Introduction
Fracture healing is a multi-level and multi-path
process regulated by systemic and local factors, involving numerous
types of cells and growth factors, such as mesenchymal cells and
transforming growth factor-β1 (TGF-β1) (1–3).
Fracture non-union or delayed union is a common complication in
orthopedics that causes physical and mental pain, as well as a
notable economic burden to patients and society. Long-term clinical
studies have revealed that, in the process of fracture healing,
callus volume and formation rate are higher in patients with
fractures combined with traumatic brain injury than in patients
with simple fractures. Heterotopic ossification occurs in patients
with craniocerebral trauma combined with a fracture, and fracture
healing is accelerated (4,5). Numerous cytokines and neuropeptide
factors, such as bone morphogenetic protein (BMP), TGF-β1, vascular
endothelial growth factor, insulin-like growth factor, fibroblast
growth factor and calcitonin gene-related peptides, can
significantly promote osteoblast proliferation, thereby
accelerating fracture healing (6–9).
Although there have been advances in the study of osteoblasts in
accelerated fracture healing, the underlying mechanisms remain
unclear. Elucidating the mechanism by which fracture healing is
accelerated may facilitate the development of effective clinical
treatments for patients with delayed fracture healing.
During craniocerebral injury, hypoxia promotes the
expression of hypoxia-inducible factor-1α (HIF-1α), which can
increase cell viability, and promote adhesion, migration and
angiogenesis. HIF-1α is the most important hypoxia receptor and
hypoxia-induced transcription factor known at present (10,11).
An increase in HIF-1α activity can increase the viability of cells
in hypoxic environments and promote cell adhesion, migration and
angiogenesis (12,13). HIF-1α can promote stem cell
migration to ischemic and hypoxic sites by regulating the
expression levels of surface molecules as a result of interaction
between a number of ligands and receptors including stromal
cell-derived factor-1 (SDF-1), a downstream gene of HIF-1α, which
binds to its specific receptor C-X-C chemokine receptor type 4
(CXCR4) to form a pair of coupling molecules (14,15).
SDF-1 is a CXC-type chemokine produced by mesenchymal stem cells
(MSCs). SDF-1 and its unique receptor CXCR4 constitute the
biological axis of SDF-1/CXCR4 (16). SDF-1 and CXCR4 are expressed in
numerous types of cells and tissues, where they serve a key role in
tissue repair and a variety of physiological and pathological
processes, including organogenesis, revascularization and response
to tissue injury (17,18). However, their role in the process
of accelerated fracture healing during craniocerebral injury and
the regulatory effect of HIF-1α on SDF-1/CXCR4 in MSCs remains
unclear. In the present study, MSCs, which serve a key role in
fracture healing, were used as a model to investigate the role of
the HIF-1α/SDF-1/CXCR4 signaling axis in accelerated fracture
healing during craniocerebral injury in vitro.
Materials and methods
Mouse bone marrow mesenchymal stem
cell (BMSC) culture
A total of 10 C57 mice (male; age, 4 weeks; weight,
18–20 g) were purchased from Shanghai SLAC Laboratory Animal Co.,
Ltd. Mice were housed in specific-pathogen-free conditions at room
temperature (22±1°C) with relative humidity (50±5%), 12-h
light/dark cycles, and free access to food and water. BMSCs were
isolated as previously described (19). C57 mice were sacrificed via
cervical dislocation and soaked in 75% alcohol for 10 min at room
temperature. Under sterile conditions, the bilateral leg bones were
extracted and separated from the surrounding muscle. The harvested
tissue was placed in cold complete DMEM/F12 (DMEM/F12+10% FBS+1%
penicillin/streptomycin/amphotericin B solution; Gibco; Thermo
Fisher Scientific, Inc.). The culture medium was aspirated
repeatedly with a needle and syringe to rinse the marrow cavity.
Cells were dispersed by repeated pipetting, and the cell suspension
was passed through a 400-mesh sterile mesh and centrifuged at 200 ×
g for 3 min, after which the supernatant was discarded. The cells
were resuspended in complete DMEM/F12, and the density was adjusted
to 5×105 cells/ml. Subsequently, the cell suspension (4
ml) was inoculated into a T-25 flask and cultured in a 5%
CO2 incubator at 37°C. After 24 h, the flask was shaken
horizontally to suspend unattached cells and the supernatant was
discarded. Fresh medium was then added to the culture flask and
changed every 2–3 days until the primary cells grew to a confluence
of 80–90%.
Cells were then sub-cultured as follows: Supernatant
media was aspirated, and the cells were washed with 1 ml PBS and
treated with 1 ml 0.25% trypsin for 2 min at 37°C. Cell morphology
was observed by light microscopy (magnification, ×100). When the
cells were rounded, 1 ml complete medium was added to terminate
digestion, and the cells were gently pipetted to detach them from
the bottom of the bottle completely. The cell suspension was
transferred to a centrifuge tube and centrifuged at 200 × g for 3
min at room temperature, and the supernatant was removed. The cells
were resuspended in complete DMEM/F12 and inoculated into a new
culture flask at a ratio of 1:2. After passage, the cells were
cultured in the same manner as the primary culture until the next
generation.
Flow cytometry and MSC sorting
Third generation MSC cells were digested with 0.25%
trypsin for 2 min at 37°C, and a single cell suspension was
obtained by adjusting the cell density to 1×106
cells/ml. The cell suspension was incubated for 30 min at room
temperature with PBS, anti-CD29-FITC (1.5 µl; cat. no. ab21845;
Abcam), anti-CD44-FITC (2 µg; cat. no. ab25064; Abcam) or
anti-CD45-FITC (10 µl; cat. no. ab27287; Abcam), and analyzed using
a Coulter Epics XL-MCL flow cytometer (BD Biosciences) with EXPO32
software (version 1.2; BD Biosciences).
