Introduction
According to 2018 statistics by The World Cancer
Research Fund, endometrial cancer (EC) is the 15th most common
malignant tumor worldwide, the incidence rate of which is still on
the rise (1). Patients with EC are
often diagnosed when the disease is still confined to the uterus
(2). Standard treatment consists
of primary hysterectomy and bilateral salpingo-oophorectomy, often
using minimally invasive approaches (laparoscopic or robotic). The
5-year overall survival rate ranges between 74 and 91% in patients
with EC without metastatic disease (2), whereas patients with stage III or IV
EC exhibit 5-year overall survival rates of 57–65 and 20–26%,
respectively (3). Numerous
patients with EC suffer from cancer recurrence and eventually
succumb to the disease, and patients with EC relapse have poor
prognosis (4). Unfortunately,
effective treatments are still rare for patients with advanced and
relapsed EC; therefore, there is an urgent requirement to develop
new EC treatment strategies.
Increasing evidence has suggested that long
non-coding RNAs (lncRNAs) and microRNAs (miRNAs/miRs) play
important roles in the occurrence and development of several types
of human cancer by affecting the malignant phenotypes of tumor
cells, such as proliferation, invasion and apoptosis, including
with breast, lung and ovarian cancer (5–8). In
carcinoma cells, lncRNAs and miRNA can function as tumor
suppressors or oncogenes. A recent study demonstrated that
increased expression of the lncRNA HOX transcript antisense RNA may
promote cell proliferation and inhibit apoptosis by binding to
phosphatase and tensin homolog in EC, suggesting the important
roles that lncRNAs serve in EC (9). Previous studies have demonstrated
that small nucleolar RNA host gene 12 (SNHG12), a lncRNA
transcribed from human chromosome 1 and located at the p35.3, is
overexpressed in a few types of human cancer (10–12).
SNHG12 has been reported to enhance the proliferative and migratory
abilities of ovarian cancer cells by sponging miR-129 to upregulate
SOX4 (13). It has also been
suggested to play a pivotal role in the development of cervical
cancer by sponging miR-125b (14).
Recently, miRNAs have been demonstrated to be involved in the
malignant behaviors of cancer by regulating cell growth and
metastasis, including miR-4429 (15). lncRNA LINC00313 has been reported
to modulate papillary thyroid cancer tumorigenesis by sponging
miR-4429. In addition, miR-4429 can repress tumor progression and
epithelial-mesenchymal transition by targeting cyclin-dependent
kinase 6 (CDK6) in clear cell renal cell carcinoma, sensitize
cervical cancer cells to irradiation by downregulating RAD51
recombinase (RAD51), and prevent gastric cancer progression by
targeting methyltransferase like 3, thus suppressing m6A-induced
stabilization of SEC62 homolog, preprotein translocation factor
(16). However, it is not clear
whether SNHG12 and miR-4429 act as oncogenes or anti-oncogenes in
EC. Therefore, the present study aimed to examine the biological
function and possible molecular mechanism of SNHG12 and miR-4429 in
the tumorigenesis of EC.
Materials and methods
Cell culture
The human EC cell lines Ishikawa, KLE, RL95-2 and
AN3 CA were purchased from Shanghai Cell Collection and the human
normal endometrial stromal cell line ESC (BNCC340262) was obtained
from BeNa Culture Collection. Cells were cultured in DMEM
(Sigma-Aldrich; Merck KGaA) supplemented with 15% FBS
(Sigma-Aldrich; Merck KGaA) at 37°C and 5% CO2.
Cell transfection
The sequences of short hairpin RNAs (shRNAs)
targeting SNHG12 (shRNA-SNHG12-1 and shRNA-SNHG12-2) and their
negative control (NC) sequence (shRNA-NC), were designed and
synthesized by Shanghai GeneChem Co., Ltd, and cloned into the
pSUPER-retro-puromycin plasmids (cat. no. VEC-pRT-0002;
OligoEngine), according to the manufacturer's instructions. The
SNHG12 overexpression plasmid (OV) was established by inserting the
full-length human SNHG12 cDNA (Aksomics Inc.) into the pcDNA3.1
vector (Invitrogen; Thermo Fisher Scientific, Inc.), whereas an
empty vector served as the NC (OV-NC). miR-4429 mimics (cat. no.
miR10018944-1-5) and corresponding NC mimics (mimic-NC; cat. no.
miR1N0000002-1-5) were purchased from Guangzhou RiboBio Co., Ltd.
After RL95-2 cells (1×106) were transfected with 50 nM
plasmids, including shRNA-SNHG12-1, shRNA-SNHG12-2, shRNA-NC,
OV-SNHG12, OV-NC, miR-4429 mimics and mimic-NC, using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) at 37°C, the transfection efficiency was
evaluated at 48 h after transfection using reverse
transcription-quantitative PCR (RT-qPCR).
RT-qPCR
Total RNA was extracted from EC cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). Subsequently, cDNA was synthesized using a cDNA Synthesis
kit (Takara Biotechnology Co., Ltd.). The reverse transcription
conditions were as follows: 25°C for 10 min, 48°C for 30 min and a
final step at 95°C for 5 min. RT-qPCR was conducted using
SYBR-Green Master Mix (Takara Biotechnology Co., Ltd.) on an ABI
7500 fast PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.) with a thermocycling profile of initial denaturation for 10
min at 95°C, followed by 40 cycles at 95°C for 20 sec, 60°C for 30
sec and 72°C for 20 sec. The expression levels of SNHG12 and
miR-4429 were normalized to the levels of GAPDH or U6,
respectively. All reactions were repeated at least three times. The
sequences of PCR primers are presented in Table I. Data were analyzed using the
2−ΔΔCq method (17) to
calculate relative levels.
 | Table I.Primer sequences for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences for reverse
transcription-quantitative PCR.
| Gene | Forward primer
(5′→3′) | Reverse primer
(5′→3′) |
|---|
| SNHG12 |
TCTGGTGATCGAGGACTTCC |
ACCTCCTCAGTATCACACACT |
| GAPDH |
CTCTGCTCCTCCTGTTCGAC |
CGACCAAATCCGTTGACTCC |
| miR-4429 |
GGCCAGGCAGTCTGAGTTG |
GGGAGAAAAGCTGGGCTGAG |
| U6 |
GCTTCGGCAGCACATATACTAAAAT |
CGCTTCACGAATTTGCGTGTCAT |
Cell Counting Kit-8 (CCK-8) assay
RL95-2 cells (2×103 cells/well) were
seeded into a 96-well plate and incubated at 37°C in a humidified
atmosphere containing 5% CO2. A total of 24, 48 and 72 h
post-transfection, 10 µl CCK-8 solution (Dojindo Molecular
Technologies, Inc.) was added to each well for 4 h. The absorbance
(450 nm) of reduced water-soluble tetrazolium salt was evaluated
using a microplate reader.
