Introduction
Inflammatory bowel diseases (IBDs), such as Crohn's
disease (CD) and ulcerative colitis (UC), are chronic inflammatory
disorders of the gastrointestinal tract (1). Several factors have been identified
that contribute to the disease pathogenesis, such as genetic
factors, host immunity and environmental factors, including the
intestinal microbiota (2).
However, the precise etiology of IBD remains unclear, and
treatments for IBD have limited efficacy; therefore, it is
necessary to explore novel, effective and safe therapies to treat
IBD.
Smoking is a controversial factor in colitis
etiology that has been discussed for decades; although it has been
shown to exacerbate CD, clinical studies have suggested it may
reduce the severity of UC (3–5).
Although the specific mechanisms underlying these effects remain
unclear, a number of studies have proposed possible explanations
for the effect of smoking on immune system reactivity and gut
barrier integrity (6,7). As a potential beneficial compound,
nicotine has anti-inflammatory effects through the activation of
the nicotinic acetylcholine receptor (8,9), and
several clinical studies suggest that nicotine could improve gut
motility in smokers (10,11). The α7 subunit of the nicotinic
acetylcholine receptor (α7nAChR) has been reported to play a
crucial role in the responsiveness of macrophages, dendritic cells
and other immune cells. Moreover, a specific agonist of α7nAChR
shows anti-inflammatory effects in various experimental animal
models (9,12).
According to research on the anti-inflammatory
effects of α7nAChR in models of sepsis and acute inflammation,
activation of α7nAChR in macrophages could be an effective pathway
towards suppression of pro-inflammatory cytokine production
(13,14). In addition, stimulation of α7nAChR
induces STAT3 activation and subsequent protection of macrophages
from endoplasmic reticulum stress-induced apoptosis, which affects
the activity of the M2-type macrophage subset (15). Furthermore, Wazea et al
(16) showed that the
anti-atherogenic role of α7nAChR was due to attenuation of
macrophage oxidative stress. In patients with CD, an inflammatory
macrophage population present in the intestine produces large
amounts of pro-inflammatory cytokines, including interleukin
(IL)-23, tissue necrosis factor-α (TNF-α) and IL-6 (17). This work demonstrates that α7nAChR
serves a crucial role in the regulation of macrophage activation
and is suggestive of a potential role for macrophage α7nAChR in
inflammatory diseases such as colitis. Indeed, some studies suggest
that α7nAChR plays a negative role in the progression of colitis
(18,19). However, treatment with
pharmacological agonists of α7nAChR in colitis induced intolerable
side effects in an experimental mouse model, and large doses of
such agonists may have disruptive effects on colon integrity and
induce anxiety-like behavior (20,21).
So it was hypothesized that a reduced dosage of α7nAChR agonist,
combined with other drugs, could reduce side effects and improve
therapeutic effects.
SHP2, first identified as a proto-oncoprotein, is a
tyrosine phosphatase family member. A number of previous studies
have reported that SHP2 serves an important role in immune
regulation. For example, upregulation of SHP2 in CD4+
T-cells causes Th17 cell loss in simian immunodeficiency virus
infection (22). In addition, SHP2
activation is enhanced in Helicobacter pylori infection,
which suppresses interferon-γ signaling (23). In dextran sulfate sodium
(DSS)-induced colitic model mice, SHP2 phosphorylation levels are
elevated, disrupting macrophage IL-10/STAT3 signaling and its
negative effect on development of inflammation (24). Clinical data has revealed that
macrophage-restricted SHP2 activation is associated with IBD
(24). To develop a more potential
therapy the activation of α7nAChR, the effects of treatment with
PNU282987 (an α7nAChR agonist) on DSS-induced mouse colitis were
evaluated. Furthermore, an optimized treatment using a combination
of α7nAChR agonist PNU282987 and SHP2 inhibitor SHP099 is
demonstrated in experimental colitis in mice.
Materials and methods
Mice
A total of 112 male C57BL/6 (B6) mice (age, 6–8
weeks; weight, 22–25 g) were purchased from Shanghai SLAC
Laboratory Animal Co., Ltd., Mice were maintained under specific
pathogen-free conditions, all mice were provided free access to
food and water, and conditions were a temperature of 23±2°C,
humidity of 40–70% and a 12-h light/dark cycle. The procedures
involving mice were carried out using protocols approved by the
Ethics Committee of the First Affiliated Hospital of Soochow
University (Suzhou, China).
Antibodies and reagents
The following primary antibodies were used:
Anti-CD68 (Bio-Rad Laboratories, Inc., cat. no. MCA1957T) and
anti-α7nAChR (Sigma-Aldrich; Merck KGaA, cat. no. M220). PNU282987
was purchased from Sigma-Aldrich (Merck KGaA, cat. no. P6499),
methyllycaconitine citrate (MLA) was purchased from Sigma-Aldrich
(Merck KGaA, cat. no. M168) and SHP099 was from Selleck Chemicals
(cat. no. S8278). All drugs were dissolved in PBS, and PBS was used
as vehicle control. DSS (molecular mass, 36,000-50,000, cat. no.
9011-18-1) was purchased from MP Biomedicals, LLC and dissolved in
the drinking water at 3% (w/v) concentration.
DSS-induced experimental colitis
model
Mice were provided 3% (wt/vol) DSS dissolved in the
drinking water for 7 days. Weight loss was monitored daily. At the
end of treatment, mice were euthanized by 100% CO2
inhalation with 30% volume/minute flow rate; death was verified by
cervical dislocation, colons were excised and colon length from the
end of the cecum to the anus was recorded. In PNU282987-treated
group, PNU282987 was dissolved in PBS and mice were intraperitoneal
injection (i.p.) followed by DSS administration. For inhibiting
a7nAChR, MLA was pre-administrated (i.p.) 1 day before PNU282987
treatment. PBS was used as vehicle control.
Histological analysis
Colons from mice treated with or without 3% (wt/vol)
DSS were excised and fixed in 4% paraformaldehyde at 4°C overnight,
then colons were dehydrated with graduated ethanol (70,80,85,90,95
and 100%) and embedded in paraffin. Paraffin-embedded colons were
sectioned into 5 µM slices and stained with hematoxylin and eosin,
as described previously (25).
Histological score was assessed (25) based on leukocyte infiltration: (0,
no infiltration; 1: Basolateral; 2: Infiltration to muscularis
mucosae; 3: Infiltration to submucosa), epithelial cell disruption
(0: No epithelium disruption; 1: Crypt hyperplasia; 2: Mild crypt
disruption; 3: Loss of crypt in large area). The histological score
was conducted blind by two of the authors. Immunofluorescence
analysis. Mouse colon specimens were fixed in 4%
paraformaldehyde at 4°C overnight, dehydrated with 30 and 20%
sucrose solution, then embedded with O.C.T. compound (Sakura
Finetek USA, Inc.). Frozen sections (7 µM) were blocked with 10%
normal goat serum (Beyotime Institute of Biotechnology) diluted
with 1X PBS (1 ml normal goat serum supplied with 9 ml PBS) for 1 h
at room temperature, then incubated with primary antibody to
α7nAChR (1:200) or CD68 antibody (1:200 dilution) at 4°C overnight,
and fluorophore-coupled secondary antibodies (Mouse IgG2a
Cross-Adsorbed Secondary Antibody Alexa Fluor 488 (cat no. A-21131;
Thermo Fisher Scientific, Inc., 1:500); Rabbit anti-Rat IgG (H +
L), Texas Red® (cat no. PA1-28571; Thermo Fisher
Scientific, Inc., 1:500) for 1 h in the dark at room temperature.
Sections were counterstained with 200 nM DAPI for 5 min and mounted
in Fluorescent Mounting Medium. Fluorescent images were captured
using a TCS SP8, laser-scanning confocal fluorescence microscope
(Leica Microsystems GmbH).
