Although previous studies have discovered numerous
RNA species, protein coding RNAs account for only the minority of
the transcriptome (1,2). The majority of RNA transcripts are
actually comprised of non-coding RNAs (1,2).
Noncoding RNA can be divided into two categories according to
nucleotide (nt) length: i) Long non-coding RNA (lncRNA; >200
nt); and ii) short noncoding RNA (<200 nt) (3). The majority of previous studies
initially considered lncRNA to be a type of transcriptional noise
(4). However, further research
over the past decade revealed that lncRNAs serve an important roles
in a wide of range of biological functions, including cell growth
and developmental processes (5).
There are several methods used for classifying lncRNAs, one of
which categorizes lncRNAs based on their position relative to
protein-coding genes: i) Intergenic lncRNA; ii) intron lncRNA; iii)
overlapping lncRNA; and iv) antisense lncRNA (6). Currently, although the regulation of
gene expression of short non-coding RNAs, such miRNAs, has been
relatively well studied, reports on the effects of lncRNAs on gene
expression remain insufficient (7). However, numerous previous studies
have indicated that lncRNAs serve a variety of roles in regulating
cell signaling, such as the JAK/STAT, PI3K/Akt and Wnt signaling
pathway (8–10).
A well-known lncRNA, growth arrest-specific
transcript (GAS) 5, has recently become the focus of research
attention. The GAS1-5 paralogs constitute the GAS multigene family
(11). GAS1-5 proteins share a
common N-terminal domain but exhibit different C-terminal
extensions (12). GAS5a and GAS5b
are produced by the alternative splicing of 12 exons at the 1q25
locus in the GAS5 gene (13).
Previous studies have reported that GAS5 serves an anti-cancer
effect in various types of cancer, including gastric cancer
(14), lung cancer (15), hepatocellular carcinoma (16) and renal cancer (17). The accumulation of GAS5 has been
found to inhibit cancer growth, invasion and metastasis, and
enhance cell sensitivity to chemotherapy drugs (18–21).
Since elucidating the underlying molecular mechanism of GAS5
function is becoming a subject of intense investigation, the
present review aims to discuss the current knowledge on the
GAS5-mediated regulation of cell signaling (Table I).
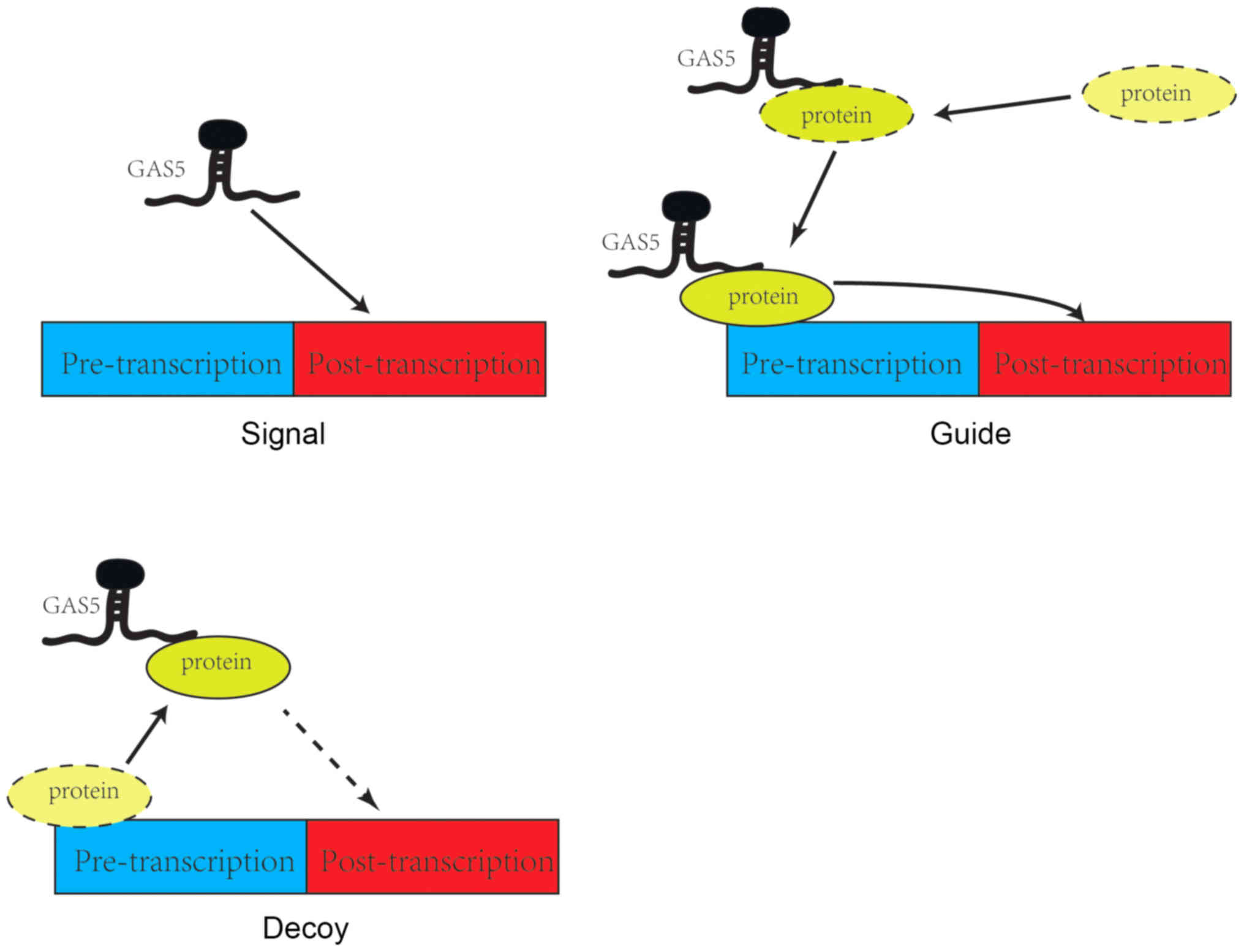
There are three modes of action by which GAS5
participates in signaling pathways. Firstly, GAS5 can serve as a
signaling protein, where it is specifically transcribed following
different triggers and then participates in signal transmission in
a number of pathways as a signaling node (22). For instance, experiments have
indicated that after DNA damage in human colorectal tissue cells,
GAS5 increases the expression of snoRNA, and GAS5 snoRNA activates
the p53 signaling pathway and responds to DNA damage. GAS5 serves
as a signaling protein in the aforementioned process and directly
participates in the regulation of p53 signaling pathway (23). Secondly, GAS5 can act as a decoy,
where it can behave as a molecular sponge that binds directly to
target RNA or proteins, thereby blocking their downstream functions
(24). For example, GAS5 binds to
miR-21 to prevent its inhibitory action on PTEN and PDCD4, thereby
upregulating the expression of PTEN and PDCD4 (25). Thirdly, GAS5 can serve a guide role
as a transport protein, where upon binding, it guides the protein
to a specific DNA sequence where it can regulate the transcription
of downstream molecules (26).