Wound-healing assay
RL95-2 cells (5×104 cells/well) were
seeded in a 6-well plate and cultured for 24 h. When cells reached
~80% confluence, the cell monolayer was scratched with a 100-µl
pipette tip to create an artificial homogenous wound, the cells
were then washed twice with serum-free DMEM. The cells were
subsequently cultured in serum-free DMEM for 72 h. The degree of
scratch healing was observed with images captured in each group at
0 and 72 h using a light microscope (Olympus Corporation). The cell
migration rate (%) was calculated using the following equation:
[Initial distance (0 h)-final distance (72 h)]/Initial distance (0
h) ×100.
Transwell invasion assay
A Transwell Matrigel invasion assay was performed
using a 24-well Transwell plate. Briefly, a 200-µl cell suspension
(5×104 cells/well) in serum-free DMEM was added into the
upper chamber, which was coated with Matrigel, and 600 µl DMEM
supplemented with 10% FBS was added to the lower chamber. After 48
h, a cotton swab was used to wipe the cells above the Matrigel
layer, whereas the cells below the membrane were fixed with 4%
methanol for 10 min at room temperature and stained with 0.1%
crystal violet solution for 15 min. The numbers of invasive cells
in five random fields (magnification, ×200) were counted using a
light microscope (Olympus Corporation).
Cell cycle assay
After transfection, RL95-2 cells were collected and
fixed in precooled 70% ethanol in PBS for 12 h at 4°C. After
washing with precooled PBS, the cells were resuspended in 1 mg/ml
propidium iodide (PI; Sigma-Aldrich; Merck KGaA) and 0.5 mg/ml
RNase A (Thermo Fisher Scientific, Inc.) in PBS for 25 min at 37°C
in the dark. Finally, cell cycle analysis was conducted using the
FACSCalibur flow cytometer and ModFit LT™ software (version 3.1;
Verity Software House, Inc.).
Dual luciferase reporter assay
Using starBase version 3.0 (http://starbase.sysu.edu.cn/index.php), SNHG12 was
discovered to be a target of miR-4229. For the dual-luciferase
reporter assay, SNHG12 3′-untranslated region (UTR) sequences were
amplified and cloned downstream of the luciferase reporter gene in
pMIR-REPORT luciferase vectors (Thermo Fisher Scientific, Inc.).
Cells (5×104 cells/well) were plated in 24-well plates
and cultured until the cells were ~50% confluent. After 48 h of
co-transfection with 50 nM miR-4429 mimics or mimic-NC and 30 ng/ml
wild-type (WT) or mutant (MUT) firefly luciferase reporter plasmids
at room temperature using Lipofectamine® 2000, cells
were collected. The mutated binding site of the SNHG12 3′-UTR was
created using a QuikChange Multi Site-Directed Mutagenesis kit
(Stratagene; Agilent Technologies, Inc.). After 48 h of culture,
the luciferase activities were measured with a dual-luciferase
reporter system (Promega Corporation), and Renilla
luciferase activity was used for normalization. All experiments
were performed in triplicate.
Western blotting
After transfection, RL95-2 cells were lysed using a
protein lysis buffer (RIPA; Beyotime Institute of Biotechnology)
and the protein concentrations were measured using a micro
bicinchoninic acid protein assay kit (Beyotime Institute of
Biotechnology). The proteins (40 µg) were separated by SDS-PAGE on
10% gels and transferred onto polyvinylidene fluoride membranes,
then blocked with 5% non-fat milk in 20 mM TBS-0.1% Tween for 2 h
at room temperature. The membranes were then incubated with primary
antibodies, including anti-matrix metalloproteinase (MMP)2
(1:1,000; cat. no. ab92536; Abcam), anti-MMP9 (1:1,000; cat. no.
ab137867; Abcam) and anti-GAPDH (1:10,000; cat. no. ab181603;
Abcam) at 4°C overnight, followed by incubation with goat
anti-rabbit horseradish peroxidase-conjugated immunoglobulin G
secondary antibodies (1:5,000; cat. no. ab6721; Abcam) for 2 h at
room temperature. Subsequently, membranes were visualized using an
ECL kit (Invitrogen; Thermo Fisher Scientific, Inc.), and imaged
with a gel imaging system (Bio-Rad Laboratories, Inc.). GAPDH
protein levels were used for normalization. ImageJ software
(version 1.47; National Institutes of Health) was used to analyze
and semi-quantify the relative protein levels.
Statistical analysis
All experiments were performed at least three times.
All data are presented as the mean ± SD. An unpaired Student's
t-test and one-way analysis of variance (ANOVA) followed by a post
hoc Tukey's test were used to analyze the statistical differences,
as appropriate. The analyses were performed using SPSS 19.0
software. P<0.05 was considered to indicate a statistically
significant difference.
