Introduction
Lung cancer remains the leading cause of
cancer-associated death worldwide, and the mortality rate in 2019
was higher in males (24%) compared with females (23%) in 2019 in
USA (1). Non-small cell lung
cancer (NSCLC) is a primary form of lung cancer that accounts for
>80% of all lung cancer cases worldwide (2). NSCLC has two major subtypes based on
clinical pathology, squamous cell carcinoma and adenocarcinoma
(3). The pathogenesis of NSCLC is
complex and poorly understood, and a previous study recorded from
Brazil that the rates of recurrence (over 50%) and mortality
(69.7%) following surgery and different lines of chemotherapy
remain extremely high (4). A
previous study suggested that novel targets (ROS1 and RET fusions)
would benefit NSCLC treatment (5).
Therefore, further research on potential therapeutic targets
associated with NSCLC remains a necessity.
Long non-coding RNAs (lncRNAs) are RNA molecules
>200 nucleotides in length with no or limited protein-coding
capacity (6). lncRNAs are involved
in multiple biological processes, including cell cycle, cell
differentiation and myogenesis, by acting as microRNAs sponges or
antisense modulators of mRNA transcripts (7). Recently, lncRNAs have been implicated
in several types of cancer, including NSCLC. For example, lncRNA
prostate cancer associated transcript 6 acted as an oncogene in
NSCLC, contributing to tumor cell growth, migration and invasion
(8). Another previous study
suggested that small nucleolar RNA host gene 1 was upregulated in
NSCLC and promoted tumor cell viability, proliferation and
metastasis in NSCLC (9). In
addition, lncRNA VPS9D1-antisense RNA 1 (AS1) was overexpressed in
NSCLC, which was associated with high rates of metastasis and short
survival time within ~10 years (10). Altogether, these previous studies
suggested that lncRNAs played essential roles in the progression of
NSCLC. Tumor protein 73 AS1 (TP73-AS1) has been found to be
involved in several types of cancer (11,12).
However, the role of TP73-AS1 in the development of NSCLC remains
unclear.
MicroRNAs (miRNAs) are non-coding RNA molecules,
~18-22 nucleotides in length (13). MiRNAs interact with downstream
mRNAs, leading to mRNA degradation or translation repression.
Therefore, miRNAs modulate diverse cellular processes, including
cell differentiation, proliferation and development (14,15).
Aberrant increased or decreased expression of miRNAs has been
implicated in different human diseases, particularly cancer
(16). Previous studies
demonstrated that miR-34a-5p acted as a tumor suppressor in various
types of cancer, such as colorectal cancer (17), cervical cancer (18) and pancreatic carcinoma (19). However, the role of miR-34a-5p in
lung cancer is unknown.
Tripartite motif-containing 29 (TRIM29) is a member
of the TRIM protein family, which contains multiple zinc finger
motifs and a leucine zipper motif (20). The TRIM29 gene is located on
chromosome 11q23 and plays a role in numerous biological processes,
including DNA assembly, protein repair and cancer progression
(21,22). Notably, TRIM29 not only serves as a
tumor inhibitor, such as in invasive breast cancer (23), but also acts as an oncogene in
gastric cancer (24), thyroid
carcinoma (25) and bladder cancer
(26). Nevertheless, the potential
role played by TRIM29 in NSCLC is poorly understood.
The present study examined the expression pattern of
TP73-AS1 in NSCLC and investigated the role of TP73-AS1 in the
progression and drug chemoresistance of NSCLC. It also provided a
potential pathway to explain the mechanism of TP73-AS1 action in
NSCLC to further understand the pathogenic processes of NSCLC.
Materials and methods
Lung cancer specimens
A total of 50 lung cancer specimens and paired
normal tissues within 5 cm of the tumor were collected from
patients at The First People's Hospital of Tianmen. Patients'
sample collection and 5-year overall survival investigation were
conducted between January 2013 and November 2018. All specimens
were promptly placed in liquid nitrogen following surgery, then
stored at −80°C. All patients provided written informed consent
prior to surgery. The present study was approved by the Ethics
Committee of The First People's Hospital of Tianmen. Clinical
variables were obtained from the medical records of the patients,
including age, sex, tumor size, and prognosis, to determine the
association between TP73-AS1 expression and clinicopathological
characteristics (Table I). The
demarcation of high expression or low expression of TP73-AS1 was
performed according to the mean value of TP73-AS1 expression in all
cases. The clinical stages of the NSCLC tissue samples were
confirmed according to the tumor-node-metastasis (TNM)
classification of the 7th edition of the American Joint Committee
on Cancer/Union for International Cancer Control (27).
 | Table I.Association between TP73-AS1
expression level and clinicopathological characteristics of
patients with non-small cell lung cancer. |
Table I.
Association between TP73-AS1
expression level and clinicopathological characteristics of
patients with non-small cell lung cancer.
|
|
| TP73-AS1
expression |
|---|
|
|
|
|
|---|
|
Characteristics | No. of
patients | High, n | Low, n | P-value |
|---|
| Age, years |
|
|
| 0.156 |
|
<60 | 27 | 11 | 16 |
|
|
≥60 | 23 | 14 | 9 |
|
| Sex |
|
|
| 0.774 |
|
Male | 29 | 15 | 14 |
|
|
Female | 21 | 10 | 11 |
|
| Tumor size, cm |
|
|
| 0.009a |
|
<5 | 31 | 11 | 20 |
|
| ≥5 | 19 | 14 | 5 |
|
| Histology |
|
|
| 0.569 |
|
Squamous | 28 | 13 | 15 |
|
|
Adenoma | 22 | 12 | 10 |
|
|
Differentiation |
|
|
| 0.089 |
|
Well/moderately | 26 | 10 | 16 |
|
|
Poorly | 24 | 15 | 9 |
|
| Lymph node
metastasis |
|
|
| 0.007a |
| No | 33 | 12 | 21 |
|
|
Yes | 17 | 13 | 4 |
|
| TNM stage |
|
|
| 0.021a |
|
I+II | 20 | 6 | 14 |
|
|
III+IV | 30 | 19 | 11 |
|
Chromogenic in situ hybridization
(CISH) assay
CISH was performed using the ZytoDot CISH
Implementation kit with the commercially available digoxigenin
(DIG)-labeled ZytoDot CISH probes (ZytoVision GmbH). Tissue
preparation (3–5 µm microtome sections), pre-treatment,
hybridization and quenching were performed according to the
manufacturer's instructions prior to the detection of hybridized
DIG-labeled probes. NSCLC tissue microarray (TMA) manufacturing was
outsourced to Outdo Biotech, Co., Ltd. The TMA consisted of 50
pairs of NSCLC tissues and matched, adjacent, normal tissues
(within 5 cm of the tumor). Probes labeled with DIG result in
permanent diaminobenzidine (DAB) brown-colored, distinct,
dot-shaped signals, which are clearly distinguishable from the
background counterstained with hematoxylin.
Cell lines and culture
The HCC827, H522, and H23 NSCLC cell lines, and the
16HBE bronchial epithelial cell were purchased from BeNa Culture
Collection. HCC827, H522 and H23 were cultured in RPMI-1640
supplemented with 10% FBS (both from Gibco; Thermo Fisher
Scientific, Inc.) at 37°C containing 5% CO2. 16HBE cells
were cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.) and
10% FBS at 37°C containing 5% CO2.