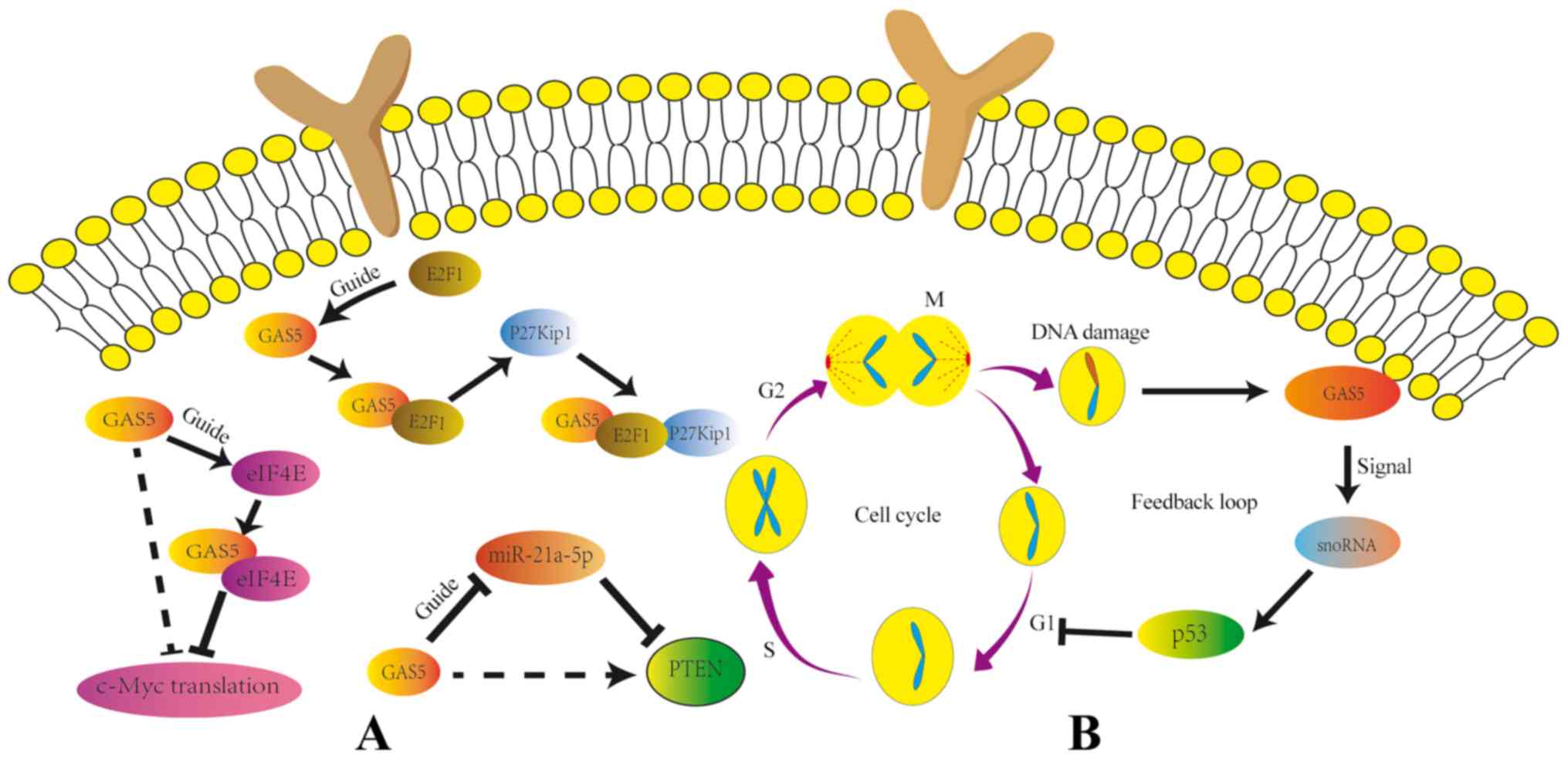
GAS5 has been previously reported to promote the binding of E2F
transcription factor 1 (E2F1) to the promoter of the
cyclin-dependent kinase inhibitor 1B (P27kip1) gene through
guidance to activate P27kip1 (27)
(Fig. 1).
GAS5 is involved in PTEN regulation through a number
of mechanisms. PTEN is an important tumor suppressor that serves an
important role in cell growth, apoptosis and migration (34). GAS5 may indirectly facilitate the
translation of PTEN by repressing miR-21a-5p (35). Additionally, in papillary thyroid
carcinoma, GAS5 can serve as a molecular sponge for miR-222-3p,
which activates the PTEN/AKT pathway (36). Similarly, GAS5 has been found to
sponge miR-222 to indirectly suppress PTEN expression in breast
cancer (37).
p53 is an extensively studied tumor suppressor gene
that regulates cell cycle progression and prevents carcinogenesis
(38). p53 is therefore termed the
‘guardian of the genome’ (39).