HIF-1α siRNA transfection
MSC cells were seeded in 6-well culture plates
(5×105 cells/well) overnight. The inoculated cells were
washed twice with PBS, and 500 µl serum-free DMEM/F12 (Gibco;
Thermo Fisher Scientific, Inc.) was added. The HilyMax-HIF-1α siRNA
transfection mixture (Dojindo Molecular Technologies, Inc.) and
siRNA control were premixed (0.2 µg/µl), added to the MSCs and
mixed. siRNA and siRNA control were obtained from Sangon Biotech
Co., Ltd. After incubating the cells in the cell culture incubator
for 6 h at 37°C, the medium was replaced with fresh medium. The
cells were trypsinized and collected for subsequent experiments
after transfection for 48 h. The sequences of siRNAs used are
presented in Table I.
 | Table I.Sequences of siRNA. |
Table I.
Sequences of siRNA.
| siRNA | Sequence (5′→3′) |
|---|
| HIF-1α-Mus-762 | F:
CUGAUAACGUGAACAAAUATT |
|
| R:
UAUUUGUUCACGUUAUCAGTT |
| HIF-1α-Mus-1102 | F:
CAUUCCUCAUCCGUCAAAUTT |
|
| R:
AUUUGACGGAUGAGGAAUGTT |
|
HIF-1α-Mus-1238 | F:
GGCCGCUCAAUUUAUGAAUTT |
|
| R:
AUUCAUAAAUUGAGCGGCCTT |
Reverse transcription-quantitative
(RT-q) PCR analysis
Total RNA was extracted from cells with TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.). cDNA synthesis was
performed using the TOYOBO ReverTra Ace qPCR RT kit, according to
the manufacturer's protocol. qPCR was performed using the KAPA
SYBR-Green Supermix PCR kit (Kapa Biosystems). RT-qPCR primers were
obtained from Sangon Biotech Co., Ltd.; sequences are listed in
Table II. The reaction was
started at 95°C for 5 min, followed by 40 cycles of 95°C for 30
sec, 61°C for 30 sec, and 72°C for 30 sec. Relative gene expression
levels were measured using the cycle threshold values and the
2−ΔΔCq method (20).
| Table II.Sequences of reverse
transcription-quantitative PCR primers. |
Table II.
Sequences of reverse
transcription-quantitative PCR primers.
| Gene | Sequence
(5′→3′) |
|---|
| Stromal
cell-derived | F:
GCATCAGTGACGGTAAACCA |
| factor-1 | R:
TCTTCAGCCGTGCAACAATC |
| C-X-C
chemokine | F:
CTAAGGAGCATGACGGACAA |
| receptor type
4 | R:
ATTTCCCAAAGTACCAGTCAGC |
| GAPDH | F:
TGACCTCAACTACATGGTCTACA |
|
| R:
CTTCCCATTCTCGGCCTTG |
Western blotting and antibodies
Cells treated with siRNA were harvested for protein
extraction using the RIPA reagent (Pierce; Thermo Fisher
Scientific, Inc.). The protein concentration was determined via BCA
protein assay reagent. After measuring protein concentration,
50–100 µg protein was subjected to 10% SDS-PAGE gel and transferred
to a PVDF membrane. The membrane was blocked with 5% non-fat milk
powder for 1 h at room temperature and incubated with primary
antibodies against HIF-1α (1:1,000; cat. no. ab179483; Abcam),
SDF-1 (1:1,000; cat. no. ab25117; Abcam), and CXCR4 (1:1,000; cat.
no. ab124824; Abcam) at 4°C overnight, followed by goat-anti-rabbit
(1:2,000; cat. no. ab205718; Abcam) secondary antibody for 1 h at
37°C. The chemiluminescent signaling was detected via ECL reagents
(Pierce; Thermo Fisher Scientific, Inc.).
Cell proliferation assay
Cell proliferation was assessed using Cell Counting
Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.). Briefly, MSCs
were seeded on 96-well microplates at a density of 1×104
cells/well. The cells were transfected with HilyMax-siRNA and
cultured for 0, 24 or 48 h at 37°C. A total of 5 µl CCK-8 solution
was then added to each well and incubated at 37°C for an additional
2 h. Optical density (OD) was determined at a wavelength of 450
nm.
Migration assay
Migration assays were performed using 24-well plate
with permeable Transwell inserts (pore size 0.4 mm; Corning, Inc.).
BMSCs were suspended in serum-free DMEM/F12 medium
(5×104 cells/well) and equal amounts of cell suspension
(200 µl) was added to the upper chambers on the migration plate.
The same amount (200 µl) of complete medium (DMEM/F12+10% FBS) with
different treatment was added to the lower chamber of the migration
plate separately. There were two sets of experiments. In the first,
cells were divided into the following treatment groups for 24 h at
37°C: 0 ng/ml SDF-1+0 µM AMD3100 (a specific antagonist that blocks
the binding of SDF-1 to CXCR4; cat. no. A5602; Sigma-Aldrich; Merck
KGaA); 10 ng/ml SDF-1+0 µM AMD3100; 100 ng/ml SDF-1+0 µM AMD3100;
100 ng/ml SDF-1+1 µM AMD3100; and 100 ng/ml SDF-1+10 µM AMD3100.
For the second experiment, cells were divided into the following
treatment groups for 24 h at 37°C: NC (siRNA-NC), siRNA
(HIF-1α-siRNA), SDF-1 (siRNA-NC+100 ng/ml SDF-1), and siRNA and
SDF-1 group (HIF-1α siRNA+100 ng/ml SDF-1). After 24 h, the
migrated cells on the bottom of the insert were fixed with 4%
paraformaldehyde solution for 24 h at room temperature and stained
with crystal violet (1%) for 15 min at room temperature. The cells
were processed as described in the cell migration assay, and
migrated cells were counted, and the relative number was calculated
with light microscopy (magnification, ×100).
