Introduction
Osteosarcoma (OS), also known as osteogenic sarcoma,
is the most common primary malignant bone tumor with a high
potential for lung metastasis, and is commonly formed in the bones
of arms and legs of children and adolescents (1,2).
Recently, the continuous improvement of technologies employed for
auxiliary examination and treatment have increased the 5-year
survival rate of OS from 20 to 70% in the past 30 years, especially
the use of novel chemotherapy agents (3,4).
However, currently approved agents can cause notable side effects
(5,6). Therefore, it is urgent to develop a
novel agent with high efficiency and low toxicity in the treatment
of OS.
Piceatannol
(3,3′,4,5′-tetrahydroxy-trans-stilbene; Pice), a phenolic
compound and an analog of resveratrol, first isolated from the
seeds Euphorbia lagascae (7), and naturally occurs in various
sources, including red wine, grapes, sugar cane, peanuts and
rhubarb (8–10). Previous studies reported that Pice
possesses a variety of pharmacological properties involved in
anti-inflammatory (11,12) and immunosuppressive features, and
is a promising chemopreventive agent with anticancer activity
against different types of tumors (12). it is well documented that Pice
could suppress proliferation, migration, and metastasis in a
variety of cancers, including breast, prostate and bladder cancer
(13–15). However, the therapeutic effects and
molecular mechanism of Pice in OS cells remains unclear.
MicroRNAs (miRNAs/miRs) are a class of small
non-coding RNAs (21–23 nucleotides), which negatively regulate gene
expression at the post-transcription level through inhibiting
translation or inducing RNA degradation (16,17).
Many miRNAs have been identified in mammals, and some of them are
expressed in a tissue-specific and developmental stage specific
manner (18). It is well reported
that some miRNAs have been identified to function as oncogenes or
tumor suppressor genes in cancers, and could inhibit cell signaling
pathways to mediate various biological processes, such as cell
differentiation, apoptosis, proliferation and migration (19,20).
Increasing evidence revealed that natural-derived agents exert
anticancer properties in various cancers via mediating the
expression of miRNAs (21–23). A previous study demonstrated that
Pice induces apoptosis in colorectal cancer cell lines via
upregulation of miR-129 (24).
Based on this, we speculated that Pice may suppress the growth of
human OS cells through modulating miRNA expression.
In this study, we aimed to investigate the
therapeutic effects of Pice on OS cells and explored the underlying
molecular mechanism by examining Pice-induced changes of miRNA
expression profiles. Our findings suggested that Pice may exert
anticancer effects on OS cells through mediating the
miR-21/PTEN/AKT signaling pathway and act as a potential
chemopreventive agent for the treatment of OS.
Materials and methods
Cell culture and treatments
The human OS cell lines MG-63 and Saos-2 were
obtained from the American Type Culture Collection, and cultured in
RPMI-1640 medium (Invitrogen; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (FBS; Sigma-Aldrich; Merck
KGaA), 100 IU/ml penicillin, and 100 mg/ml streptomycin, at 37°C in
a humidified atmosphere containing 5% CO2. The cells
treated with different concentrations (0, 10, 25, 50, 100 and 200
µM) at 37°C of Pice (Sigma-Aldrich; Merck KGaA) for 24 h and then
the cells were collected for further measurements.
MTT assay
The antiproliferative effects of Pice against OS
cells was measured using an MTT assay. Briefly, the cells
(1×104) were seeded into 96-well plate overnight at
37°C, and then the cells were treated with or without 10–200 µM of
Pice. At 24 h following Pice treatment, the cells were washed with
PBS and recovered in fresh medium. After incubation for 48 h at
37°C, 20 µl of MTT solution (Sigma-Aldrich; Merck KGaA) was added
to each well, the concentration of MTT was 0.5 g/ml. The cells were
incubated for 4 h at 37°C subsequently. The absorbance of the
samples was read at 490 nm (Sunrise™; Tecan Group Ltd.). Each
experiment was performed in triplicate and the data were presented
as mean ± standard deviation.
Apoptosis analysis by flow
cytometry
MG-63 or Saos-2 cells (1×106) were
harvested after treatment with Pice, washed in ice-cold PBS, and
fixed in 70% ice-cold ethanol in PBS for 30 min at 4°C. Then, the
cells were harvested and were double stained with 5 µl Annexin
V-fluorescein isothiocyanate and 1 µl of propidium iodide for 30
min at 4°C. The stained cells were analyzed with EPICS XL-MCL
FACScan (Becton Dickinson). The MultiCycle Software for Windows
(version 3.11; Phoenix Flow Systems) was used to analyze the
experimental data.
miRNA microarray analysis
Total RNAs were isolated from OS cells treated with
Pice (100 µM) using TRIzol® (Thermo Fisher Scientific,
Inc.) according to manufacturer's instructions. The quantity and
integrity of RNA samples was evaluated using a 2100 Bioanalyzer
(Agilent Technologies, Inc.). The miRNA ULS™ Labeling Kit (Kreatech
Diagnostics; Leica Biosystems) was used to label the total RNA (2.5
µg) according to the manufacturer's protocols. Labeled miRNA
targets were hybridized with OneArray® Hybridization
System. Microarray images were obtained using the Axon GenePix
4000B microarray scanner (Axon Instruments). Then, the scanning
images imported into the GenePix Pro6.0 program (Axon Instruments)
for grid alignment and data extraction. The miRNAs with intensities
≥50 were used to calculate a normalization factor in all samples.
Normalization was performed using median normalization. The miRNA
expression profiles were determined using MEV software (version
4.6; TIGR, Microarray Software Suite 4).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
After treatment with Pice, total RNA was extracted
from cultured cells using TRIzol according to the manufacturer's
protocol. cDNA was synthesized using the High Capacity cDNA
Synthesis Kit (Applied Biosystems; Thermo Fisher Scientific, Inc.)
with miRNA-specific primers. The miR-21 primer was obtained from
Guangzhou Ribobio Co., Ltd. The miR-21 primer sequences were as
follows: stem-loop RT primer:
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTCAACA-3′; forward
5′-GCCCGCTAGCTTATCAGACTGATG-3′ and reverse 5′-GTGCAGGGTCCGAGGT-3′.
Expression of U6 RNA served to normalize the expression of miR-21.
The U6 primer sequences were as follows: forward
5′-CTCGCTTCGGCAGCACA-3′ and reverse 5′-AACGCTTCACGAATTTGCGT-3′.
qPCR was performed by an Applied Biosystems 7500 Real-Time PCR
machine with miRNA-specific primer by TaqMan Gene Expression Assay
(Applied Biosystems; Thermo Fisher Scientific, Inc.). Thermocycling
conditions were: 95°C for 5 min, followed by 40 cycles of 95°C for
30 sec and 55°C for 25 sec, and extension at 72°C for 10 sec. All
reactions were performed in triplicate. The miRNAs relative
expression was analyzed with the 2−ΔΔCq method (25).
Transfection
MG-63 or Saos-2 cells (5×103) cells were
seeded into each well of 6-well plate, cultured in the RPMI-1640
medium containing 10% FBS, and treated with or without Pice. Then,
the cells were transfected with miR-21 mimics
(5′-UAGCUUAUCAGACUGAUGUUGA-3), miR-21 inhibitor
(5′-UCAACAUCAGUCUGAUAAGCUA-3′) or mimics negative control (NC,
5′-UUCUCCGAACGUGUCACGUTT-3′) inhibitor negative control (NC,
5′-CAGUACUUUUGUGUAGUACAA-3′), synthesized by Guangzhou Ribobio Co.,
Ltd., at a final concentration of 50 µM using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. At 48 h
after transfection, the cells were harvested and proliferation,
apoptosis and caspase-3 activity were evaluated by MTT assay, flow
cytometry and colorimetric activity analysis, respectively. Target
gene analyses of miR-21. A bioinformatics method was used to
predict the potential target genes of miR-21. Web-based software of
TargetScan 7.0 (targetscan.org/) and miRanda (microrna.org/) databases were used to predict the
targets of miR-21.