The predominant role of p53 is to maintain genome stability and
avoid mutation. A number of lncRNAs have been found to regulate the
p53 network (40,41). Previous studies have indicated that
reduced GAS5 expression is associated with cell cycle arrest via
increased p53 expression, breast cancer gene type 1 and growth
damage and DNA damage inducible α expression (40–42).
In colorectal cancer cell lines, p53 expression has been previously
found to associate closely with GAS5-derived snoRNA levels
(23). Therefore, GAS5 may
regulate p53 expression via a feedback loop, the mechanism of which
requires further investigation. The feedback pathway is activated
after DNA damage as GAS5 derives snoRNA, which directly promotes
the expression of p53 and inhibits the G1 phase in the
middle stage of cell division, which inhibits DNA damage in cells
undergoing mitosis (Fig. 2B).
mTOR is involved in numerous signaling pathways,
including cell proliferation, differentiation, autophagy and
apoptosis (43). A regulatory
relationship between mTOR and GAS5 has been previously found
(44,45). Several studies have demonstrated
that GAS5 is expressed at reduced levels in gastric cancer cells
(46,47). GAS5 overexpression has been
revealed to inhibit the proliferation of gastric cancer cells by
reducing mTOR expression, suggesting that GAS5 exerts its
biological function through the mTOR pathway (48). One study reported that the
expression level of GAS5 in gastric cancer is negatively associated
with that of miR-106a-5p. Overexpression of miR-106a-5p reverses
the biological effects induced by GAS5 overexpression, which occurs
via Akt/mTOR (49). This shows
that GAS5 overexpression serves a role in inhibiting proliferation
in gastric cancer cells by inhibiting the function of miR-106a-5p,
thereby reducing mTOR expression (49). In patients with esophageal cancer,
GAS5 expression has been demonstrated to be reduced. In esophageal
cancer cell lines, GAS5 overexpression has been found to inhibit
proliferation, migration, reduce PI3K expression and AKT and mTOR
phosphorylation. The use of PI3K agonists reversed these
aforementioned effects without affecting the expression of GAS5
(50,51). This suggests that GAS5 mediates its
effects through the PI3K/mTOR pathway in esophageal cancer cells
(50,51). Additionally, a previous study on
glioma cells revealed that GAS5 inhibits excessive autophagy
induced by cisplatin exposure by activating the mTOR signaling
pathway (52). It was found that
treatment with rapamycin (mTOR antagonist) can eliminate
GAS5-mediated inhibition of autophagy caused by cisplatin and
reduce GAS5 sensitivity to cisplatin (52).
In hepatocellular carcinoma (HCC) tissues, GAS5,
PDCD4 and PTEN levels were reduced whilst miR-21 levels were
increased compared with those in adjacent normal tissue (60). Hepatocellular carcinoma (HCC) is a
common tumor with high malignancy and poor prognosis. Furthermore,
the expression levels of GAS5 and miR-21 are associated with the
clinical features of HCC (60).
Lower GAS5 expression levels are correlated with higher miR-21
expression levels (60). Compared
with patients with liver cancer with low GAS5 expression levels,
patients with liver cancer with high GAS5 expression levels survive
longer (60). GAS5 acts as a
negative regulator in liver cancer through miR-21 and its targets,
which are involved in the migration and invasion of cancer cells
(61). This indicates that GAS5
may be a target for the treatment of HCC (61). Additionally, increased expression
of GAS5 may lead to increased corticotropin-releasing hormone
receptor 1 expression and long-term activation of the
hypothalamic-pituitary-adrenal axis, possibly due to the inhibition
of miR449a by GAS5 (62).
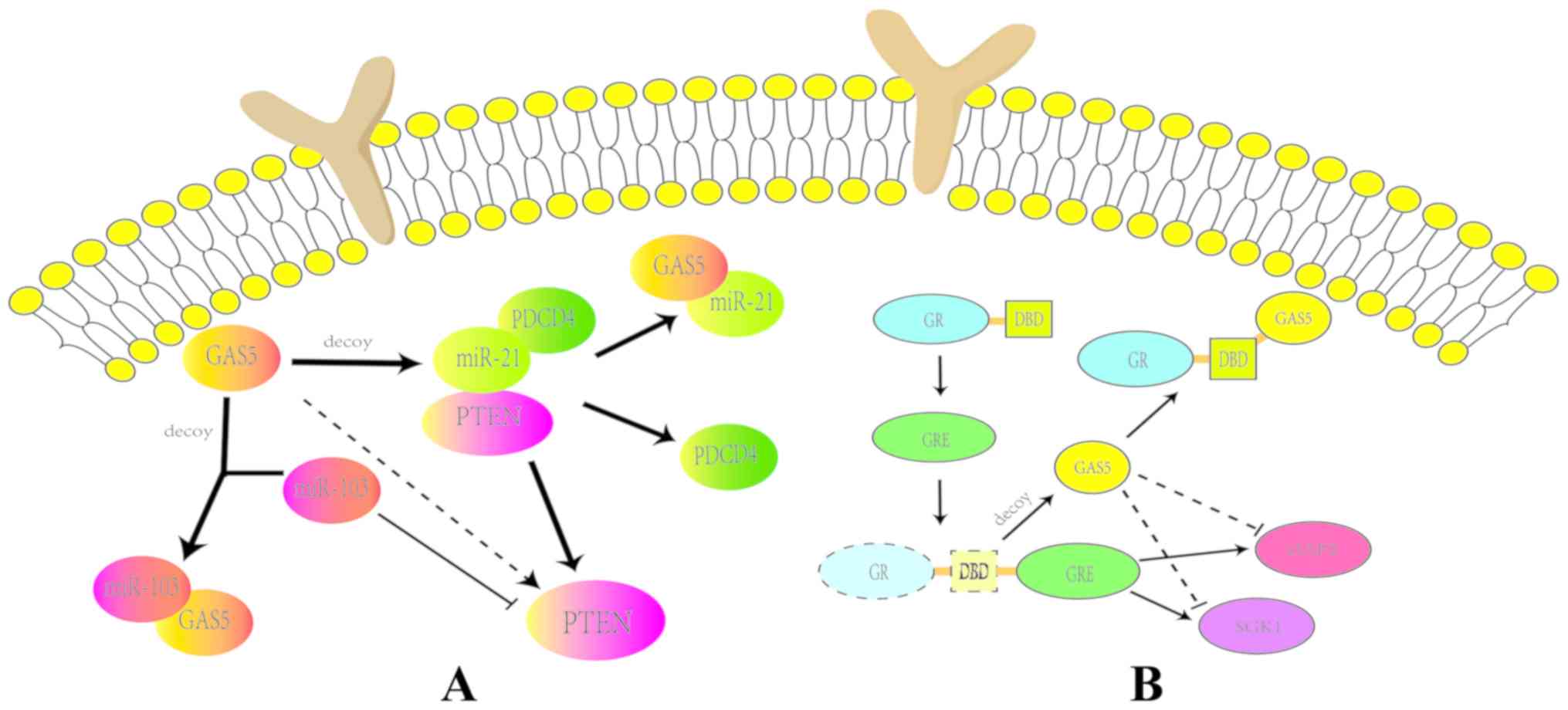
Glucocorticoid receptors (GRs) are important members
of the nuclear-receptor family and common transcription regulators
depending on hormone levels (63).
