Introduction
Severe trauma, tumor resection, cancer or congenital
diseases can result in large segmental bone defects (1). Synthetic bone graft substitutes
combined with bioactive molecules, such as growth factors, peptides
and small molecules, can improve the delivery of bone precursor
cells, as well as subsequent bone formation and metabolism
(2). Bioactive molecules are
required in patients with larger bone defects (>2 cm) (2). For this purpose, growth factors,
peptides and small molecules are currently being evaluated at the
pre-clinical and clinical levels (2). Based on previous findings, combining
Neuropeptide Y (NPY) with the appropriate corresponding bone
scaffold may bring about effective clinical translations and novel
insights into the future of bone transplantation.
NPY, a 36-amino acid peptide, is a neurotransmitter
located in the brain and autonomic nervous system (3). As a typical regulator of appetite and
energy balance, NPY serves an important role in the central nervous
system (4), as well as a
protective role in chronic bone loss-induced stress (5). NPY, which is stored and released
together with noradrenaline in the process of nerve stimulation, is
part of the sympathetic arm of the peripheral nervous system
(4). NPY has been identified in
several cell types, including osteoblasts and adipocytes (6–8), and
has previously been demonstrated to regulate the balance of bone
mass in both the central and peripheral regions (9,10).
NPY receptors are members of the G protein-coupled
receptor superfamily, and five subtypes (Y1, 2, 4, 5 and 6) are
currently acknowledged in humans (11). However, in rodents, bone mass is
regulated by the Y1 and Y2 receptors only (12). In previous studies, inhibiting the
Y1 receptor increased the mRNA expression levels of osterix and
runt-related transcription factor 2 (Runx2), two transcription
factors essential for osteoblastic differentiation and bone
formation (13). Y1 inhibition can
also promote the activity of Runx2 and osterix, and thus may
contribute to osteoblastic differentiation and the expression of
osteocalcin (OCN), alkaline phosphatase (ALP), bone sialoprotein
(BSP) and collagen 1α (14).
However, whether NPY itself is able to modulate osteoblasts via
Runx2, osterix and bone morphogenetic protein (BMP) signaling
remains to be elucidated.
Igwe et al (15) revealed that NPY is expressed in
osteoblasts and serves an important role in osteogenic
differentiation. They also identified higher NPY mRNA expression in
bone fractions highly abundant in mature osteoblasts and
osteocytes. In a previous study, treatment of osteoblasts with NPY
decreased the expression of markers of osteoblast differentiation
(9). Collectively, the
aforementioned studies focused on cellular proliferation and
osteogenesis as a result of the exogenous administration of NPY.
However, little is known of the effects of NPY alterations at the
genetic level. In the present study, the aim of the study was to
investigate the effects of regulating NPY by an autocrine mechanism
(coincident with the regulation of Runx2 and osterix) using the
MC3T3-E1 cell line.
In a previous report, non-tumorous MC3T3-E1 cells
were extracted from the skull and underwent a typical osteoblastic
differentiation process in vitro (16). In the present study, experiments
were conducted by transfecting MC3T3-E1 cells with NPY small
interfering RNA (siRNA) or overexpression plasmids to alter its
expression level; this method demonstrated the autocrine, rather
than the exocrine mechanism by which NPY regulates osteoblastic
function. The osteoblastic ability of MC3T3-E1 cells was
significantly enhanced following NPY overexpression, resulting in
an increase in the expression of ALP, OCN, Runx2 and osterix mRNA
levels at different time points, compared with those in untreated
cells. In conclusion, this study demonstrated that NPY signaling
serves a positive role in osteogenic differentiation of MC3T3-E1
cells by upregulating Runx2 and osterix in vitro. This will
help to solve the clinical problem of large bone defects caused by
severe trauma, tumor resection, cancer or congenital diseases in
the future.
Materials and methods
Cell isolation, culture and
treatment
The MC3T3-E1 cell line was purchased from the Cell
Bank of Union Medical University (http://sbm.pumc.edu.cn/). MC3T3-E1 cells were cultured
in α-minimum essential medium (MEM with 10% Fetal Bovine Serum,
both Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10
mmol β-glycerophosphate, 50 µg/ml ascorbic acid and 10 mmol
dexamethasone, and maintained at 37°C in 5% CO2. The
culture medium was renewed every 2–3 days.