Isolation of intestinal lamina propria
macrophages
Briefly, mice were treated with 3% (wt/vol) DSS for
7 days to induce colitis, and the large intestines were removed and
carefully cleaned of their mesentery and fat tissue. The large
intestine was then opened longitudinally, washed of fecal contents,
cut into 0.5 cm sections, and subjected to two sequential 20-min
incubations at 37°C in Hanks balanced salt solution (HBSS) plus 5%
FBS (Gibco; Thermo Fisher Scientific) and 5 mM EDTA at 37°C with
gentle agitation to remove epithelial cells. After each incubation
step, media containing epithelial cells and tissue debris was
discarded. The remaining tissue was minced with ophthalmic scissors
and incubated for 30 min in HBSS plus 5% FBS, 1 mg/ml collagenase
VIII (Sigma-Aldrich; Merck KGaA) and 0.1 mg/ml DNase I
(Sigma-Aldrich; Merck KGaA) at 37°C with gentle agitation. Cell
suspensions were collected and filtered through a 70-µm strainer
and pelleted by centrifugation at 300 × g at room temperature for
10 min. Cells were incubated with CD45-Percific Blue (diluted in
PBS with 1:1,000), CD11b-FITC (diluted in PBS with 1:1,000) and
F4/80-PE (diluted in PBS with 1:1,000) antibody at room temperature
in dark for 30 min, after washing twice with PBS, cells were
resuspended in PBS with 1×106 cells/ml. Colonic
macrophages were sorted from CD45 positive cells, CD11b and F4/80
double positive gates by using flow cytometry (BD FACSAria™ III
sorter). Purity of colonic macrophages was >85% analyzed with BD
FACSCalibur™ with CD11b and F4/80 double positive gates. Purified
colonic macrophages were cultured in RPMI-1640 (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml
streptomycin (Gibco; Thermo Fisher Scientific, Inc.).
Generation of bone marrow derived
macrophages (BMDMs)
Mice were euthanized and their femurs and tibias
were isolated and rinsed in 70% (vol/vol) ethanol. Bone marrow
cells were flushed out with cold PBS and passed through 40 µM
filters, then centrifuged at 2,500 × g for 5 min at room
temperature. Pellets were suspended in 1 ml ACK red blood cell
lysis buffer (Sangon Biotech Co., Ltd.) at room temperature for 1
min, then neutralized with 20 ml PBS. After centrifugation at 2,500
× g for 10 min at room temperature, the isolated bone marrow cells
were resuspended in complete DMEM (supplemented with 10% FBS, 100
U/ml penicillin and 100 µg/ml streptomycin) containing 10 ng/ml
macrophage colony stimulating factor (PeproTech, Inc.) and cultured
at 37°C in a 5% CO2 incubator to allow for
differentiation for 7 days.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
RNA was isolated from 30 mg of colon tissue.
Biopsies were weighed and homogenized using liquid nitrogen and
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) was used to isolate total RNA following the manufacturer's
instructions. cDNA was reverse transcribed from 1 µg total RNA with
the ReverTra Ace qPCR RT kit (Toyobo Life Science). qPCR was
performed with SYBR® Green Realtime PCR Master Mix
(Toyobo Life Science) and an ABI 7500 instrument (Applied
Biosystems; Thermo Fisher Scientific, Inc.), using the following
primers were: α7nAChR, forward 5′-TCCTCATGTCAGCCCCAAAC-3′,
reverse 5′-AGACCGACAGCTACAGGGAT-3′; IL-1β, forward
5′-AAGGGGACATTAGGCAGCAC-3′, reverse 5′-ATGAAAGACCTCAGTGCGGG-3′;
IL-6, forward 5′-TGCCTTCTTGGGACTGATGC-3′, reverse
5′-TGAAGTCTCCTCTCCGGACT-3′; TNF-α, forward
5′-GCACCACCATCAAGGACTCA-3′, reverse 5′-TGTGAGGAAGGCTGTGCATT-3′;
Il-12 p35, forward 5′-ATGATGACCCTGTGCCTTGG-3′, reverse
5′-CACCCTGTTGATGGTCACGA-3′; IL-23 p19, forward
5′-TGGAGCAACTTCACACCTCC-3′, reverse 5′-GGCAGCTATGGCCAAAAAGG-3′;
IL-10, forward 5′-CCTCGTTTGTACCTCTCTCCG-3′, reverse
5′-CAAGTGTGGCCAGCCTTAGA-3′; GAPDH, forward
5′-CCTTCCGTGTTCCTACC-3′, reverse 5′-CAACCTGGTCCTCAGTGTA-3′. Gene
transcripts were normalized against the loading control GAPDH, and
relative expression levels were calculated using the
2−ΔΔCq method (26).
ELISA: Colon (0.5 cm) was excised from mice
with sterile scissors, washed three times in cold PBS plus 100
µg/ml gentamycin, 100 U/ml penicillin and 100 µg/ml streptomycin.
The colons slices were cultured with RPMI-1640 medium supplemented
with 10% FBS and 100 µg/ml gentamycin at 37°C, 5% CO2
for 24 h. Supernatant was harvested for cytokine measurement.
Cytokine production from colons was analyzed with Quantikine ELISA
kits (R&D Systems; Cat. no. MLB00C, M6000B, MTA00B, M1270 and
M1000B respectively), following the manufacturer's instructions.
Cytokine production was normalized according to total colon tissue
weight. Each treatment contained 3–4 mice.
Western blot analysis
Colon proteins were extracted as described
previously (25). Briefly, RIPA
buffer supplemented with protease inhibitors (Sangon Biotech Co.,
Ltd.) was used as lysis buffer. Colon proteins were separated by
10% SDS-PAGE, and the gels were then electro-transferred onto
nitrocellulose filter membranes (Whatman; Cytiva). The membranes
were incubated with antibodies against α7nAChR (Sigma-Aldrich;
Merck KGaA, cat. no. M220, 1:1,000), phosphorylated (p-)SHP2
(Abcam; cat. no. ab62322, 1:500), SHP2 (Abcam, cat. no. ab131541,
1:500), p-STAT3 (Cell Signaling Technology, Inc.; cat. no. 9145,
1:1,000); STAT3 (Santa Cruz Biotechnology, Inc., cat. no. sc-482,
1:100), p-Jak2 (Abcam, cat. no. ab32101, 1:1,000); Jak2 (cat. no.
ab108596, Abcam, 1:1,000) or GAPDH (Abcam, cat. no. ab181602,
1:2,000) overnight at 4°C. The membranes were then incubated with
an IRDye 800CW-conjugated secondary antibody (Rockland
Immunochemicals Inc. 1:20,000) for 1 h at room temperature. Images
were acquired using an Odyssey infrared imaging system
(Odyssey® CLx Imaging System, LI-COR Biosciences) and
ImageJ software v1.52 (NIH) was used for analysis.
Intracellular ROS assay
Intracellular oxidative stress was measured using
dichlorodihydrofluorescein diacetate oxidation. Macrophages were
incubated with 100 µl of 1X DCFH-DA/media solution (Sangon Biotech
Co., Ltd.) for 1 h at 37°C. After washed with DPBS three times, the
fluorescence was read using a fluorometric plate reader
(FlexStation 3, Molecular Devices, LLC) at 480 nm/530 nm.
Statistical analysis
Data are presented as the mean ± SD. All experiments
were repeated three times. One-way ANOVA followed by Dunnett's
multiple comparison post hoc test was performed when comparing all
experimental groups with the control group, whereas Tukey's post
hoc test was used for comparing the means of all treatments to the
mean of every other treatment, using GraphPad software version 6.0
(GraphPad Software, Inc.). P<0.05 was considered to indicate a
statistically significant difference.
Results
α7nAChR expression in DSS-induced
mouse colitis
As a nicotinic acetyl cholinergic receptor, α7nAChR
is expressed widely in the central and peripheral nervous systems;
previous studies have shown that activation of α7nAChR could
alleviate inflammation (12,27,28).