Following hormone binding, GRs regulate the expression of target
genes, including apoptosis genes cIAP2 and SGK1, by interacting
with the glucocorticoid response element (GRE) (64). Via the aforementioned signaling
pathways, GR can regulate the process of cell apoptosis. GAS5 and
GR interact as GAS5 is considered to be a sponge for GR, suggesting
that GAS5 is a possible factor for the control and maintenance of
glucocorticoid sensitivity and drug resistance (65). Previous studies have demonstrated
that GAS5 competes with GRE to bind the DNA-binding domain (DBD)
sites on GRs (66,67). This process blocks GR binding to
the GRE, consequently repressing the transcriptional activation of
target genes (Fig. 3B) (68).
Activation of the AKT pathway occurs through a
variety of signaling pathways, including those of MAPK, PI3K/AKT
and mTOR. The AKT pathway is closely associated with the
proliferative and invasive abilities of tumor cells. GAS5 serves an
important role in the regulation of the AKT pathway (69,70).
Previous studies have reported that GAS5 interacts with the PDCD4
protein (71–73). In cardiomyocytes, GAS5
competitively binds to miR-21, thereby positively regulating PDCD4
expression. The enhanced expression of PDCD4 in turn inhibits the
PI3K/AKT signaling pathway (74).
This indicates that GAS5 regulates the expression of PDCD4 by
targeting miR-21 and indirectly inhibiting the PI3K/AKT signaling
pathway (74). Inhibiting the
expression of GAS5 activates the PI3K/AKT signaling pathway and
increases the expression of its downstream proteins matrix
metallopeptidase 9 and tumor protein p53 in human trophoblast cell
lines HTR-8/SVneo and JEG-3 (75).
GAS5 competitively binds to miR-532-5p such that silencing GAS5
expression upregulates the expression of miR-532-5p to activate the
PI3K/AKT signaling pathway in myocardial ischemia reperfusion
injury (76). This indicates that
GAS5 regulates the PI3K/AKT signaling pathway by binding with
miR-532-5p (76). Furthermore, in
ischaemic brain injury, GAS5 enhances the expression level of PTEN
by regulating the expression of miR-21, thereby inhibiting the
PI3K/AKT signaling pathway (77).
In cardiomyocytes, GAS5 knockdown has been found to suppress the
expression of Rho associated coiled-coil containing protein kinase
1 (ROCK1), activating the PI3K/AKT signaling pathway, which in turn
suppress the expression of glycogen synthase kinase 3β (GSK-3β).
miR-335 can bind to GAS5 and inhibit its expression. When miR-335
is inhibited, the expression of GAS5 is increased and the
aforementioned effects are abolished (78). This suggests that GAS5 exerts its
biological function through the miR-335/ROCK1/AKT/GSK-3β axis
(78). Furthermore, under hypoxic
conditions, cell survival rate is reduced whereas apoptosis ability
is enhanced. GAS5 expression in myocardial cells is increased and
the PI3K/AKT signaling pathway is suppressed. Inhibiting the
expression of GAS5 improves the viability of cells and inhibits
cell apoptosis. Additionally, the expression of miR-222-3p, a
target gene of GAS5, was found to be downregulated under hypoxic
conditions (79). Overexpression
of miR-222-3p reversed the effects of GAS5 on cell survival and
apoptosis under hypoxic conditions by activating the PI3K/AKT
signaling pathway (79).
In short, GAS5 exerts its functions via three modes
of action, namely signal, decoy and guide. In lung cancer, GAS5 can
enhance the effect of gefitinib by inhibiting epidermal growth
factor and downregulating the expression of insulin-like growth
factor receptor 1 (22). GAS5 is
underexpressed in esophageal cancer cells, and overexpression of
GAS5 inhibits the proliferation and migration of tumor cells, and
inhibits the expression of PI3K and mTOR signaling pathways
(35). In hepatocellular
carcinoma, GAS5 is involved in downregulating the invasion and
migration of liver cancer cells by interacting with miR-21, and the
level of GAS5 is positively correlated with the survival rate of
patients with liver cancer (39).
In myocardial ischemia-reperfusion injury, GAS5 can combine with
miR-532-5p to inhibit the PI3K/AKT signaling pathway, and silencing
GAS5 expression can upregulate miR-532-5p to activate the PI3K/AKT
signaling pathway to reduce myocardial damage (47). GAS5 can act as a sponge for GR to
block the GR/GRE/cIAP2 and SGK1 signaling pathways, and achieve the
effect of controlling and maintaining glucocorticoid sensitivity
and drug resistance (43).