Results
lncRNA SNHG12 is upregulated in EC
cell lines, and SNHG12 silencing prevents proliferation, migration
and invasion, and induces cell cycle arrest of RL95-2 cells
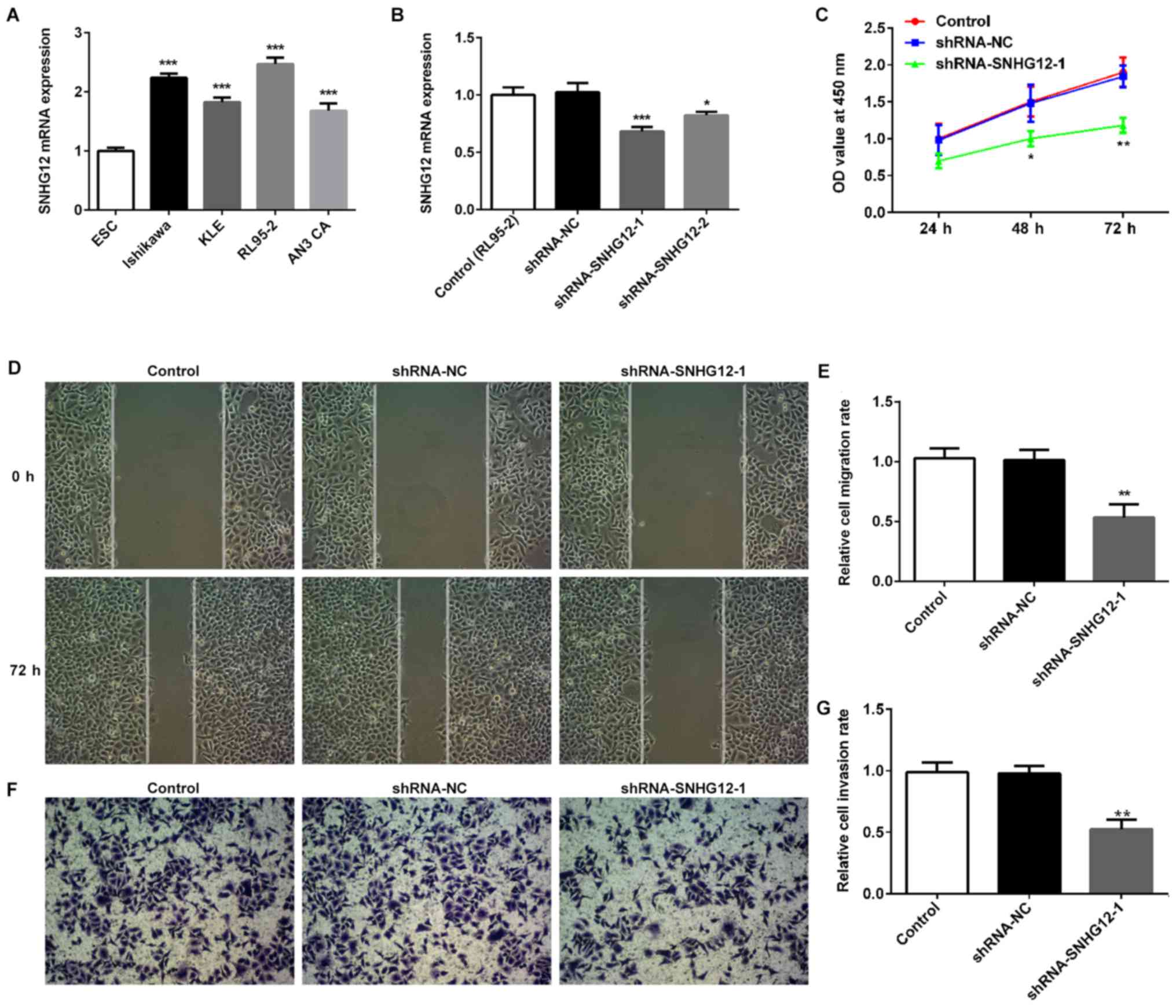
The expression of SNHG12 in EC cell lines was
examined to determine whether SNHG12 was involved in EC. Results
from RT-qPCR demonstrated that SNHG12 was higher in four EC cell
lines, including Ishikawa, KLE, RL95-2 and AN3 CA cells, compared
with the normal EC cell line, ESC (Fig. 1A). Subsequently, SNHG12 was
silenced in RL95-2 cells, as that cell line possessed the highest
SNHG12 mRNA levels, using shRNA-SNHG12-1 and shRNA-SNHG12-2
(Fig. 1B). As shRNA-SNHG12-1 had a
greater inhibitory effect, shRNA-SNHG12-1 was selected for
subsequent studies. Silencing SNHG12 significantly suppressed the
proliferation of RL95-2 cells (Fig.
1C). Wound-healing assays demonstrated that SNHG12 depletion
effectively prevented RL95-2 cells from entering the wounded area
after 72 h, compared with the shRNA-NC and untransfected control
groups, thus suggesting that shRNA-SNHG12-1 was able to inhibit
cell migration (Fig. 1D and E). A
Transwell Matrigel assay was also used to evaluate the invasion of
RL95-2 cells transfected with shRNA-SNHG12-1. The present study