Statistical analysis
Results are representative of three independent
experiments. GraphPad Prism (version 7.0; GraphPad Software, Inc.)
and SPSS software (version 19; IBM Corp.) were used for statistical
analysis of cell proliferation, as well as the results of the
Transwell assay and RT-qPCR. Differences between two groups were
analyzed using an unpaired t-test. Comparisons between multiple
groups were analyzed using one-way ANOVA followed by Tukey's post
hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Morphological identification of
MSCs
The present study determined whether the
HIF-1α/SDF-1/CXCR4-axis serves a role in fracture combined with
traumatic brain injury in an in vitro BMSC model. MSCs
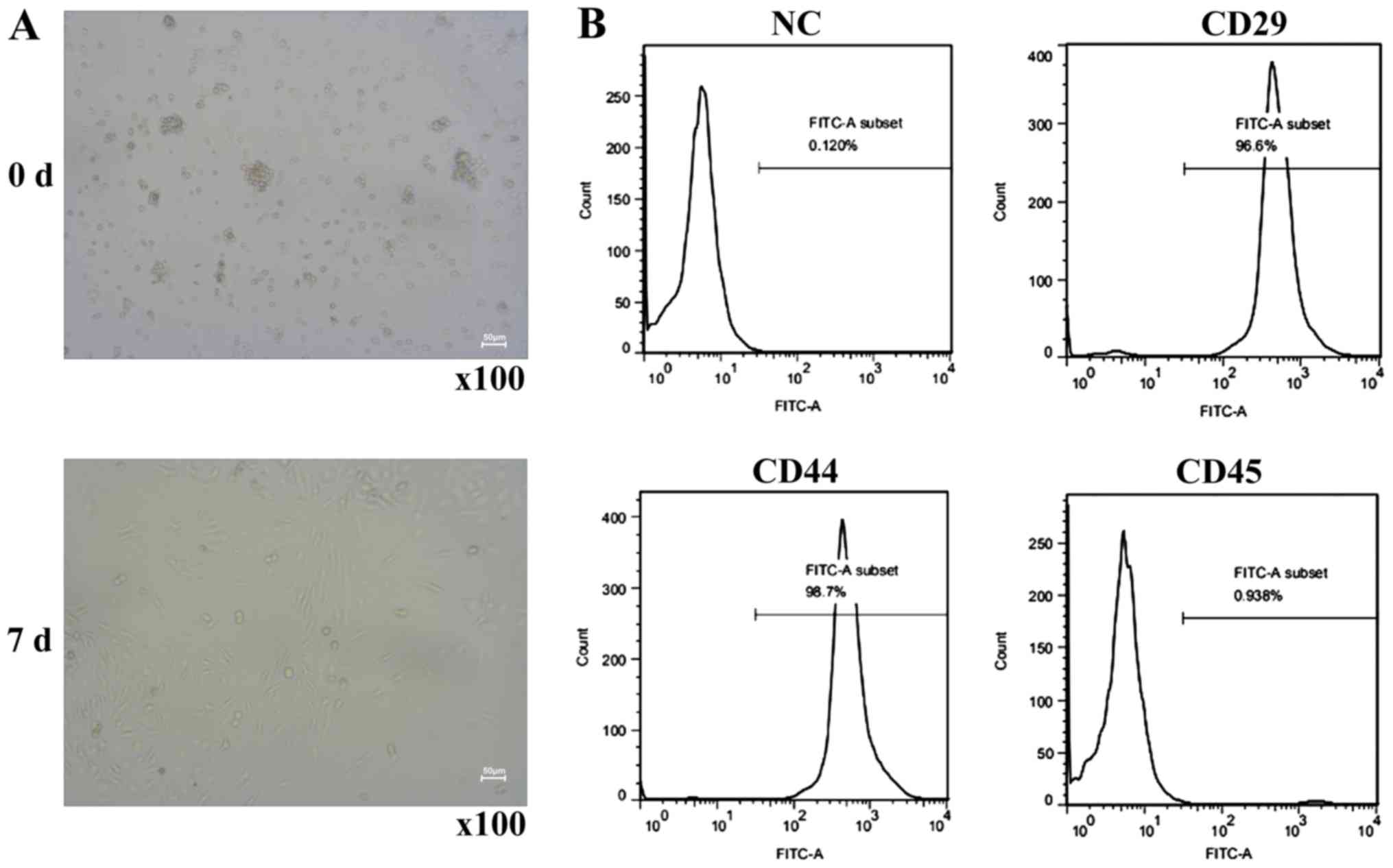
exhibited a typical spindly morphology (Fig. 1A). Cells in the third passage were
characterized via flow cytometry. The results demonstrated that the
expression levels of CD29 and CD44 on the surface of BMSCs were
>95%, whereas the expression level of the hematopoietic stem
cell marker CD45 was <5% (Fig.
1B), which was consistent with the characteristics of
BMSCs.