GAS5 serves a key role in various signaling pathways
and has a role in regulating gene expression. For example, GAS5 can
act as a guide to regulate the translation of P27Kip1, PTEN and
c-Myc whilst regulating the expression of p53 by acting as a
signaling molecule to form a feedback loop (18). GAS5 expression is closely
associated with the degree of mTOR expression. Overexpression of
GAS5 reduces the expression of mTOR and miRNA-106a-5p in a
PI3K-dependent manner (34).
Furthermore, GAS5 interacts with miRNA primarily through sponging.
In this process, GAS5 compete to bind to a key gene or protein in a
pathway to alter their expression levels (37). Additionally, GAS5 can serve as a
molecular sponge in the GRE pathway, where it competes with GRE for
the DBD structure on GRs (43).
GAS5 is also involved in the regulation of AKT, since GAS5
inhibition activates the AKT signaling pathway (45). Furthermore, PDCD4, miR-532-5p and
ROCK1 all serve important roles in the regulation process of this
pathway. Increasing the expression of GAS5 commonly inhibits the
development of tumors and enhances the therapeutic effect of drugs
on tumor cells.
In conclusion, these data indicate GAS5 serves key
roles in miRNA, p53, mTOR, GRE and AKT signaling pathways. However,
the mechanism of action of GAS5 remains unclear and further
experiments are required to elucidate its functions. In gastric
cancer, lung cancer, hepatocellular carcinoma and renal cancer, the
role of GAS5 is similar to that of tumor suppressor genes. However,
whether it is expressed at low levels only in these specific
diseases or expressed at low levels in all diseases requires
further research. Fortunately, there are continuous novel
developments in this field.
Thanks to Mr. Wei Hu (Department of Andrology, The
First Affiliated Hospital of University of South China) for
assisting with the preparation of the current review.
The present study was supported by the National
Natural Science Foundation of China (grant no. 81402100), the
Foundation for ‘Young Medical Talents’ of Jiangsu Province (grant
no. QNRC2016840), the ‘Six Talent Peaks Project’ in Jiangsu
Province (grant no. WSW-007) and the Natural Science Foundation of
Hunan Province (grant no. 2019JJ40269).
Data sharing is not applicable to this article, as
no datasets were generated or analyzed during the current
study.
YZ completed the majority of the current review,
including the preparation of the manuscript. BC provided specific
academic guidance and assisted in improving the manuscript. All
authors read and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Philippe N, Bou Samra E, Boureux A,
Mancheron A, Rufflé F, Bai Q, De Vos J, Rivals E and Commes T:
Combining DGE and RNA-sequencing data to identify new polyA+
non-coding transcripts in the human genome. Nucleic Acids Res.
42:3049–2832. 2014. View Article : Google Scholar
|
|
2
|
Pauli A, Valen E and Schier AF:
Identifying (non-)coding RNAs and small peptides: Challenges and
opportunities. Bioessays. 37:103–112. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shi Q and Yang X: Circulating MicroRNA and
long noncoding RNA as biomarkers of cardiovascular diseases. J Cell
Physiol. 231:751–755. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xu S, Gong Y, Yin Y, Xing H and Zhang N:
The multiple function of long noncoding RNAs in osteosarcoma
progression, drug resistance and prognosis. Biomed Pharmacother.
127:1101412020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ricciuti B, Mencaroni C, Paglialunga L,
Paciullo F, Crinò L, Chiari R and Metro G: Long noncoding RNAs: New
insights into non-small cell lung cancer biology, diagnosis and
therapy. Med Oncol. 33:182016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yousefi H, Maheronnaghsh M, Molaei F,
Mashouri L, Reza Aref A, Momeny M and Alahari SK: Long noncoding
RNAs and exosomal lncRNAs: Classification, and mechanisms in breast
cancer metastasis and drug resistance. Oncogene. 39:953–974. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Panir K, Schjenken JE, Robertson SA and
Hull ML: Non-coding RNAs in endometriosis: A narrative review. Hum
Reprod Update. 24:497–515. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang DW, Zhang X, Qian GB, Jiang MJ, Wang
P and Wang KZ: Downregulation of long noncoding RNA LOC101928134
inhibits the synovial hyperplasia and cartilage destruction of
osteoarthritis rats through the activation of the Janus
kinase/signal transducers and activators of transcription signaling
pathway by upregulating IFNA1. J Cell Physiol. 234:10523–10534.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu Q and Yi X: Down-regulation of long
noncoding RNA MALAT1 protects hippocampal neurons against excessive
autophagy and apoptosis via the PI3K/Akt Signaling Pathway in rats
with epilepsy. J Mol Neurosci. 65:234–245. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang L, Fang Y, Cheng X, Lian YJ and Xu
HL: Silencing of long noncoding RNA SOX21-AS1 relieves neuronal
oxidative stress injury in mice with Alzheimer's disease by
upregulating FZD3/5 via the Wnt Signaling Pathway. Mol Neurobiol.
56:3522–3537. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rolli E, Ragni E, de Medina-Redondo M,
Arroyo J, de Aldana CR and Popolo L: Expression, stability, and
replacement of glucan-remodeling enzymes during developmental
transitions in Saccharomyces cerevisiae. Mol Biol Cell.
22:1585–1598. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ragni E, Fontaine T, Gissi C, Latgè JP and
Popolo L: The Gas family of proteins of Saccharomyces cerevisiae:
Characterization and evolutionary analysis. Yeast. 24:297–308.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schneider C, King RM and Philipson L:
Genes specifically expressed at growth arrest of mammalian cells.
Cell. 54:787–793. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dong X, Gao W, Lv X, Wang Y, Wu Q, Yang Z,
Mao G and Xing W: Association between lncRNA GAS5, MEG3, and PCAT-1
polymorphisms and cancer risk: A Meta-Analysis. Dis Markers.