Caspase-3 activity
Caspase-3 activity was determined using a
colorimetric activity assay kit according to manufacturer's
instructions (Bio Vison). After treatment with Pice (100 µM) for 24
h, OS cells were harvested by centrifugation at 1,000 × g for 10
min at 4°C and incubated in lysis buffer (cat. no. P0013K; Beyotime
Institute of Biotechnology) on ice for 15 min at RT. Then, the
lysate was centrifuged at 13,000 × g and 4°C for 15 min, the
protein concentration was measured using a BCA Protein Assay Kit
(Beyotime Institute of Biotechnology) according to the
manufacturer's instructions. The lysates (10 µl) were incubated
with 10 µl of 0.2 mM Ac-DEVD-pNA in 80 µl of reaction buffer at
37°C for 2 h. The samples were measured with a microplate reader
(Model 680, Bio-Rad Laboratories, Inc.) at an absorbance of 405
nm.
Luciferase reporter assay
The human phosphatase and tensin homolog (PTEN)
3′-untranslated region (3′-UTR) containing complementary sequences
for the seed sequence of miR-21 was amplified by PCR and cloned
into the firefly luciferase expressing vector pMIR-REPORT (Ambion;
Thermo Fisher Scientific, Inc.; wild-type pMIR-REPORT-PTEN-3′-UTR,
wt). A mutant of the 3′-UTR with a mutation (mut) of complementary
sequences for the seed sequence of miR-21
(pMIR-REPORT-PTEN-mut-3′-UTR) was performed using the QuikChange II
Site-Directed Mutagenesis Kit (Stratagene; Agilent Technologies,
Inc.). The MG-63 cells were seeded into 24-well plate and
transfected with wt or mut reporter vector, together with miR-21
mimics or miR-21 inhibitor using Lipofectamine® 2000.
The pRL-TK plasmid (Promega Corporation) was used as a normalizing
control. At 48 h after transfection, the luciferase activity was
determined with the Dual-Light luminescent reporter gene assay
system (Applied Biosystems; Thermo Fisher Scientific, Inc.), and
the relative firefly luciferase activity was normalized with
Renilla luciferase. Each determination was performed in
triplicate.
Western blot analysis
After 48 h following transfection, total protein
from cells was isolated using radioimmunoprecipitation assay buffer
with protease inhibitor Cocktail (Pierce; Thermo Fisher Scientific,
Inc.). The protein concentration was determined using a BCA protein
assay kit. Total proteins (20 µg) were separated via 10% SDS-PAGE
and then transferred onto to polyvinylidene difluoride membranes
(BD Pharmingen; BD Biosciences). The membranes were blocked with 5%
non-fat milk at 4°C overnight, and incubated with primary
antibodies against PTEN (cat. no. sc-7974, 1:1,000, Santa Cruz
Biotechnology, Inc.), phosphorylated (p)-AKT (cat. no. sc-7985-R,
1:1,000, Santa Cruz Biotechnology, Inc.) and AKT (cat. no. sc-5298,
1:1,000, Santa Cruz Biotechnology, Inc.) at 4°C overnight. β-actin
(cat. no. A-5441, 1:1,000, Sigma-Aldrich; Merck KGaA) served as an
internal control. Horseradish peroxidase-conjugated (cat. no.
sc-2031, 1:5,000, Santa Cruz Biotechnology, Inc.) antibodies were
used as the secondary antibodies, incubating with the secondary
antibody for 1 h at room temperature. The protein bands were
scanned on the using the ChemiDocXRS + Imaging System (Bio-Rad
Laboratories, Inc.). The band intensity was quantified using
Quantity One v4.6.2 software (Bio-Rad Laboratories, Inc.). All the
experiments were performed in triplicate.
Statistical analysis
The SPSS 14.0 software (SPSS, Inc.) was used to
analyze the data. Numerical data presented as the mean ± standard
deviation. The difference between two groups was analyzed with a
Student's t-test. Multiple groups were compared using one-way
analysis of variance with Tukey's post hoc tests. P<0.05 was
considered to indicate a statistically significant difference and
P<0.01 indicated a highly statistically significant
difference.
Results
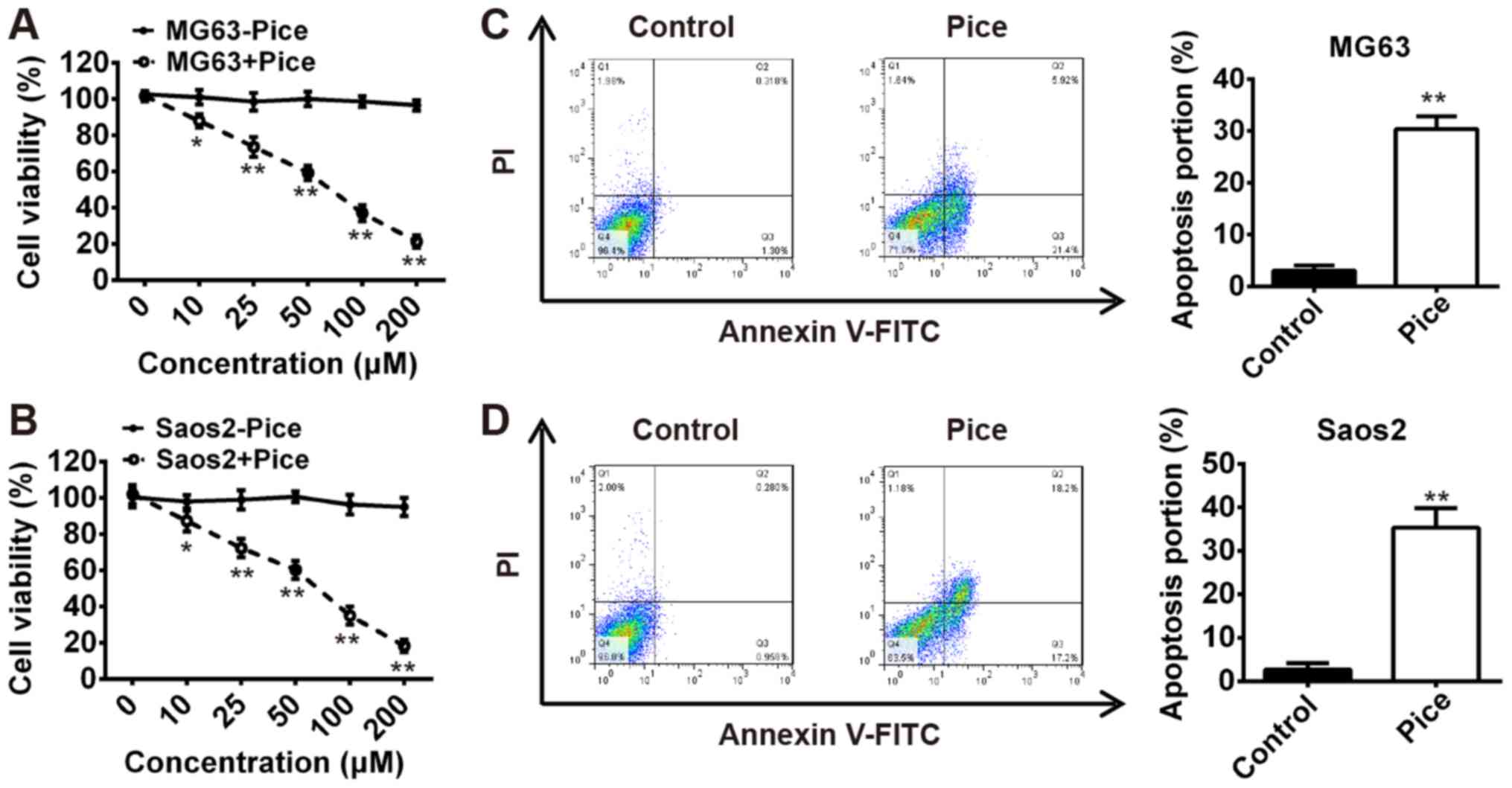
Pice suppresses OS cell growth
To examine the effects of Pice on the growth of OS
cells, MG-63 or Saos-2 cells were treated with Pice at different
concentrations (10–200 µM) for 24 h, and then the cell viability
was measured using an MTT assay. Our results demonstrated that
treatment with 10–200 µM Pice significantly inhibited Saos-2 and
MG-63 cell proliferation in a dose-dependent manner compared with
the control (P<0.01; Fig. 1A and
B). To explore whether the reduction in cell viability was
associated with cell apoptosis, we performed flow cytometric
analysis to examine apoptosis of MG-63 and Saos-2 cells treated
with Pice. As presented in Fig. 1C and
D, compared with the control group, the number apoptotic cells
was significantly increased for both cell lines after treatment
with 100 µM Pice for 24 h (P<0.01). These data indicated that
Pice may be a potential therapeutic agent in the treatment of
OS.
