Introduction
Acute myeloid leukemia (AML) is a frequently
diagnosed malignant clonal disease of hematopoietic stem cells that
shows high heterogeneity (1). The
disease primarily manifests via differentiation and maturation
disorders of myeloid progenitor cells, abnormal proliferation,
inhibited malignant clonal leukemia cell apoptosis and abnormal
hematopoiesis (2). AML is the most
common type of acute leukemia in adults, accounting for
approximately 80% of cases, and the incidence of AML is increasing
annually, thus seriously affecting human health (3,4).
According to statistics obtained between 2006 and 2010, the 5-year
overall survival rate of patients with AML was approximately 21.4%
and the 5-year overall survival rate for elderly patients was
<10%, due to poor tolerability to chemotherapy (5,6).
Therefore, it is important to identify novel treatment strategies
to improve complete remission rates and prolong disease-free
survival time for patients with AML. In the past few years, AML
treatment strategies have progressed slowly, which may be
associated with the increased resistance of AML cells to
chemotherapeutic drugs (7).
However, reducing the resistance of AML cells to chemotherapy is
difficult and has become a key focus of the treatment of the
disease.
MicroRNAs (miRNAs) are a class of highly conserved
non-coding endogenous small molecules, which can bind to the
3′-untranslated regions (UTRs) of mRNAs to interfere with
translation. As a result, miRNAs participate in a series of
biological processes including cell proliferation, differentiation
and apoptosis (8,9). Previous studies have reported that
miRNAs are differentially expressed in tumor cells, and miRNA
expression is associated with the regulation of cancer cell
biological processes and the modulation of oncogene/anti-oncogene
expression during tumor formation and development (10,11).
Similarly, miRNAs also play crucial roles during the pathogenesis,
diagnosis and treatment of leukemia (12). A previous study reported that
multiple factors are involved in the regulation of multidrug
resistance in leukemia (13).
Szymczyk et al (14)
revealed that low miR-34a expression was closely related to the
sensitivity of patients with chronic lymphocytic leukemia to purine
nucleoside analogs, including cladribine and fludarabine. Another
previous study reported that miR-130a expression was upregulated in
patients with t(8;21) AML and AML cell lines, but the expression of
miR-130a decreased significantly once patients with t(8;21) AML
achieved complete remission (15).
Therefore, the aforementioned studies suggested that low miR-34a
expression contributes to the decrease of clonal leukemic
cells.
miR-130a has been reported to promote cancer growth
(16). Yin et al (17) reported that the upregulation of
miR-130a was highly correlated with advanced clinical stage and
lymph node metastasis in cervical cancer. Therefore, inhibition of
miR-130a expression may represent a treatment strategy with
potentially antileukemic effects. At present, few studies have
investigated the role of miR-130a in AML cell drug resistance. In
the present study, the effects of miR-130a on the sensitivity of
AML cells to Adriamycin (Adr) were investigated. AML cell viability
and invasion were assessed following miR-130a overexpression and
knockdown. The aim of the present study was to identify a potential
therapeutic target for drug-resistant AML, to promote the
sensitivity of AML cells to Adr and improve the prognosis of
patients with AML.
Materials and methods
Cell culture
The AML cell line HL-60 was purchased from the
German Collection of Microorganisms and Cell Cultures GmbH (DSMZ
no. ACC-3). HL-60 cells (1×106 cells/ml) were maintained
in RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.)
containing 10% heat-inactivated fetal bovine serum (FBS;
Invitrogen; Thermo Fisher Scientific, Inc.), 100 µg/ml streptomycin
(Invitrogen; Thermo Fisher Scientific, Inc.) and 100 µg/ml
penicillin (Invitrogen; Thermo Fisher Scientific, Inc.) in a
humidified incubator at 37°C with 5% CO2. Cells in the
logarithmic phase of growth were used for subsequent
experiments.
Cell treatment
HL-60 cells were stimulated using
5-aza-2′-deoxycytidine (5-Aza-dC; 2.5 µmol/l; Sigma-Aldrich; Merck
KGaA). Control cells were incubated with DMSO (Sigma-Aldrich; Merck
KGaA). After incubation for 3 days at 37°C, cells were treated with
Adriamycin (Adr; Sigma-Aldrich; Merck KGaA) at different
concentrations (0.00, 0.01, 0.10, 1.00, 5.00 or 10.00 µmol/l) for
24 h at 37°C.
Cell transfection
miR-130a mimic (50 nM; 5′-CAGUGCAAUGUUAAAAGGGCAU-3′;
cat. no. 4464066; Thermo Fisher Scientific, Inc.) and miR-130a
inhibitor (50 nM; 5′-AUGCCCUUUUAACAUUGCACUG-3′; cat. no. 4464084;
Thermo Fisher Scientific, Inc.) were transfected into HL-60 cells
(2×104 cells/well) alone or in combination with Adr (0.1
µmol/l) using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, inc.), according to the manufacturer's protocol.
Negative control cells were transfected with 50 nM negative
inhibitor (5′-UUUGUACUACACAAAAGUACUG-3′; cat. no. AM17010; Thermo
Fisher Scientific, Inc.) and 50 nM negative mimic
(5′-CAGUACUUUUGUGUAGUACAAA-3′; cat. no. 4464058; Thermo Fisher
Scientific, Inc.), while untransfected cells served as controls. At
24 h post-transfection, the cells were collected for subsequent
experiments.