However, the effects and associated mechanisms of receptor
signaling in IBD is not completely understood. Immunofluorescence
analysis was used to investigate the expression of α7nAChR in
colonic tissues of mice, and the results demonstrated that α7nAChR
expression was highly induced in inflammatory sites within the
colon but was only sparsely expressed in healthy colon (Fig. 1A). Macrophages serve a crucial role
in intestinal homeostasis (19),
and expression of their specific surface marker, CD68, was more
abundant in the inflamed compared with in the healthy colon
(Fig. 1A); the merged panel
demonstrates that these two proteins shared a degree of
co-localization, which suggested that α7nAChR on macrophages may
serve a crucial role in colitis. To clarify α7nAChR expression
pattern after colitis onset, western blotting was used to analyze
the protein expression levels of α7nAChR at day 0, 3, 5, 7 after
DSS administration in mice. α7nAChR expression was significantly
increased after 5 days of DSS treatment (Fig. 1B). To evaluate whether
colitis-induced α7nAChR protein expression was due to reduced
degradation or increased receptor synthesis, mRNA expression levels
of α7nAChR in mice were assessed using RT-qPCR. The results showed
α7nAChR mRNA levels have a similar expression pattern as protein
levels in the mouse colon after DSS treatment (Fig. 1C). These data demonstrated that
α7nAChR is highly expressed in the inflamed colon and suggested
that the receptor on macrophages may play a critical role in
colitis.
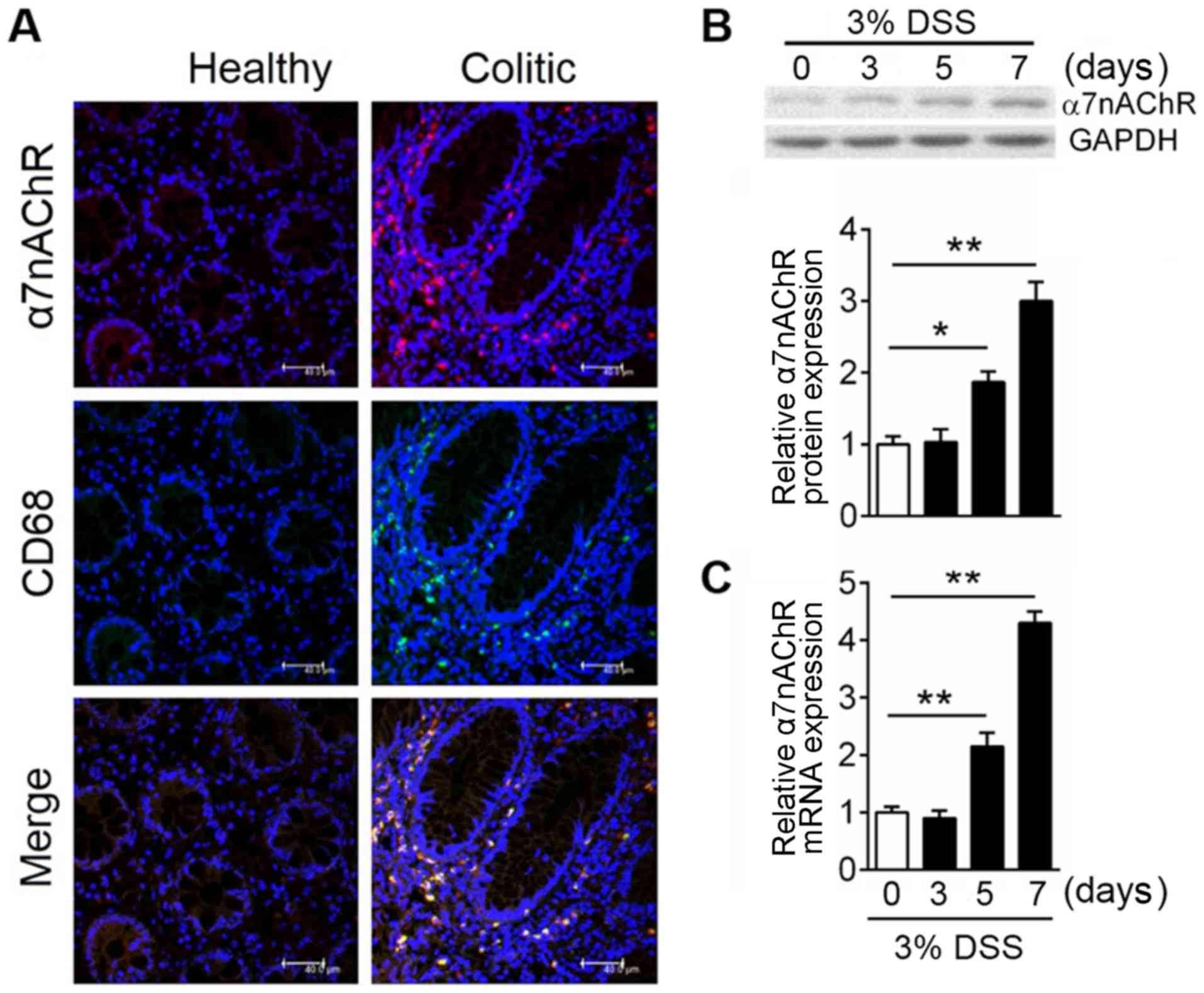 | Figure 1.Macrophages drive increased α7nAChR
expression in IBD. In the experimental model, colitis was induced
with 3% DSS in C57BL/6 mice, mice were sacrificed at the indicated
time points and the colon was excised for protein and mRNA
expression analysis. (A) Representative images of
immunofluorescence analyses of colonic mucosal tissue in healthy
and colitic mice. α7nAChR is in red; CD68 is in green expressions;
DAPI is in blue. Co-localization of the α7nAChR and macrophage
markers are displayed in merged images. Scale bar, 40 µM. Relative
expression levels of α7nAChR (B) protein and (C) mRNA were analyzed
by western blotting and reverse transcription-quantitative PCR,
respectively, in DSS-induced colitis mice at days 0, 3, 5 and 7;
GAPDH was used as internal reference for both. Data are expressed
as the mean ± SD of three independent experiments; n=5–7
mice/group; *P<0.05, **P<0.01. α7nAChR, α7 nicotinic
acetylcholine receptor; DSS, dextran sulfate sodium; IBD,
inflammatory bowel disease. |
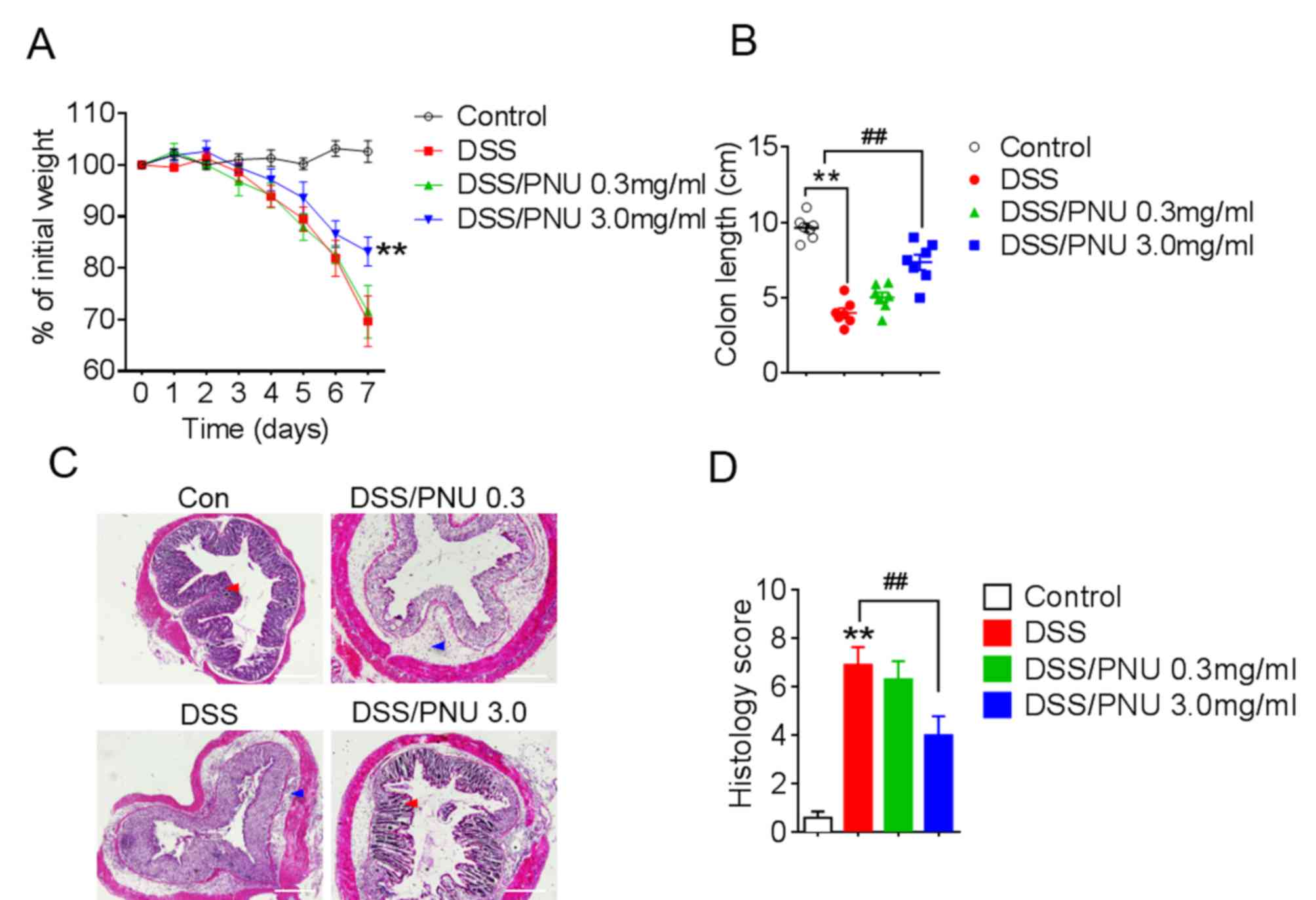
Treatment with α7nAChR specific
agonist PNU282987 ameliorates DSS-induced experimental colitis in
mice
To investigate the role of α7nAChR in colitis, the
α7nAChR specific agonist PNU282987 (0.3 or 3 mg/kg) was
administered to mice by i.p. injection daily, following DSS
administration. PNU282987 was dissolved in PBS and PBS was used as
vehicle control. Pathological changes were observed in each group;
progressive weight loss occurred in colitic mice treated with DSS,
but the extent of weight loss was reduced with a 3 mg/kg of
PNU282987 treatment in colitic mice compared with DSS+PBS-treated
group (Fig. 2A). Moreover, the
protective effects of PNU282987 were decreased by pre-treatment
with MLA for 1 day, the antagonist of α7nAChR (Fig. S1). Colon shortening was lessened
in colitic mice treated with PNU282987 at 3 mg/kg (Fig. 2B). Histological analysis of colons
from colitic mice treated with the PBS control showed severe
inflammation with loss of crypts (Fig.