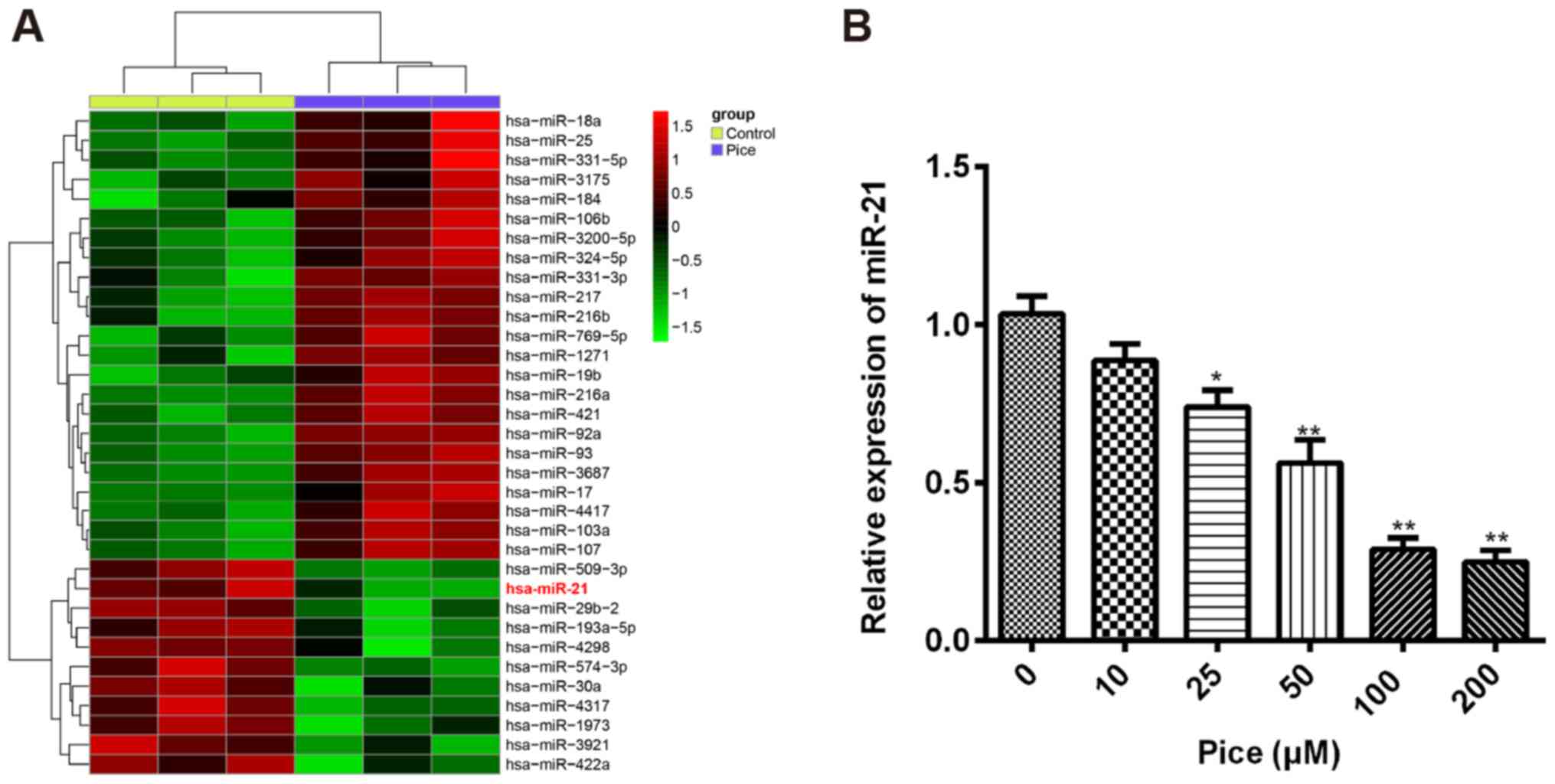
Pice induces the aberrant expression
of miRNAs in OS cells
Increasing evidence revealed that miRNAs are
associated a variety of biological and pathological processes,
including cellular differentiation, proliferation, apoptosis and
carcinogenesis (26,27). Recently, Pice has been identified
to exhibit anti-cancer effects on colorectal cancer cell lines
through upregulation of miR-129 (24). To investigate whether Pice
possessed the suppressive effect in OS cells via modulating miRNA
expression, microarray analysis was performed to measure the miRNA
expression profiles in OS cells after treatment with Pice (100 µM)
for 24 h. We observed that a large set of miRNAs expression were
altered after Pice treatment compared with the control, and that
miR-21 was the most significantly downregulated in human OS cells
after Pice treatment (Fig. 2A).
miR-21 has been reported to act as an oncogene in OS through
promoting proliferation and invasion, and suppressing apoptosis of
OS cells (28–30). Based on these studies, we further
measured the miR-21 levels in OS cells treated with Pice. The
Saos-2 cells were treated with 10–200 µM of Pice for 24 h and then
the expression of miR-21 was analyzed using RT-qPCR. The results
showed that Pice treatment reduced miR-21 expression in a
dose-dependent manner in OS cells (Fig. 2B). These results suggested that
Pice may exert anticancer effects through regulating miR-21
expression in human OS cells.
Overexpression of miR-21 attenuates
the therapeutic effects of Pice
To determine whether Pice exhibits the anticancer
effects on OS cells via modulating miR-21 expression, MG-63 and
Saos-2 cells were treated with or without 100 µM of Pice for 24 h
and transfected with miR-21 mimics or mimic NC, and cell viability
and apoptosis were measured by an MTT assay and flow cytometric
analysis, respectively. As shown in Fig. 3A and B, transfection with miR-21
mimic/inhibitor resulted in the upregulation and downregulation of
miR-21 in MG63 and Saos-2 cells, respectively, compared with the
NC-transfected cells (P<0.01). It was found that Pice treatment
significantly suppressed the viability of MG-63 and Saos-2 cells
compared with control, but overexpression of miR-21 significantly
increased cell viability compared with the Pice-treated group
(P<0.01; Fig. 3C and F).