2020:67234872020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shi X, Sun M, Liu H, Yao Y, Kong R, Chen F
and Song Y: A critical role for the long non-coding RNA GAS5 in
proliferation and apoptosis in non-small-cell lung cancer. Mol
Carcinogen. 54 (Suppl 1):E1–E12. 2015. View
Article : Google Scholar
|
|
16
|
Zhao P, Cui X, Zhao L, Liu L and Wang D:
Overexpression of Growth-Arrest-Specific Transcript 5 improved
cisplatin sensitivity in hepatocellular carcinoma through Sponging
miR-222. DNA Cell Biol. 39:724–732. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang W, Zhang K, Li L, Ma K, Hong B, Gong
Y and Gong K: Discovery and validation of the prognostic value of
the lncRNAs encoding snoRNAs in patients with clear cell renal cell
carcinoma. Aging (Albany NY). 12:4424–4444. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hu G, Lou Z and Gupta M: The long
non-coding RNA GAS5 cooperates with the eukaryotic translation
initiation factor 4E to regulate c-Myc translation. PLoS One.
9:e1070162014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu Y, Zhang L, Wang Y, Li H, Ren X, Wei F,
Yu W, Wang X, Zhang L, Yu J and Hao X: Long noncoding RNA HOTAIR
involvement in cancer. Tumor Biol. 35:9531–9538. 2014. View Article : Google Scholar
|
|
20
|
Yan L, Zhou J, Gao Y, Ghazal S, Lu L,
Bellone S, Yang Y, Liu N, Zhao X, Santin AD, et al: Regulation of
tumor cell migration and invasion by the H19/let-7 axis is
antagonized by metformin-induced DNA methylation. Oncogene.
34:3076–3084. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li S, Wang Q, Qiang Q, Shan H, Shi M, Chen
B, Zhao S and Yuan L: Sp1-mediated transcriptional regulation of
MALAT1 plays a critical role in tumor. J Cancer Res Clin Oncol.
141:1909–1920. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang L, Wang C and Shen X: lncRNA GAS5
suppresses ER stress-induced apoptosis and inflammation by
regulating SERCA2b in HG-treated retinal epithelial cell. Mol Med
Rep. 22:1072–1080. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Krell J, Frampton AE, Mirnezami R, Harding
V, De Giorgio A, Roca Alonso L, Cohen P, Ottaviani S, Colombo T,
Jacob J, et al: Growth arrest-specific transcript 5 associated
snoRNA levels are related to p53 expression and DNA damage in
colorectal cancer. PLoS One. 9:e985612014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qian X, Xu C, Zhao P and Qi Z: Long
non-coding RNA GAS5 inhibited hepatitis C virus replication by
binding viral NS3 protein. Virology. 492:155–165. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang Z, Zhu Z, Watabe K, Zhang X, Bai C,
Xu M, Wu F and Mo YY: Negative regulation of lncRNA GAS5 by miR-21.
Cell Death Differ. 20:1558–1568. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu X, Jiao T, Wang Y, Su W, Tang Z and
Han C: Long non-coding RNA GAS5 acts as a molecular sponge to
regulate miR-23a in gastric cancer. Minerva Med. 2016.(Epub sahead
of print).
|
|
27
|
Luo G, Liu D, Huang C, Wang M, Xiao X,
Zeng F, Wang L and Jiang G: LncRNA GAS5 inhibits cellular
proliferation by targeting P27Kip1. Mol Cancer Res.
15:789–799. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Eilebrecht S, Wilhelm E, Benecke BJ, Bell
B and Benecke AG: HMGA1 directly interacts with TAR to modulate
basal and Tat-dependent HIV transcription. RNA Biol. 10:436–444.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Aminian K, Mashayekhi F, Mirzanejad L and
Salehi Z: A functional genetic variant in GAS5 lncRNA (rs145204276)
modulates p27Kip1 expression and confers risk for
gastric cancer. Br J Biomed Sci. 76:83–85. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu Y, Zhao J, Zhang W, Gan J, Hu C, Huang
G and Zhang Y: lncRNA GAS5 enhances G1 cell cycle arrest via
binding to YBX1 to regulate p21 expression in stomach cancer. Sci
Rep. 5:101592015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dong S, Qu X, Li W, Zhong X, Li P, Yang S,
Chen X, Shao M and Zhang L: The long non-coding RNA, GAS5, enhances
gefitinib-induced cell death in innate EGFR tyrosine kinase
inhibitor-resistant lung adenocarcinoma cells with wide-type EGFR
via downregulation of the IGF-1R expression. J Hematol Oncol.
8:432015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu SD, Meng WX, Xu L, Chi C, Sun X and
Liu HY: GAS5 promotes myocardial apoptosis in myocardial
ischemia-reperfusion injury via upregulating LAS1 expression. Eur
Rev Med Pharmacol Sci. 22:8447–8453. 2018.PubMed/NCBI
|
|
33
|
Cao Q, Wang N, Qi J, Gu Z and Shen H: Long
non-coding RNA-GAS5 acts as a tumor suppressor in bladder
transitional cell carcinoma via regulation of chemokine (C-C motif)
ligand 1 expression. Mol Med Rep. 13:27–34. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xing Y, Lin NU, Maurer MA, Chen H, Mahvash
A, Sahin A, Akcakanat A, Li Y, Abramson V, Litton J, et al: Phase
II trial of AKT inhibitor MK-2206 in patients with advanced breast
cancer who have tumors with PIK3CA or AKT mutations, and/or PTEN
loss/PTEN mutation. Breast Cancer Res. 21:782019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gao ZQ, Wang JF, Chen DH, Ma XS, Wu Y,
Tang Z and Dang XW: Long non-coding RNA GAS5 suppresses pancreatic
cancer metastasis through modulating miR-32-5p/PTEN axis. Cell
Biosci. 7:662017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang XF, Ye Y and Zhao SJ: lncRNA Gas5
acts as a ceRNA to regulate PTEN expression by sponging miR-222-3p
in papillary thyroid carcinoma. Oncotarget. 9:3519–3530. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gu J, Wang Y, Wang X, Zhou D, Shao C, Zhou
M and He Z: Downregulation of lncRNA GAS5 confers tamoxifen
resistance by activating miR-222 in breast cancer. Cancer Lett.