2C, red arrow) and infiltration of leukocytes (Fig. 2C, blue arrow), whereas this damage
was alleviated by treatment with PNU282987 (Fig. 2C). Histological scores were
obtained based on colonic epithelial cell disruption and leukocyte
infiltration, and the scores in the colitic mice were significantly
lower after activation of α7nAChR by PNU282987 at 3 mg/kg (Fig. 2D). These results demonstrated that
PNU282987 may exert a protective effect in DSS-induced mouse
colitis.
Cytokine expression in colons of
colitic mice treated with α7nAChR agonist PNU282987
Cytokines serve a crucial role in the pathology of
colitis (1,2); therefore, IL-1β, IL-6, TNF-α, IL-12,
IL-23 and IL-10 levels in colonic tissue were analyzed by ELISA.
Compared with untreated control colitic mice, colitic mice treated
with 3 mg/kg PNU282987 exhibited a significant reduction in IL-1β,
IL-6, TNF-α, IL-12 and IL-23 concentrations, whereas IL-10 levels
were significantly increased after treatment with PNU282987
(Fig. 3A-F). In DSS alone-treated
group, IL-10 level was also enhanced possibly due to the macrophage
or a small amount of Treg cell activation. TGF-α and IL-22
production showed no significant differences between mice treated
with PNU282987 or PBS; these two cytokines play a negative role in
IBD development (data not shown).
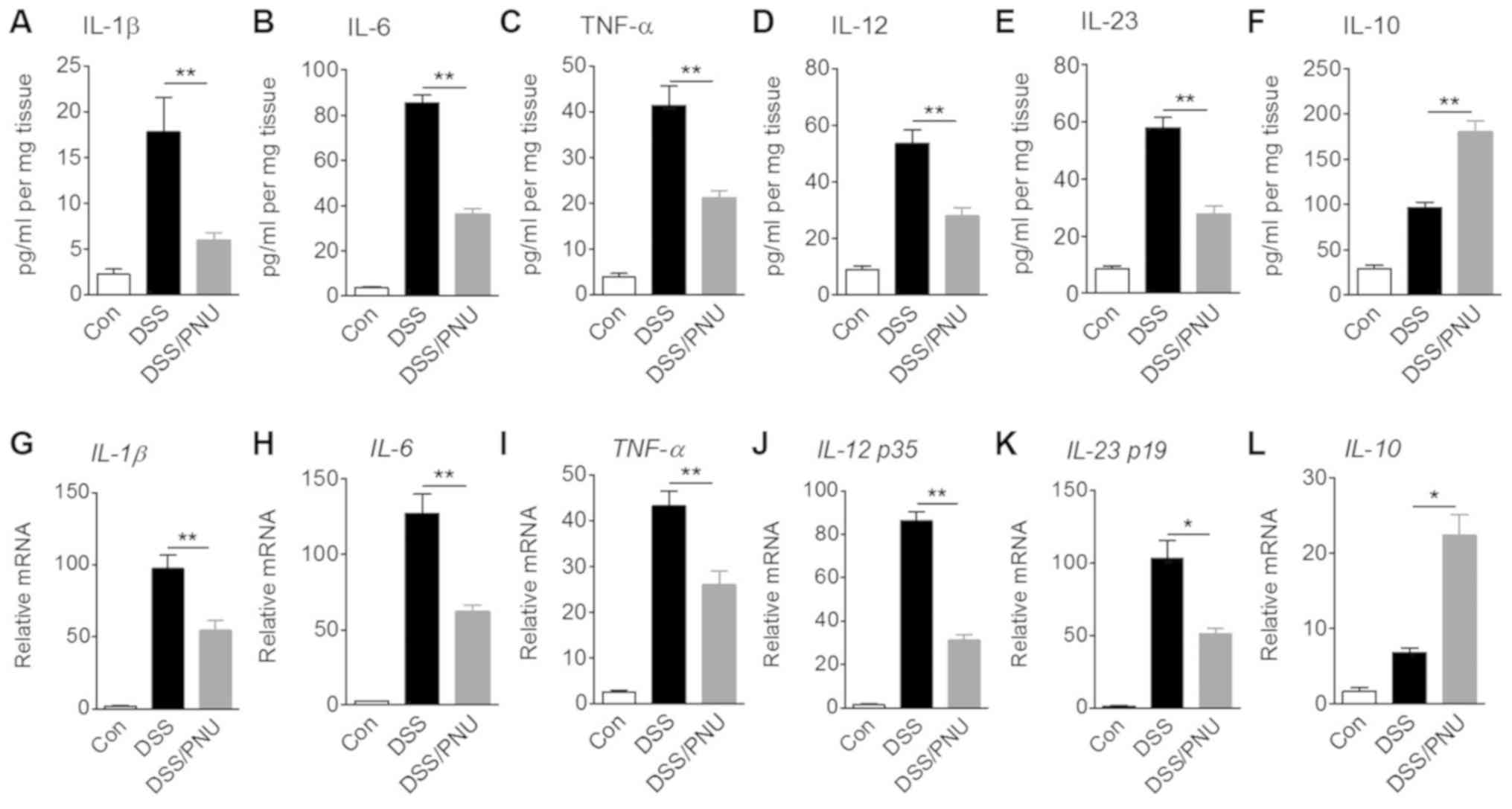 | Figure 3.Effects of α7nAChR stimulation with
PNU282987 on protein concentrations and mRNA expression levels of
cytokines. C57BL/6 mice were treated with 3% DSS for 7 days, and
intraperitoneally administered with PNU282987 (3 mg/kg;
n=3-4/group) daily. Colonic samples were collected and cultured for
24 h. Supernatants of tissue culture were harvested and (A) IL-1β,
(B) IL-6, (C) TNF-α, (D) IL-12, (E) IL-23 and (F) IL-10 protein
concentrations were determined by ELISA. Colonic macrophages were
sorted from colonic lamina propria cells of mice and mRNA
expression levels of (G) IL-1β, (H) IL-6, (I) TNF-α, (J) IL-12 p35,
(K) IL-23 p19 and (L) IL-10 were measured by reverse
transcription-quantitative PCR. Data are displayed as the mean ± SD
of three independent experiments. *P<0.05, **P<0.01. α7nAChR,
α7 nicotinic acetylcholine receptor; DSS, dextran sulfate sodium;
IL, interleukin; PNU, PNU282987; TNF-α, tissue necrosis
factor-α. |
One of the functions of macrophages is maintenance
of intestinal homeostasis through cytokine production. In
DSS-induced colitic mice treated with PNU282987 or with PBS,
colonic macrophages were sorted >85% purity by flow cytometry
using CD11b and F4/80 antibody staining (Fig. S2). IL-1β, IL-6, TNF-α, IL-12 p35,
IL-23 p19 and IL-10 mRNA expression levels in macrophages were
measured. In colitic mice treated with PNU282987, mRNA expression
levels of IL-1β, IL-6, TNF-α, IL-12 p35 and IL-23 p19 were
significantly decreased compared with colitic mice treated with
PBS, whereas IL-10 mRNA expression was significantly raised in
colonic macrophages of colitic mice treated with PNU282987
(Fig. 3G-L). Also due to the
activation of macrophages in the colitic colon, IL-10 mRNA level
was enhanced in the DSS-treated group compared with the control.