Cell transfection
Small interfering (si)RNA against TP73-AS1
(si-TP73-AS1, #1: 5′-TAAGGTTATCCGAATAACGGTATCGTT-3′; #2:
5′-CCTGCTGCCTCTCCAAGAGACTGCTATTA-3′ and #3:
5′-GCAGTCGGGGCTGACGGCGG-3′), miR-34a-5p mimic [miR-34a-5p:
(5′-UGGCAGUGUCUUAGCUGGUUGU-3′)], miR-34a-5p inhibitor
(anti-miR-34a-5p: 5′-ACAACCAGCTAAGACACUTCCA-3′) and their negative
controls [si-NC (5′-CCTCCACGTCACGTATAGTGACATT-3′), miR-NC
(5′-UUCUCCGAACGUGUCACGUTT-3′) and anti-NC
(5′-UUCUCCGAACGUGUCACGUTT-3′), respectively] were synthesized by
Guangzhou RiboBio Co., Ltd. TP73-AS1 overexpression vector, TRIM29
overexpression (pcDNA-TRIM29) and pcDNA empty vector were obtained
from Sangon Biotech Co., Ltd. Transient transfection with 40 nM of
the indicated oligonucleotides or 1 µg plasmids was performed using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Subsequent experiments were conducted 48 h
following transfection.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and cDNA was
synthesized using the PrimeScript RT reagent kit (Takara
Biotechnology Co., Ltd.) for TP73-AS1 and TRIM29, or with the One
Step PrimeScript miRNA cDNA Synthesis kit (Takara Biotechnology
Co., Ltd.) for miR-34a-5p. RT-qPCR was then performed using the
SYBR Premix Ex Taq II kit (Takara Biotechnology Co., Ltd.) on an
ABI 7900 system (Applied Biosystems; Thermo Fisher Scientific,
Inc.), according to the manufacturer's instructions. Expression
levels were calculated using the 2−ΔΔCq method (28) and normalized to GAPDH or small
nuclear RNA U6. Primers sequences were as follows: TP73-AS1
forward, 5′-CCGGTTTTCCAGTTCTTGCA-3′ and reverse,
5′-GCCTCACAGGGAAACTTCATG-3′; TRIM29 forward,
5′-TTGCATGTTCCAGGAGCACAAGAAT-3′, and reverse,
5′-CAATGCACCAAATTCCTGCAGAAACA-3′; GAPDH forward,
5′-ACCACAGTCCATGCCATCAC-3′; and reverse,
5′-TCCACCACCCTGTTGCTGTA-3′. The primers for miR-34a-5p (forward,
5′-ACACTCCAGCTGGGTGGCAGTGTCTTAGC-3′; and reverse,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGACAACCA-3′) and U6 (forward,
5′-CTCGCTTCGGCAGCACA-3′; and reverse, 5′-AACGCTTCACGAATTTGCGT-3′)
were purchased from Guangzhou RiboBio Co., Ltd.
Cell proliferation and drug resistance
analysis
Cell proliferation and viability was performed using
a Cell Counting Kit-8 (CCK-8; Beyotime Institute of Biotechnology).
Transfected cells were seeded into 96-well plates (2,000
cells/well) and 10 µl CCK-8 solution was added into each well at
different time points post-transfection (0, 24, 48 and 72 h) for
another 2 h. The absorbance at 450 nm was measured using a
microplate reader (Bio-Rad Laboratories, Inc.). Cells were planted
into 96-well plates (10,000 cells/well) and then treated with
varying concentrations of cisplatin (DDP) (0, 1, 2, 4, 8, 16, 32,
64 and 128 µg/ml) for 48 h, and the concentration of DDP resulting
in 50% inhibition of growth (IC50) was obtained from the
dose-response curve.
Cell apoptosis analysis
Apoptosis was assessed using flow cytometry with a
Cell Apoptosis kit (Invitrogen; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. A total of
1×106 transfected HCC827 and H522 cells were washed with
cold PBS and re-suspended in 1X Annexin binding buffer. Next, 5 µl
fluorescein isothiocyanate (FITC)-conjugated Annexin V and 1 µl
propidium iodide at 100 µg/ml were added to 100 µl cell suspension
for 15 min at room temperature. Apoptotic cells were analyzed using
the S3™ Cell Sorter (Bio-Rad Laboratories, Inc.) and FlowJo
software v7.6 (FlowJo LLC).
Cell migration and invasion
analysis
Cell migration and invasion was investigated using
the Transwell assay, and 24-well chambers (Corning Inc.). For
migration, transfected HCC827 and H522 cells (10,000 cells/well) in
serum-free RPIM medium were placed into the top of chambers without
Matrigel™ (Corning Inc.). For invasion assays, chambers were
pre-coated with Matrigel™ at 4°C overnight, and then the
transfected cells (40,000 cell/well) were seeded into the upper
chambers coated with Matrigel™. In either assay, RPMI medium
containing 10% FBS was added to the bottom chambers. After a 24-h
incubation, the migrated or invaded cells on the bottom surface
were fixed with 90% methanol for 20 min and stained with crystal
violet (Beyotime Institute of Biotechnology) for 10 min at room
temperature. Finally, three randomly selected fields were chosen to
evaluate the number of cells under a light microscope
(magnification, ×100; Olympus Corporation).
Bioinformatics analysis
The microRNA.org
(version: 2010; http://www.microrna.org/microrna/getDownloads.do)
and Starbase (version: 3.0; http://starbase.sysu.edu.cn/agoClipRNA.php?source=circRNA)
online tools were used to predict the targets and binding sites
between lncRNA and miRNA, or miRNA and mRNA, respectively. Then,
the expression of these predicted miRNAs was detected in cells with
TP73-AS1 knockdown to monitor which miRNAs were upregulated.
Similarly, the expression of predicted mRNAs was detected in cells
with miR-34a-5p enrichment to monitor which mRNAs were
downregulated, using RT-qPCR assay as detailed above.
Dual-luciferase reporter assay
The wild-type (WT) and mutant (MUT) 3′untranslated
region (3′UTR) sequences of TP73-AS1 and TRIM29 containing putative
binding sites with miR-34a-5p (400 bp, containing 200 bp sequences
before or after binding sites) were amplified by PCR and introduced
into the luciferase reporter vector PGL3 (Promega Corporation) to
generate fusion plasmids, referred to as TP73-AS1-WT, TP73-AS1-MUT,
TRIM29-WT and TRIM29-MUT. These fusion plasmids were transfected
into HCC827 and H522 cells together with miR-34a-5p mimic or miR-NC
using Lipofectamine® 2000 reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). After a 48-h transfection, cells were
collected and luciferase activity was detected using the dual Glo™
Luciferase Assay System (Promega Corporation), according to the
manufacturer's protocol. Relative luciferase activity was expressed
as normalization of Renilla luciferase activity to firefly
luciferase activity.
Western blot analysis
Total protein was isolated using RIPA lysis buffer
(Beyotime Institute of Biotechnology) and quantified using BCA
Protein Assay kit (Beyotime Institute of Biotechnology). Equal
amounts (40 µg) of protein were electrophoresed using 12% SDS-PAGE,
then transferred onto PVDF membranes on ice (Bio-Rad Laboratories,
Inc.). Next, the membranes were blocked with 5% skimmed milk for 1
h at room temperature and incubated with primary antibodies
overnight at 4°C. The following day, the membranes were washed with
Tris-buffered saline + Tween-20 buffer, and incubated with a
secondary antibody for 2 h. The protein blots were then visualized
using an enhanced chemiluminescence kit (Beyotime Institute of
Biotechnology) on a ChemiDoc MP imaging system (Bio-Rad
Laboratories, Inc.). All antibodies were purchased from Abcam,
including primary antibodies against TRIM29 (cat. no. ab244380;
1:1,000) and GAPDH (cat. no. ab181602; 1:10,000) and the
HRP-conjugated secondary antibody (cat no. ab205718; 1:5,000). The
protein bands were quantified using the ImageJ software (version
1.46; National Institutes of Health).
Statistical analysis
All experiments were performed at least three times.
Differences between two groups were assessed using paired and
unpaired Student's t-test. ANOVA was used for multi-group
comparisons, followed by Tukey's post hoc test. Overall survival
was evaluated using the Kaplan-Meier method and analyzed by
log-rank test. The linear association between variables was
analyzed using Pearson correlation coefficient. Statistical
analysis was conducted using SPSS v17.0 software (SPSS, Inc.). Data
are presented as the mean ± SD. P<0.05 was considered to
indicate a statistically significant difference.