434:1–10. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
de Oliveira GAP, Petronilho EC, Pedrote
MM, Marques MA, Vieira TCRG, Cino EA and Silva JL: The status of
p53 oligomeric and aggregation states in cancer. Biomolecules.
10:5482020. View Article : Google Scholar
|
|
39
|
Silva JL, Lima CGS, Rangel LP, Ferretti
GDS, Pauli FP, Ribeiro RCB, da Silva TB, da Silva FC and Ferreira
VF: Recent synthetic approaches towards small molecule reactivators
of p53. Biomolecules. 10:6352020. View Article : Google Scholar
|
|
40
|
Mazar J, Rosado A, Shelley J, Marchica J
and Westmoreland TJ: The long non-coding RNA GAS5 differentially
regulates cell cycle arrest and apoptosis through activation of
BRCA1 and p53 in human neuroblastoma. Oncotarget. 8:6589–6607.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dong P, Xiong Y, Yue J, J B Hanley S,
Kobayashi N, Todo Y and Watari H: Exploring lncRNA-Mediated
regulatory networks in endometrial cancer cells and the tumor
microenvironment: Advances and challenges. Cancers (Basel).
11:2342019. View Article : Google Scholar
|
|
42
|
Ye D, Bao Z, Yu Y, Han Z, Yu Y, Xu Z, Ma
W, Yuan Y, Zhang L, Xu Y, et al: Inhibition of cardiomyocyte
differentiation of human induced pluripotent stem cells by
Ribavirin: Implication for its cardiac developmental toxicity.
Toxicology. 435:1524222020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zou Z, Tao T, Li H and Zhu X: mTOR
signaling pathway and mTOR inhibitors in cancer: Progress and
challenges. Cell Biosci. 10:312020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li H, Liu Y, Huang J, Liu Y and Zhu Y:
Association of genetic variants in lncRNA GAS5/miR-21/mTOR axis
with risk and prognosis of coronary artery disease among a Chinese
population. J Clin Lab Anal.e234302020.(Epub ahead of print).
|
|
45
|
Yacqub-Usman K, Pickard MR and Williams
GT: Reciprocal regulation of GAS5 lncRNA levels and mTOR inhibitor
action in prostate cancer cells. Prostate. 75:693–705. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Esfandi F, Salehnezhad T, Taheri M,
Afsharpad M, Hafez AA, Oskooei VK and Ghafouri-Fard S: Expression
assessment of a panel of long non-coding RNAs in gastric
malignancy. Exp Mol Pathol. 113:1043832020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhang N, Wang AY, Wang XK, Sun XM and Xue
HZ: GAS5 is downregulated in gastric cancer cells by promoter
hypermethylation and regulates adriamycin sensitivity. Eur Rev Med
Pharmacol Sci. 20:3199–3205. 2016.PubMed/NCBI
|
|
48
|
Li Y, Gu J and Lu H: The GAS5/miR-222 axis
regulates proliferation of gastric cancer cells through the
PTEN/Akt/mTOR Pathway. Dig Dis Sci. 62:3426–3437. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Dong S, Zhang X and Liu D: Overexpression
of long noncoding RNA GAS5 suppresses tumorigenesis and development
of gastric cancer by sponging miR-106a-5p through the Akt/mTOR
pathway. Biol Open. 8:bio0413432019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang G, Sun J, Zhao H and Li H: Long
Non-Coding RNA (lncRNA) Growth Arrest Specific 5 (GAS5) suppresses
esophageal squamous cell carcinoma cell proliferation and migration
by inactivating phosphatidylinositol 3-kinase (PI3K)/AKT/Mammalian
target of rapamycin (mTOR) signaling pathway. Med Sci Monit.
24:7689–7696. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Huang J, Li Y, Lu Z, Che Y, Sun S, Mao S,
Lei Y, Zang R, Li N, Sun N and He J: Long non-coding RNA GAS5 is
induced by interferons and plays an antitumor role in esophageal
squamous cell carcinoma. Cancer Med. 7:3157–3167. 2018. View Article : Google Scholar
|
|
52
|
Huo JF and Chen XB: Long noncoding RNA
growth arrest-specific 5 facilitates glioma cell sensitivity to
cisplatin by suppressing excessive autophagy in an mTOR-dependent
manner. J Cell Biochem. 120:6127–6136. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Rustagi Y, Jaiswal HK, Rawal K, Kundu GC
and Rani V: Comparative characterization of cardiac development
specific microRNAs: Fetal regulators for future. PLoS One.
10:e01393592015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Witwer KW and Halushka MK: Toward the
promise of microRNAs-Enhancing reproducibility and rigor in
microRNA research. RNA Biol. 13:1103–1116. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Shen X, Zhang Y, Wu X, Guo Y, Shi W, Qi J,
Cong H, Wang X, Wu X and Ju S: Upregulated lncRNA-PCAT1 is closely
related to clinical diagnosis of multiple myeloma as a predictive
biomarker in serum. Cancer Biomark. 18:257–263. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Jing Z, Gao L, Wang H, Chen J, Nie B and
Hong Q: Long non-coding RNA GAS5 regulates human B lymphocytic
leukaemia tumourigenesis and metastasis by sponging miR-222. Cancer
Biomark. 26:385–392. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wang W, Jia YJ, Yang YL, Xue M, Zheng ZJ,
Wang L and Xue YM: lncRNA GAS5 exacerbates renal tubular epithelial
fibrosis by acting as a competing endogenous RNA of miR-96-5p.
Biomed Pharmacother. 121:1094112020. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Chen D, Guo Y, Chen Y, Guo Q, Chen J, Li
Y, Zheng Q, Jiang M, Xi M and Cheng L: lncRNA growth
arrest-specific transcript 5 targets miR-21 gene and regulates
bladder cancer cell proliferation and apoptosis through PTEN.