HIF-1α knockdown suppresses MSC
proliferation in vitro
Next, the effects of HIF-1α on the migration of MSCs
were investigated. In order to assess the effect of HIF-1α
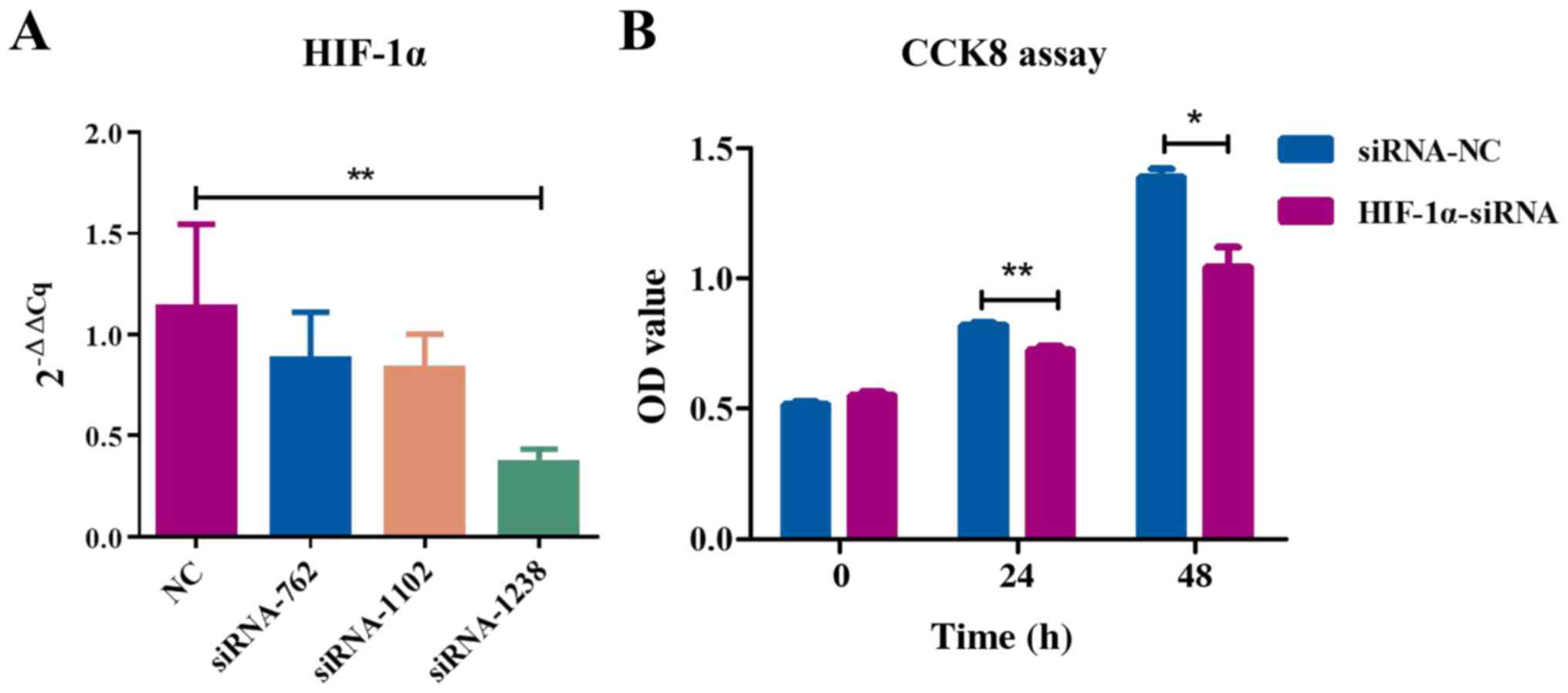
knockdown on cell proliferation, CCK-8 was used to detect
proliferation at different time-points. The OD value of
HIF-1α-knockdown cells was significantly decreased at 450 nm
(Fig. 2), indicating that HIF-1α
knockdown inhibits MSC proliferation.
SDF-1 promotes cell migration by
binding to CXCR4
In order to confirm the effect of SDF-1 in MSCs, its
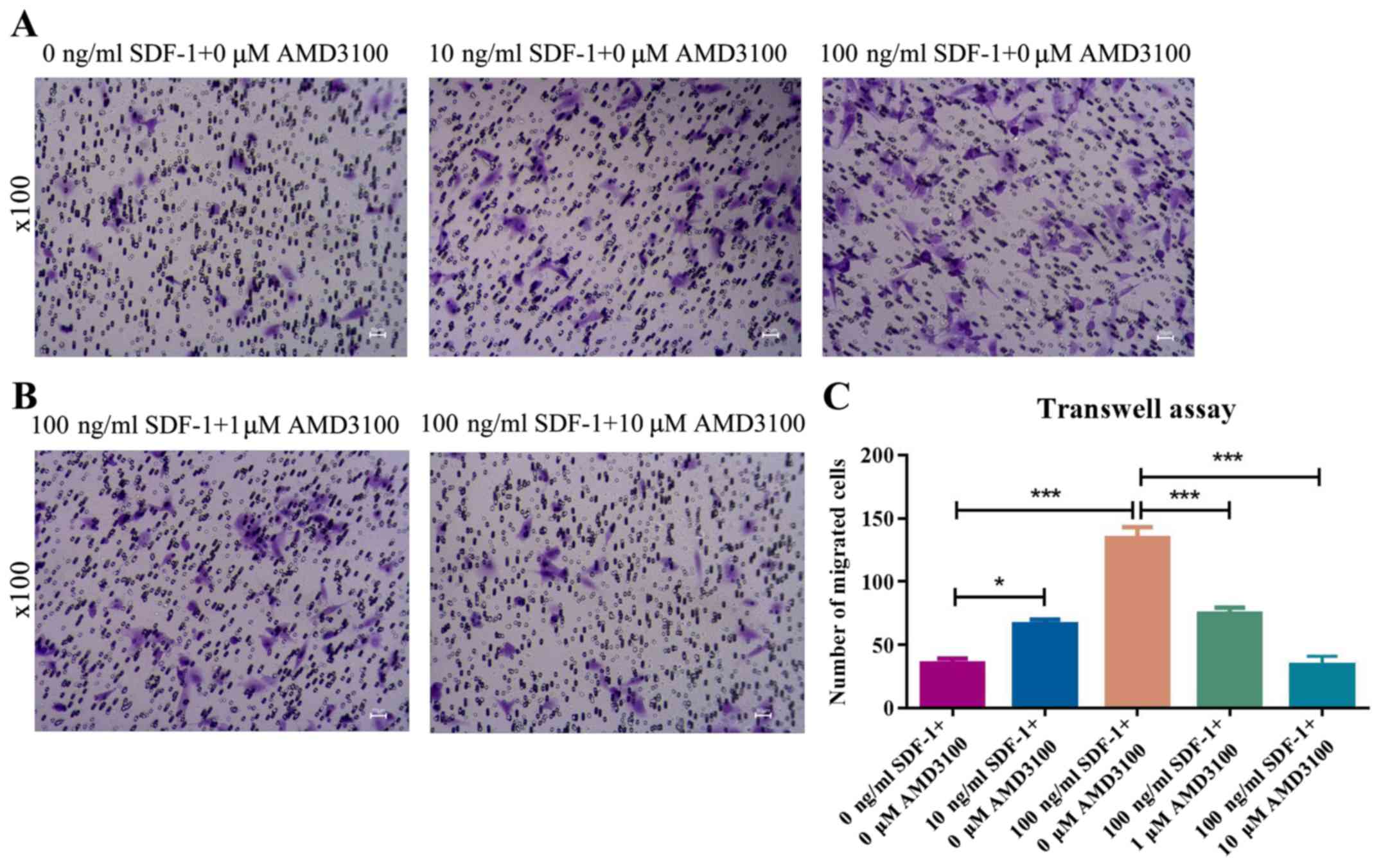
impact on cell migration was analyzed. Treatment of MSCs with