Results
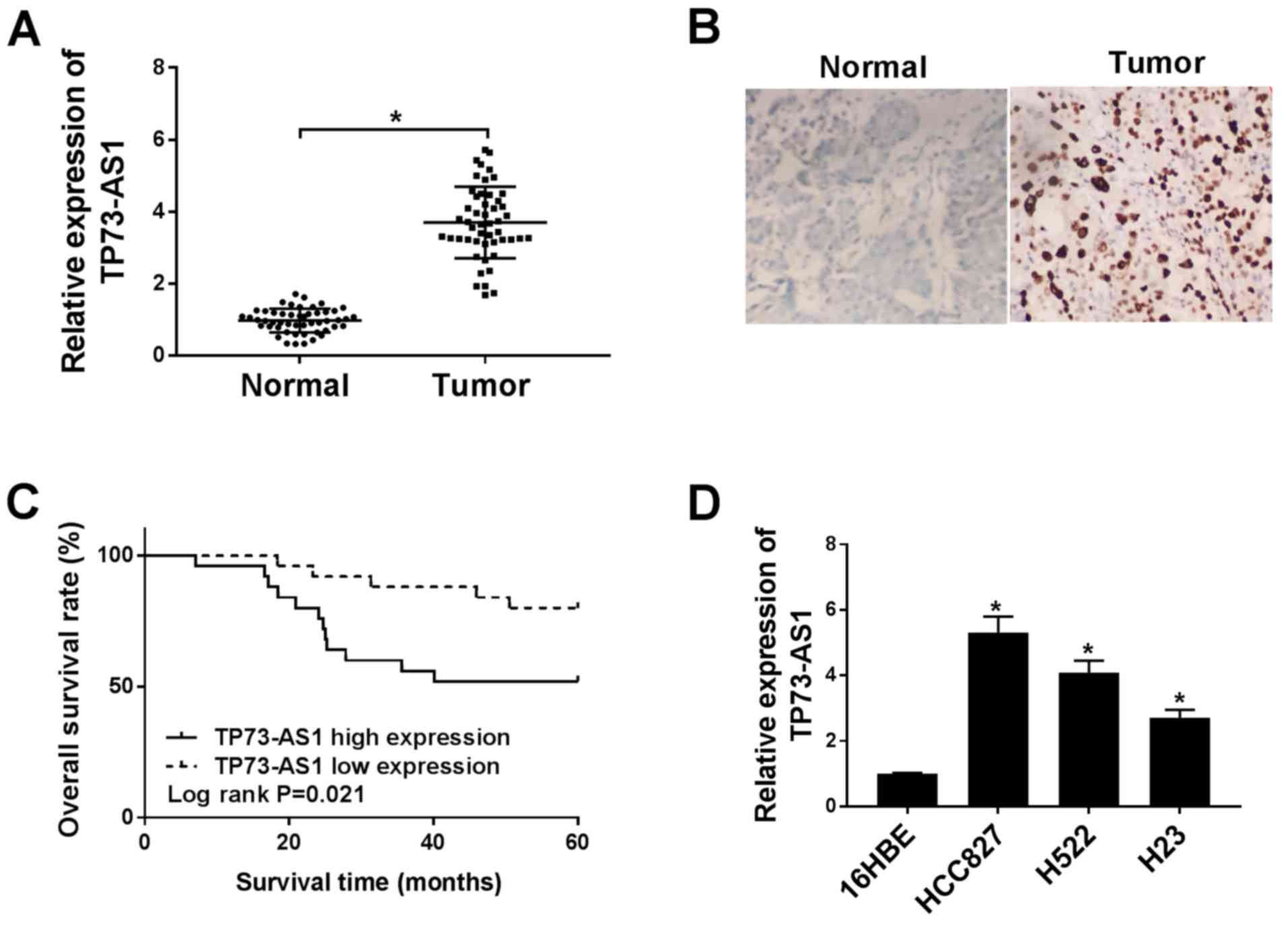
TP73-AS1 is upregulated in NSCLC
tissue and cell lines
Statistical analysis of clinicopathologic features
demonstrated that high TP73-AS1 expression was significantly
associated with large tumor size, positive lymph node metastasis
and advanced TNM stage (Table I).
To determine whether the expression of TP73-AS1 was altered in
NSCLC, RT-qPCR was performed in tumor samples and normal tissue
(n=50 in each group). Compared with that in normal tissue, TP73-AS1
was expressed in tumor tissues at significantly higher levels
(Fig. 1A). CISH was also used to
detect the expression of TP73-AS1 in NSCLC tumor tissues relative
to adjacent normal tissues, which revealed a high level of TP73-AS1
in NSCLC tissues (Fig. 1B).
Moreover, Kaplan-Meier curves and log-rank analysis indicated that
the overall survival rate of patients with NSCLC presenting with
high TP73-AS1 levels was lower, compared with patient with low
TP73-AS1 expression levels (Fig.
1C).
TP73-AS1 was also highly expressed in NSCLC cell
lines, including HCC827, H522, and H23, compared with that in the
16HBE bronchial epithelial cell line (Fig. 1D). In particular, expression of
TP73-AS1 was significantly higher in the HCC827 and H522 cell lines
relative to the H23 cell line. Therefore, the HCC827 and H522 cell
lines were selected for subsequent experiments. Altogether, these
findings indicated that increased TP73-AS1 expression levels could
play a role in NSCLC progression.
TP73-AS1 knockdown inhibits
proliferation, migration and invasion but induced apoptosis of
NSCLC cells, and enhances sensitivity to DDP
To determine the potential role of TP73-AS1 in NSCLC
cells, si-TP73-AS1 was transfected into HCC827 and H522 cells for
functional analysis. The mRNA expression level of TP73-AS1 was
reduced following transfection with si-TP73-AS1#1, si-TP73-AS1#2
and si-TP73-AS1#3 transfection, and the largest decrease was in the
si-TP73-AS1#1 group, in both cell lines (Fig. 2A and B). Therefore, si-TP73-AS1#1
was used in subsequent experiments.
 | Figure 2.TP73-AS1 knockdown inhibits
proliferation, migration and invasion, and promoted apoptosis and
drug sensitivity. The transfection efficiency of TP73-AS1 was
validated using reverse transcription-quantitative PCR in (A)
HCC827 and (B) H522 cells. *P<0.05 vs. si-NC. TP73-AS1 knockdown
suppressed cell proliferation in (C) HCC827 and (D) H522 cells. (E)
TP73-AS1 knockdown promoted cell apoptosis. (F and G) TP73-AS1
knockdown inhibited cell migration and invasion (magnification,
×100). Cell viability was detected in (H) HCC827 and (I) H522 cells
following si-TP73-AS1 transfection and DDP treatment. (J) TP73-AS1
knockdown reduced the IC50 following DDP treatment in
lung cancer cell lines. *P<0.05. TP73-AS1, tumor protein 73
antisense RNA 1; DDP, cisplatin; si, small interfering; OD, optical
density; NC, negative control; PI, propidium iodide; FITC,
fluorescein isothiocyanate. |
Cell proliferation was significantly decreased
following si-TP73-AS1 transfection (Fig. 2C and D). By contrast, apoptosis was
significantly increased in HCC827 and H522 cells transfected with
si-TP73-AS1 (Fig. 2E). In
addition, the numbers of migrating and invading cells were
significantly reduced in the NSCLC cell lines following
transfection with si-TP73-AS1 (Fig.
2F-G). In addition, the viability of HCC827 and H525 cells
transfected with si-TP73-AS1 was reduced with increasing DDP
concentrations, compared with that in the si-NC-transfected cells.
(Fig. 2H and I). Furthermore,
TP73-AS1 knockdown significantly reduced IC50 of DPP in
NSCLC cells (Fig. 2J). Taken
together, these results demonstrated that TP73-AS1 knockdown
inhibited the growth of NSCLC cells, and increased their
sensitivity to DDP.