These results are consistent with previous reports that suggest
acute DSS-induced colitis development depends on macrophage
activation (29,30).
SHP2 is activated in DSS-induced mouse
colonic macrophages
SHP2 has previously been identified as a detrimental
factor in the colonic immune system (24). To study SHP2 activation in colonic
macrophages and the effect of treatment with PNU282987, mice were
treated with DSS and colonic macrophages were isolated. JAK2 and
STAT3 phosphorylation levels were notably upregulated in the
colitic model mice, whereas a markedly decreased level was seen in
the PNU282987-treated group (Fig.
4A). Moreover, SHP2 phosphorylation levels were also enhanced
in the colitic group and were unchanged in the colitic group
treated with PNU282987, indicating that PNU282987 has no direct
effect on SHP2 activation (Fig.
4A). As STAT3 is involved in both α7nAChR and SHP2 pathways,
activating α7nAChR or inhibiting SHP2 leads to upregulating the
level of p-STAT3, which is beneficial for recovery in colitis
(16,24). To investigate the possible role of
SHP2 in PNU282987 effects on macrophages, SHP2 expression in BMDMs
was knockdown by three siRNA, and the third siRNA show the highest
efficacy on SHP2 knockdown (knockdown efficacy was verified by
western blot; Fig. 4B). Cells were
incubated with PNU282987 followed with LPS challenge which is a
classical regent to inducing inflammation in macrophages. In BMDMs
which had been challenged with LPS and treated with PNU282987,
TNF-α and IL-6 cytokine production was significantly reduced in
cells depleted of SHP2 or PNU282987 treatment compared with LPS
only administration group. In addition, a combination of siRNA-SHP2
and PNU282987 decreased TNF-α and IL-6 cytokine protein level
(Fig. 4C and D). These results
indicated that inhibiting SHP2 activation may have a combined
effect with PNU282987 in suppression of inflammatory
macrophages.
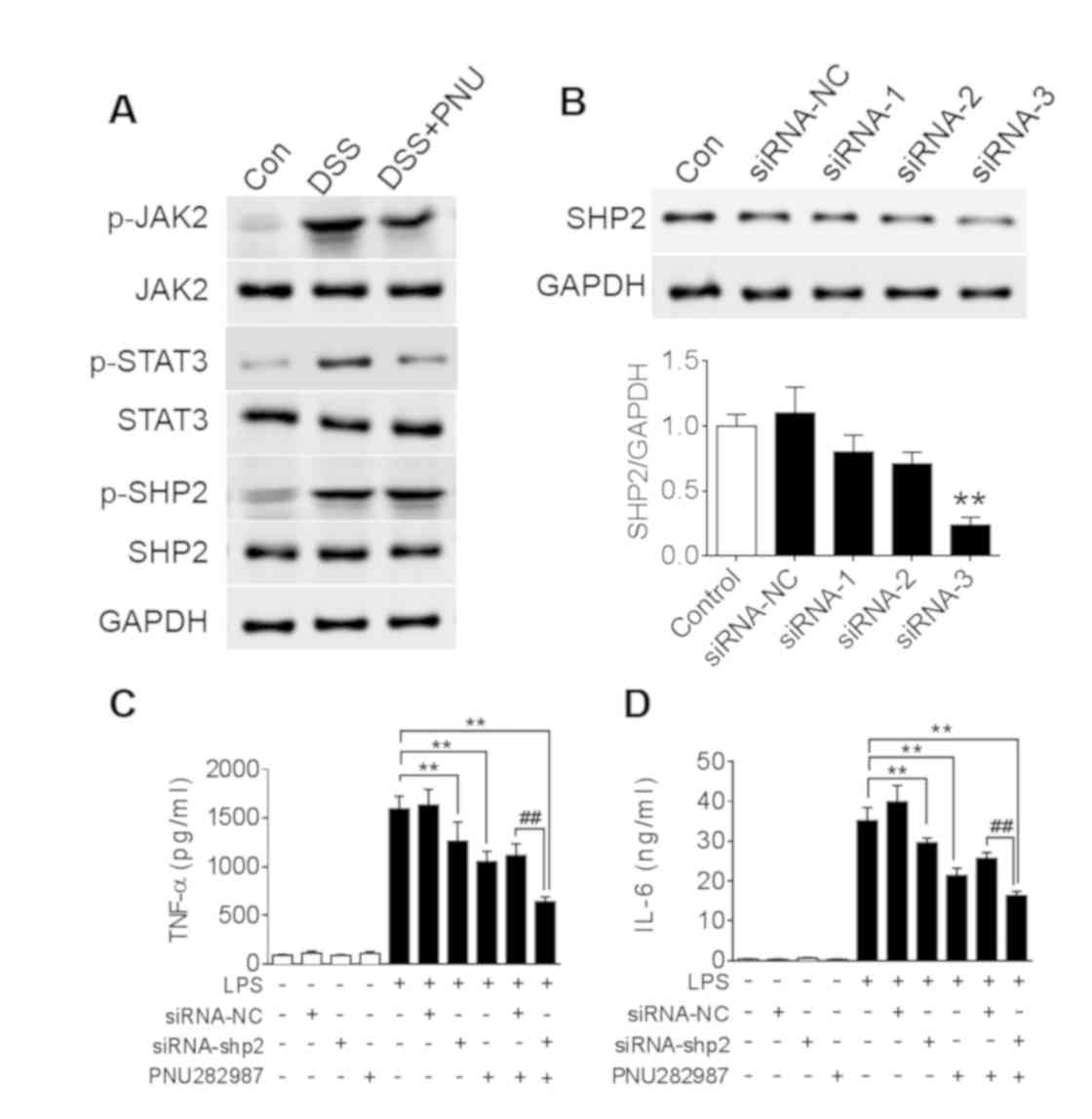 | Figure 4.SHP2 is activated in DSS-induced
colitis. (A) C57BL/6 mice were treated with 3% DSS for 7 days and
intraperitoneally administered PNU282987 (3 mg/kg, n=3-4/group)
daily; JAK2, STAT3 and SHP2 phosphorylation levels in the colon
were analyzed after DSS treatment with or without PNU282987
treatment. (B) SHP2 protein level was analyzed with western blot to
verify the efficacy of siRNA depletion. **P<0.01 vs. control
group; Subsequently, BMDMs were treated with siRNA-3 to silence
SHP2 expression, then treated with PNU282987 (10 µM) followed with
LPS for 24 h, supernatant was harvested to determine the
concentrations of (C) TNF-α and (D) IL-6 by ELISA. Data are
presented as the mean ± SD of three independent experiments.