various concentrations of SDF-1 caused a dose-dependent increase in
cell migration (Fig. 3A). Cell
migration induced by SDF-1 was abrogated by AMD3100, indicating
that AMD3100 inhibits MSC migration via obstructing SDF-1 binding
with CXCR4 (Fig. 3B).
Collectively, these results indicated that SDF-1 promoted MSC
migration via binding to CXCR4, which was impaired by AMD3100.
HIF-1α knockdown inhibits MSC
migration via SDF-1/CXCR4 signaling
It was then determined whether knockdown of HIF-1α
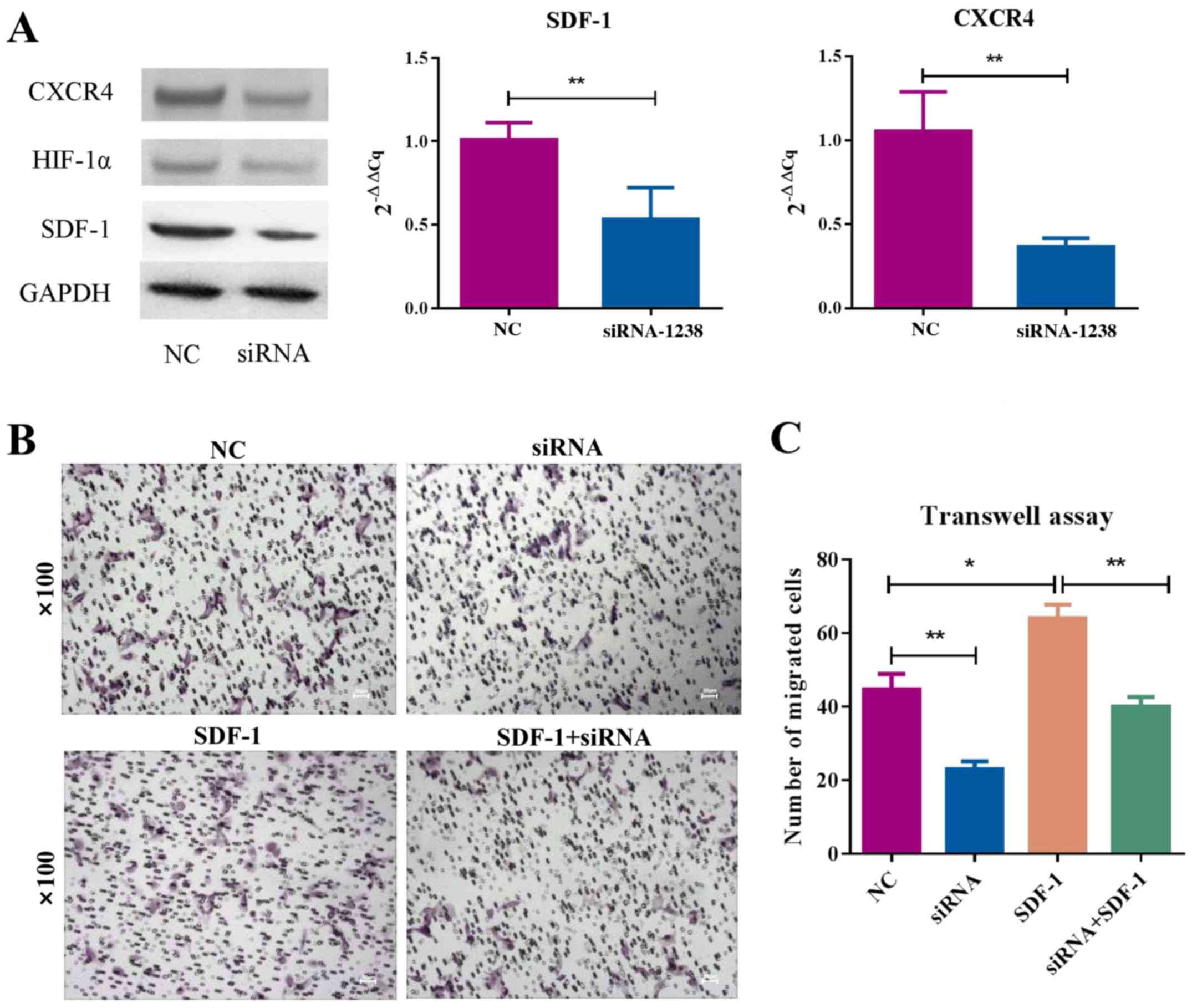
impairs the chemotactic function of SDF-1/CXCR4 signaling in MSCs.