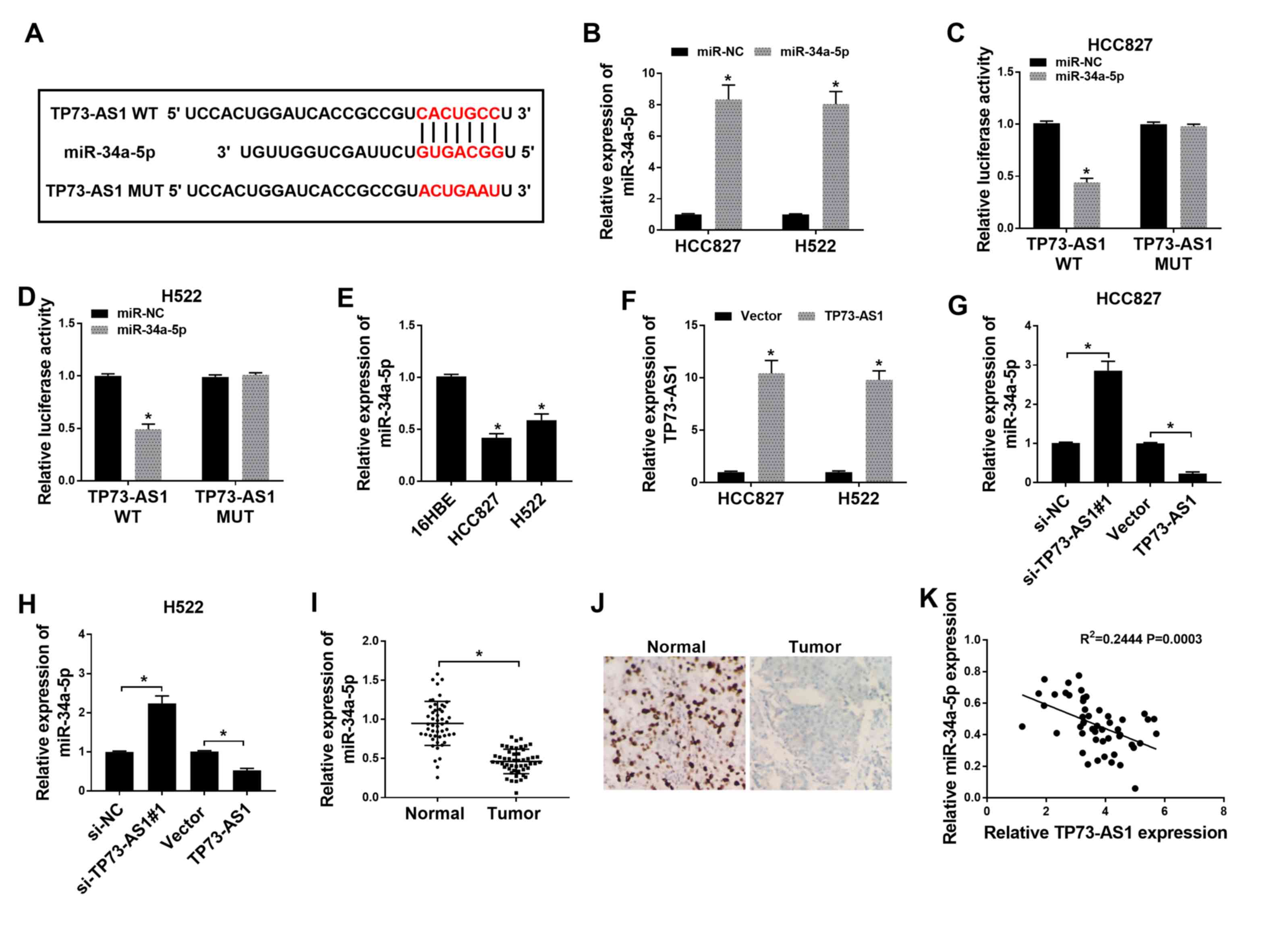
TP73-AS1 targets miR-34a-5p and
negatively regulates the expression of miR-34a-5p in NSCLC
cells
To explore the mechanism underlying the effect of
TP73-AS1 on NSCLC cell lines, the target miRNAs of TP73-AS1 were
predicted using microRNA.org and were subsequently
screened according to their expression levels. The sectional
predicted target miRNAs of TP73-AS1 are shown in Table SI. A significant increase in
miR-34a-3p expression was observed following TP73-AS1 knockdown
compared with that in cells transfected with si-NC, while there was
no significant difference in the other predicted target miRNAs
(Fig. S1). The binding sites
between TP73-AS1 and miR-34a-5p were identified using microRNA.org (Fig.
3A). The expression of miR-34a-5p was subsequently found to be
increased in HCC827 and H522 cells transfected with miR-34a-5p
mimics, compared with that in cells transfected with miR-NC
(Fig. 3B). In a dual-luciferase
reporter assay, luciferase activity decreased in HCC827 and H522
cells co-transfected with TP73-AS1-WT and miR-34a-5p. However,
there was no difference in luciferase activity in cells
co-transfected with TP73-AS1-MUT and miR-34a-5p (Fig. 3C and D).
The expression of miR-34a-5p in NSCLC cells was
measured, and was found to be significantly downregulated in HCC827
and H522 cells, compared with that in 16HBE cells (Fig. 3E). The efficiency of TP73-AS1
overexpression was subsequently verified. The expression of
TP73-AS1 was significantly increased in HCC827 and H522 cells
transfected with TP73-AS1, compared with that in cells transfected
with the empty vector (Fig. 3F).
Further analysis suggested that TP73-AS1 knockdown in HCC827 and
H522 cells increased miR-34a-5p expression, while TP73-AS1
overexpression reduced miR-34a-5p expression (Fig. 3G and H). Furthermore, the
expression of miR-34a-5p was significantly lower in NSCLC tumor
tissues compared with that in normal tissues (Fig. 3I), which was also confirmed using
CISH (Fig. 3J). In addition,
Pearson correlation coefficient analysis demonstrated that
miR-34a-5p expression was negatively associated with TP73-AS1
expression in NSCLC tissues (Fig.
3K). These results demonstrated that miR-34a-5p was targeted by
TP73-AS1.
miR-34a-5p inhibition reverses the
effects of TP73-AS1 knockdown on proliferation, apoptosis,
migration, invasion and drug resistance in NSCLC cells
To determine whether TP73-AS1 exerted its effects by
targeting miR-34a-5p, the HCC827 and H522 cell lines were
transfected with si-TP73-AS1 and anti-miR-34a-5p. Transfection with
anti-miR-34a-5p significantly reduced the expression of miR-34a-5p
in HCC827 and H522 cells, compared with that in cells transfected
with anti-NC (Fig. 4A). In a CCK-8
assay, cell proliferation was inhibited following si-TP73-AS1
transfection, compared with that in cells transfected with si-NC.
However, this effect was revered following transfection with
si-TP73-AS1+anti-miR-34a-5p, in both HCC827 and H522 cells
(Fig. 4B and C). Apoptosis was
significantly increased by TP73-AS1 knockdown but was significantly
decreased by anti-miR-34a-5p (Fig.
4D). The number of migrating and invading cells was inhibited
by TP73-AS1 knockdown, but rescued by miR-34a-5p inhibition
(Fig. 4E and F). Furthermore, the
IC50 of DDP was reduced following TP73-AS1 knockdown. By
contrast, anti-miR-34a-5p increased the IC50 value in
NSCLC cells (Fig. 4G and H). Taken
together, these findings suggest that TP73-AS1 knockdown blocked
proliferation, migration and invasion, accelerated apoptosis, and
reduced resistance to DDP, by upregulating miR-34a-5p in NSCLC
cells.