Cancer Med. 9:2846–2858. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lyu K, Xu Y, Yue H, Li Y, Zhao J, Chen L,
Wu J, Zhu X, Chai L, Li C, et al: Long noncoding RNA GAS5 acts as a
tumor suppressor in laryngeal squamous cell carcinoma Via miR-21.
Cancer Manag Res. 11:8487–8498. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Tu ZQ, Li RJ, Mei JZ and Li XH:
Down-regulation of long non-coding RNA GAS5 is associated with the
prognosis of hepatocellular carcinoma. Int J Clin Exp Pathol.
7:4303–4309. 2014.PubMed/NCBI
|
|
61
|
Hu L, Ye H, Huang G, Luo F, Liu Y, Liu Y,
Yang X, Shen J, Liu Q and Zhang J: Long noncoding RNA GAS5
suppresses the migration and invasion of hepatocellular carcinoma
cells via miR-21. Tumor Biol. 37:2691–2702. 2016. View Article : Google Scholar
|
|
62
|
Nemoto T, Kakinuma Y and Shibasaki T:
Impaired miR449a-induced downregulation of Crhr1 expression in
low-birth-weight rats. J Endocrinol. 224:195–203. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Weikum ER, Okafor CD, D'Agostino EH,
Colucci JK and Ortlund EA: Structural analysis of the
glucocorticoid receptor ligand-binding domain in complex with
triamcinolone acetonide and a fragment of the atypical coregulator,
small heterodimer partner. Mol Pharmacol. 92:12–21. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Tani H, Torimura M and Akimitsu N: The RNA
degradation pathway regulates the function of GAS5 a non-coding RNA
in mammalian cells. PLoS One. 8:e556842013. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Lucafo M, De Iudicibus S, Di Silvestre A,
Pelin M, Candussio L, Martelossi S, Tommasini A, Piscianz E,
Ventura A and Decorti G: Long noncoding RNA GAS5: A novel marker
involved in glucocorticoid response. Curr Mol Med. 15:94–99. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Gharesouran J, Taheri M, Sayad A,
Ghafouri-Fard S, Mazdeh M and Omrani MD: The growth arrest-specific
transcript 5 (GAS5) and nuclear receptor subfamily 3 Group C Member
1 (NR3C1): Novel markers involved in multiple sclerosis. Int J Mol
Cell Med. 7:102–110. 2018.PubMed/NCBI
|
|
67
|
Ketab FNG, Gharesouran J, Ghafouri-Fard S,
Dastar S, Mazraeh SA, Hosseinzadeh H, Moradi M, Javadlar M,
Hiradfar A, Rezamand A, et al: Dual biomarkers long non-coding RNA
GAS5 and its target, NR3C1, contribute to acute myeloid leukemia.
Exp Mol. Pathol. 114:1043992020.
|
|
68
|
Kino T, Hurt DE, Ichijo T, Nader N and
Chrousos GP: Noncoding RNA gas5 is a growth arrest- and
starvation-associated repressor of the glucocorticoid receptor. Sci
Signal. 3:ra82010. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Chen L, Ren P, Zhang Y, Gong B, Yu D and
Sun X: Long non-coding RNA GAS5 increases the radiosensitivity of
A549 cells through interaction with the miR-21/PTEN/Akt axis. Oncol
Rep. 43:897–907. 2020.PubMed/NCBI
|
|
70
|
Dong Z, Li S, Wang X, Si L, Ma R, Bao L
and Bo A: lncRNA GAS5 restrains CCl4-induced hepatic
fibrosis by targeting miR-23a through the PTEN/PI3K/Akt signaling
pathway. Am J Physiol Gastrointest Liver Physiol. 316:G539–G550.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Shen Z and She Q: Association between the
deletion allele of Ins/Del polymorphism (Rs145204276) in the
promoter region of GAS5 with the risk of atherosclerosis. Cell
Physiol Biochem. 49:1431–1443. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhu L, Zhu Q, Wen H, Huang X and Zheng G:
Mutations in GAS5 affect the transformation from benign prostate
proliferation to aggressive prostate cancer by affecting the
transcription efficiency of GAS5. J Cell Physiol. 234:8928–8940.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Liu K, Liu C and Zhang Z: lncRNA GAS5 acts
as a ceRNA for miR-21 in suppressing PDGF-bb-induced proliferation
and migration in vascular smooth muscle cells. J Cell Biochem.
120:15233–15240. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Zhou XH, Chai HX, Bai M and Zhang Z:
lncRNA-GAS5 regulates PDCD4 expression and mediates myocardial
infarction-induced cardiomyocytes apoptosis via targeting MiR-21.
Cell Cycle. 19:1363–1377. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Zheng D, Hou Y, Li Y, Bian Y, Khan M, Li
F, Huang L and Qiao C: Long non-coding RNA Gas5 is associated with
preeclampsia and regulates biological behaviors of trophoblast via
MicroRNA-21. Front Genet. 11:1882020. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Han Y, Wu N, Xia F, Liu S and Jia D: Long
non-coding RNA GAS5 regulates myocardial ischemia-reperfusion
injury through the PI3K/AKT apoptosis pathway by sponging
miR-532-5p. Int J Mol Med. 45:858–872. 2020.PubMed/NCBI
|
|
77
|
Li J, Lv H and Che YQ: Long non-coding RNA
Gas5 potentiates the effects of microRNA-21 Downregulation in
response to ischaemic brain injury. Neuroscience. 437:87–97. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Wu N, Zhang X, Bao Y, Yu H, Jia D and Ma
C: Down-regulation of GAS5 ameliorates myocardial
ischaemia/reperfusion injury via the miR-335/ROCK1/AKT/GSK-3β axis.
J Cell Mol Med. 23:8420–8431. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
He J, Dai H, Zhao Q, Guo J and Chen C:
LNCRNA Gas5 suppression protected Hl-1 cells against hypoxia injury
by sponging miR-222-3p. Exp Mol Pathol. 115:1044362020. View Article : Google Scholar : PubMed/NCBI
|