The mRNA and protein levels of SDF-1 and CXCR4 were decreased in
MSCs stably transfected with siHIF-1α compared with NC group
(Fig. 4A). Moreover, HIF-1α
knockdown inhibited MSC migration; replenishment of SDF-1 increased
cell migration (Fig. 4B and C).
Thus, HIF-1α likely regulates MSC migration via SDF-1/CXCR4
signaling.
Discussion
In the present study, the proliferation rate of MSCs
transfected with HIF-1α siRNA significantly decreased at 24 and 48
h. MSCs are essential for the repair of bone defects, and improving
the effects of MSCs in bone is of clinical interest. In patients
with skull injuries, ischemia and hypoxia surrounding brain tissue
often occur, and the skull bone repair process must be adapted to
the hypoxic environment (10,21).
HIF-1α can maintain homeostasis of oxygen under hypoxia, allowing
the body to adapt to hypoxia (22). HIF-1α can induce the formation of
blood vessels and bone tissue via numerous factors, including
vascular endothelial growth factor, angiopoietin, platelet growth
factor and transforming growth factor, making it a potential
candidate for the treatment of bone defects and other types of
disease (23,24). HIF-1α is a regulator of
BMP2-induced chondrogenic differentiation, osteogenic
differentiation and endochondral bone formation (25). HIF-1α and BMP2 have been revealed
to synergistically induce MSC differentiation and promote the
expansion of the proliferating chondrocyte zone (25). HIF-1 also serves an important role
in coupling osteogenesis and angiogenesis during skeletal
development and bone regeneration, and HIF-1α overexpression may
represent a therapeutic option to improve cellular functions of
MSCs in the treatment of critical-sized bone defects (26). Thus, HIF-1α may improve the
cellular functions of MSCs; however, the underlying molecular
mechanisms are still unclear.
SDF-1 is a protein secreted primarily by MSCs. When
SDF-1 specifically binds to CXCR4 on a number of types of cell
surface, activated SDF-1/CXCR4 can stimulate downstream signaling
pathways associated with the regulation of stem cell mobilization
and migration (27), induce MSC
migration to the damaged site and participate in injury repair
(28,29). Activation of the ERK-AKT pathway
mediates SDF1-induced cell migration. SDF1/CXCR4 signaling is
required for MSC homing and retention to their niche in the bone
marrow (30). Secretion of SDF-1
has been revealed to increase during bone regeneration, allowing
CXCR4-expressing MSCs to enter the stretch region with the
concentration gradient of SDF-1 following induction via the
SDF-1/CXCR4 molecular axis, and to participate in new bone
formation and cardiovascular generation (31–33).
In order to confirm the effect of SDF-1 in MSCs, the present study
analyzed the chemotactic effect of the SDF-1/CXCR4 axis on MSCs.
SDF-1 was demonstrated to promote the migration of MSCs in a
dose-dependent manner. This result is similar to that reported by
Zhou et al (25), who
demonstrated that HIF-1α can regulate cell migration by influencing
the expression levels of CXCR4. Additionally, by using AMD3100 to
block the binding of SDF-1 to CXCR4, SDF-1-induced cell migration
in the present study was abrogated, indicating that AMD3100
inhibits MSC migration by obstructing SDF-1 binding with CXCR4.
Therefore, the SDF-1/CXCR4 axis serves a key role in MSC
function.
SDF-1/CXCR4 signaling has been revealed to play an
important role in cell migration, chemotaxis as well as other
biological behaviors. SDF-1-induced transendothelial behavior has
been revealed to be positively associated with the density of cell
surface receptor CXCR4 and is affected by a number of factors
(34). HIF-1α is a central
transcription factor of hypoxia-specific gene expression levels
(30). HIF-1α can bind to the
SDF-1 promoter and specifically regulate the expression levels of
SDF-1 (30,35). Hypoxia can upregulate the
expression levels of CXCR4 in human monocytes and macrophages, as
well as endothelial and tumor cells, via the activation of HIF-1α
(36). The hypoxia/HIF-1α/CXCR4
pathway may be a mechanism of regulating the migration of different
types of cells under hypoxia (37). A number of CXCR4-positive stem
cells are involved in angiogenesis in locally damaged areas in a
hypoxic environment. The adhesion, migration and homing of these
CXCR4-positive stem cells are initiated via binding of SDF-1,
containing hypoxia response elements, to CXCR4; the expression
levels of SDF-1 are regulated by HIF-1α (38). The present study therefore
determined whether HIF-1α can recruit MSCs and promote osteogenesis
via the SDF-1/CXCR4 molecular axis. Following HIF-1α knockdown, it
was demonstrated that the mRNA and protein levels of SDF-1 and
CXCR4 in MSCs were significantly decreased. HIF-1α gene silencing
inhibited the migration of MSCs (P<0.05). This confirmed that
SDF-1 promotes the migration of MSC cells and indicated that HIF-1α
acts on MSCs via the SDF-1/CXCR4 molecular axis. However, further
investigation is required to elucidate the mechanisms underlying
the effect of HIF-1α on oxidative stress in the healing process of
skull injury, which may involve complex signaling pathways.