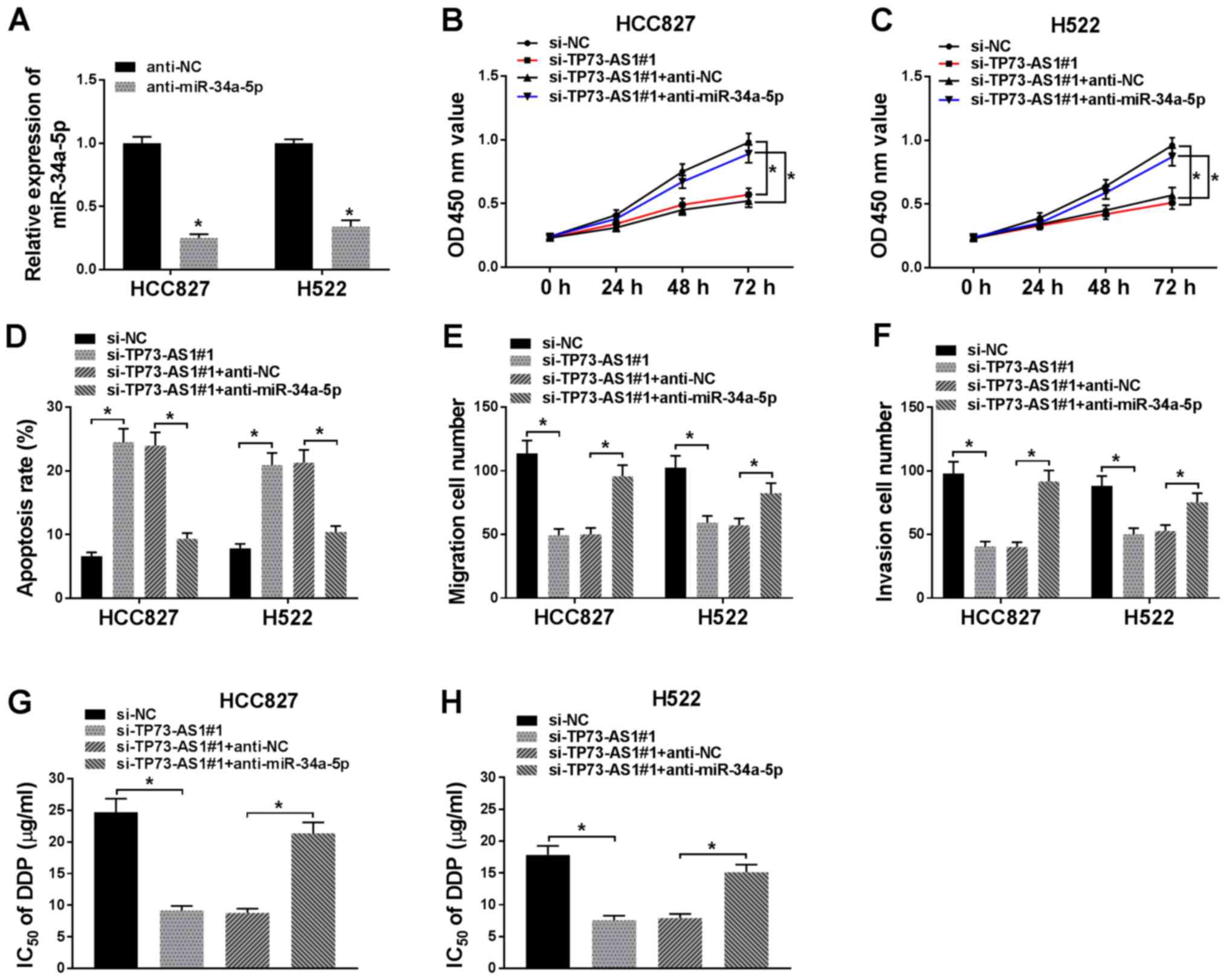 | Figure 4.Inhibition of miR-34a-5p reverses the
effect of TP73-AS1 knockdown in NSCLC cells. (A) Transfection of
anti-miR-34a-5p decreased miR-34a-5p expression in HCC827 and H522
cells. Transfection with anti-miR-34a-5p transfection reverses the
effect of si-TP73-AS1 on (B and C) proliferation, (D) apoptosis,
(E) migration, (F) invasion and (G and H) DDP resistance in NSCLC
cells. *P<0.05. TP73-AS1, tumor protein 73 antisense RNA 1;
NSCLC, non-small cell lung cancer; miR, microRNA; si, small
interfering; DDP, cisplatin; OD, optical density; NC, negative
control. |
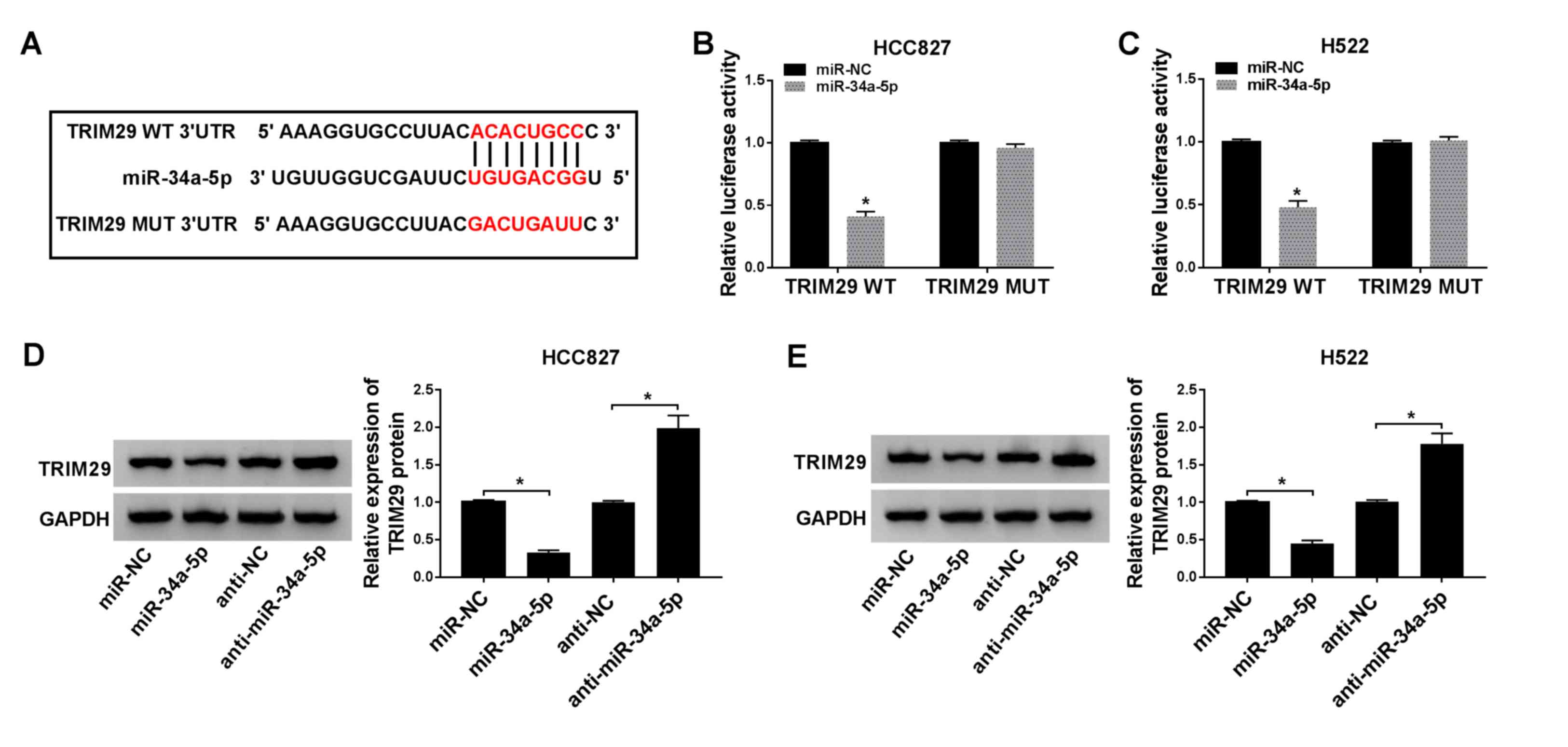
TRIM29 is a miR-34a-5p target in NSCLC
cells
To determine whether miR-34a-5p functioned by
regulating downstream genes, potential target mRNAs were obtained
using Starbase. The sectional predicted target mRNAs of miR-34a-5p
are shown in Table SII. A
significant decrease was found in TRIM29 expression in cells
following miR-34a-5p transfection compared with that in cells
transfected with miR-NC (Fig.
S2). A binding site was predicted between miR-34a-5p and TRIM29
3′UTR (Fig. 5A). In a
dual-luciferase reporter assay, luciferase activity was
significantly reduced in HCC827 and H522 cells co-transfected with
TRIM29-WT and miR-34a-5p. By contrast, TRIM29-MUT and miR-34a-5p
transfection did not affect luciferase activity compared with that
in the control group (Fig. 5B and
C). The expression of TRIM29 at the protein level was
suppressed following transfection with miR-34a-5p mimic, but
enhanced by anti-miR-34a-5p transfection, in both HCC827 and H522
cells (Fig. 5D and E). These
findings suggested that TRIM29 was a target for miR-34a-5p.