Cell Counting Kit-8 (CCK-8) assay
HL-60 cell viability following treatment and
transfection was determined using the CCK-8 assay (Beyotime
Institute of Biotechnology), according to the manufacturer's
protocol. Briefly, cells in the logarithmic phase of growth were
seeded (1×104 cells/well) into 96-well plates. After
transfection or treatment for 24 h, 10 µl CCK-8 reagent was added
to each well and incubated for 2 h at 37°C with 5% CO2.
The optical density of each well was assessed at a wavelength of
450 nm using an ELX800 microplate reader (BioTek Instruments,
Inc.).
Reverse transcription-quantitative PCR
(RT-qPCR)
RT-qPCR was performed to determine the expression
level of miR-130a in HL-60 cells after treatment and transfection.
Total RNA was extracted from HL-60 cells using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocol. Total RNA quality and integrity were
evaluated using a NanoDrop-2000c spectrophotometer (Thermo Fisher
Scientific, Inc.) and 1% modified agarose gel electrophoresis.
Subsequently, total RNA (1 µg) was reverse transcribed into cDNA
using the SuperScript III First-Strand Synthesis system
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. qPCR was performed using the SYBR Premix
Ex Taq™ II (Takara Biotechnology) in the ABI Prism 7500
Fast Real-time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. The
following primer pair was synthesized by Shanghai GenePharma Co.,
Ltd. and used for qPCR: miR-130a forward,
5′-GATGCTCTCAGTGCAATGTTA-3′ and reverse,
5′-CTCTGTCTCTCGTCTTGTTGGTAT-3′; and U6 forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′.
The following thermocycling conditions were used for qPCR: Initial
denaturation at 95°C for 5 min, 40 cycles at 95°C for 10 sec and
60°C for 45 sec, and a final extension step at 72°C for 10 min.
miRNA levels were quantified using the 2−ΔΔCq method
(18) and normalized to the
internal reference gene U6.
Transwell invasion assay
HL-60 cell invasion after transfection was analyzed
using Transwell invasion chambers (Costar; Corning, Inc.).
Transfected cells (1×104 cells/well) were suspended in
serum-free RPMI-1640 media, and plated into the upper chambers.
Transwell membranes were pre-coated with Matrigel® (BD
Biosciences) at 37°C for 4 h. RPMI-1640 media containing 10% FBS
was plated in the lower chambers. After incubation for 24 h at
37°C, non-invading cells were removed from the Transwell membrane
using cotton swabs, and invading cells were fixed in 4%
paraformaldehyde for 15 min at room temperature. Subsequently,
invading cells were stained using 0.1% crystal violet solution for
20 min at room temperature and observed using an Eclipse TS-100
inverted microscope (magnification, ×200; Nikon Corporation).
Western blotting (WB)
WB was performed to determine the expression levels
of EMT-related proteins in HL-60 cells after transfection. Total
protein was extracted from transfected cells using RIPA lysis
buffer (Beijing Solarbio Biotech Co., Ltd.) according to the
manufacturer's protocol. Total protein was quantified using the
bicinchoninic protein assay kit (Pierce; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. Protein (50 µg per
lane) was separated via 12% SDS-PAGE and transferred onto PVDF
membranes (EMD Millipore). After blocking with 5% non-fat milk for
1 h at room temperature, the membranes were incubated overnight at
4°C with primary antibodies targeted against: E-Cadherin (1:20,000;
cat. no. ab40772; Abcam), N-Cadherin (1:1,000; cat. no. ab18203;
Abcam), Vimentin (1:2,000; cat. no. ab92547; Abcam) and GAPDH
(1:1,000; cat. no. ab8245; Abcam). Subsequently, the membranes were
incubated at 37°C for 2 h with horseradish peroxidase-conjugated
rabbit anti-mouse IgG H&L (1:7,000; cat. no. ab3728; Abcam) and
horseradish peroxidase-conjugated goat anti-rabbit IgG H&L
(1:7,000; cat. no. ab6721; Abcam) secondary antibodies. Protein
bands were visualized using an ECL detection reagent (EMD
Millipore). GAPDH was used as the loading control. The data were
analyzed via densitometry using ImageJ software (version 1.46;
National Institutes of Health) and normalized to the expression
level of the internal control.
Statistical analysis
SPSS software (version 20; IBM Corp.) was used to
perform statistical analyses. Data are presented as the mean ±
standard deviation. Comparisons among groups were analyzed using
one-way or two-way ANOVA followed by Tukey's post-hoc test. All
experiments were performed in triplicate. P<0.05 was considered
to indicate a statistically significant difference.