Expression plasmids and
transfection
MC3T3-E1 cells (2.0×106 cells/well) were
seeded into 6-well plates (2 ml/well) and cultured at 37°C (5%
CO2) until 70–80% confluence. In order to study the
subsequent biological effects of NPY on MC3T3-E1 cells, three
different groups of siRNA (final concentration: 10 nM) and an NPY
overexpression plasmid (final concentration: 25 nM) were designed
by Wuhan Qingke Innovation Biotech Co., Ltd., along with a
scrambled siRNA group (Wuhan Qingke Innovation Biotech Co., Ltd.)
and an empty vector group (Wuhan Qingke Innovation Biotech Co.,
Ltd.), which acted as the negative controls. The three NPY siRNA
target sequences (siRNA1, siRNA2 and siRNA3) are displayed in
Table I. Then, untransfected cells
were used as the control group for subsequent experiments. The
cells were transfected with NPY siRNA and overexpression plasmids;
NPY siRNA transfection was conducted for 48 h using RFect siRNA
transfection reagent (Changzhou Baidai Biotechnology Co., Ltd.),
and the NPY overexpression plasmid was transfected using RFect DNA
transfection reagent (Changzhou Baidai Biotechnology Co., Ltd.),
also for 48 h. Material synthesis and reagent preparation used in
RFect siRNA and plasmid DNA transfection were performed according
to the instructions of the transfection kits. Statistical analysis
results showed that NPY siRNA3 had the highest interference
efficiency, and as a result, siRNA3 was used for subsequent
experimentation. The cells were then cultured for 7 days in the
same conditions as aforementioned.
 | Table I.Primers used for reverse
transcription-quantitative PCR. |
Table I.
Primers used for reverse
transcription-quantitative PCR.
| Gene name | Primer sequences
(5′→3′) | GenBank number |
|---|
| NPY | Sense:
CTCGTGTGTTTGGGCATTC | NM_023456 |
|
| Antisense:
TAGTGTCGCAGAGCGGAGTA |
|
| siRNA1 | Sense:
CCAAACAAUAGACAUGCUUdTdT | NM_000909.6 |
|
| Antisense:
AAGCAUGUCUAUUGUUUGGdTdT |
|
| siRNA2 | Sense:
GCCCUUUGUGAUCUAUCAAdTdT | NM_000909.6 |
|
| Antisense:
UUGAUAGAUCACAAAGGGCdTdT |
|
| siRNA3 | Sense:
CCAAGCGAAUCAACAUCAUdTdT | NM_000909.6 |
|
| Antisense:
AUGAUGUUGAUUCGCUUGGdTdT |
|
| Runx2 | Sense:
GACGAGGCAAGAGTTTCACC | NM_009820 |
|
| Antisense:
GGACCGTCCACTGTCACTTT |
|
| OCN | Sense:
CAAGCAGGGAGGCAATAAGG | NM_007541 |
|
| Antisense:
CGTCACAAGCAGGGTTAAGC |
|
| GAPDH | Sense:
TCCACCACCCTGTTGCTGTA | NM_002046.3 |
|
| Antisense:
ACCACAGTCCATGCCATCAC |
|
| ALP | Sense:
TGGTCACAGCAGTTGGTAGC | NM_001287172.1 |
|
| Antisense:
CTGAGATTCGTCCCTCGCTG |
|
| Osterix | Sense:
GATGGCGTCCTCTCTGCTTG | NM_130458.3 |
|
| Antisense:
TCTTTGTGCCTCCTTTCCCC |
|
Reverse transcription-quantitative
(RT-q)PCR analysis
At 4 and 7 days post-transfection, MC3T3-E1 cells
were collected and gene expression was detected by RT-qPCR. Total
RNA was isolated using TRIzol® reagent (Tiangen Biotech
Co., Ltd.) and quantified using the NanoDrop™ 2000 (Thermo Fisher
Scientific, Inc.). The RNA was reverse transcribed into cDNA using
the RevertAid First Strand cDNA Synthesis kit (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol; the
primer sequences for NPY, Runx2, OCN, ALP, osterix and GADPH (Wuhan
Qingke Innovation Biotechnology Co., Ltd.) are displayed in
Table I. The FastStart Universal
SYBR-Green Master (Rox) was then used to perform qPCR with the
LightCycler® 480 Instrument II system (both Roche
Diagnostics), under the following thermocycling conditions: 95°C
for 5 min, 44 cycles at 95°C for 15 sec, and then 72°C for 20 sec.
Melting curve analysis was used to confirm the specificity of
transcription amplification. Fold changes in target gene expression
were quantified using the 2−∆∆Cq method (17) after normalization to the internal
reference gene (GAPDH).
Western blot analysis
The cells were harvested and lysed with RIPA lysis
buffer containing phenylmethane sulfonyl fluoride and protease
inhibitors (both Sigma-Aldrich; Merck KGaA). The concentration of
each sample was determined by a bicinchoninic acid protein assay.