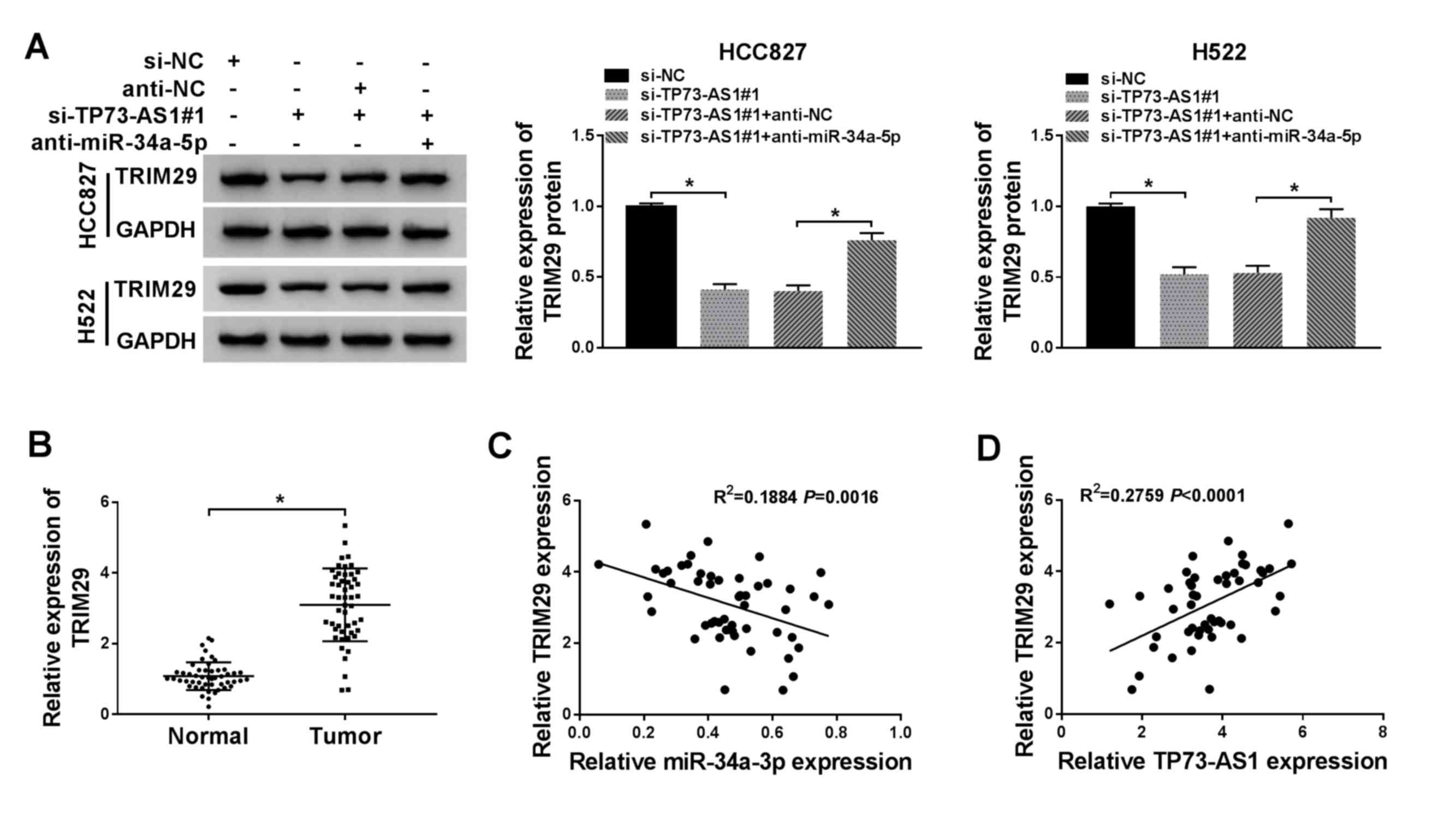
TP73-AS1 knockdown inhibits TRIM29
expression by increasing the expression of miR-34a-5p in NSCLC
cells
To determine whether TRIM29 expression was modulated
by TP73-AS1 and miR-34a-5p, the expression levels of TRIM29 in
HCC827 and H522 cells were investigated under different
transfection conditions. The protein expression level of TRIM29
decreased following si-TP73-AS1 transfection. However, TRIM29
expression was restored following si-TP73-AS1+anti-miR-34a-5p
co-transfection (Fig. 6A). In
addition, the expression of TRIM29 was significantly higher in
NSCLC tumor tissues compared with that in normal tissues (Fig. 6B). Pearson's correlation
coefficient analysis demonstrated that TRIM29 expression was
negatively associated with miR-34a-5p expression (Fig. 6C), but positively associated with
TP73-AS1 expression (Fig. 6D).
These results suggested that the expression of TRIM29 was regulated
by TP73-AS1 through miR-34a-5p.
TRIM29 overexpression reverses the
effects of miR-34a-5p enrichment on proliferation, apoptosis,
migration, invasion and drug resistance in NSCLC cells
To further investigate the mechanism underlying the
effects of TP73-AS1 on NSCLC cells, miR-34a-5p, miR-NC,
miR-34a-5p+pcDNA-TRIM29 or miR-34a-5p+pcDNA were introduced into
HCC827 and H522 cells. Transfection with pcDNA-TRIM29 led to a
significant increase in TRIM29 protein expression compared with
that in cells transfected with pcDNA (Fig. 7A). A CCK-8 assay demonstrated that
NSCLC cell proliferation was reduced by miR-34a-5p mimic, but
promoted following miR-34a-5p+pcDNA-TRIM29 transfection (Fig. 7B and C). Apoptosis was
significantly increased following miR-34a-5p mimic transfection.
However, the rates of apoptosis were reduced following
miR-34a-5p+pcDNA-TRIM29 co-transfection, in both HCC827 and H522
cells (Fig. 7D). The numbers of
migrating and invading cells were significantly reduced by
miR-34a-5p mimic transfection and significantly increased by TRIM29
overexpression (Fig. 7E and F).
Furthermore, the IC50 of DDP was significantly decreased
with miR-34a-5p mimic. By contrast, IC50 values were
significantly increased by TRIM29 overexpression (Fig. 7G and H). Taken together, this
indicated that miR-34a-5p overexpression inhibited NSCLC cell
growth and drug resistance by downregulating TRIM29.
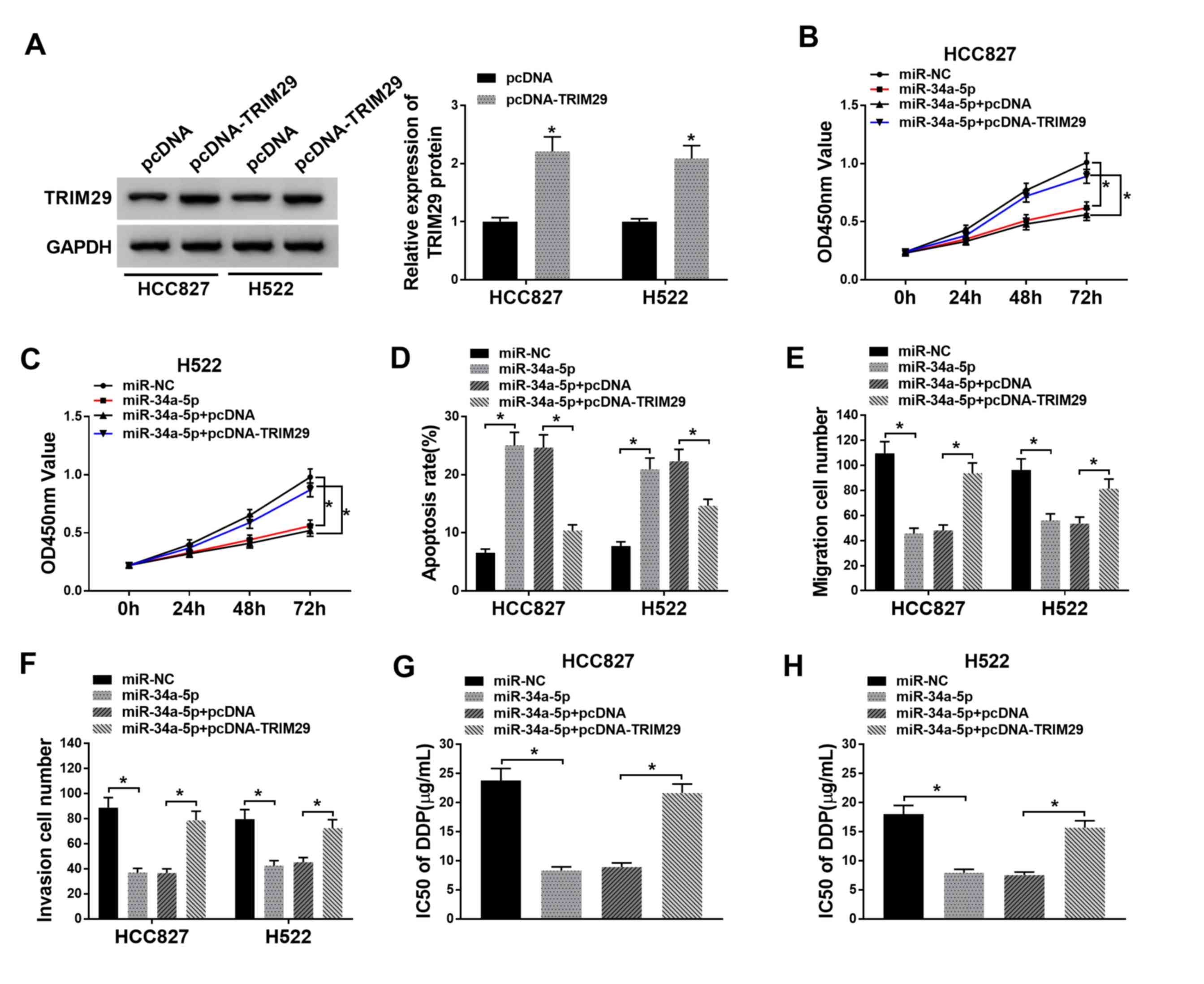 | Figure 7.Overexpression of TRIM29 reverses the
effects of miR-34a-5p mimics on proliferation, apoptosis,
migration, invasion and DPP resistance in NSCLC cells. (A)
Transfection with pcDNA-TRIM29 increased TRIM29 expression in
HCC827 and H522 cell, as indicated by western blot analysis. CCK-8
assay found that miR-34a-5p+pcDNA-TRIM29 co-transfection restored
cell proliferation vs. miR-34a-5p transfection alone in (B) HCC827
and (C) H522 cells. (D) TRIM29 overexpression inhibited apoptosis
in NSCLC cells. Cell (E) migration and (F) invasion was suppressed
following miR-34a-5p overexpression, but recovered after TRIM29
overexpression. DDP IC50 was reduced following miR-34a-5p
transfection, but increased after miR-34a-5p + pcDNA-TRIM29
transfection in (G) HCC827 and (H) H522 cells. *P<0.05. TRIM29,
tripartite motif-containing 29; NSCLC, non-small cell lung cancer;
miR, microRNA; NC, negative control; OD, optical density. |
Discussion
Substantial progress has been made into the
management and treatment of lung cancer; however, it remains the
leading cause of cancer-related mortality worldwide (29). The identification of additional
biomarkers are still required for the treatment of NSCLC. In the
present study, TP73-AS1 was highly expressed in NSCLC cells.