During craniocerebral injury healing, the hypoxic
state of surrounding brain tissue induces an increase in the
secretion of HIF-1α (39), which
accelerates the fracture repair process via chemotaxis due to the
SDF-1/CXCR4 axis. In the present study, a gene silencing plasmid
was successfully constructed based on the HIF-1α gene sequence, and
the regulatory association between HIF-1α and SDF-1/CXCR4 was
elucidated. Silencing of HIF-1α decreased MSC migration, as well as
the mRNA and protein levels of SDF-1 and CXCR4 in MSCs. Due to the
wide distribution and diversity of roles of SDF-1 and CXCR4, they
may represent prognostic biomarkers or therapeutic targets for a
number of neurological diseases.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Shanghai
Excellent Young Medical Talents Training Program (grant no.
2018YQ46), the National Natural Science Foundation of China (grant
no. 81501897), the Shanghai Municipal Commission of Health and
Family Planning of Science and Research Fund (grant no. 20154Y0070)
and K.C. Wong Education Foundation.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WX and YX conceived the study. WX, YX, ZL, RH and XZ
performed the experiments and contributed to writing the
manuscript. WX, YX and YW analyzed the data. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Animal Research Ethics
Committee of TongRen Hospital, School of Medicine, Shanghai
JiaoTong University (approval no. 170004).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
van Baardewijk LJ, van der Ende J,
Lissenberg-Thunnissen S, Romijn LM, Hawinkels LJA, Sier CFM and
Schipper IB: Circulating bone morphogenetic protein levels and
delayed fracture healing. Int Orthop. 37:2767–527. 2013. View Article : Google Scholar
|
|
2
|
Hara Y, Ghazizadeh M, Shimizu H, Matsumoto
H, Saito N, Yagi T, Mashiko K, Mashiko K, Kawai M and Yokota H:
Delayed expression of circulating TGF-β1 and BMP-2 levels in human
nonunion long bone fracture healing. J Nippon Med Sch. 84:12–18.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hankenson KD, Zimmerman G and Marcucio R:
Biological perspectives of delayed fracture healing. Injury. 45
(Suppl 2):S8–S15. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Seemann R, Graef F, Garbe A, Keller J,
Huang F, Duda G, Schmidt-Bleek K, Schaser KD and Tsitsilonis S:
Leptin-deficiency eradicates the positive effect of traumatic brain
injury on bone healing: Histological analyses in a combined trauma
mouse model. J Musculoskelet Neuronal Interact. 18:32–41.
2018.PubMed/NCBI
|
|
5
|
Huang H, Cheng WX, Hu YP, Chen JH, Zheng
ZT and Zhang P: Relationship between heterotopic ossification and
traumatic brain injury: Why severe traumatic brain injury increases
the risk of heterotopic ossification. J Orthop Translat. 12:16–25.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu S, Liu Y, Jiang L, Li Z, Lee S, Liu C,
Wang J and Zhang J: Recombinant human BMP-2 accelerates the
migration of bone marrow mesenchymal stem cells via the
CDC42/PAK1/LIMK1 pathway in vitro and in vivo. Biomater Sci.
7:362–372. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Manzano-Moreno FJ, Medina-Huertas R,
Ramos-Torrecillas J, García-Martínez O and Ruiz C: The effect of
low-level diode laser therapy on early differentiation of
osteoblast via BMP-2/TGF-β1 and its receptors. J Craniomaxillofac
Surg. 43:1926–1932. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang T, Guo S and Zhang H: Synergistic
effects of controlled-released BMP-2 and VEGF from nHAC/PLGAs
scaffold on osteogenesis. BioMed Res Int.
2018:35164632018.PubMed/NCBI
|
|
9
|
Quarles LD: Fibroblast growth factor 23
and α-Klotho co-dependent and independent functions. Curr Opin
Nephrol Hypertens. 28:16–25. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wan C, Shao J, Gilbert SR, Riddle RC, Long
F, Johnson RS, Schipani E and Clemens TL: Role of HIF-1alpha in
skeletal development. Ann N Y Acad Sci. 1192:322–326. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tirpe AA, Gulei D, Ciortea SM, Crivii C
and Berindan-Neagoe I: Hypoxia: Overview on hypoxia-mediated
mechanisms with a focus on the role of HIF genes. Int J Mol Sci.
20:61402019. View Article : Google Scholar
|
|
12
|
Kanichai M, Ferguson D, Prendergast PJ and
Campbell VA: Hypoxia promotes chondrogenesis in rat mesenchymal
stem cells: A role for AKT and hypoxia-inducible factor
(HIF)-1alpha. J Cell Physiol. 216:708–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim A and Ma JY: Rhaponticin decreases the
metastatic and angiogenic abilities of cancer cells via suppression
of the HIF1α pathway. Int J Oncol. 53:1160–1170. 2018.PubMed/NCBI
|
|
14
|
Luo Y, Cai J, Xue H, Miura T and Rao MS:
Functional SDF1 alpha/CXCR4 signaling in the developing spinal
cord. J Neurochem. 93:452–462. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Knerlich-Lukoschus F, von der Ropp-Brenner
B, Lucius R, Mehdorn HM and Held-Feindt J: Spatiotemporal CCR1,
CCL3(MIP-1α), CXCR4, CXCL12(SDF-1α) expression patterns in a rat
spinal cord injury model of posttraumatic neuropathic pain. J
Neurosurg Spine. 14:583–597. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Merino JJ, Gutiérrez-Fernández M,
Rodríguez-Frutos B, Alvarez-Grech J, Alcalde ME, Vallejo-Cremades
MT and Díez-Tejedor E: CXCR4/SDF-1α-chemokine regulates
neurogenesis and/or angiogenesis within the vascular niche of
ischemic rats; however, does SDF-1α play a role in repair? Int J
Stroke. 6:466–467. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Petit I, Jin D and Rafii S: The
SDF-1-CXCR4 signaling pathway: A molecular hub modulating
neo-angiogenesis. Trends Immunol. 28:299–307. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wen J, Zhang JQ, Huang W and Wang Y:
SDF-1α and CXCR4 as therapeutic targets in cardiovascular disease.