demonstrated that silencing SNHG12 could significantly suppress
invasion of RL95-2 cells (Fig. 1F and
G). In addition, the effects of shRNA-SNHG12-1 treatment on the
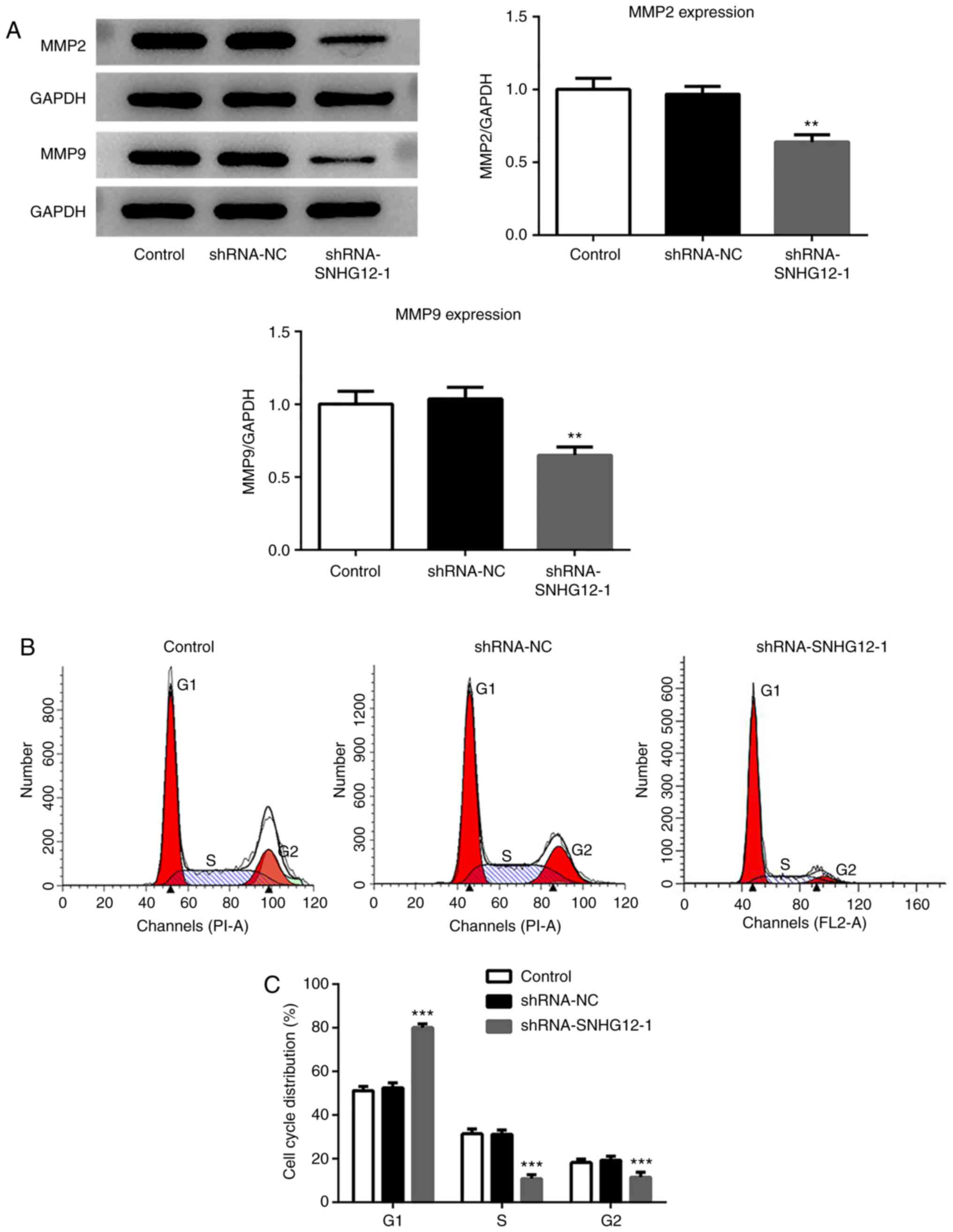
expression of proteins related to RL95-2 cell migration were
assessed. The western blotting results demonstrated that the
expression of MMP2 and MMP9 were significantly decreased in the
shRNA-SNHG12-1 group when compared with the untransfected control
and shRNA-NC groups (Fig. 2A).
Flow cytometry was performed to determine the
effects of SNHG12 on the progression of the RL95-2 cell cycle. As
presented in Fig. 2B and C,
shRNA-SNHG12-1 treatment led to a significant increase in the
proportion of cells at the G1 phase, and significantly
reduced arrest in the S and G2 phases compared with the
untransfected control and shRNA-NC cells.
miR-4429 targets SNHG12 in RL95-2
cells
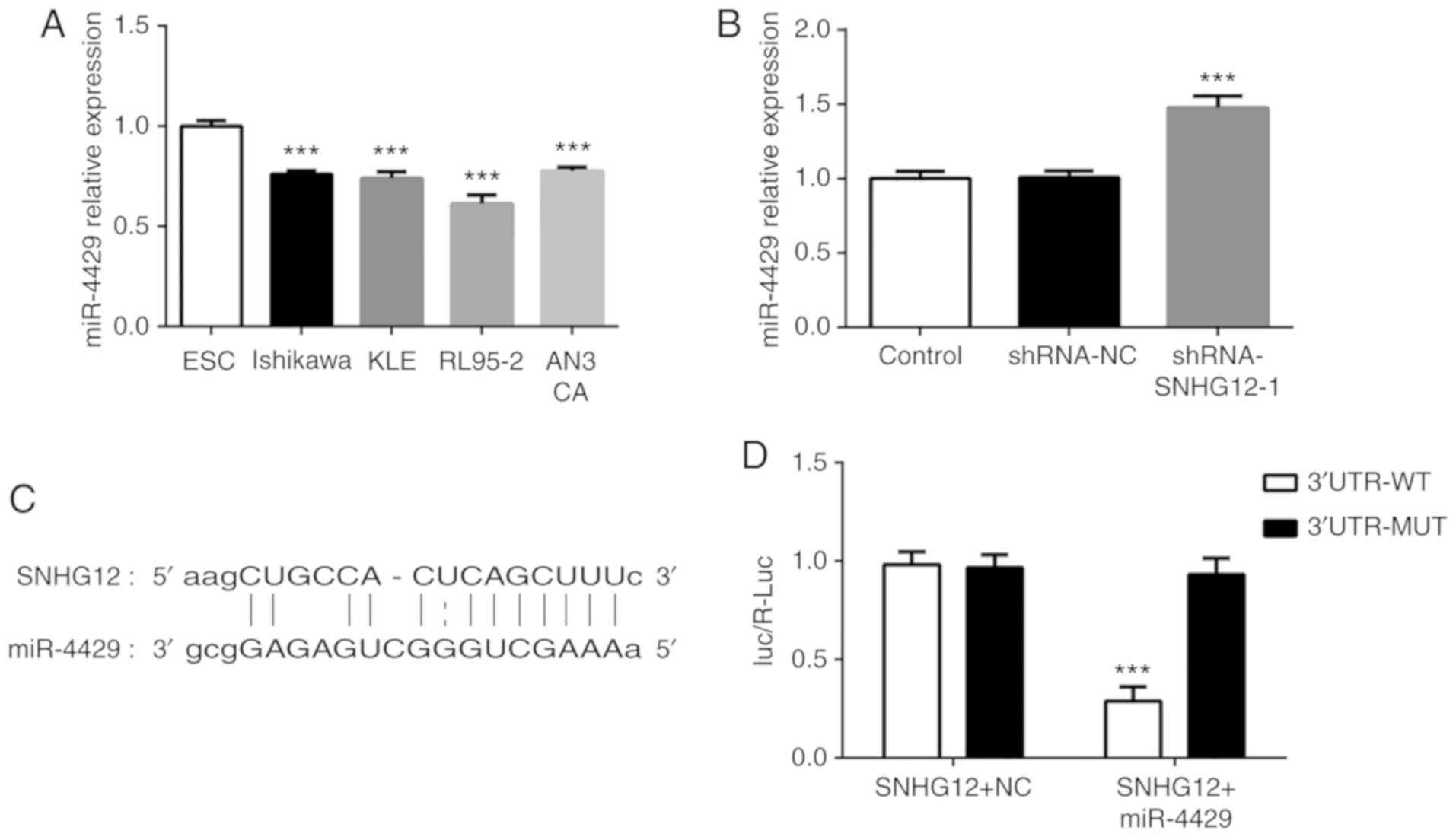
It was determined that miR-4429 was expressed at low
levels in four EC cell lines compared with ESC cells (Fig. 3A). Notably, silencing SNHG12
markedly increased the level of miR-4429 in RL95-2 cells (Fig. 3B). The binding sequences of SNHG12
for miR-4429 are presented in Fig.