Moreover, the Pice-induced apoptosis effect was significantly
impaired by overexpression of miR-21 in MG-63 and Saos-2 cells
compared with Pice-treated group (P<0.01; Fig. 3D and G). Additionally, it was
observed that Pice significantly increased caspase-3 activity
compared with control, but this enhancement was rescued by
overexpression of miR-21 in MG-63 and Saos-2 cells (P<0.01;
Fig. 3E and H). Taken together,
these data suggested that the therapeutic effects of Pice were
attenuated by overexpression of miR-21 in OS cells.
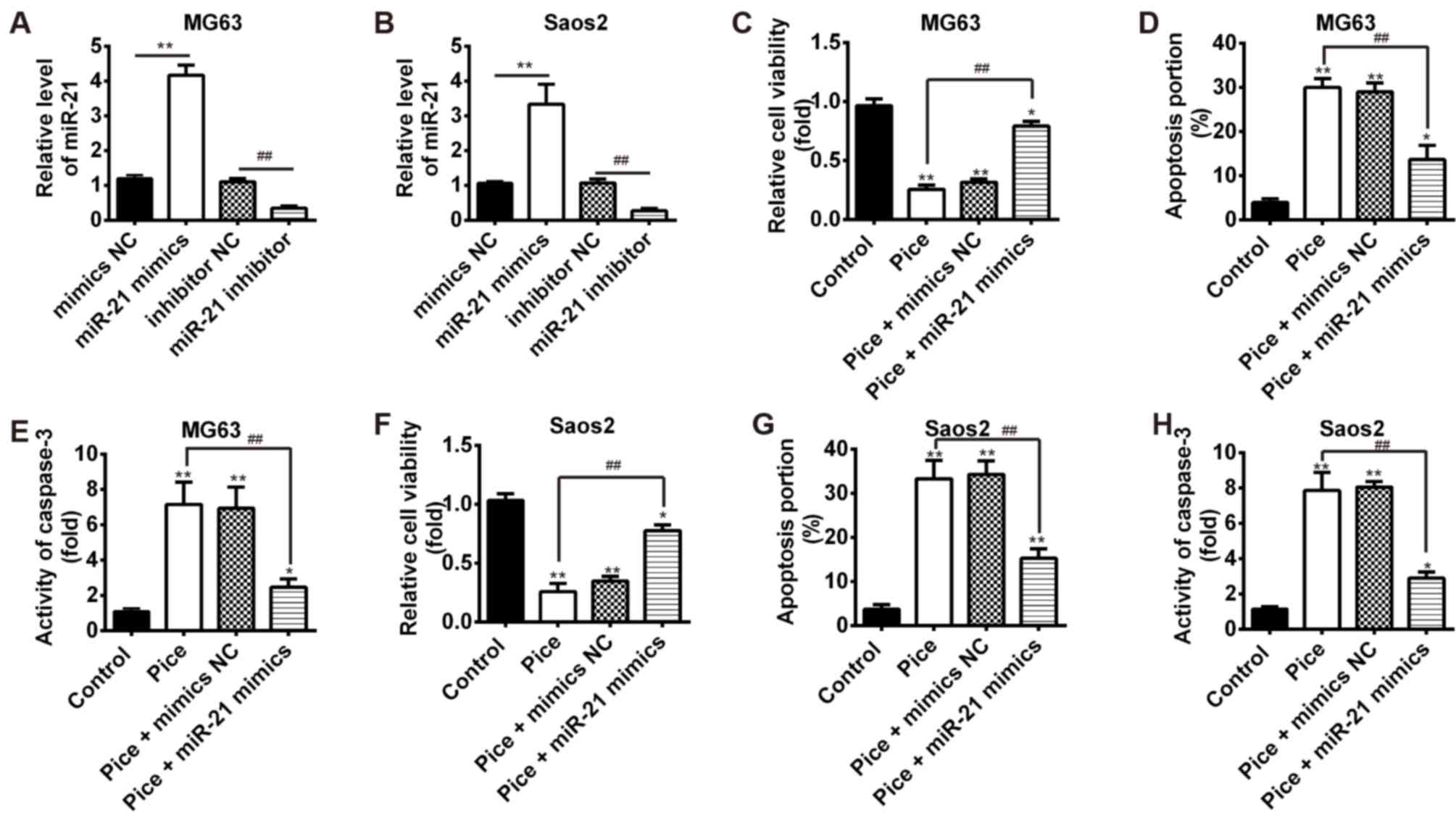 | Figure 3.Overexpression of miR-21 attenuates
the therapeutic effects of Pice. The MG-63 or Saos-2 cells were
transfected with miR-21 mimics or mimic NC after treatment with or
without 100 µM of Pice for 24 h. (A and B) MG-63 and Saos-2 cells
were transfected with miR-21 mimic/inhibitor or mimic/inhibitor NC.
The expression of miR-21 was measured using reverse
transcription-quantitative polymerase chain reaction analysis
(**P<0.01, ##P<0.01). (C-E) MTT assay,
flow cytometric analysis and colorimetric activity assay kit were
employed to measure cell viability, apoptosis and caspase-3
activity in MG-63 cells, respectively. (F-H) The cell viability,
apoptosis and caspase-3 activity were determined in Saos-2 cells
using an MTT assay, flow cytometric analysis and colorimetric
activity assay kit, respectively. Data were presented as the mean ±
standard deviation of three independent experiments. *P<0.05,
**P<0.01 vs. control. ##P<0.01. miR, microRNA; NC,
negative control; Pice, Piceatannol. |
PTEN is a target of miR-21 in OS
cells
It is reported that the protein PTEN has been
reported to act as a functional target of miR-21 in various human
cancer cells, including lung cancer, esophageal cancer and lung
squamous carcinoma (31–33). However, whether PTEN is a direct
target of miR-21 in OS cells remains to be elucidated. To identify
the target of miR-21 in OS cells, we performed TargetScan analysis
to predict the target genes of miR-21 and identified PTEN as a
potential target of miR-21 (Fig.
4A). To verify this bioinformatic predication, we established
the luciferase reporter plasmids containing the wt or mut 3′-UTR
segments of PTEN (Fig. 4A). A
luciferase reporter assay showed that miR-21 mimic significantly
inhibited the luciferase activity compared with the mimic NC, but
miR-21 inhibitor significantly enhanced the luciferase activity
compared with the inhibitor NC (P<0.01; Fig. 4B). Additionally, miR-21 mimic or
inhibitor did not affect the luciferase activity in the cells
transfected with pMIR-REPORT-PTEN-mut-3′-UTR (Fig. 4B). To further confirm that the PTEN
expression is regulated by miR-21, we performed the RT-qPCR and
western blotting to detect PTEN mRNA and protein levels,
respectively. The results showed that overexpression of miR-21
significantly inhibited the PTEN mRNA and protein levels compared
with NC, but knockdown of miR-21 significantly increased the PTEN
mRNA and protein levels (P<0.01; Fig. 4C and D). These results suggested
that miR-21 suppresses PTEN by targeting its 3′-UTR in OS
cells.