Proteins from each group (30 µg/lane) were resolved via SDS-PAGE on
a 10% gel, and subsequently transferred to PVDF membranes. After
blocking with 5% BSA (Sigma-Aldrich; Merck KGaA) at room
temperature for 1 h in a phosphate buffered solution (PBS with 0.1%
Tween-20; pH 7.5) and washing in TBS with 0.05% Tween-20 (TBST),
the membranes were incubated with anti-NPY (1:1,000; cat. no.
DF6431; Affinity Biosciences), anti-OCN (1:500; cat. no.
PAA471Mu01; Cloud-Clone Corp.), anti-ALP (1:1,500; cat. no.
11187-1-AP; ProteinTech Group, Inc.), anti-osterix (1:1,000; cat.
no. bs-1110R; BIOSS) and anti-Runx2 (1:500; cat. no. PAB011Mu01;
Cloud-Clone Corp.) antibodies overnight at 4°C. After rinsing with
TBST, the membranes were all incubated with a secondary antibody
(horseradish peroxidase-linked guinea pig anti-rabbit IgG
polyclonal; 1:2,000; cat. no. SAA544Rb59; Cloud-Clone Corp.) at 4°C
for 1 h, and then developed using a chemiluminescence detection
system (Pierce; Thermo Fisher Scientific, Inc.). The data were
analyzed using ImageJ software (Version 1.8.0; National Institutes
of Health); the relative protein expression level was calculated as
the grey value ratio of the target protein to GAPDH.
Statistical analysis
The experimental data are presented as the mean ± SD
of three independently performed experiments. One-way ANOVA was
used to analyze the overall differences between multiple groups.
The Scheffe (Fig. 1A and B) or
least significant difference post hoc tests were used to evaluate
the differences between groups using SPSS 18.0 software (SPSS,
Inc.); representative bar graphs were created in GraphPad Prism
v.6.0 (GraphPad Software, Inc.). P<0.05 was considered to
indicate a statistically significant difference.
Results
Screening of NPY-specific siRNA
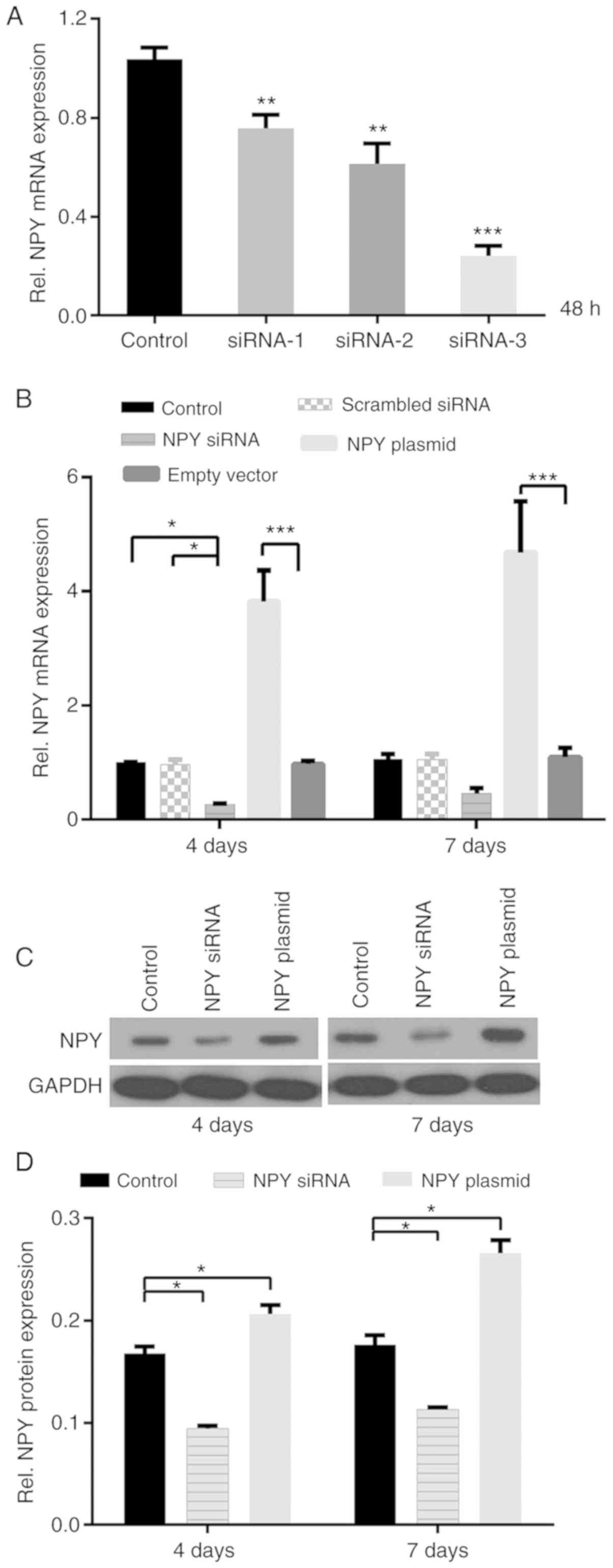
Comparison of the control, siRNA1, siRNA2 and siRNA3
groups was conducted by one-way ANOVA followed by Scheffe post hoc
test, and the differences among the groups were statistically
significant (P<0.01 or P<0.001). NPY siRNA3 had the highest
interference efficiency of >75%, and as a result, siRNA3 was
used for subsequent experimentation (Fig. 1A).