Results
5-Aza-dC increases miR-130a expression
levels in Adr-treated AML cells
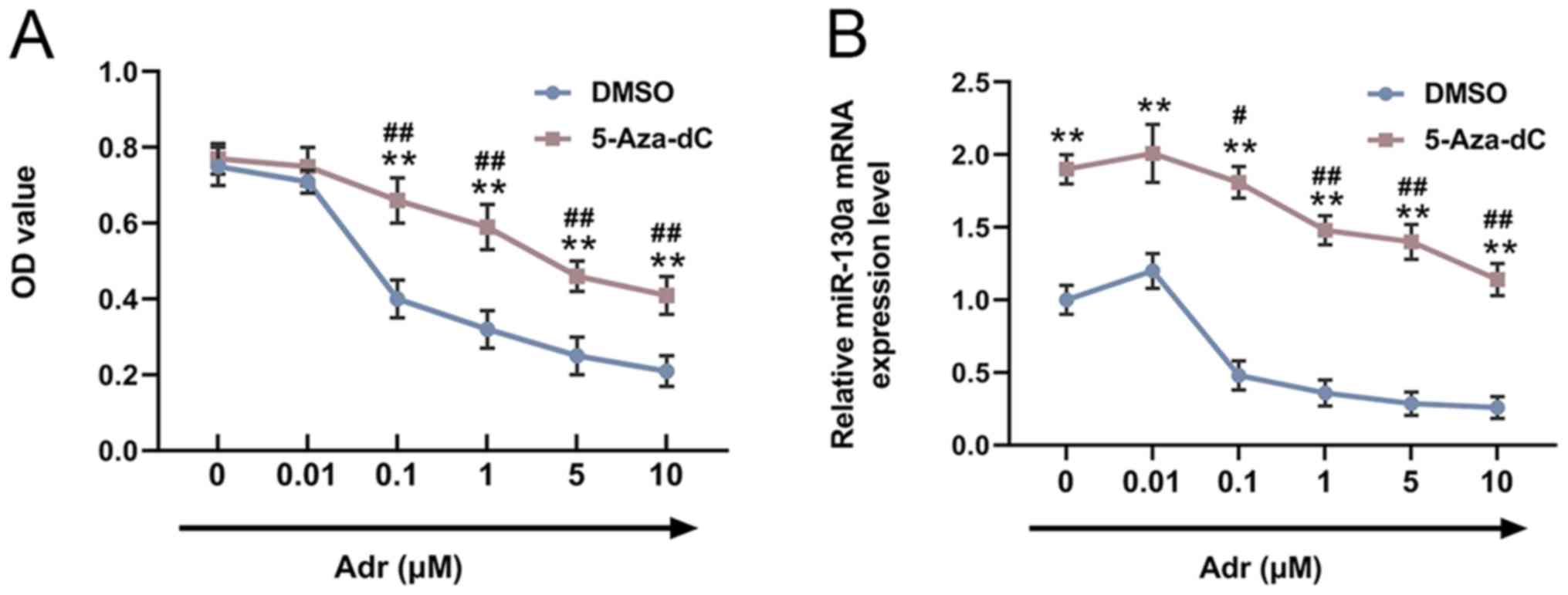
In order to explore the effects of 5-Aza-dC on the
viability of Adr-treated AML cells, HL-60 cells were pre-stimulated
with 5-Aza-dC or DMSO, and subsequently treated with different
concentrations of Adr (0.00, 0.01, 0.10, 1.00, 5.00 or 10.00
µmol/l). Compared with the control cells, pre-treatment with
5-Aza-dC increased the viability of Adr-treated HL-60 cells, and
cell viability decreased in a dose-dependent manner with increasing
Adr concentrations (P<0.001; Fig.
1A). miR-130a expression in HL-60 cells was measured by
RT-qPCR. The results suggested that miR-130a expression in
Adr-treated cells was increased by 5-Aza-dC, and similarly,
miR-130a expression levels decreased in a dose-dependent manner
with increasing Adr concentrations (P<0.05; Fig. 1B).
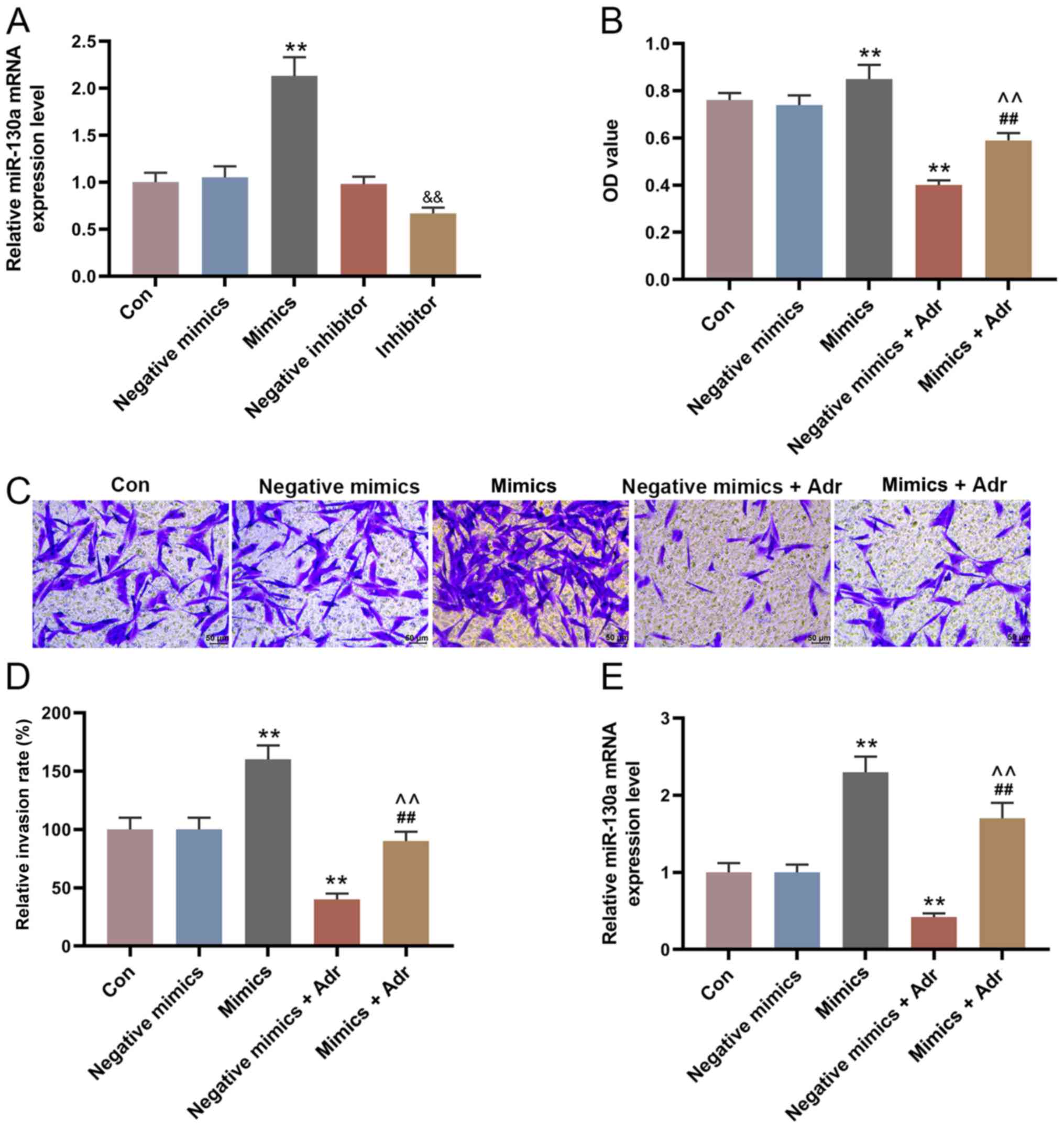
miR-130a overexpression reduces the
sensitivity of AML cells to Adr
The expression of miR-130a in the mimic group was
higher compared with the negative mimic group, and miR-130a