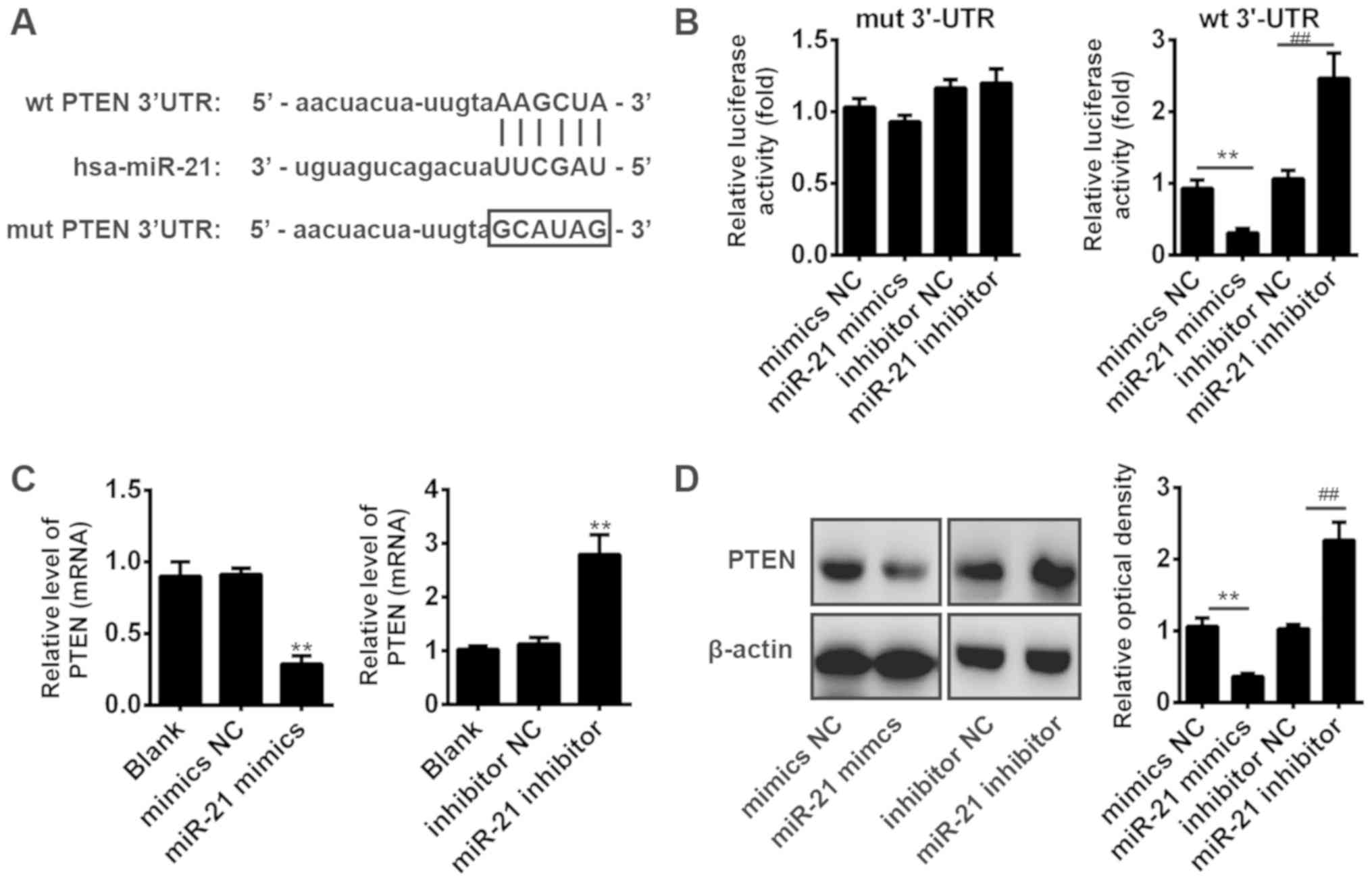 | Figure 4.PTEN is a direct target of miR-21 in
osteosarcoma cells. (A) The PTEN 3′-UTR region containing the wt or
mut binding site for miR-21. (B) The Saos-2 cells were
co-transfected with miR-21 mimics/inhibitor or NC oligos and
plasmid pMIR-REPORT-PTEN-3′-UTR (wt, mut). The relative firefly
luciferase activity normalized with Renilla luciferase was
measured 48 h after transfection. **P<0.01,
##P<0.01. (C) Saos-2 cells were transfected with
miR-21 mimic/inhibitor or corresponding mimic/inhibitor NC, and
reverse transcription-quantitative polymerase chain reaction was
used to measure PTEN mRNA level. **P<0.01 vs. NC. (D) Western
blot analysis was conducted to detect the PTEN protein level in
Saos-2 cells after transfection with miR-21 mimic/inhibitor or
corresponding mimic/inhibitor NC; β-actin was used as an internal
control. **P<0.01, ##P<0.01. Data were presented
as the mean ± standard deviation of three individual experiments.
Hsa, homo sapiens; miR, microRNA; mut, mutant; NC, negative
control; PTEN, phosphatase and tensin homolog; wt, wild-type;
3′-UTR, 3′-untranslated region. |
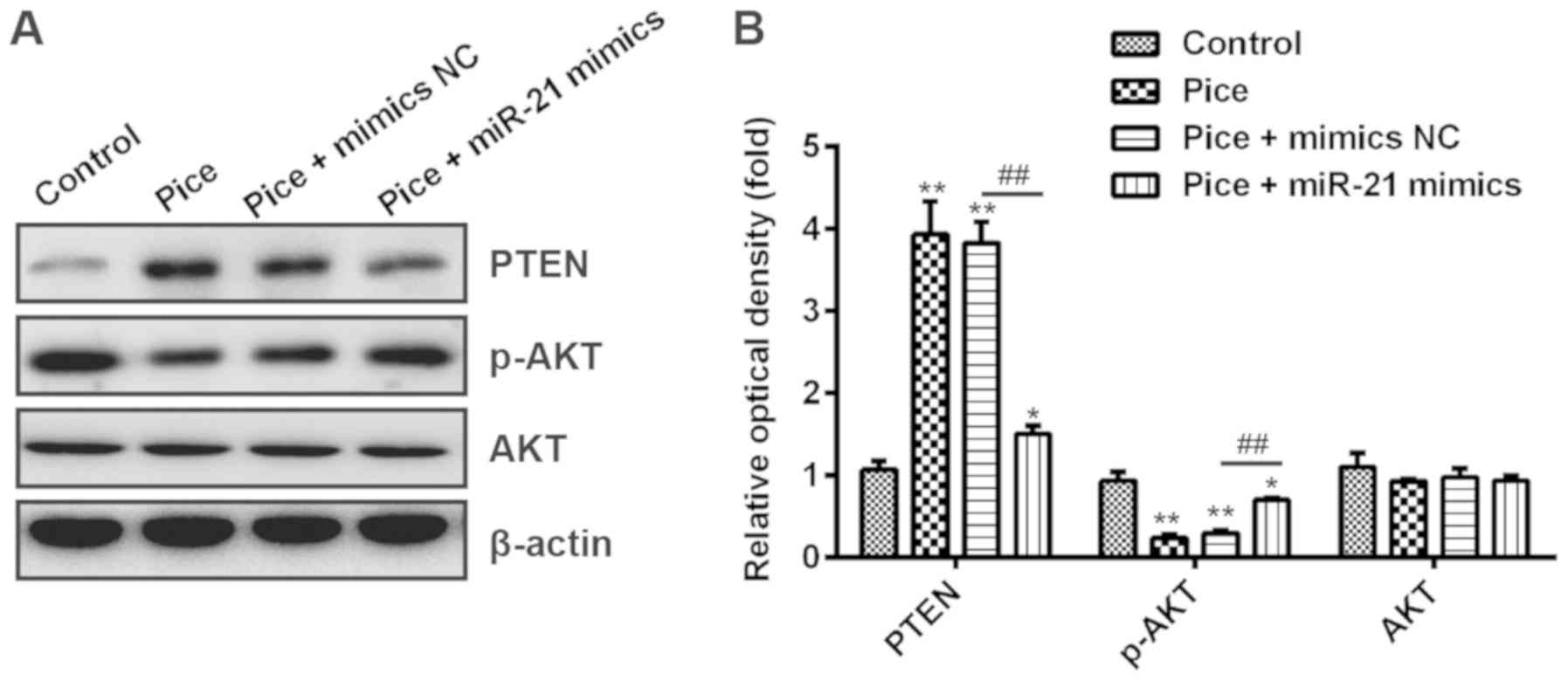
Pice blocks the PTEN/AKT signaling
pathway via modulating the expression of miR-21 in OS cells
It has been reported that the PI3K/AKT signaling
pathway plays a key role in cell survival, and possesses a
protective effect on tumorigenesis-associated apoptosis in cancer
cells (34). Moreover, AKT was
negatively regulated by PTEN, which serves a key role in a variety
of diseases through modulating cell proliferation, survival,
apoptosis and metabolism (35). A
recent study has demonstrated that miR-21 mediates the
proliferation, apoptosis, migration, invasion and the cell cycle of
human esophageal cancer cells, through targeting key proteins of
the PTEN/PI3K/AKT signaling pathway (36). Based on this background, we
hypothesized that Pice may also modulate the PTEN/AKT signaling
pathway via downregulation of miR-21 in OS cells. To investigate
this hypothesis, Saos-2 cells were transfected with or without
miR-21 mimics after treatment with or without Pice, and Western
blot analysis was used to measure the expression of PTEN and AKT.