**P<0.01 vs. LPS treatment; ##P<0.01
LPS+PNU282987+si-shp2 vs. LPS+PNU282987+si-NC. BMDMs, bone marrow
derived macrophages; DSS, dextran sulfate sodium; IL-6,
interleukin-6; LPS, lipopolysaccharide; PNU, PNU282987; TNF-α,
tissue necrosis factor-α. |
Inhibition of SHP2 with SHP099 and
treatment with PNU282987 may synergistically suppress macrophages
and colonic inflammation
SHP2 is a ubiquitously expressed tyrosine
phosphatase and previous studies reported that it has an inhibitory
effect on the JAK/STAT signaling pathway (24). To investigate whether inhibiting
SHP2 has a synergistic effect in combination with PNU282987, BMDMs
were treated with SHP099, a selective pharmacological inhibitor of
SHP2, and PNU282987 simultaneously. Co-treatment with SHP099 and
PNU282987 resulted in a significant decrease in TNF-α and IL-6
expression compared with PNU282987 only treatment in LPS-induced
macrophages, whereas no significant decrease was identified in
TNF-α and only a slight decrease in IL-6 production with SHP099
treatment alone (Fig. 5A).
Excessive accumulation of reactive oxygen species (ROS) can induce
further damage in inflamed colonic tissue (31). Consistent with the cytokine
production data, ROS levels induced by LPS in macrophages were
significantly decreased by a combination of SHP099 and PNU282987
treatments, whereas there were no differences identified between
the SHP099-alone treatment group (Fig.
5B).
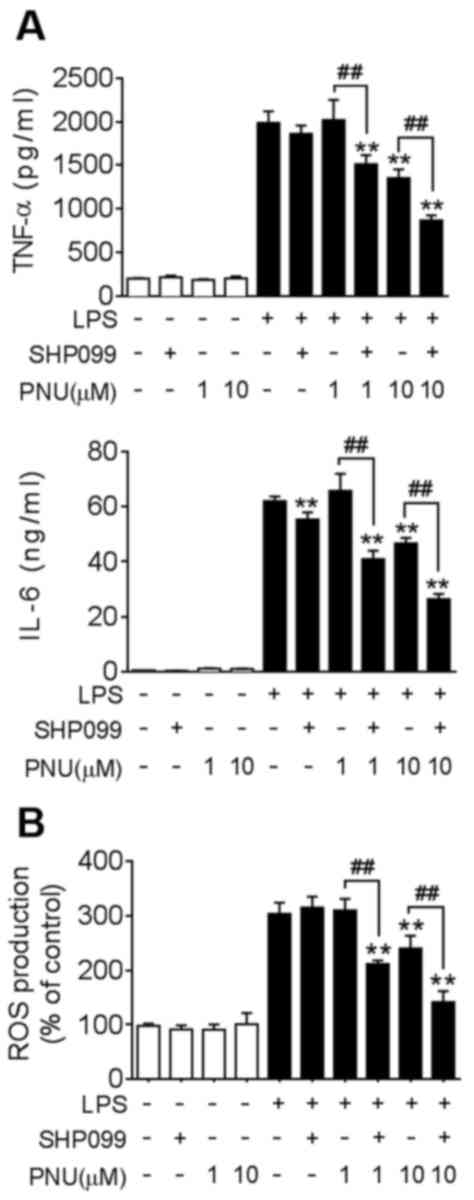 | Figure 5.Co-treatment with SHP099 and
PNU282987 synergistically suppresses cytokine production and ROS
levels in macrophages. BMDMs were pre-treated with SHP099 (10 µM),
PNU282987 (1 or 10 µM), or a combination of SHP099 and PNU282987
for 30 min, followed by LPS (100 ng/ml). (A) TNF-α and IL-6 protein
levels in BMDMs, analyzed after LPS-treatment for 24 h. (B) ROS
levels were quantified in BMDMs using an assay to detect
fluorescent carboxyl DCFH-DA after LPS-treatment for 4 h. Data are
displayed as the mean ± SD of three independent experiments.
**P<0.01 vs. LPS treatment, ##P<0.01 LPS+PNU282987
vs. LPS + PNU282987+SHP099. IL-6, interleukin-6; LPS,
lipopolysaccharide; PNU, PNU282987; ROS, reactive oxygen species;
TNF-α, tissue necrosis factor-α. |
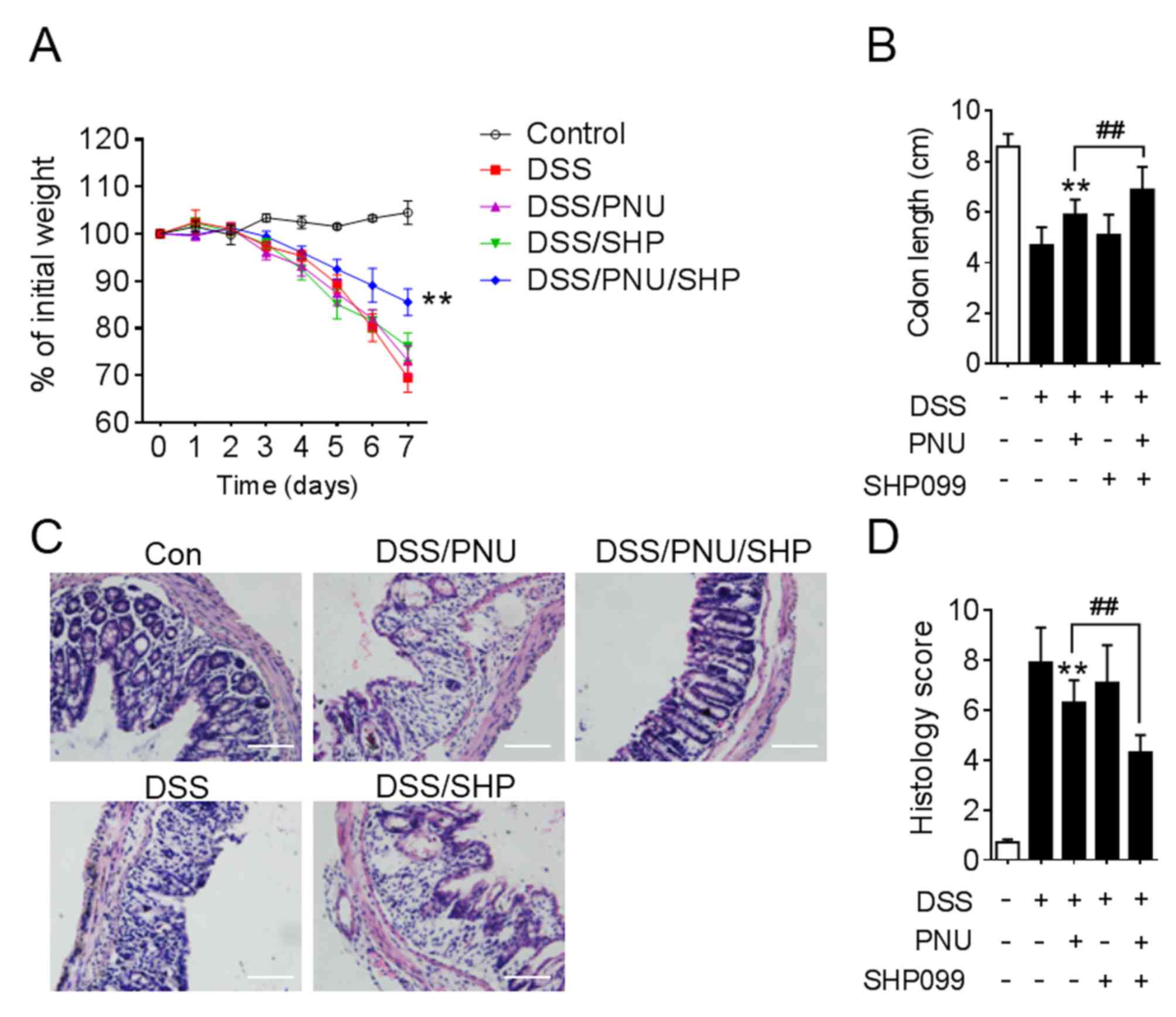
To further investigate the effect of SHP099 and
PNU282987 in the experimental animal model, DSS-induced colitic
mice were treated with PNU282987 combined with SHP099.
Administration of DSS lead to weight loss, colon shortening and
severe colonic disruption. The aim of the present study was to find
a combined drug that could lower the dose of PNU282987 and protect
mice from DSS-induced colitis, so 0.3 mg/kg PNU282987 or 1 mg/kg
SHP099 treatment alone were chosen, which had no significant effect
on colitis. However, when 0.3 mg/kg PNU282987 was combined with
SHP099 administration, this resulted in attenuated weight loss and
colon shortening (Fig. 6A and B).