expression was decreased in the miR-130a inhibitor group compared
with the negative inhibitor group (P<0.001; Fig. 2A). To investigate the effect of
miR-130a overexpression on the sensitivity of AML cells to Adr,
miR-130a mimic and negative mimic were transfected into HL-60 cells
alone or in combination with Adr (0.1 µmol/l). The results
suggested that miR-130a overexpression increased HL-60 cell
viability, compared with the control group, and alleviated the
inhibitory effect of Adr on cell viability (P<0.001; Fig. 2B). Moreover, the Transwell assay
results indicated that Adr significantly decreased the number of
invasive HL-60 cells in the negative mimic group, which was
partially reversed by miR-130a overexpression (P<0.001; Fig. 2C and D). The RT-qPCR results
suggested that the expression of miR-130a in HL-60 cells
transfected with negative mimic was significantly inhibited by Adr
(P<0.001; Fig. 2E). Moreover,
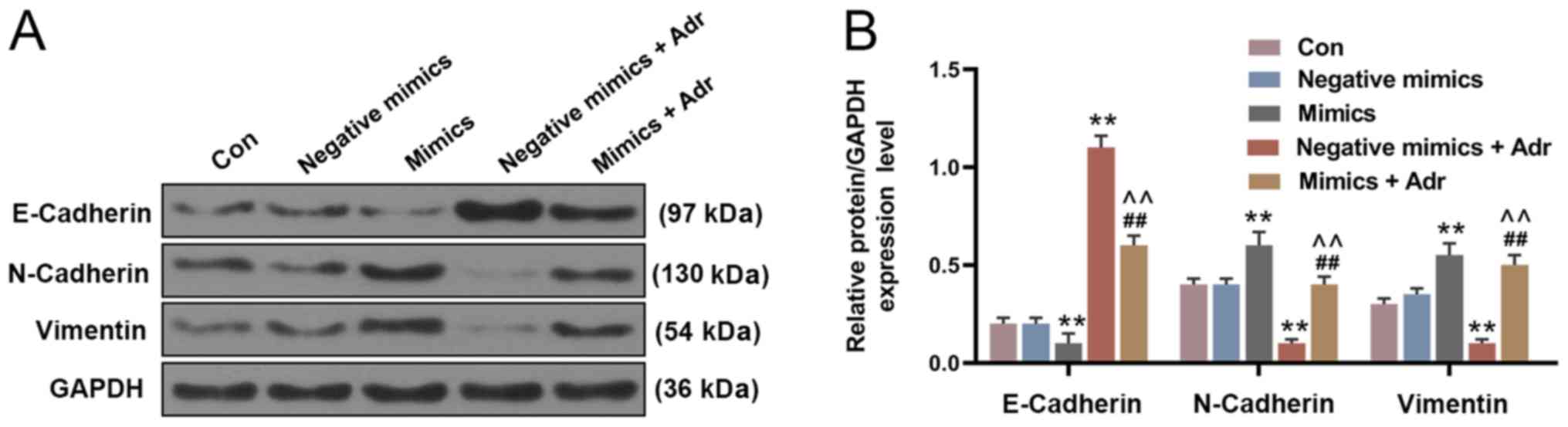
miR-130a overexpression significantly decreased the expression of
E-Cadherin, and increased the expression of N-Cadherin and Vimentin
in HL-60 cells compared with the negative mimic control group.
