFKT007).
Availability of data and materials
The datasets generated and/or analyzed during the
current study are not publicly available due to confidentiality of
another study in our group, but are available from the
corresponding author on reasonable request.
Authors' contributions
SW, PW, RT, PY, XY, YL, QS, FN, JH, RZ, YT and JM
were involved in conception and design of the study, as well as
acquisition, analysis and interpretation of data. JM, YT, RZ and SW
drafted and revised the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Medical Ethics
Committee of Xi'an Jiaotong University Health Science Center, in
accordance with the Declaration of Helsinki. Informed consent was
obtained from all subjects.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yue WH, Wang HF, Sun LD, Tang FL, Liu ZH,
Zhang HX, Li WQ, Zhang YL, Zhang Y, Ma CC, et al: Genome-wide
association study identifies a susceptibility locus for
schizophrenia in Han Chinese at 11p11. 2. Nat Genet. 43:1228–1231.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schizophrenia Working Group of the
Psychiatric Genomics Consortium, . Biological insights from 108
schizophrenia-associated genetic loci. Nature. 511:421–427. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Misiak B, Stramecki F, Gawęda Ł, Prochwicz
K, Sąsiadek MM, Moustafa AA and Frydecka D: Interactions between
variation in candidate genes and environmental factors in the
etiology of schizophrenia and bipolar disorder: A systematic
review. Mol Neurobiol. 55:5075–5100. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
O'donovan MC, Craddock N, Norton N,
Williams H, Peirce T, Moskvina V, Nikolov I, Hamshere M, Carroll L,
Georgieva L, et al: Identification of loci associated with
schizophrenia by genome-wide association and follow-up. Nat Genet.
40:1053–1055. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Harrison PJ: ZNF804A: Insights from the
first genome-wide significant schizophrenia gene. Biol Psychiatry.
82:6–7. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Williams HJ, Norton N, Dwyer S, Moskvina
V, Nikolov I, Carroll L, Georgieva L, Williams NM, Morris DW, Quinn
EM, et al: Fine mapping of ZNF804A and genome-wide significant
evidence for its involvement in schizophrenia and bipolar disorder.
Mol Psychiatry. 16:429–441. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Girgenti MJ, LoTurco JJ and Maher BJ:
ZNF804a regulates expression of the schizophrenia-associated genes
PRSS16, COMT, PDE4B, and DRD2. PLoS One. 7:e324042012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Milanesi E, Maj C, Bocchio-Chiavetto L and
Maffioletti E: Nanomedicine in psychiatry: New therapeutic
opportunities from research on small RNAs. Drug Dev Res.
77:453–457. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schizophrenia Psychiatric Genome-Wide
Association Study (GWAS) Consortium, . Genome-wide association
study identifies five new schizophrenia loci. Nat Genet.
43:969–976. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Siegert S, Seo J, Kwon EJ, Rudenko A, Cho
S, Wang W, Flood Z, Martorell AJ, Ericsson M, Mungenast AE and Tsai
LH: The schizophrenia risk gene product miR-137 alters presynaptic
plasticity. Nat Neurosci. 18:1008–1016. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu S, Zhang R, Nie F, Wang X, Jiang C, Liu
M, Valenzuela RK, Liu W, Shi Y and Ma J: MicroRNA-137 inhibits
EFNB2 expression affected by a genetic variant and is expressed
aberrantly in peripheral blood of schizophrenia patients.
EBioMedicine. 12:133–142. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
D'Souza UM, Russ C, Tahir E, Mill J,
McGuffin P, Asherson PJ and Craig IW: Functional effects of a
tandem duplication polymorphism in the 5′flanking region of the
DRD4 gene. Biol Psychiatry. 56:691–697. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Warburton A, Breen G, Rujescu D, Bubb VJ
and Quinn JP: Characterization of a REST-regulated internal
promoter in the schizophrenia genome-wide associated gene MIR137.
Schizophr Bull. 41:698–707. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
O'Brien NL, Fiorentino A, Curtis D, Rayner
C, Petrosellini C, Al Eissa M, Bass NJ, McQuillin A and Sharp SI:
Rare variant analysis in multiply affected families, association
studies and functional analysis suggest a role for the ITGBeta4
gene in schizophrenia and bipolar disorder. Schizophr Res.
199:181–188. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yuan JS, Reed A, Chen F and Stewart CN Jr:
Statistical analysis of real-time PCR data. BMC Bioinformatics.
7:852006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chang TC and Mendell JT: MicroRNAs in
vertebrate physiology and human disease. Annu Rev Genomics Hum
Genet. 8:215–239. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guillin O, Abi-Dargham A and Laruelle M:
Neurobiology of dopamine in schizophrenia. Int Rev Neurobiol.
78:1–39. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li B, Chen R, Chen L, Qiu P, Ai X, Huang
E, Huang W, Chen C, Liu C, Lin Z, et al: Effects of DDIT4 in
methamphetamine-induced autophagy and apoptosis in dopaminergic
neurons. Mol Neurobiol. 54:1642–1660. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Korecka JA, van Kesteren RE, Blaas E,
Spitzer SO, Kamstra JH, Smit AB, Swaab DF, Verhaagen J and Bossers
K: Phenotypic characterization of retinoic acid differentiated
SH-SY5Y cells by transcriptional profiling. PLoS One. 8:e638622013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xie HR, Hu LS and Li GY: SH-SY5Y human
neuroblastoma cell line: In vitro cell model of dopaminergic
neurons in Parkinson's disease. Chin Med J (Engl). 123:1086–1092.