Histopathologically, in the PNU282987 and SHP099 co-treatment
groups, mice displayed markedly mitigated mucosal damage and less
leukocyte infiltration compared with PNU282987 treatment alone
(Fig. 6C). Statistical analysis of
histological scores supported these conclusions (Fig. 6D).
Discussion
Several studies have indicated that activating
α7nAChR can alleviate colonic inflammation to a certain degree
(18,19). The present study demonstrated that
macrophage-specific expression of α7nAChR was induced in inflamed
colons of DSS-induced colitis in mice. Intestinal integrity in the
colitic mice was improved by a large dose of the α7nAChR agonist
PNU282987, accompanied by side effects such as disrupted colon
integrity and anxiety-like behavior (20,21).
More importantly, inhibiting SHP2 with SHP099 was found to have a
synergistic effect with PNU282987 on DSS-induced colitis. 0.3 mg/kg
PNU282987 have no effect when treated colitic mice alone, while it
has significantly protective effect combined with SHP099
administration.
Previous studies have also shown that α7nAChR
activation is associated with alleviation of inflammation in
LPS-induced endotoxin shock, accompanied by reduced
pro-inflammatory cytokine production and leukocyte infiltration
(9,16). Consistent with these studies,
results from the present study demonstrated that stimulating
α7nAChR with its selective agonist PNU282987 significantly
attenuated DSS-induced acute colitis, which is accompanied by
colonic shortening and weight loss in mice. Histological analysis
of colon sections showed immune cell infiltration and tissue damage
of the epithelial layer induced by DSS, which was markedly reduced
after PNU282987 treatment compared with PBS-treated control mice.
This evidence indicated that PNU282987 may have a protective
effect, maintaining colonic homeostasis at large doses. In the
present study, PNU282987 administration following DSS-induced
colitis caused a marked drop in TNF-α, IL-1β, IL-6, IL-12 and IL-23
levels, whereas IL-10 was increased. IL-10 is a cytokine that
modulates both innate and adaptive immunity, primarily by exerting
anti-inflammatory effects. In inflammatory colon, IL-10 is produced
by many cell types including both classical (M1) or alternative
(M2) macrophages. In colitic colon of DSS-treated mice, macrophages
exhibit M1 phenotype, while PNU282987 could alter macrophages into
M2 phenotype (32). M2 macrophages
produce more IL-10 than the M1 phenotype and this may be the reason
why IL-10 levels were significantly increased after treatment with
PNU282987 in colitic mice. By contrast, Tasaka et al
(19) found that TNF-α and IL-1β
levels were not changed after PNU282987 treatment in colitic mice.
This disparity may have been due to differences in the measurement
of cytokines from the colon (directly lysed colon vs. supernatant
of colon culture). These cytokines are mainly produced by
macrophages and are positively associated with the development of
IBD, so a degree of manipulation of cytokine levels may help to
improve IBD pathology. These findings have shown that the relative
mRNA expression level of various cytokines in colonic macrophages
was consistent with protein levels in colon culture.
In the present study, it was observed that combined
use of PNU282987 and SHP099 presented an anti-inflammatory effect
in LPS-induced macrophages. Abnormal macrophage activation is the
major cause of tissue damage in colitis, and excessive ROS
accumulation causes further disruption to tissue and increased
pathology (1,2). Thus, a combination of these two drugs
may have potential effect on protecting mice against DSS-induced
colitis.
It was also found that treatment with SHP099 alone
could not reverse the pathological deterioration in DSS-induced
colitic mice, the possible reasons include that the treatment
dosage used was not high enough in these DSS-induced colitic mice,
thus inhibiting SHP2 alone may not represent a good method of
treating colitis. However, SHP099 exhibits a promising
synergistical effect with PNU282987 on the inflammatory
response.
The present study demonstrated that IL-10 expression
in colon cultures is significantly increased in DSS-induced colitis
in mice treated with PNU282987. Based on other reports, STAT3
phosphorylation is crucial for IL-10 expression in macrophages
induced by LPS, and STAT3 in innate cells has been shown to protect
mice from colitis (33,34). A previous study identified SHP2 as
an inhibitory molecule for STAT3 phosphorylation in macrophages and
overexpression of SHP2 exacerbates colonic inflammation via
disrupting the IL-10-STAT3 axis (20). Other findings indicate that SHP2
reduces JAK2/STAT3 signaling through promotion of JAK2
phosphorylation at Y570, resulting in amelioration of TGF-α induced
fibroblast activation (35).
Therefore, enhanced STAT3 activation may contribute to the combined
potential therapy of a7nAChR activation and inhibiting SHP2
inhibition. Recently, SHP099 was developed as an allosteric
inhibitor of SHP2 and showed an anti-proliferative effect in
several cancers, such as non-small-cell lung cancer and hematologic
malignancy (36). However, there
are far fewer studies of SHP099 in inflammatory disease. The
present study demonstrated that there was a synergistic effect
between SHP099 and PNU282987 on DSS-induced colitis in mice. The
observed colitis symptoms, including weight loss, colon shortening,
colonic epithelium disruption and leukocyte infiltration were
attenuated by co-treatment with SHP099 and PNU282987.
In conclusion, this study has described a combined
effect between a selective agonist of α7nAChR, PNU282987, and a
tyrosine phosphatase SHP2 inhibitor, SHP099, to protect mice from
DSS-induced colitis. To the best of our knowledge, this data
represents the first demonstration that treatment with these two
compounds is effective in alleviating colonic inflammation,
including leukocyte infiltration and inflammatory cytokine release.
This evidence supports the notion that selectively stimulating
α7nAChR while inhibiting SHP2 might represent a new and attractive
therapy for colonic inflammation.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from The
Natural Science Foundation of Jiangsu Province (grant no.
BK20170372), and Primary Research & Development Plan of Jiangsu
Province (grant no. BE2018659).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JX and GZ performed most of the experiments and
statistical analysis, SG performed cell experiments, JS, FH and ZH
advised on IBD pathology and experimental design, JX, GZ and CX
designed the study. JX wrote the manuscript and all authors
reviewed and approved the final manuscript.
Ethics approval and consent to
participate
The procedures involving mice were carried out using
protocols approved by the Ethics Committee of the First Affiliated
Hospital of Soochow University (Suzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kaser A, Zeissig S and Blumberg RS:
Inflammatory bowel disease. Annual review of immunology.
28:573–621. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maloy KJ and Powrie F: Intestinal
homeostasis and its breakdown in inflammatory bowel disease.
Nature. 474:298–306. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Opstelten JL, Plassais J, van Mil SW,
Achouri E, Pichaud M, Siersema PD, Oldenburg B and Cervino AC: Gut
microbial diversity is reduced in smokers with crohn's disease.
Inflamm Bowel Dis. 22:2070–2077. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yadav P, Ellinghaus D, Rémy G,
Freitag-Wolf S, Cesaro A, Degenhardt F, Boucher G, Delacre M,
International IBD GC, Peyrin-Biroulet L, et al: Genetic factors
interact with tobacco smoke to modify risk for inflammatory bowel
disease in humans and mice. Gastroenterology. 153:550–565. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ueno A, Jijon H, Traves S, Chan R, Ford K,
Beck PL, Iacucci M, Fort Gasia M, Barkema HW, Panaccione R, et al:
Opposing effects of smoking in ulcerative colitis and Crohn's
disease may be explained by differential effects on dendritic
cells. Inflamm Bowel Dis. 20:800–810. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Browning KN, Verheijden S and Boeckxstaens
GE: The vagus nerve in appetite regulation, mood, and intestinal
inflammation. Gastroenterology. 152:730–744. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ingram JR, Rhodes J, Evans BK and Thomas
GA: Preliminary observations of oral nicotine therapy for
inflammatory bowel disease: An open-label phase I–II study of
tolerance. Inflamm Bowel Dis. 11:1092–1096. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Galle-Treger L, Suzuki Y, Patel N,
Sankaranarayanan I, Aron JL, Maazi H, Chen L and Akbari O:
Nicotinic acetylcholine receptor agonist attenuates ILC2-dependent
airway hyperreactivity. Nat Commun. 7:132022016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim H, Kim SR, Je J, Jeong K, Kim S, Kim
HJ, Chang KC and Park SW: The proximal tubular α7 nicotinic
acetylcholine receptor attenuates ischemic acute kidney injury
through Akt/PKC signaling-mediated HO-1 induction. Exp Mol Med.