However, Adr treatment displayed the opposite effect to miR-130a
overexpression on EMT-related protein expression. Adr treatment
following miR-130a overexpression reversed the effects of Adr on
EMT-associated protein expression levels (P<0.001; Fig. 3).
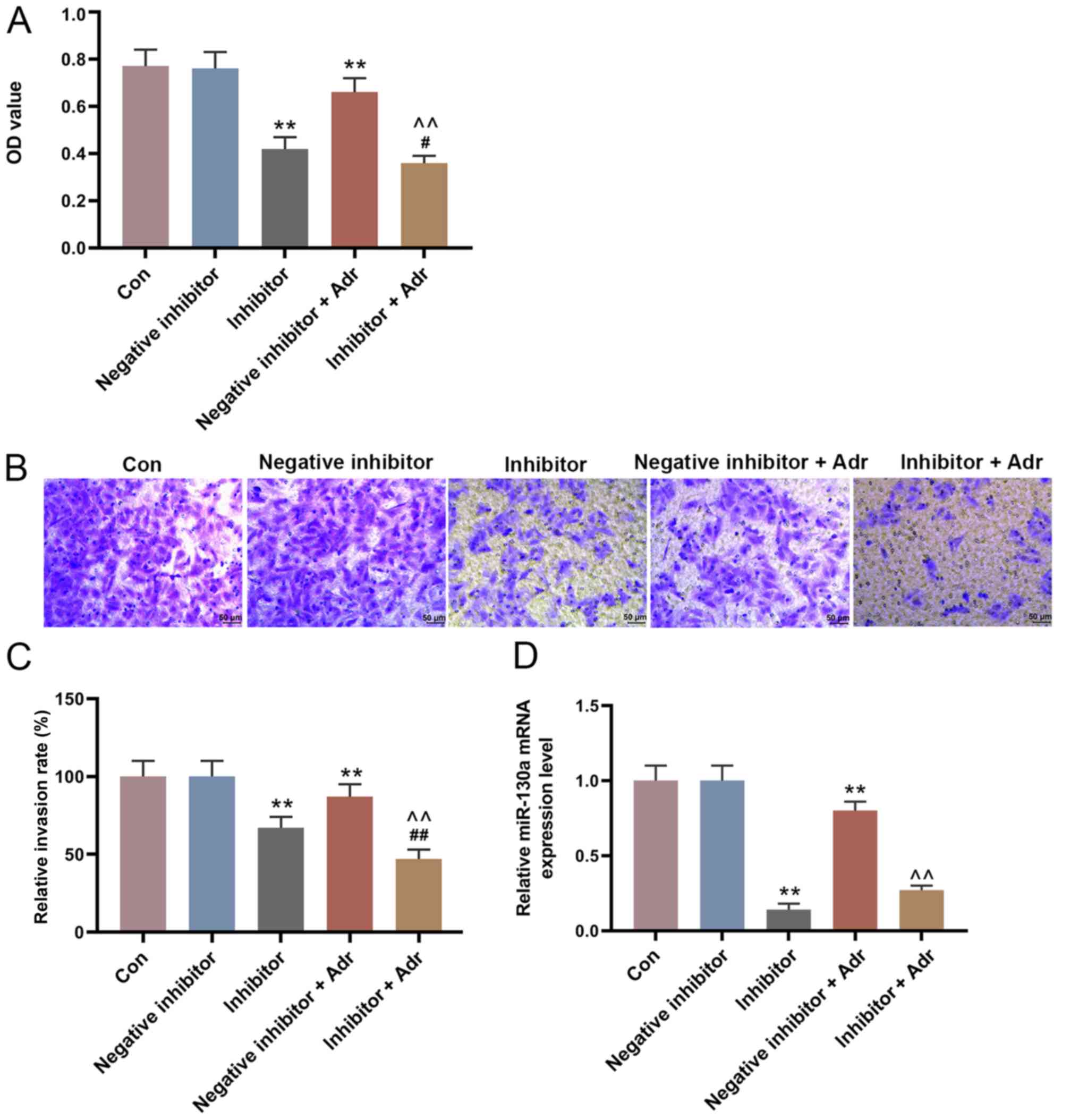
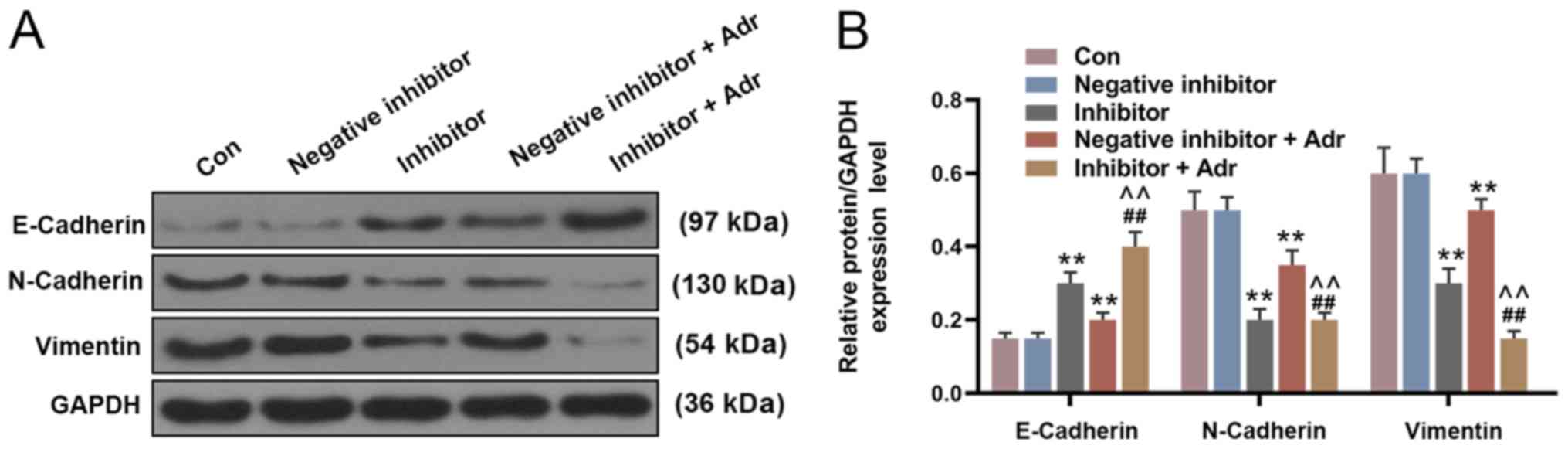
miR-130a knockdown increases the
sensitivity of AML cells to Adr
To investigate the effect of miR-130a knockdown on
the sensitivity of AML cells to Adr, miR-130a inhibitor and
negative inhibitor were transfected into HL-60 cells alone or in
combination with Adr (0.1 µmol/l). miR-130a knockdown not only
directly reduced the viability of HL-60 cells compared with the
negative inhibitor group, but also enhanced the inhibitory effect
of Adr on cell viability (P<0.05; Fig. 4A). Additionally, miR-130a knockdown
further promoted Adr-induced inhibition of cell invasion,
intensifying the anti-cell invasion function of Adr (P<0.001;
Fig. 4B and C). Furthermore,
miR-130a inhibitor promoted the inhibitory effect of Adr on
miR-130a expression (P<0.001; Fig.
4D). The WB results showed similar Adr-induced effects on
EMT-related protein expression. miR-130a knockdown increased
E-Cadherin expression levels and decreased the expression levels of
N-Cadherin and Vimentin in HL-60 cells compared with the negative
inhibitor group. In addition, Adr in combination with miR-130a
inhibitor displayed increased regulatory effects on EMT-related
protein expression compared with either treatment alone
(P<0.001; Fig. 5).
Discussion
AML is a malignant clonal disease of hematopoietic
stem cells, and is often accompanied by a variety of genetic
alterations, such as patients with t(8;21) (q22;q22)
[RUNX1/RUNX1T1], inv(16)(p13q22) [CBFB/MYH11] and t(15;17)(q24;q21)
[PML/RARA] have a favorable prognosis with good response to
treatment and complete remissions (19); IDH1 and IDH2 mutations are
recurring genetic changes in AML, they constitute a poor prognostic
factor in CN-AML with mutated NPM1 without FLT3-ITD (20). Furthermore, the poor prognosis of
patients with AML is usually related to the presence of
multidrug-resistant leukemia cells (21,22).