Functional analysis demonstrated that TP73-AS1 downregulation
inhibited NSCLC cell growth and resistance to DDP. An interaction
between miR-34a-5p, TP73-AS1 and TRIM29 was also identified. Thus,
TP73-AS1 promoted tumor cell growth, invasion and drug resistance
to DDP through the modulation of TRIM29 and miR-34a-5p in NSCLC
cells.
Numerous molecules and genes that are abnormally
expressed in tumor tissues significantly affect the occurrence and
development of tumors. Previous studies have found that TP73-AS1
was associated with the development of NSCLC. For example, Zhu
et al (30) found that
TP73-AS1 was expressed at high levels in NSCLC tumor tissues and
was associated with poor survival. In addition, increased TP73-AS1
endogenous levels promoted migration and invasion of NSCLC cells
(30). Zhang et al
(31) found that inhibition of
TP73-AS1 reduced NSCLC cell proliferation, inhibited cell cycle
progression in vitro, and blocked tumor growth in
vivo. Furthermore, a previous study demonstrated that TP73-AS1
promoted temozolomide resistance in glioblastoma multiform cancer
stem cells (32). However, whether
TP73-ASa could affect DDP resistance in NSCLC remains unclear. In
the present study, TP73-AS1 was also upregulated in NSCLC tissues
and cells. Moreover, TP73-AS1 knockdown suppressed cell growth,
migration and invasion, and inhibited drug resistance to DDP. These
findings demonstrated that TP73-AS1 functioned as a tumor promoter
in NSCLC. Reactive oxygen species (ROS) are a vital biological
product of regular cell metabolism (33). ROS play an important role in
numerous biological processes, such as cell survival,
differentiation and apoptosis (33). Notably, apoptosis induced by
intracellular production of ROS has been recognized as one of the
basic antitumor mechanisms of DDP (34,35).
Therefore, further investigations should focus on the association
between ROS production and TP73-AS1 dysregulation in NSCLC.
miR-34a-5p was identified as a target of TP73-AS1 in
the present study. A previous study demonstrated that miR-34a-5p
was involved in the inhibition of lung cancer progression and
metastasis induced by curcumin (36). Another study, based on miRNA
microarray and quantitative PCR, found that miR-34a-5p was
upregulated in NSCLC tumor tissues treated with luteolin, resulting
in the inhibition of tumorigenesis (37). Moreover, miR-34a was downregulated
in lung cancer cells, which enhanced their sensitivity to DDP
(38). miR-34a overexpression also
enhances DDP sensitivity in gastric cancer (39). In agreement with these previous
studies, miR-34a-5p was downregulated in NSCLC cells in the present
study. In addition, miR-34a-5p overexpression reduced
proliferation, migration, invasion and drug resistance to DDP,
whilst inducing apoptosis in NSCLC cells. These results provide
insight into the role of miR-34a-5p in the growth and DDP
resistance of NSCLC cells.
Further experiments suggested that TRIM29 was
targeted by miR-34a-5p. Previous studies have documented that
TRIM29 functions as a tumor suppressor in several types of cancer,
including breast cancer and hepatocellular carcinoma (23,40).
By contrast, TRIM29 appears to function as a tumor promoter in
lung-related cancers, and accelerates the progression of lung
cancer (41–43). Moreover, Liu et al (43) also found that TRIM29 downregulation
could augment DDP chemosensitivity in lung cancer cells. Similarly,
TRIM29 was upregulated in of NSCLC tissues and cell lines in the
present study. TRIM29 overexpression reversed the inhibitory effect
of miR-34a-5p on progression and DDP resistance in NSCLC. In
conclusion, TRIM29 enhanced the growth and DDP resistance to DDP of
NSCLC cells.
In summary, the expression of TP73-AS1 was increased
in NSCLC tissues and cells, leading to poor overall survival.
TP73-AS1 promoted NSCLC progression and resistance to DDP, which
was accomplished partially by modulating the
TP73-AS1/miR-34a-5p/TRIM29 axis. Thus, the present study suggested
that TP73-AS1 might represent a potential biomarker in the
treatment of NSCLC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MS and HC designed the study and performed the
experiments. WL and CC collated and analyzed the data. SL and HC
were involved in performing the experiments. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethical Review
Committee of The First People's Hospital of Tianmen. All patients
provided written informed consent prior to the start of the
study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Papaetis GS, Roussos C and Syrigos KN:
Targeted therapies for non-small cell lung cancer. Curr Pharm Des.
13:2810–2831. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fan WD, Zhang XQ, Guo HL, Zeng WW, Zhang
N, Wan QQ, Xie WY, Cao J and Xu CH: Bioinformatics analysis reveals
connection of squamous cell carcinoma and adenocarcinoma of the
lung. Asian Pac J Cancer Prev. 13:1477–1482. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Younes RN, Pereira JR, Fares AL and Gross
JL: Chemotherapy beyond first-line in stage IV metastatic non-small
cell lung cancer. Rev Assoc Med Bras (1992). 57:686–691. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gainor JF and Shaw AT: Novel targets in
non-small cell lung cancer: ROS1 and RET fusions. Oncologist.
18:865–875. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Spizzo R, Almeida MI, Colombatti A and
Calin GA: Long non-coding RNAs and cancer: A new frontier of
translational research? Oncogene. 31:4577–4587. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dimartino D, Colantoni A, Ballarino M,
Martone J, Mariani D, Danner J, Bruckmann A, Meister G, Morlando M
and Bozzoni I: The long non-coding RNA lnc-31 interacts with Rock1
mRNA and mediates its YB-1-dependent translation. Cell Rep.
23:733–740. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shi X, Liu Z, Liu Z, Feng X, Hua F, Hu X,
Wang B, Lu K and Nie F: Long noncoding RNA PCAT6 functions as an
oncogene by binding to EZH2 and suppressing LATS2 in non-small-cell
lung cancer. EBioMedicine. 37:177–187. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lu Q, Shan S, Li Y, Zhu D, Jin W and Ren
T: Long noncoding RNA SNHG1 promotes non-small cell lung cancer
progression by up-regulating MTDH via sponging miR-145-5p. FASEB J.