Our results showed that Pice treatment resulted in PTEN
upregulation and p-AKT downregulation compared with the control,
but overexpression of miR-21 significantly reduced PTEN expression
and increased p-AKT expression in Pice-treated cells after
transfection with miR-21 mimics compared with transfection with
mimics NC (P<0.01; Fig. 5A and
B). Collectively, our data suggested that Pice blocks the
PTEN/AKT signaling pathway via inhibiting miR-21 expression in OS
cells.
Discussion
Accumulating evidence revealed that Pice has been
identified to act as anticancer agent in various cancers via
suppressing proliferation, migration, and metastasis (13–15).
However, whether Pice exerts such anti-cancer effects when used in
the context of the treatment of human OS remains unclear. In
present study, our results demonstrated that Pice suppresses
proliferation and in a dose-dependent manner prompts apoptosis in
OS cells. More importantly, we found that Pice alters miRNAs
expression in human OS cells and reduces miR-21 expression, which
was significantly downregulated in a dose-dependent manner.
Moreover, the therapeutic effects of Pice on OS cells were
attenuated by overexpression of miR-21. Additionally, we verified
that PTEN is a direct target of miR-21 and Pice blocks the PTEN/AKT
signaling pathway via suppressing miR-21 expression in OS cells.
These data suggested that Pice may exert therapeutic effects on OS
cells via modulating miR-21/PTEN/AKT signaling pathway and function
as a promising therapeutic agent in the treatment of OS.
Previous studies uncovered that resveratrol confers
an anticancer effect in different cancers through repressing cell
growth (37,38). Increasing evidence demonstrated
that Pice, a natural analog of resveratrol, induces apoptosis and
cell cycle arrest in human melanoma (39). Additionally, Pice has been reported
to act as an anti-tumor agent in leukemia cells (40,41).
In the present study, the results showed that Pice inhibits
proliferation and induces apoptosis in a dose-dependent manner in
both Saos-2 and MG-63 cells. These data suggested that Pice may be
a potential therapeutic agent in the treatment of OS.
Mounting evidence demonstrated that miRNAs
negatively regulate their target genes through inducing mRNA
cleavage, translational arrest and a combination of the two, mainly
by direct targeting of the 3′-UTRs of mRNAs (17,42,43).
It is extensively reported that miRNAs can function as tumor
suppressors or oncogenes by targeting genes involved in tumor cell
proliferation, apoptosis, differentiation and metastasis (44). One study demonstrated that
resveratrol and its analogs were used as attractive miRNA-mediated
chemopreventive and therapeutic strategy in prostate cancer
(45). Ke et al (46) revealed that resveratrol and Pice
reduce the expression of miR-183, resulting in attenuated
osteoclastogenesis. In addition, Pice has been identified to
suppress colorectal cancer growth via upregulation of miR-129.
Therefore, to investigate whether Pice exerts anticancer effects on
OS cells via mediating miRNAs, we used microarray analysis to
determine the miRNA expression profiles of OS cells after treatment
with Pice. We found that Pice induces the aberrant expression of
miRNAs and miR-21 was the most significantly downregulated in OS
cells. Moreover, Pice decreases miR-21 in a dose-dependent manner
as verified by RT-qPCR. miR-21 has been identified to act as an
oncogene in OS via promoting OS cell proliferation and invasion,
and suppressing apoptosis (28–30).
Then, we further investigated whether the therapeutic effects of
Pice were modulated by miR-21 expression; we observed that
overexpression of miR-21 attenuates the therapeutic effects of Pice
via promoting cell growth and inhibiting caspase-3 activity. Taken
together, these results suggested that Pice harbored the anticancer
effects on OS cells may suppress miR-21 expression. However, the
potential molecular mechanism require further investigation.
Previously, many studies demonstrated that numerous
miRNAs are upregulated or downregulated in OS, and are often
associated with the entire process of tumor development (44,47).
The relationship between miRNA expression and the prognosis of
patients with OS has been extensively reported. Cheng et al
(48) conducted a meta-analysis of
the prognostic significance miRNAs in OS, and indicated that a
number of other miRNAs are either upregulated (miR-214, miR-29, and
miR-148a) or downregulated (miR-382, miR-26a, miR-195, and miR-124)
in OS. However, these miRNAs (miR-214, miR-29, miR-148a, miR-382,
miR-26a, miR-195, and miR-124) were not detected in our microarray
analysis; whether these miRNAs are involved in the anticancer
effects of Pice on OS is yet to be determined.
Several studies have confirmed that miR-21 inhibits
PTEN expression by directly targeting its 3′-UTR in a variety of
cancer cells (31–33). Consistent with previous reports,
our data showed that PTEN is a functional target of miR-21 in OS
cells. PTEN, a tumor suppressor gene, plays an important role in
many types of solid tumors through modulating cell apoptosis and
the cell cycle (33), and its
tumor suppressor activity is dependent on its lipid phosphatase
activity, which negatively modulates the PI3K/AKT/mTOR pathway
(49). Therefore, we speculated
that Pice may regulate the PTEN/AKT signaling pathway via
inhibiting miR-21 expression. Our results demonstrated that Pice
treatment resulted in PTEN upregulation and p-AKT downregulation,
but overexpression of miR-21 significantly decreased PTEN
expression and increased p-AKT expression in Pice-treated OS cells.
These data indicated that Pice could modulate the PTEN/AKT
signaling pathway via suppressing miR-21 expression, suggesting
Pice may exert anticancer effects on OS cells via modulating
miR-21/PTEN/AKT signaling pathway.
Various natural products have been reported to
prevent or treat tumors, due to their effects on cellular defenses
or by targeting the key transcription factors, such as nuclear
factor-κB, signal transducers, activator protein and activators of
transcription and others (50–53).
The differential effects of natural products from plants in tumor
cells may be due to different abilities to induce specific
apoptotic pathways, modify the levels of major metabolic enzymes,
or induce detoxifying enzymes and tumor suppressor genes (54–56).