Adr, a topoisomerase inhibitor, shows an anti-tumor role during AML
by restraining the synthesis of double-stranded DNA to induce
leukemic cell apoptosis (23,24).
In the present study, Adr treatment downregulated the expression
level of miR-130a in AML cells. miR-130a overexpression increased
the viability and invasion of Adr-treated AML cells, while miR-130a
knockdown enhanced the inhibitory effects of Adr on AML cells. In
addition, Adr displayed a dose-dependent relationship with miR-130a
expression, such that increasing concentrations of Adr decreased
the expression levels of miR-130a. The results suggested that
miR-130a upregulation may be an important cause of Adr resistance
in AML, whereas miR-130a downregulation may serve an effective
therapeutic strategy to increase the sensitivity of AML cells to
Adr.28
In previous studies, miR-130a has been reported to
promote or inhibit tumor growth in different types of cancer. For
example, Kong et al (25)
reported that miR-130a-3p expression levels were reduced in human
breast cancer tissues, and miR-130a-3p overexpression decreased the
proliferation and metastasis of breast cancer cells. Previous
findings suggested that miR-130a is closely related to drug
resistance and that it serves as an intermediary in drug
resistance-related signaling pathways (26). For example, Liu et al
(27) reported that miR-130a-3p
activation decreased the migration and invasion of
gemcitabine-resistant hepatoma cells. In the present study,
5-Aza-dC was used to induce Adr resistance in AML cells, and
miR-130a expression was upregulated as a result, suggesting that
miR-130a may be involved in tumor cell resistance to Adr. The
results also suggested that miR-130a overexpression increased the
viability and invasion of AML cells, indicating that miR-130a
overexpression reversed the anti-AML effects of Adr. Consistent
with the present study, Ding et al (15) reported that miR-130a was abnormally
upregulated in adult patients with AML, and miR-130a knockdown
increased the sensitivity of AML cells to etoposide. Moreover,
another study reported that miR-130a overexpression stimulated the
propagation of ovarian cancer cells to accelerate disease
progression (16).