3C. It was observed in the luciferase reporter assay that
miR-4429 significantly decreased the luciferase activity of SNHG12
WT but had no influence on that of SNHG12 MUT (Fig. 3D). These findings indicated that
SNHG12 may negatively affect miR-4429 expression in EC cells.
miR-4429 suppresses cell proliferation
and induces cell cycle arrest at the S phase via SNHG12 in RL95-2
cells
miR-4429 was overexpressed in RL95-2 cells by
transfection with miR-4429 mimic (Fig.
4A) and overexpression of SNHG12 was also induced in RL95-2
cells using OV-SNHG12 plasmids (Fig.
4B). The rescue assays were performed to evaluate whether
miR-4429 regulates EC procession by targeting SNHG12. As presented
in Fig. 4C-F, the migration and
invasion of RL95-2 cells were significantly decreased upon
transection with the miR-4429 mimic; however, they were rescued by
overexpression of SNHG12. Additionally, the miR-4429 mimic
suppressed the proliferation of RL95-2 cells, which was then
partially reversed by OV-SNHG12 (Fig.
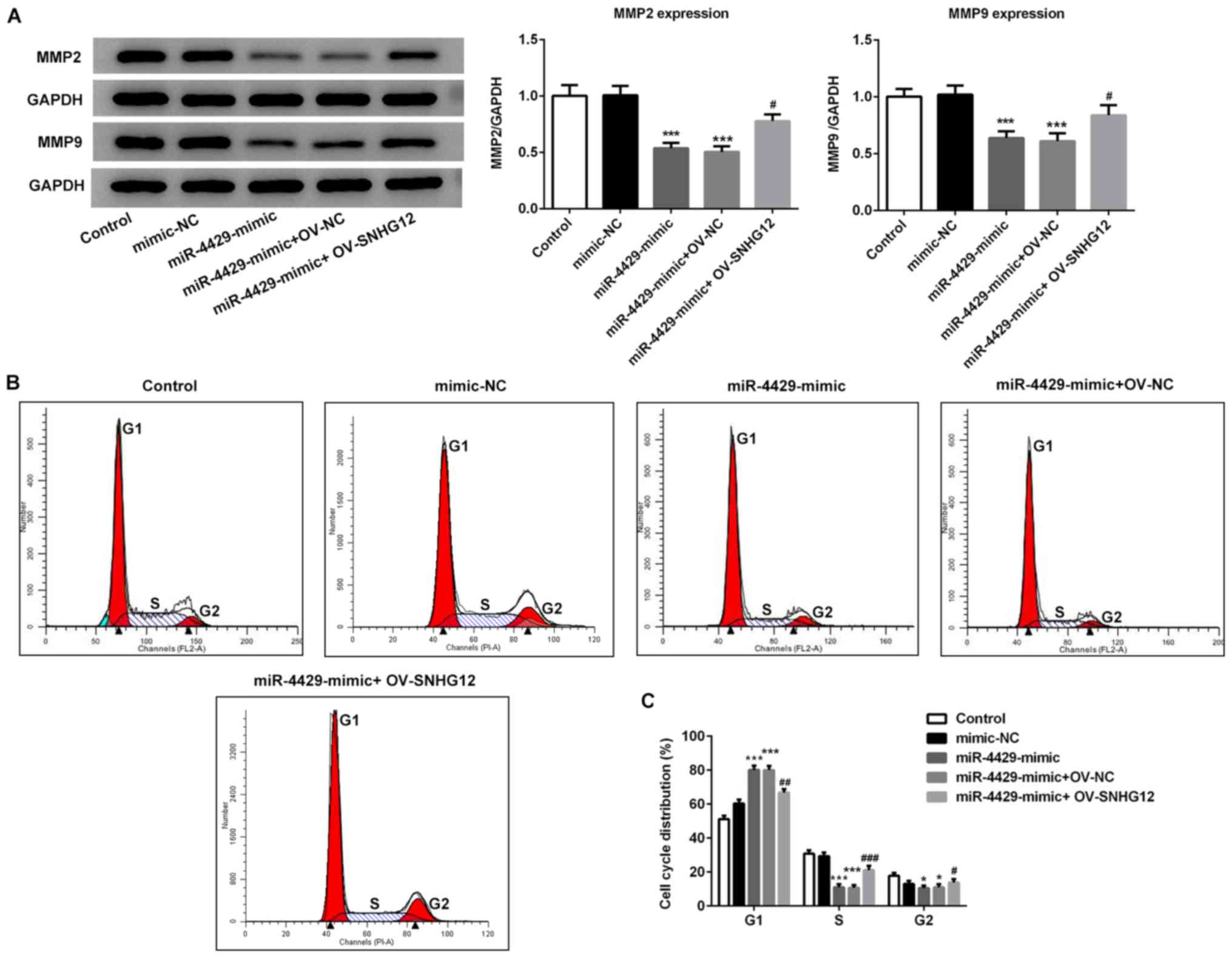
4G). Furthermore, the expression levels of proteins related to
migration in response to miR-4429 overexpression were detected by
western blotting. As presented in Fig.