2010.PubMed/NCBI
|
|
22
|
Chen Y, Song YX and Wang ZN: The
microRNA-148/152 family: Multi-faceted players. Mol Cancer.
12:432013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Santarelli DM, Beveridge NJ, Tooney PA and
Cairns MJ: Upregulation of dicer and microRNA expression in the
dorsolateral prefrontal cortex Brodmann area 46 in schizophrenia.
Biol Psychiatry. 69:180–187. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang J, Chen T and Shan G: miR-148b
regulates proliferation and differentiation of neural stem cells
via Wnt/β-catenin signaling in rat ischemic stroke model. Front
Cell Neurosci. 11:3292017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Deans PJM, Raval P, Sellers KJ, Gatford
NJF, Halai S, Duarte RRR, Shum C, Warre-Cornish K, Kaplun VE, Cocks
G, et al: Psychosis risk candidate ZNF804A localizes to synapses
and regulates neurite formation and dendritic spine structure. Biol
Psychiatry. 82:49–61. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhou Y, Dong F, Lanz TA, Reinhart V, Li M,
Liu L, Zou J, Xi HS and Mao Y: Interactome analysis reveals
ZNF804A, a schizophrenia risk gene, as a novel component of protein
translational machinery critical for embryonic neurodevelopment.
Mol Psychiatry. 23:952–962. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen J, Lipska BK, Halim N, Ma QD,
Matsumoto M, Melhem S, Kolachana BS, Hyde TM, Herman MM, Apud J, et
al: Functional analysis of genetic variation in
catechol-O-methyltransferase (COMT): Effects on mRNA, protein, and
enzyme activity in postmortem human brain. Am J Hum Genet.
75:807–821. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Levite M: Dopamine and T cells: Dopamine
receptors and potent effects on T cells, dopamine production in T
cells, and abnormalities in the dopaminergic system in T cells in
autoimmune, neurological and psychiatric diseases. Acta Physiol
(Oxf). 216:42–89. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kiss LE and Soares-da-Silva P: Medicinal
chemistry of catechol O-methyltransferase (COMT) inhibitors and
their therapeutic utility. J Med Chem. 57:8692–8717. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Antonova E, Kumari V, Morris R, Halari R,
Anilkumar A, Mehrotra R and Sharma T: The relationship of
structural alterations to cognitive deficits in schizophrenia: A
voxel-based morphometry study. Biol Psychiatry. 58:457–467. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gao S, Cheng J, Li G, Sun T, Xu Y, Wang Y,
Du X, Xu G and Duan S: Catechol-O-methyltransferase gene promoter
methylation as a peripheral biomarker in male schizophrenia. Eur
Psychiatry. 44:39–46. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Basu S and Dasgupta PS: Dopamine, a
neurotransmitter, influences the immune system. J Neuroimmunol.
102:113–124. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Egan MF, Goldberg TE, Kolachana BS,
Callicott JH, Mazzanti CM, Straub RE, Goldman D and Weinberger DR:
Effect of COMT Val108/158 Met genotype on frontal lobe function and
risk for schizophrenia. Proc Natl Acad Sci USA. 98:6917–6922. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kitazawa M, Ohnuma T, Takebayashi Y,
Shibata N, Baba H, Ohi K, Yasuda Y, Nakamura Y, Aleksic B, Yoshimi
A, et al: No associations found between the genes situated at 6p22.
1, HIST1H2BJ, PRSS16, and PGBD1 in Japanese patients diagnosed with
schizophrenia. Am J Med Genet B Neuropsychiatr Genet 159B. 456–464.
2012. View Article : Google Scholar
|
|
35
|
Stefansson H, Ophoff RA, Steinberg S,
Andreassen OA, Cichon S, Rujescu D, Werge T, Pietiläinen OP, Mors
O, Mortensen PB, et al: Common variants conferring risk of
schizophrenia. Nature. 460:744–747. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Harris LW, Pietsch S, Cheng TM, Schwarz E,
Guest PC and Bahn S: Comparison of peripheral and central
schizophrenia biomarker profiles. PLoS One. 7:e463682012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ander BP, Barger N, Stamova B, Sharp FR
and Schumann CM: Atypical miRNA expression in temporal cortex
associated with dysregulation of immune, cell cycle, and other
pathways in autism spectrum disorders. Mol Autism. 6:372015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chivet M, Javalet C, Laulagnier K, Blot B,
Hemming FJ and Sadoul R: Exosomes secreted by cortical neurons upon
glutamatergic synapse activation specifically interact with
neurons. J Extracell Vesicles. 3:247222014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Glebov K, Löchner M, Jabs R, Lau T, Merkel
O, Schloss P, Steinhäuser C and Walter J: Serotonin stimulates
secretion of exosomes from microglia cells. Glia. 63:626–634. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wood MJ, O'Loughlin AJ and Samira L:
Exosomes and the blood-brain barrier: Implications for neurological
diseases. Ther Deliv. 2:1095–1099. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ohno S and Kuroda M: Exosome-mediated
targeted delivery of miRNAs. Methods Mol Biol. 1448:261–270. 2016.
View Article : Google Scholar : PubMed/NCBI
|