The effects of miR-130a knockdown on Adr-resistant
AML cells were also investigated. Contrasting to the effect of
miR-130a overexpression, miR-130a knockdown significantly decreased
the viability of AML cells, and promoted the anti-AML effects of
Adr. Based on the results, it was hypothesized that miR-130a
knockdown increased the sensitivity of AML cells to Adr. A previous
study demonstrated that the expression level of miR-130a was
significantly reduced in patients with CML with poor prognosis, and
that a low expression was associated with a short overall survival
time (28). Moreover, Feng et
al (29) reported that
miR-130a overexpression reduced cisplatin-sensitivity of cervical
cancer cells, and a low expression of miR-130a restored the
chemoresistance of the cancer cells to cisplatin. The present study
and the aforementioned previous studies suggested that miR-130a
upregulation was related to the pathogenesis and progression of
AML, and also contributed to the occurrence of Adr resistance.
To further verify the effects of miR-130a on AML
chemoresistance, the present study detected the expression of
EMT-related proteins, including E-Cadherin, N-Cadherin and
Vimentin. EMT is a phenotypic transition, during which cells
transform from an epithelial state to a mesenchymal state, in terms
of cell functional characteristics and morphology (30,31).
The transition of epithelial cells into mesenchymal cells results
in increased invasive and migratory abilities, which can lead to
metastasis during cancer progression (32,33).
Cancer cells have shown increased levels of N-cadherin and
Vimentin, and decreased levels of E-cadherin during the transition
from epithelial cell to mesenchymal cell (34,35).
In the present study, Adr promoted E-cadherin expression, and
downregulated N-cadherin and Vimentin expression in AML cells.
However, miR-130a overexpression inhibited Adr-induced E-cadherin
expression, and upregulated N-cadherin and Vimentin expression. By
contrast, miR-130a knockdown indicated the opposite effect and
promoted Adr-induced effects. The results suggested that miR-130a
overexpression increased AML cell Adr resistance by promoting EMT,
whereas miR-130a downregulation increased the sensitivity of AML
cells to Adr by regulating the expression of EMT-related proteins.
However, the present study had a number of limitations. A key
limitation of the present study was that investigations were only
conducted using one cell line; therefore, the results of the
present study need to be verified in multiple cell lines and in
in vitro models.
In conclusion, the expression level of miR-130a was
increased in Adr-resistant AML cells, and miR-130a knockdown
promoted the inhibitory effects of Adr on AML cell viability,
invasion and EMT. Therefore, miR-130a knockdown increased the
sensitivity of AML cells to Adr and may serve as a therapeutic
target for Adr-resistant AML.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HL conceived and designed the study. ML, JZ and YL
acquired, analyzed and interpreted the data. HL drafted and
critically revised the manuscript for important intellectual
content. All authors provided final approval of the version to be
published. ML agreed to be held accountable for all aspects of the
work in ensuring that questions related to the accuracy or
integrity of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
Adr
|
Adriamycin
|
|
AML
|
acute myeloid leukemia
|
|
CCK-8
|
Cell Counting Kit-8
|
|
RT-qPCR
|
reverse transcription-quantitative
PCR
|
|
EMT
|
epithelial-mesenchymal transition
|
|
WB
|
western blotting
|
|
miR
|
microRNA
|
References
|
1
|
Li S, Mason CE and Melnick A: Genetic and
epigenetic heterogeneity in acute myeloid leukemia. Curr Opin Genet
Dev. 36:2810–106. 2016. View Article : Google Scholar
|
|
2
|
Juliusson G and Hough R: Leukemia. Prog
Tumor Res. 43:87–100. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
De Kouchkovsky I and Abdul-Hay M: Acute
myeloid leukemia: A comprehensive review and 2016 update. Blood
Cancer J. 6:e4412016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yamamoto JF and Goodman MT: Patterns of
leukemia incidence in the United States by subtype and demographic
characteristics, 1997–2002. Cancer Causes Control. 19:379–390.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pulte D, Gondos A and Brenner H: Expected
long-term survival of patients diagnosed with acute myeloblastic
leukemia during 2006–2010. Ann Oncol. 21:335–341. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Meyers J, Yu Y, Kaye JA and Davis KL:
Medicare fee-for-service enrollees with primary acute myeloid
leukemia: An analysis of treatment patterns, survival, and
healthcare resource utilization and costs. Appl Health Econ Health
Policy. 11:275–286. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bose P, Vachhani P and Cortes JE:
Treatment of relapsed/refractory acute myeloid leukemia. Curr Treat
Options Oncol. 18:172017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
McCall MN, Kim MS, Adil M, Patil AH, Lu Y,
Mitchell CJ, Leal-Rojas P, Xu J, Kumar M, Dawson VL, et al: Toward
the human cellular microRNAome. Genome Res. 27:1769–1781. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mohr AM and Mott JL: Overview of microRNA
biology. Semin Liver Dis. 35:3–11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rupaimoole R and Slack FJ: MicroRNA
therapeutics: Towards a new era for the management of cancer and
other diseases. Nat Rev Drug Discov. 16:203–222. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Acunzo M, Romano G, Wernicke D and Croce
CM: MicroRNA and cancer-a brief overview. Adv Biol Regul. 57:1–9.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang X, Chen H, Bai J and He A: MicroRNA:
An important regulator in acute myeloid leukemia. Cell Biol Int.