50:402018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bonaz BL and Bernstein CN: Brain-gut
interactions in inflammatory bowel disease. Gastroenterology.
144:36–49. 2014. View Article : Google Scholar
|
|
11
|
Severs M, Mangen MJ, van der Valk ME,
Fidder HH, Dijkstra G, van der Have M, van Bodegraven AA, de Jong
DJ, van der Woude CJ, Romberg-Camps MJ, et al: Smoking is
associated with higher disease-related costs and lower
health-related quality of life in inflammatory bowel disease. J
Crohns Colitis. 11:342–352. 2017.PubMed/NCBI
|
|
12
|
Kong W, Kang K, Gao Y, Liu H, Meng X, Cao
Y, Yang S, Liu W, Zhang J, Yu K and Zhao M: GTS-21 protected
against lps-induced sepsis myocardial injury in mice through
α7nAChR. Inflammation. 41:1073–1083. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li B, Alli R, Vogel P and Geiger TL: IL-10
modulates DSS-induced colitis through a macrophage-ROS-NO axis.
Mucosal Immunol. 7:869–878. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lin Y, Yang X, Yue W, Xu X, Li B, Zou L
and He R: Chemerin aggravates DSS-induced colitis by suppressing M2
macrophage polarization. Cell Mol Immunol. 11:355–366. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yi L, Lyn YJ, Peng C, Zhu RL, Bai SS, Liu
L, Wang PX, Zhou H and Dong Y: Sinomenine inhibits fibroblast-like
synoviocyte proliferation by regulating α7nAChR expression via
ERK/Egr-1 pathway. Int Immunopharmacol. 56:65–70. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wazea SA, Wadie W, Bahgat AK and El-Abhar
HS: Galantamine anti-colitic effect: Role of alpha-7 nicotinic
acetylcholine receptor in modulating Jak/STAT3, NF-κB/HMGB1/RAGE
and p-AKT/Bcl-2 pathways. Sci Rep. 8:51102018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu L, Zhou Y, Zhou Z, Liu Y, Bai Y, Xing X
and Wang X: Nicotine induces the production of IL-1β and IL-8 via
the α7 nAChR/NF-κB pathway in human periodontal ligament cells: An
in vitro study. Cell Physiol Biochem. 34:423–431. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Grandi A, Zini I, Flammini L, Cantoni AM,
Vivo V, Ballabeni V, Barocelli E and Bertoni S: α7 nicotinic
agonist AR-R17779 protects mice against 2,4,6-trinitrobenzene
sulfonic acid-induced colitis in a spleen-dependent way. Front
Pharmacol. 8:8092017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tasaka Y, Yasunaga D, Kiyoi T, Tanaka M,
Tanaka A, Suemaru K and Araki H: Involvement of stimulation of α7
nicotinic acetylcholine receptors in the suppressive effect of
tropisetron on dextran sulfate sodium-induced colitis in mice. J
Pharmacol Sci. 127:275–283. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Snoek SA, Verstege MI, van der Zanden EP,
Deeks N, Bulmer DC, Skynner M, Lee K, Te Velde AA, Boeckxstaens GE
and de Jonge WJ: Selective alpha7 nicotinic acetylcholine receptor
agonists worsen disease in experimental colitis. Br J Pharmacol.
160:322–333. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Anshul AP and Jerrel LY: Activation of the
α7 Nicotinic ACh Receptor Induces Anxiogenic Effects in Rats Which
Is Blocked by a 5-HT1a Receptor Antagonist.
Neuropharmacology. 70:35–42. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bixler SL, Sandler NG, Douek DC and
Mattapallil JJ: Suppressed Th17 levels correlate with elevated
PIAS3, SHP2, and SOCS3 expression in CD4 T cells during acute
simian immunodeficiency virus infection. J Virol. 87:7093–7101.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang YC, Chen CL, Sheu BS, Yang YJ, Tseng
PC, Hsieh CY and Lin CF: Helicobacter pylori infection activates
Src homology-2 domain-containing phosphatase 2 to suppress IFN-γ
signaling. J Immunol. 193:4149–4158. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xiao P, Zhang H, Zhang Y, Zheng M, Liu R,
Zhao Y, Zhang X, Cheng H, Cao Q and Ke Y: Phosphatase Shp2
exacerbates intestinal inflammation by disrupting macrophage
responsiveness to interleukin-10. J Exp Med. 216:337–349. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xiao J, Shao L, Shen J, Jiang W, Feng Y,
Zheng P and Liu F: Effects of ketanserin on experimental colitis in
mice and macrophage function. Int J Mol Med. 37:659–668. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Harwani SC, Ratcliff J, Sutterwala FS,
Ballas ZK, Meyerholz DK, Chapleau MW and Abboud FM: Nicotine
mediates CD161a+ renal macrophage infiltration and premature
hypertension in the spontaneously hypertensive rat. Circ Res.
119:1101–1115. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Inoue T, Abe C, Sung SS, Moscalu S,
Jankowski J, Huang L, Ye H, Rosin DL, Guyenet PG and Okusa MD:
Vagus nerve stimulation mediates protection from kidney
ischemia-reperfusion injury through α7nAChR+ splenocytes. J Clin
Invest. 126:1939–1952. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lv Z, Wang Z, Luo L, Chen Y, Han G, Wang
R, Xiao H, Li X, Hou C, Feng J, et al: Spliceosome protein Eftud2
promotes colitis-associated tumorigenesis by modulating
inflammatory response of macrophage. Mucosal Immunol. 12:1164–1173.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xia Y, Tian LM, Liu Y, Guo KS, Lv M, Li
QT, Hao SY, Ma CH, Chen YX, Tanaka M, et al: Low dose of
cyanidin-3-O-glucoside alleviated dextran sulfate sodium-induced
colitis, mediated by CD169+ macrophage pathway. Inflamm Bowel Dis.
25:1510–1521. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang Z, Li S, Cao Y, Tian X, Zeng R, Liao
DF and Cao D: Oxidative stress and carbonyl lesions in ulcerative
colitis and associated colorectal cancer. Oxid Med Cell Longev.
2016:98752982016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pinheiro NM, Santana FPR, Almeida RR,
Guerreiro M, Martins MA, Caperuto LC, Câmara NOS, Wensing LA, Prado
VF, Tibério IFLC, et al: Acute lung injury is reduced by the
α7nAChR agonist PNU-282987 through changes in the macrophage
profile. FASEB J. 31:320–332. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang G, Xu B, Shi F, Du M, Li Y, Yu T and
Chen L: Protective effect of methane-rich saline on acetic
acid-induced ulcerative colitis via blocking the TLR4/NF-κB/MAPK
pathway and promoting IL-10/JAK1/STAT3-mediated anti-inflammatory
response. Oxid Med Cell Longev. 2019:78503242019.PubMed/NCBI
|
|
34
|
Qian T, Hong J, Wang L, Wang Z, Lu Z, Li
Y, Liu R and Chu Y: Regulation of CD11b by HIF-1α and the STAT3
signaling pathway contributes to the immunosuppressive function of
B cells in inflammatory bowel disease. Mol Immunol. 111:162–171.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zehender A, Huang J, Gyorfi AH, Matei AE,
Trinh-Minh T, Xu X, Li YN, Chen CW, Lin J, Dees C, et al: The
tyrosine phosphatase SHP2 controls TGFbeta-induced STAT3 signaling
to regulate fibroblast activation and fibrosis. Nat Commun.
9:32592018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dardaei L, Wang HQ, Singh M, Fordjour P,
Shaw KX, Yoda S, Kerr G, Yu K, Liang J, Cao Y, et al: SHP2
inhibition restores sensitivity in ALK-rearranged non-small-cell
lung cancer resistant to ALK inhibitors. Nat Med. 4:512–517. 2018.
View Article : Google Scholar
|