Resveratrol, a polyphenol, which has been found in various plants,
including grapes, passion fruit, white tea, and Japanese knotweed,
displays a wide spectrum of biological activity (50). Previously, polyphenolic compounds
were reported to exhibit anti-cancer effects in cancers, including
green tea polyphenol, honokiol (HNK), and Pice (50). HNK is a small organic molecule
purified from magnolia species and has demonstrated antitumor
activities in a variety of tumor cell lines (57). HNK inhibited the growth and
proliferation of oral squamous cell carcinoma cells in vitro
(58). Recently, Yang et al
(59) reported that HNK inhibits
proliferation and induces apoptosis through modulating the
miR-21/PTEN/PI3K/AKT signaling pathway in human OS cells. In the
present study, our results revealed that Pice exerts anticancer
effects on OS cells via regulating miR-21/PTEN/AKT signaling
pathway. These data indicated that plant-derived polyphenolic
compounds may exert anticancer effects in various cancers by
mediating miRNA expression. In subsequent experiments, we will
further verify whether the anticancer effects of plant-derived
polyphenolic compounds are associated with miRNA-mediated signaling
pathways.
In conclusion, our results revealed that Pice
suppressed cell proliferation in a dose-dependent manner and
induces the apoptosis of OS cells. Meanwhile, we verified that Pice
induces the aberrant expression of miRNAs in human OS cells, and
miR-21 was the most significantly downregulated. Most importantly,
the therapeutic effects of Pice on OS cells were weakened by
overexpression of miR-21 via blocking the PTEN/AKT signaling
pathway. Taken together, our findings indicated that the molecular
mechanism underlying the observed Pice-induced apoptosis could be
regulated by a miR-21/PTEN/AKT axis in human OS cells.
Acknowledgements
Not applicable.
Funding
This study was supported by the Scientific research
project of Shanghai science and technology commission (grant no.
18401902400) and the three-year action plan for further
accelerating the development of TCM in Shanghai (grant no.
ZY(2018–2020)-FWTX-4020).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
MZ and YW performed the experiments, contributed to
data analysis and wrote the paper. MZ and YW analyzed the data. YW
made substantial contributions to the concept of the study,
contributed to data analysis and acquired experimental materials.
Both authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ottaviani G and Jaffe N: The Epidemiology
of Osteosarcoma. Pediatric and Adolescent Osteosarcoma. Jaffe N,
Bruland OS and Bielack S: Springer US; Boston, MA: pp. 3–13.
2010
|
|
2
|
Wang H, Tang M, Ou L, Hou M, Feng T, Huang
YE, Jin Y, Zhang H and Zuo G: Biological analysis of cancer
specific microRNAs on function modeling in osteosarcoma. Sci Rep.
7:53822017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bielack SS, Marina N, Ferrari S, Helman
LJ, Smeland S, Whelan JS and Reaman GH: Osteosarcoma: The same old
drugs or more? J Clin Oncol. 26:3102–3103; author reply 3104–3105.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Epis MR, Giles KM, Beveridge DJ,
Richardson KL, Candy PA, Stuart LM, Bentel J, Cohen RJ and Leedman
PJ: miR-331-3p and Aurora Kinase inhibitor II co-treatment
suppresses prostate cancer tumorigenesis and progression.
Oncotarget. 8:55116–55134. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Longhi A, Errani C, De Paolis M, Mercuri M
and Bacci G: Primary bone osteosarcoma in the pediatric age: State
of the art. Cancer Treat Rev. 32:423–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Marina N, Gebhardt M, Teot L and Gorlick
R: Biology and therapeutic advances for pediatric osteosarcoma.
Oncologist. 9:422–441. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li Z, Yang X, Dong S and Li X: DNA
breakage induced by piceatannol and copper(II): Mechanism and
anticancer properties. Oncol Lett. 3:1087–1094. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cantos E, Espín JC, Fernández MJ, Oliva J
and Tomás-Barberán FA: Postharvest UV-C-irradiated grapes as a
potential source for producing stilbene-enriched red wines. J Agric
Food Chem. 51:1208–1214. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Matsuda H, Tomohiro N, Hiraba K, Harima S,
Ko S, Matsuo K, Yoshikawa M and Kubo M: Study on anti-Oketsu
activity of rhubarb II. Anti-allergic effects of stilbene
components from Rhei undulati Rhizoma (dried rhizome of Rheum
undulatum cultivated in Korea). Biol Pharm Bull. 24:264–267. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rimando AM, Kalt W, Magee JB, Dewey J and
Ballington JR: Resveratrol, pterostilbene, and piceatannol in
vaccinium berries. J Agric Food Chem. 52:4713–4719. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim YH, Kwon H-S, Kim DH, Cho HJ, Lee HS,
Jun JG, Park JH and Kim JK: Piceatannol, a stilbene present in
grapes, attenuates dextran sulfate sodium-induced colitis. Int
Immunopharmacol. 8:1695–1702. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jin CY, Moon DO, Lee KJ, Kim MO, Lee JD,
Choi YH, Park YM and Kim GY: Piceatannol attenuates
lipopolysaccharide-induced NF-kappaB activation and
NF-kappaB-related proinflammatory mediators in BV2 microglia.
Pharmacol Res. 54:461–467. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Song NR, Hwang MK, Heo YS, Lee KW and Lee
HJ: Piceatannol suppresses the metastatic potential of MCF10A human
breast epithelial cells harboring mutated H-ras by inhibiting MMP-2
expression. Int J Mol Med. 32:775–784. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jayasooriya RGPT, Lee YG, Kang CH, Lee KT,
Choi YH, Park SY, Hwang JK and Kim GY: Piceatannol inhibits
MMP-9-dependent invasion of tumor necrosis factor-α-stimulated
DU145 cells by suppressing the Akt-mediated nuclear factor-κB
pathway. Oncol Lett. 5:341–347. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kuo PL and Hsu YL: The grape and wine
constituent piceatannol inhibits proliferation of human bladder
cancer cells via blocking cell cycle progression and inducing
Fas/membrane bound Fas ligand-mediated apoptotic pathway. Mol Nutr
Food Res. 52:408–418. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao JJ, Yang J, Lin J, Yao N, Zhu Y,
Zheng J, Xu J, Cheng JQ, Lin JY and Ma X: Identification of miRNAs
associated with tumorigenesis of retinoblastoma by miRNA microarray
analysis. Childs Nerv Syst. 25:13–20. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhuang LK, Xu GP, Pan XR, Lou YJ, Zou QP,
Xia D, Yan WW, Zhang YT, Jia PM and Tong JH: MicroRNA-181a-mediated
downregulation of AC9 protein decreases intracellular cAMP level
and inhibits ATRA-induced APL cell differentiation. Cell Death Dis.
5:e11612014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang B, Pan X, Cobb GP and Anderson TA:
MicroRNAs as oncogenes and tumor suppressors. Dev Biol. 302:1–12.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zamani M, Sadeghizadeh M, Behmanesh M and
Najafi F: Dendrosomal curcumin increases expression of the long
non-coding RNA gene MEG3 via up-regulation of epi-miRs in
hepatocellular cancer. Phytomedicine. 22:961–967. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu S, Fang Y, Shen H, Xu W and Li H:
Berberine sensitizes ovarian cancer cells to cisplatin through
miR-21/PDCD4 axis. Acta Biochim Biophys Sin (Shanghai). 45:756–762.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hong M, Wang N, Tan YH, Tsao S-W and Feng
Y: MicroRNAs and Chinese Medicinal Herbs: New Possibilities in
Cancer Therapy. Cancers (Basel). 7:1643–1657. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang H, Jia R, Wang C, Hu T and Wang F:
Piceatannol promotes apoptosis via up-regulation of microRNA-129
expression in colorectal cancer cell lines. Biochem Biophys Res
Commun. 452:775–781. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−ΔΔC(T)) Method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
le Sage C and Agami R: Immense promises
for tiny molecules: Uncovering miRNA functions. Cell Cycle.