41:936–945. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li Q, Song W and Wang J: TUG1 confers
Adriamycin resistance in acute myeloid leukemia by epigenetically
suppressing miR-34a expression via EZH2. Biomed Pharmacother.
109:1793–1801. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Szymczyk A, Chocholska S, Macheta A,
Szczepanek D, Hus M and Podhorecka M: Assessment of microRNA
expression in leukemic cells as predictors of sensitivity to purine
nucleoside analogs, fludarabine and cladribine, in chronic
lymphocytic leukemia patients. Cancer Manag Res. 11:5021–5031.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ding C, Chen SN, Macleod RAF, Drexler HG,
Nagel S, Wu DP, Sun AN and Dai HP: miR-130a is aberrantly
overexpressed in adult acute myeloid leukemia with t(8;21) and its
suppression induces AML cell death. Ups J Med Sci. 123:19–27. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wei H, Cui R, Bahr J, Zanesi N, Luo Z,
Meng W, Liang G and Croce CM: miR-130a deregulates PTEN and
stimulates tumor growth. Cancer Res. 77:6168–6178. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yin S, Zhang Q, Wang Y, Li S and Hu R:
MicroRNA-130a regulated by HPV18 E6 promotes proliferation and
invasion of cervical cancer cells by targeting TIMP2. Exp Ther Med.
17:2837–2846. 2019.PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lagunas-Rangel FA, Chavez-Valencia V,
Gomez-Guijosa MA and Cortes-Penagos C: Acute myeloid
leukemia-genetic alterations and their clinical prognosis. Int J
Hematol Oncol Stem Cell Res. 11:328–339. 2017.PubMed/NCBI
|
|
20
|
Paschka P, Schlenk RF, Gaidzik VI, Habdank
M, Krönke J, Bullinger L, Späth D, Kayser S, Zucknick M, Götze K,
et al: IDH1 and IDH2 mutations are frequent genetic alterations in
acute myeloid leukemia and confer adverse prognosis in
cytogenetically normal acute myeloid leukemia with NPM1 mutation
without FLT3 internal tandem duplication. J Clin Oncol.
28:3636–3643. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kater L, Claffey J, Hogan M, Jesse P,
Kater B, Strauss S, Tacke M and Prokop A: The role of the intrinsic
FAS pathway in Titanocene Y apoptosis: The mechanism of overcoming
multiple drug resistance in malignant leukemia cells. Toxicol In
Vitro. 26:119–124. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Madanat YF, Kalaycio ME and Nazha A:
Advances in acute myeloid leukemia genomics, where do we stand in
2018? Acta Med Acad. 48:35–44. 2019.PubMed/NCBI
|
|
23
|
Rivankar S: An overview of doxorubicin
formulations in cancer therapy. J Cancer Res Ther. 10:853–858.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Han YQ, Hong Y, Su XL and Wang JR:
Quercetin enhances the anti-leukemic effect of adriamycin. Zhongguo
Shi Yan Xue Ye Xue Za Zhi. 22:1555–1560. 2014.(In Chinese).
PubMed/NCBI
|
|
25
|
Kong X, Zhang J, Li J, Shao J and Fang L:
miR-130a-3p inhibits migration and invasion by regulating RAB5B in
human breast cancer stem cell-like cells. Biochem Biophys Res
Commun. 501:486–493. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang HD, Jiang LH, Sun DW, Li J and Ji
ZL: The role of miR-130a in cancer. Breast cancer. 24:521–527.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu Y, Li Y, Wang R, Qin S, Liu J, Su F,
Yang Y, Zhao F, Wang Z and Wu Q: miR-130a-3p regulates cell
migration and invasion via inhibition of Smad4 in gemcitabine
resistant hepatoma cells. J Exp Clin Cancer Res. 35:192016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu X, Zhao H, Lin Z and Zhang G:
Functional studies of miR-130a on the inhibitory pathways of
apoptosis in patients with chronic myeloid leukemia. Cancer Gene
Ther. 22:573–580. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Feng C, Ma F, Hu C, Ma JA, Wang J, Zhang
Y, Wu F, Hou T, Jiang S, Wang Y and Feng Y: SOX9/miR-130a/CTR1 axis
modulates DDP-resistance of cervical cancer cell. Cell Cycle.
17:448–458. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen T, You Y, Jiang H and Wang ZZ:
Epithelial-mesenchymal transition (EMT): A biological process in
the development, stem cell differentiation, and tumorigenesis. J
Cell Physiol. 232:3261–3272. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dominguez C, David JM and Palena C:
Epithelial-mesenchymal transition and inflammation at the site of
the primary tumor. Semin Cancer Biol. 47:177–184. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liao TT and Yang MH: Revisiting
epithelial-mesenchymal transition in cancer metastasis: The
connection between epithelial plasticity and stemness. Mol Oncol.
11:792–804. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee HH, Jung J, Moon A, Kang H and Cho H:
Antitumor and anti-invasive effect of apigenin on human breast
carcinoma through suppression of IL-6 expression. Int J Mol Sci.
20:31432019. View Article : Google Scholar
|
|
35
|
Serrano-Gomez SJ, Maziveyi M and Alahari
SK: Regulation of epithelial-mesenchymal transition through
epigenetic and post-translational modifications. Mol Cancer.
15:182016. View Article : Google Scholar : PubMed/NCBI
|