5A, the results demonstrated that transfection with miR-4429
mimic significantly decreased the expression of MMP2 and MMP9
compared with the control and mimic-NC groups. However, RL95-2
cells in the miR-4429 mimic + OV-SNHG12 group presented higher
expression levels of MMP2 and MMP9 when compared with the cells in
the miR-4429 mimic + OV-NC group. The flow cytometry data
demonstrated that the miR-4429 mimic increased the number of cells
at G1 phase, whereas the percentage of cells in the S
phase were significantly decreased compared with the mimic-NC group
(Fig. 5B and C). By contrast,
OV-SNHG12 effectively reversed cell cycle arrest at G1
and increased the number of cells at S phase compared with the
miR-4429 mimic + OV-NC group (Fig. 5B
and C). Taken together, these findings suggested that miR-4429
suppressed EC progression and induced cell cycle arrest through the
targeting of SNHG12.
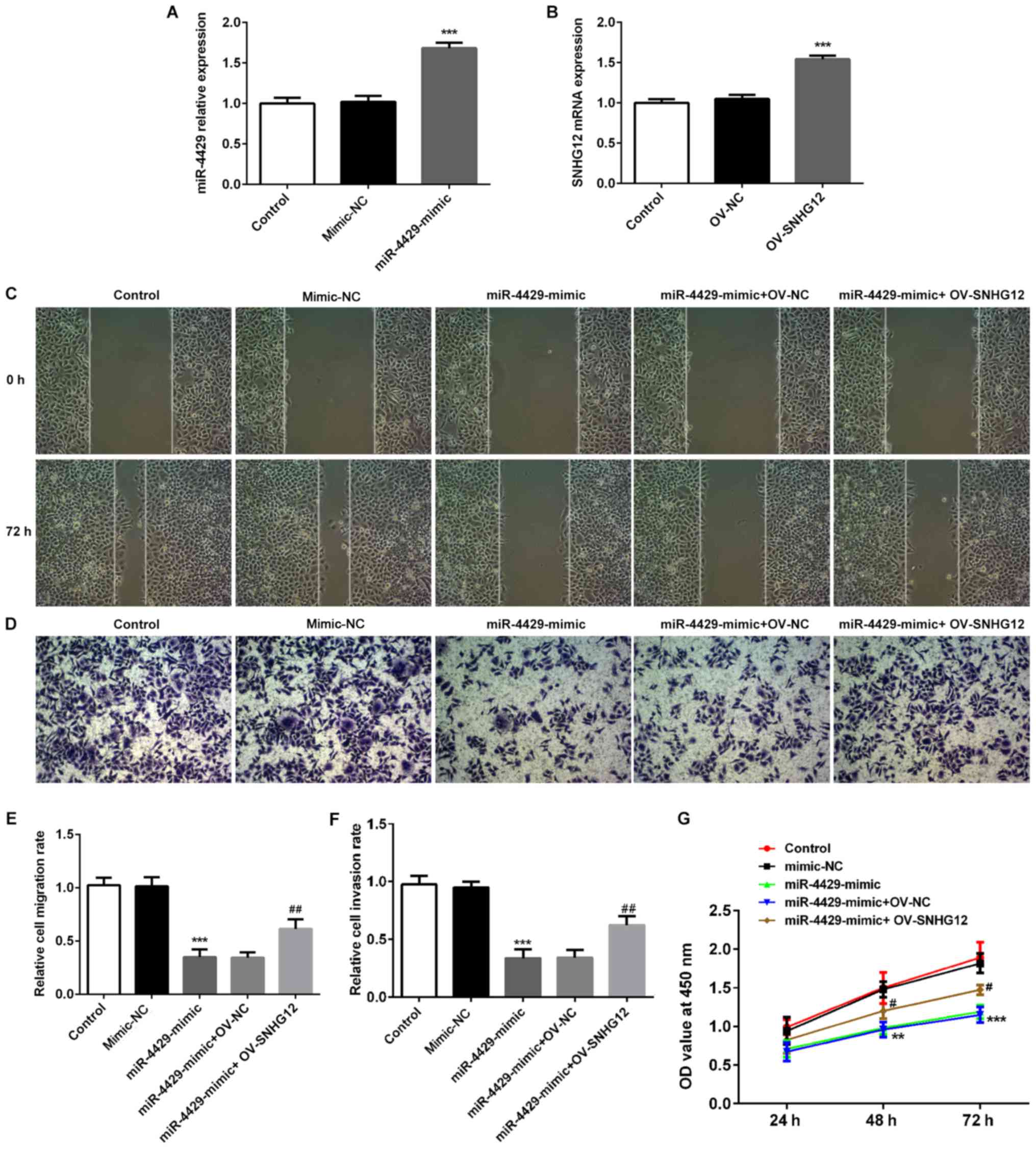 | Figure 4.Overexpression of miR-4429 inhibits
endometrial cancer migration, invasion and proliferation through
targeting SNHG12 in RL95-2 cells. (A) Expression levels of miR-4429
in RL95-2 cells transfected with a miR-4429 mimic were assessed
using RT-qPCR. ***P<0.001 vs. control and mimic-NC groups. (B)
SNHG12 expression in RL95-2 cells transfected with OV-SNHG12 was
detected using RT-qPCR. ***P<0.001 vs. control and OV-NC groups.
Data are presented as the mean ± SD (n=3). (C-F) RL95-2 cells were
transfected with mimic-NC, miR-4429 mimic, miR-4429 mimic + OV-NC
and miR-4429 mimic + OV-SNHG12, respectively. The capacities of (C
and E) migration and (D and F) invasion of RL95-2 cells were
evaluated using wound healing and Transwell assays. Magnification,
×200. (G) Proliferation of RL95-2 cells was detected by Cell
Counting Kit-8 assay. Data are represented as the mean ± SD (n=3).