5:1415–1421. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ren X, Shen Y, Zheng S, Liu J and Jiang X:
miR-21 predicts poor prognosis in patients with osteosarcoma. Br J
Biomed Sci. 73:158–162. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lv C, Hao Y and Tu G: MicroRNA-21 promotes
proliferation, invasion and suppresses apoptosis in human
osteosarcoma line MG63 through PTEN/Akt pathway. Tumour Biol.
37:9333–9342. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ziyan W, Shuhua Y, Xiufang W and Xiaoyun
L: MicroRNA-21 is involved in osteosarcoma cell invasion and
migration. Med Oncol. 28:1469–1474. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu ZL, Wang H, Liu J and Wang ZX:
MicroRNA-21 (miR-21) expression promotes growth, metastasis, and
chemo- or radioresistance in non-small cell lung cancer cells by
targeting PTEN. Mol Cell Biochem. 372:35–45. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhuang LK, Yang YT, Ma X, Han B, Wang ZS,
Zhao QY, Wu LQ and Qu ZQ: MicroRNA-92b promotes hepatocellular
carcinoma progression by targeting Smad7 and is mediated by long
non-coding RNA XIST. Cell Death Dis. 7:e22032016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu LF, Wu ZP, Chen Y, Zhu QS, Hamidi S and
Navab R: MicroRNA-21 (miR-21) regulates cellular proliferation,
invasion, migration, and apoptosis by targeting PTEN, RECK and
Bcl-2 in lung squamous carcinoma, Gejiu City, China. PLoS One.
9:e1036982014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Guo H, German P, Bai S, Barnes S, Guo W,
Qi X, Lou H, Liang J, Jonasch E, Mills GB, et al: The PI3K/AKT
Pathway and Renal Cell Carcinoma. J Genet Genomics. 42:343–353.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nakanishi A, Wada Y, Kitagishi Y and
Matsuda S: Link between PI3K/AKT/PTEN Pathway and NOX Proteinin
Diseases. Aging Dis. 5:203–211. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu YR, Qi HJ, Deng DF, Luo YY and Yang SL:
MicroRNA-21 promotes cell proliferation, migration, and resistance
to apoptosis through PTEN/PI3K/AKT signaling pathway in esophageal
cancer. Tumour Biol. 37:12061–12070. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tsunoda T, Ishikura S, Doi K, Matsuzaki H,
Iwaihara Y and Shirasawa S: Resveratrol induces luminal apoptosis
of human colorectal cancer HCT116 cells in three-dimensional
culture. Anticancer Res. 34:4551–4555. 2014.PubMed/NCBI
|
|
38
|
Fu Y, Chang H, Peng X, Bai Q, Yi L, Zhou
Y, Zhu J and Mi M: Resveratrol inhibits breast cancer stem-like
cells and induces autophagy via suppressing Wnt/β-catenin signaling
pathway. PLoS One. 9:e1025352014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Larrosa M, Tomás-Barberán FA and Espín JC:
The grape and wine polyphenol piceatannol is a potent inducer of
apoptosis in human SK-Mel-28 melanoma cells. Eur J Nutr.
43:275–284. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu WH and Chang LS: Suppression of
Akt/Foxp3-mediated miR-183 expression blocks Sp1-mediated ADAM17
expression and TNFα-mediated NFκB activation in piceatannol-treated
human leukemia U937 cells. Biochem Pharmacol. 84:670–680. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kang CH, Moon DO, Choi YH, Choi IW, Moon
SK, Kim WJ and Kim GY: Piceatannol enhances TRAIL-induced apoptosis
in human leukemia THP-1 cells through Sp1- and ERK-dependent DR5
up-regulation. Toxicol In Vitro. 25:605–612. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lee RC, Feinbaum RL and Ambros V: The C.
elegans heterochronic gene lin-4 encodes small RNAs with antisense
complementarity to lin-14. Cell. 75:843–854. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wu QB, Chen J, Zhu JW, Yin X, You HY, Lin
YR and Zhu HQ: MicroRNA 125 inhibits RKO colorectal cancer cell
growth by targeting VEGF. Int J Mol Med. 42:665–673.
2018.PubMed/NCBI
|
|
44
|
Zhang J, Yan YG, Wang C, Zhang SJ, Yu XH
and Wang WJ: MicroRNAs in osteosarcoma. Clin Chim Acta. 444:9–17.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kumar A, Rimando AM and Levenson AS:
Resveratrol and pterostilbene as a microRNA-mediated
chemopreventive and therapeutic strategy in prostate cancer. Ann N
Y Acad Sci. 1403:15–26. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ke K, Sul OJ, Rajasekaran M and Choi HS:
MicroRNA-183 increases osteoclastogenesis by repressing heme
oxygenase-1. Bone. 81:237–246. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lu J, Song G, Tang Q, Yin J, Zou C, Zhao
Z, Xie X, Xu H, Huang G, Wang J, et al: miR-26a inhibits stem
cell-like phenotype and tumor growth of osteosarcoma by targeting
Jagged1. Oncogene. 36:231–241. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Cheng D, Qiu X, Zhuang M, Zhu C, Zou H and
Liu Z: MicroRNAs with prognostic significance in osteosarcoma: A
systemic review and meta-analysis. Oncotarget. 8:81062–81074. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Song MS, Salmena L and Pandolfi PP: The
functions and regulation of the PTEN tumour suppressor. Nat Rev Mol
Cell Biol. 13:283–296. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Butler MS, Robertson AAB and Cooper MA:
Natural product and natural product derived drugs in clinical
trials. Nat Prod Rep. 31:1612–1661. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Pan M-H, Ghai G and Ho CT: Food
bioactives, apoptosis, and cancer. Mol Nutr Food Res. 52:43–52.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Aravindaram K and Yang NS:
Anti-inflammatory plant natural products for cancer therapy. Planta
Med. 76:1103–1117. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tosetti F, Noonan DM and Albini A:
Metabolic regulation and redox activity as mechanisms for
angioprevention by dietary phytochemicals. Int J Cancer.
125:1997–2003. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Shammas MA, Koley H, Batchu RB, Neri P,
Tassone P, Prabhala R, Anderson KC and Munshi NC: Specific Killing
of Multiple Myeloma Cancer Cells by Epigallocatechin-3-Gallate
Extracted from Green Tea. Blood. 104:24612004. View Article : Google Scholar
|
|
55
|
Kwon KH, Barve A, Yu S, Huang MT and Kong
ANT: Cancer chemoprevention by phytochemicals: Potential molecular
targets, biomarkers and animal models. Acta Pharmacol Sin.
28:1409–1421. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Singh RP, Tyagi A, Sharma G, Mohan S and
Agarwal R: Oral silibinin inhibits in vivo human bladder tumor
xenograft growth involving down-regulation of survivin. Clin Cancer
Res. 14:300–308. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ku KL, Chang PS, Cheng YC and Lien CY:
Production of stilbenoids from the callus of Arachis hypogaea: A
novel source of the anticancer compound piceatannol. J Agric Food
Chem. 53:3877–3881. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Chen XR, Lu R, Dan HX, Liao G, Zhou M, Li
XY and Ji N: Honokiol: A promising small molecular weight natural
agent for the growth inhibition of oral squamous cell carcinoma
cells. Int J Oral Sci. 3:34–42. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Yang J, Zou Y and Jiang D: Honokiol
suppresses proliferation and induces apoptosis via regulation of
the miR 21/PTEN/PI3K/AKT signaling pathway in human osteosarcoma
cells. Int J Mol Med. 41:1845–1854. 2018.PubMed/NCBI
|