Am J Cardiovasc Dis. 2:20–28. 2012.PubMed/NCBI
|
|
19
|
Liu X, Zheng L, Zhou Y, Chen Y, Chen P and
Xiao W: BMSC transplantation aggravates inflammation, oxidative
stress, and fibrosis and impairs skeletal muscle regeneration.
Front Physiol. 10:872019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Knight MN and Hankenson KD: Mesenchymal
stem cells in bone regeneration. Adv Wound Care (New Rochelle).
2:306–316. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yuan G, Peng YJ, Reddy VD, Makarenko VV,
Nanduri J, Khan SA, Garcia JA, Kumar GK, Semenza GL and Prabhakar
NR: Mutual antagonism between hypoxia-inducible factors 1α and 2α
regulates oxygen sensing and cardio-respiratory homeostasis. Proc
Natl Acad Sci USA. 110:E1788–E1796. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Maes C: Signaling pathways effecting
crosstalk between cartilage and adjacent tissues: Seminars in cell
and developmental biology: The biology and pathology of cartilage.
Semin Cell Dev Biol. 62:16–33. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Johnson RW, Schipani E and Giaccia AJ: HIF
targets in bone remodeling and metastatic disease. Pharmacol Ther.
150:169–177. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhou N, Hu N, Liao JY, Lin LB, Zhao C, Si
WK, Yang Z, Yi SX, Fan TX, Bao W, et al: HIF-1α as a regulator of
BMP2-induced chondrogenic differentiation, osteogenic
differentiation, and endochondral ossification in stem cells. Cell
Physiol Biochem. 36:44–60. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lampert FM, Kutscher C, Stark GB and
Finkenzeller G: Overexpression of Hif-1α in mesenchymal stem cells
affects cell-autonomous angiogenic and osteogenic parameters. J
Cell Biochem. 117:760–768. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kucia M, Ratajczak J, Reca R,
Janowska-Wieczorek A and Ratajczak MZ: Tissue-specific muscle,
neural and liver stem/progenitor cells reside in the bone marrow,
respond to an SDF-1 gradient and are mobilized into peripheral
blood during stress and tissue injury. Blood Cells Mol Dis.
32:52–57. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sun J, Zhao Y, Li Q, Chen B, Hou X, Xiao Z
and Dai J: Controlled release of collagen-binding SDF-1α improves
cardiac function after myocardial infarction by recruiting
endogenous stem cells. Sci Rep. 6:266832016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ji JF, He BP, Dheen ST and Tay SS:
Interactions of chemokines and chemokine receptors mediate the
migration of mesenchymal stem cells to the impaired site in the
brain after hypoglossal nerve injury. Stem cells. 22:415–427. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yu Q, Liu L, Lin J, Wang Y, Xuan X, Guo Y
and Hu S: SDF-1α/CXCR4 axis mediates the migration of mesenchymal
stem cells to the hypoxic-ischemic brain lesion in a rat model.
Cell J. 16:440–447. 2015.PubMed/NCBI
|
|
31
|
Kortesidis A, Zannettino A, Isenmann S,
Shi S, Lapidot T and Gronthos S: Stromal-derived factor-1 promotes
the growth, survival, and development of human bone marrow stromal
stem cells. Blood. 105:3793–3801. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Periyasamy-Thandavan S, Burke J, Mendhe B,
Kondrikova G, Kolhe R, Hunter M, Isales CM, Hamrick MW, Hill WD and
Fulzele S: MicroRNA-141-3p negatively modulates SDF-1 expression in
age-dependent pathophysiology of human and murine bone marrow
stromal cells. J Gerontol A Biol Sci Med Sci. 74:1368–1374. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kristocheck M, Dias LD, Ghem C, Eibel B,
Kalil RAK and Markoski MM: The homing of bone marrow stem cells is
differentially activated in ischemic and valvular heart diseases
and influenced by beta-blockers. J Transl Med. 16:1332018.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
van Buul JD, Voermans C, van Gelderen J,
Anthony EC, van der Schoot CE and Hordijk PL: Leukocyte-endothelium
interaction promotes SDF-1-dependent polarization of CXCR4. J Biol
Chem. 278:30302–30310. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sun X, Wei L, Chen Q and Terek RM:
CXCR4/SDF1 mediate hypoxia induced chondrosarcoma cell invasion
through ERK signaling and increased MMP1 expression. Mol Cancer.
9:172010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu H, Liu S, Li Y, Wang X, Xue W, Ge G
and Luo X: The role of SDF-1-CXCR4/CXCR7 axis in the therapeutic
effects of hypoxia-preconditioned mesenchymal stem cells for renal
ischemia/reperfusion injury. PLoS One. 7:e346082012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Schioppa T, Uranchimeg B, Saccani A,
Biswas SK, Doni A, Rapisarda A, Bernasconi S, Saccani S, Nebuloni
M, Vago L, et al: Regulation of the chemokine receptor CXCR4 by
hypoxia. J Exp Med. 198:1391–1402. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ceradini DJ, Kulkarni AR, Callaghan MJ,
Tepper OM, Bastidas N, Kleinman ME, Capla JM, Galiano RD, Levine JP
and Gurtner GC: Progenitor cell trafficking is regulated by hypoxic
gradients through HIF-1 induction of SDF-1. Nat Med. 10:858–864.
2004. View
Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu X, Zhou C, Li Y, Ji Y, Xu G, Wang X
and Yan J: SDF-1 promotes endochondral bone repair during fracture
healing at the traumatic brain injury condition. PLoS One.
8:e540772013. View Article : Google Scholar : PubMed/NCBI
|