32:3957–3967. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tan J and Yang L: Long noncoding RNA
VPS9D1-AS1 overexpression predicts a poor prognosis in non-small
cell lung cancer. Biomed Pharmacother. 106:1600–1606. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Varon M, Levy T, Mazor G, Ben David H,
Marciano R, Krelin Y, Prasad M, Elkabets M, Pauck D, Ahmadov U, et
al: The long noncoding RNA TP73-AS1 promotes tumorigenicity of
medulloblastoma cells. Int J Cancer. 145:3402–3413. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu G, Zhao X, Zhou J, Cheng X, Ye Z and
Ji Z: lncRNA TP73-AS1 promotes cell proliferation and inhibits cell
apoptosis in clear cell renal cell carcinoma through repressing
KISS1 expression and inactivation of PI3K/Akt/mTOR signaling
pathway. Cell Physiol Biochem. 48:371–384. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shukla GC, Singh J and Barik S: MicroRNAs:
Processing, maturation, target recognition and regulatory
functions. Mol Cell Pharmacol. 3:83–92. 2011.PubMed/NCBI
|
|
16
|
Macfarlane LA and Murphy PR: MicroRNA:
Biogenesis, function and role in cancer. Curr Genomics. 11:537–561.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gao J, Li N, Dong Y, Li S, Xu L, Li X, Li
Y, Li Z, Ng SS, Sung JJ, et al: miR-34a-5p suppresses colorectal
cancer metastasis and predicts recurrence in patients with stage
II/III colorectal cancer. Oncogene. 34:4142–4152. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhong Z, Zhou F, Wang D, Wu M, Zhou W, Zou
Y, Li J, Wu L and Yin X: Expression of KLF9 in pancreatic cancer
and its effects on the invasion, migration, apoptosis, cell cycle
distribution, and proliferation of pancreatic cancer cell lines.
Oncol Rep. 40:3852–3860. 2018.PubMed/NCBI
|
|
19
|
Sun Z, Zhang B and Cui T: Long non-coding
RNA XIST exerts oncogenic functions in pancreatic cancer via
miR-34a-5p. Oncol Rep. 39:1591–1600. 2018.PubMed/NCBI
|
|
20
|
Kosaka Y, Inoue H, Ohmachi T, Yokoe T,
Matsumoto T, Mimori K, Tanaka F, Watanabe M and Mori M: Tripartite
motif-containing 29 (TRIM29) is a novel marker for lymph node
metastasis in gastric cancer. Ann Surg Oncol. 14:2543–2549. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Masuda Y, Takahashi H, Sato S,
Tomomori-Sato C, Saraf A, Washburn MP, Florens L, Conaway RC,
Conaway JW and Hatakeyama S: TRIM29 regulates the assembly of DNA
repair proteins into damaged chromatin. Nat Commun. 6:72992015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Napolitano LM and Meroni G: TRIM family:
Pleiotropy and diversification through homomultimer and
heteromultimer formation. IUBMB Life. 64:64–71. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ai L, Kim WJ, Alpay M, Tang M, Pardo CE,
Hatakeyama S, May WS, Kladde MP, Heldermon CD, Siegel EM and Brown
KD: TRIM29 suppresses TWIST1 and invasive breast cancer behavior.
Cancer Res. 74:4875–4887. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qiu F, Xiong JP, Deng J and Xiang XJ:
TRIM29 functions as an oncogene in gastric cancer and is regulated
by miR-185. Int J Clin Exp Pathol. 8:5053–5061. 2015.PubMed/NCBI
|
|
25
|
Xu J, Li Z, Su Q, Zhao J and Ma J: TRIM29
promotes progression of thyroid carcinoma via activating P13K/AKT
signaling pathway. Oncol Rep. 37:1555–1564. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tan ST, Liu SY and Wu B: TRIM29
overexpression promotes proliferation and survival of bladder
cancer cells through NF-κB signaling. Cancer Res Treat.
48:1302–1312. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Edge SB and Compton CC: The American joint
committee on cancer: The 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Balata H, Fong KM, Hendriks LE, Lam S,
Ostroff JS, Peled N, Wu N and Aggarwal C: Prevention and early
detection for NSCLC: Advances in thoracic oncology 2018. J Thorac
Oncol. 14:1513–1527. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhu D, Zhou J, Liu Y, Du L, Zheng Z and
Qian X: lncRNA TP73-AS1 is upregulated in non-small cell lung
cancer and predicts poor survival. Gene. 710:98–102. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang L, Fang F and He X: Long noncoding
RNA TP73-AS1 promotes non-small cell lung cancer progression by
competitively sponging miR-449a/EZH2. Biomed Pharmacother.
104:705–711. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mazor G, Levin L, Picard D, Ahmadov U,
Carén H, Borkhardt A, Reifenberger G, Leprivier G, Remke M and
Rotblat B: The lncRNA TP73-AS1 is linked to aggressiveness in
glioblastoma and promotes temozolomide resistance in glioblastoma
cancer stem cells. Cell Death Dis. 10:2462019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen B, Shen Z, Wu D, Xie X, Xu X, Lv L,
Dai H, Chen J and Gan X: Glutathione peroxidase 1 promotes NSCLC
resistance to cisplatin via ROS-induced activation of PI3K/AKT
pathway. Biomed Res Int. 2019:76405472019.PubMed/NCBI
|
|
34
|
Zhang Y, Zheng S, Zheng JS, Wong KH, Huang
Z, Ngai SM, Zheng W, Wong YS and Chen T: Synergistic induction of
apoptosis by methylseleninic acid and cisplatin, the role of
ROS-ERK/AKT-p53 pathway. Mol Pharm. 11:1282–1293. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tonino SH, van Laar J, van Oers MH, Wang
JY, Eldering E and Kater AP: ROS-mediated upregulation of Noxa
overcomes chemoresistance in chronic lymphocytic leukemia.
Oncogene. 30:701–713. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jiao DM, Yan L, Wang LS, Hu HZ, Tang XL,
Chen J, Wang J, Li Y and Chen QY: Exploration of inhibitory
mechanisms of curcumin in lung cancer metastasis using a
miRNA-transcription factor-target gene network. PLoS One.
12:e01724702017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jiang ZQ, Li MH, Qin YM, Jiang HY, Zhang X
and Wu MH: Luteolin inhibits tumorigenesis and induces apoptosis of
non-small cell lung cancer cells via regulation of MicroRNA-34a-5p.
Int J Mol Sci. 19:4472018. View Article : Google Scholar
|
|
38
|
Wang X, Dong K, Gao P, Long M, Lin F, Weng
Y, Ouyang Y, Ren J and Zhang H: MicroRNA-34a sensitizes lung cancer
cell lines to DDP treatment independent of p53 status. Cancer
Biother Radiopharm. 28:45–50. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang Z, Kong Y, Yang W, Ma F, Zhang Y, Ji
S, Ma EM, Liu H, Chen Y and Hua Y: Upregulation of microRNA-34a
enhances the DDP sensitivity of gastric cancer cells by modulating
proliferation and apoptosis via targeting MET. Oncol Rep.
36:2391–2397. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xu M, Hu J, Zhou B, Zhong Y, Lin N and Xu
R: TRIM29 prevents hepatocellular carcinoma progression by
inhibiting Wnt/β-catenin signaling pathway. Acta Biochim Biophys
Sin (Shanghai). 51:68–77. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Song X, Fu C, Yang X, Sun D, Zhang X and
Zhang J: Tripartite motif-containing 29 as a novel biomarker in
non-small cell lung cancer. Oncol Lett. 10:2283–2288. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhan W, Han T, Zhang C, Xie C, Gan M, Deng
K, Fu M and Wang JB: TRIM59 promotes the proliferation and
migration of non-small cell lung cancer cells by upregulating cell
cycle related proteins. PLoS One. 10:e01425962015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu C, Huang X, Hou S, Hu B and Li H:
Silencing of tripartite motif (TRIM) 29 inhibits proliferation and
invasion and increases chemosensitivity to cisplatin in human lung
squamous cancer NCI-H520 cells. Thorac Cancer. 6:31–37. 2015.
View Article : Google Scholar : PubMed/NCBI
|