**P<0.01 and ***P<0.001 vs. control and mimic-NC groups;
#P<0.05 and ##P<0.01 vs. miR-4429 mimic
+ OV-NC group. SNHG12, small nucleolar RNA host gene 12; miR,
microRNA; RT-qPCR, reverse transcription quantitative PCR; NC,
negative control; OV, overexpression plasmid; OD, optical
density. |
Discussion
EC is the most common gynecologic malignancy. The
main treatment for EC is total hysterectomy with bilateral
salpingo-oophorectomy (18).
However, the prognosis and survival rate are not satisfactory.
Despite the roles that radiation and chemotherapy serve in EC
treatment, and the promising therapies in drug development for the
treatment of advanced/recurrent EC, no targeted therapies beyond
hormonal therapy have been approved at present (19). Furthermore, early diagnosis is key
to improving the probability of EC survival, which has a <20%
survival rate for the advanced disease at 5 years but >90%
survival rate for the early-stage disease (20). However, to the best of our
knowledge, there are no validated biological markers for its early
detection. Thus, there is an urgent need for scientifically
validated therapy with predictive biomarkers. In the present study,
it was demonstrated that SNHG12 was significantly upregulated in EC
cells, which was negatively associated with miR-4429. Therefore,
SNHG12 and miR-4429 may represent biomarkers that can be used for
early diagnosis and targeted therapy of EC.
lncRNAs, miRNAs, piwiRNAs and other non-coding RNAs
comprise ~90% of human genomes and can act as oncogenes or tumor
suppressor genes (21,22). SNHG12, also known as LNC04080, is
1,867 bases in length and is involved in a number of human tumors,
including non-small cell lung cancer, gastric cancer,
triple-negative breast cancer, hepatocellular carcinoma, colorectal
cancer, cervical cancer and ovarian cancer (23). Changes in the expression levels of
SNHG12 have been reported to be associated with viability,
proliferation, migration and invasion of tumor cells, thus
affecting the prognosis and survival of patients with cancer
(24). In the present study,
SNHG12 silencing inhibited the proliferation, migration and
invasion of EC cells, and induced cell cycle arrest in the
G1 phase, which may enhance apoptosis. In addition,
SNHG12 silencing markedly inhibited the expression of MMP2 and
MMP9, which are key molecules in cancer invasion and metastasis
(25). The present data identified
a potential mechanism on how SNHG12 depletion may inhibit cell
proliferation, invasion and migration.
SNHG12 also acts as a competitive endogenous RNA by
harboring multiple miRNA-binding sites, thereby regulating their
downstream targets by ‘sponging’ these miRNAs (23). Given that the interaction between
lncRNAs and their mRNA targets has been predicted to rely on
miRNAs, an increased density of miRNA response elements may
increase the likelihood of lncRNAs to share miRNAs to communicate
with and coregulate each other (26). The miRNAs that have been reported
to be regulated by SNHG12 include miR-195 (27) and miR-101-3p (28), which have roles in the regulation
of cell proliferation, and miR-424-5p and miR-125b, which regulate
invasion and metastasis (14).
Through the TargetScan Human prediction site, it was identified
that miR-4429 potentially interacted with SNHG12. Previous studies
have demonstrated that miR-4429 can be sponged by lncRNA LINC00313
to modulate papillary thyroid cancer tumorigenesis (16). miR-4429 was also demonstrated to
sensitize cervical cancer cells to irradiation by targeting RAD51
(29) and inhibit the progression
of clear cell renal cell carcinoma by targeting CDK6 (30). These findings demonstrated that
miR-4429 may function as a tumor suppressor gene in several types
of cancer. The present study demonstrated that miR-4429 mimic
inhibited EC cell proliferation, migration and invasion, and at
least partially inhibited cell cycle arrest. The present study also
demonstrated the inhibitory effect of miR-4429 mimic on the
proliferation of RL95-2 cells by enhancing the number of cells in
G1 phase. Notably, the number of cells in G1
phase was increased after mimic-NC transfection compared with in
the untransfected control group; it was suspected that the reason
may be the mimic-NC plasmid affected the G1 phase of the
RL95-2 cell cycle, whereas it had no significant effect on the S
phase and G2 phase. However, in general, the miR-4429
mimic plasmid had a significant effect on all phases the cell
cycle. All these suppressive effects of miR-4429 mimic on EC cells
were attenuated by SNHG12 overexpression.
In summary, the present data demonstrated that
silencing of SNHG12 was able to significantly suppress cell
viability and cause cell cycle arrest, which was similar to the
effects of the miR-4429 mimic. To the best of our knowledge, this
is the first study to identify SNHG12 as a target molecule of
miR-4429, which may serve as potential diagnostic and prognostic
biomarkers, or targets for novel therapeutic strategies for EC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PYC and MXW designed the study, wrote the manuscript
and performed the cell culture and molecular biology experiments;
BZ, SYW and HYW performed the cell culture and molecular biology
experiments and analyzed the data; and LW designed the study and
critically revised the manuscript for intellectual content. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Brooks RA, Fleming GF, Lastra RR, Lee NK,
Moroney JW, Son CH, Tatebe K and Veneris JL: Current
recommendations and recent progress in endometrial cancer. CA
Cancer J Clin. 69:2842–279. 2019.
|
|
2
|
Morice P, Leary A, Creutzberg C,
Abu-Rustum N and Darai E: Endometrial cancer. Lancet.
387:1094–1108. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Van Nyen T, Moiola CP, Colas E, Annibali D
and Amant F: Modeling endometrial cancer: Past, present, and
future. Int J Mol Sci. 19:23482018. View Article : Google Scholar
|
|
4
|
Kahlert C and Kalluri R: Exosomes in tumor
microenvironment influence cancer progression and metastasis. J Mol
Med (Berl). 91:431–437. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhou M, Zhong L, Xu W, Sun Y, Zhang Z,
Zhao H, Yang L and Sun J: Discovery of potential prognostic long
non-coding RNA biomarkers for predicting the risk of tumor
recurrence of breast cancer patients. Sci Rep. 6:310382016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou M, Xu W, Yue X, Zhao H, Wang Z, Shi
H, Cheng L and Sun J: Relapse-related long non-coding RNA signature
to improve prognosis prediction of lung adenocarcinoma. Oncotarget.
7:29720–29738. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou M, Sun Y, Sun Y, Xu W, Zhang Z, Zhao
H, Zhong Z and Sun J: Comprehensive analysis of lncRNA expression
profiles reveals a novel lncRNA signature to discriminate
nonequivalent outcomes in patients with ovarian cancer. Oncotarget.
7:32433–32448. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li J, Xue W, Lv J, Han P, Liu Y and Cui B:
Identification of potential long non-coding RNA biomarkers
associated with the progression of colon cancer. Oncotarget.
8:75834–75843. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang XH, Hu P, Xie YQ, Kang YJ and Li M:
lncRNA HOTAIR promotes endometrial carcinoma cells proliferation by
binding to PTEN via activating PI3k/Akt signaling pathway. Mol Cell
Biol. 39:e00251–e00219. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu ZB, Tang C, Jin X, Liu SH and Pi W:
Increased expression of lncRNA SNHG12 predicts a poor prognosis of
nasopharyngeal carcinoma and regulates cell proliferation and
metastasis by modulating notch signal pathway. Cancer Biomark.
23:603–613. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Feng X, Dong X, Wu D, Zhao H, Xu C and Li
H: Long noncoding RNA small nucleolar RNA host gene 12 promotes
papillary thyroid carcinoma cell growth and invasion by targeting
miR-16-5p. Histol Histopathol. 35:217–224. 2020.PubMed/NCBI
|
|
12
|
Liu Y, Zhou J, Wang S, Song Y, Zhou J and
Ren F: Long non-coding RNA SNHG12 promotes proliferation and
invasion of colorectal cancer cells by acting as a molecular sponge
of microRNA-16. Exp Ther Med. 18:1212–1220. 2019.PubMed/NCBI
|
|
13
|
Sun D and Fan XH: LncRNA SNHG12
accelerates the progression of ovarian cancer via absorbing
miRNA-129 to upregulate SOX4. Eur Rev Med Pharmacol Sci.
23:2345–2352. 2019.PubMed/NCBI
|
|
14
|
Jin XJ, Chen XJ, Zhang ZF, Hu WS, Ou RY,
Li S, Xue JS, Chen LL, Hu Y and Zhu H: Long noncoding RNA SNHG12
promotes the progression of cervical cancer via modulating
miR-125b/STAT3 axis. J Cell Physiol. 234:6624–6632. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu X, Chen R and Liu L:
SP1-DLEU1-miR-4429 feedback loop promotes cell proliferative and
anti-apoptotic abilities in human glioblastoma. Biosci Rep.
39:BSR201909942019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu WJ, Yin H, Hu JJ and Wei XZ: Long
noncoding RNA LINC00313 modulates papillary thyroid cancer
tumorigenesis via sponging miR-4429. Neoplasma. 65:933–942. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Braun MM, Overbeek-Wager EA and Grumbo RJ:
Diagnosis and management of endometrial cancer. Am Fam Physician.
93:468–474. 2016.PubMed/NCBI
|
|
19
|
Lee YC, Lheureux S and Oza AM: Treatment
strategies for endometrial cancer: Current practice and
perspective. Curr Opin Obstet Gynecol. 29:47–58. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Njoku K, Chiasserini D, Whetton AD and
Crosbie EJ: Proteomic biomarkers for the detection of endometrial
cancer. Cancers (Basel). 11:15722019. View Article : Google Scholar
|
|
21
|
Khandelwal A, Malhotra A, Jain M, Vasquez
KM and Jain A: The emerging role of long non-coding RNA in
gallbladder cancer pathogenesis. Biochimie. 132:152–160. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Malhotra A, Sharma U, Puhan S, Chandra
Bandari N, Kharb A, Arifa PP, Thakur L, Prakash H, Vasquez KM and
Jain A: Stabilization of miRNAs in esophageal cancer contributes to
radioresistance and limits efficacy of therapy. Biochimie.
156:148–157. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tamang S, Acharya V, Roy D, Sharma R,
Aryaa A, Sharma U, Khandelwal A, Prakash H, Vasquez KM and Jain A:
SNHG12: An LncRNA as a potential therapeutic target and biomarker
for human cancer. Front Oncol. 9:9012019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lan T, Ma W, Hong Z, Wu L, Chen X and Yuan
Y: Long non-coding RNA small nucleolar RNA host gene 12 (SNHG12)
promotes tumorigenesis and metastasis by targeting miR-199a/b-5p in
hepatocellular carcinoma. J Exp Clin Cancer Res. 36:112017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen P, Hu MD, Deng XF and Li B: Genistein
reinforces the inhibitory effect of cisplatin on liver cancer
recurrence and metastasis after curative hepatectomy. Asian Pac J
Cancer Prev. 14:759–764. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tan JY and Marques AC: miRNA-mediated
crosstalk between transcripts: The missing ‘linc’? Bioessays.
38:295–301. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu X, Zheng J, Xue Y, Qu C, Chen J, Wang
Z, Li Z, Zhang L and Liu Y: Inhibition of TDP43-mediated
SNHG12-miR-195-SOX5 feedback loop impeded malignant biological
behaviors of glioma cells. Mol Ther Nucleic Acids. 10:142–158.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sun Y, Liu J, Chu L, Yang W, Liu H, Li C
and Yang J: Long noncoding RNA SNHG12 facilitates the tumorigenesis
of glioma through miR-101-3p/FOXP1 axis. Gene. 676:315–321. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sun H, Fan G, Deng C and Wu L: miR-4429
sensitized cervical cancer cells to irradiation by targeting RAD51.
J Cell Physiol. 235:185–193. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pan H, Hong Y, Yu B, Li L and Zhang X:
miR-4429 inhibits tumor progression and epithelial-mesenchymal
transition via targeting CDK6 in clear cell renal cell carcinoma.
Cancer Biother Radiopharm. 34:334–341. 2019. View Article : Google Scholar : PubMed/NCBI
|