Secretion and expression of NPY in
MC3T3-E1 cells
First, NPY secretion from MC3T3-E1 cells was
verified at 4 and 7 days; NPY was consistently expressed at all
time points, which provided the basis for the following experiments
(Fig. 1B). In addition, NPY mRNA
was sustainably expressed in both the experimental and control
groups (Fig. 1B). Furthermore, the
overexpression plasmids significantly promoted mRNA expression in
MC3T3-E1 cells, which was increased by ~3.9-fold at day 4 and
4.3-fold at day 7, in comparison with the empty vector group
(P<0.001; Fig. 1B).
In order to more fully understand the role of NPY,
MC3T3-E1 cells were transfected with NPY siRNA and cultured for 48
h, and changes in NPY mRNA expression were observed over time. NPY
mRNA expression was decreased by ~0.75-fold at day 4 and ~0.55-fold
at day 7, compared with the control and scrambled siRNA groups;
there was statistical significance at day 4 (P<0.05; Fig. 1B) and no statistical significance
at day 7. Conversely, semi-quantitative analysis revealed that NPY
overexpression increased NPY protein expression by 1.2-fold at day
4 and 1.5-fold at day 7, compared with the control group
(P<0.05; Fig. 1C and D). NPY
siRNA was then used to further elucidate the role of NPY in
MC3T3-E1 cells. NPY protein expression was decreased by ~0.45-fold
and 0.35-fold at days 4 and 7 post-transfection, respectively
(P<0.05; Fig. 1C and D).
NPY is critical for osteoblastic
differentiation
The expression levels of bone-formation factors,
such as ALP and OCN, are determined by the different stages of bone
differentiation (18). In the
present study, ALP and OCN mRNA was consistently expressed in the
three siRNA groups at days 4 and 7 post-transfection (Fig. 2A and B). The effects of NPY
overexpression were then investigated using the MC3T3-E1 cell line,
and NPY overexpression was found to significantly increase ALP mRNA
expression by ~3.7- and 4.9-fold at days 4 and 7, and OCN mRNA
expression by ~4.0- and 4.3-fold at days 4 and 7, respectively,
compared with the control group (P<0.001; Fig. 2A and B).
 | Figure 2.Expression of ALP, OCN, Runx2 and
osterix mRNA in MC3T3-E1 cells. Expression of (A) ALP, (B) OCN, (C)
Runx2 and (D) osterix mRNA at days 4 and 7. *P<0.05,
***P<0.001 vs. control group; ###P<0.001 vs. siRNA
group. NPY, neuropeptide Y; ALP, alkaline phosphatase; OCN,
osteocalcin; Runx2, runt-related transcription factor 2; siRNA,
small interfering RNA. |
To further determine the effects of NPY on ALP and
OCN mRNA, MC3T3-E1 cells were transfected with NPY siRNA for 48 h,
and the effects were observed over a subsequent time period. At
days 4 and 7, ALP mRNA was decreased by ~0.56- and 0.26-fold, and
OCN mRNA was decreased by ~0.60- and 0.35-fold, respectively,
compared with the control group. The differences were statistically
significant at day 4 (P<0.05; Fig.
2A and B). The ALP mRNA were also compared between the siRNA
and overexpression groups, and overexpression was found to increase
these levels ~7.9-fold (compared with siRNA inhibition) at day 4
and ~6.5-fold at day 7, and OCN mRNA levels ~10.1-fold (compared
with siRNA inhibition) at day 4 and ~6.7-fold at day 7,
respectively (P<0.001; Fig. 2A and
B).
Semi-quantitative analysis of protein expression
revealed that OCN protein expression was increased by 1.4-fold at
day 4, and 1.6-fold at day 7 following NPY overexpression, compared
with the control group; ALP protein expression was increased by
1.6-fold at days 4 and 7 (P<0.05; Fig. 3A-C).
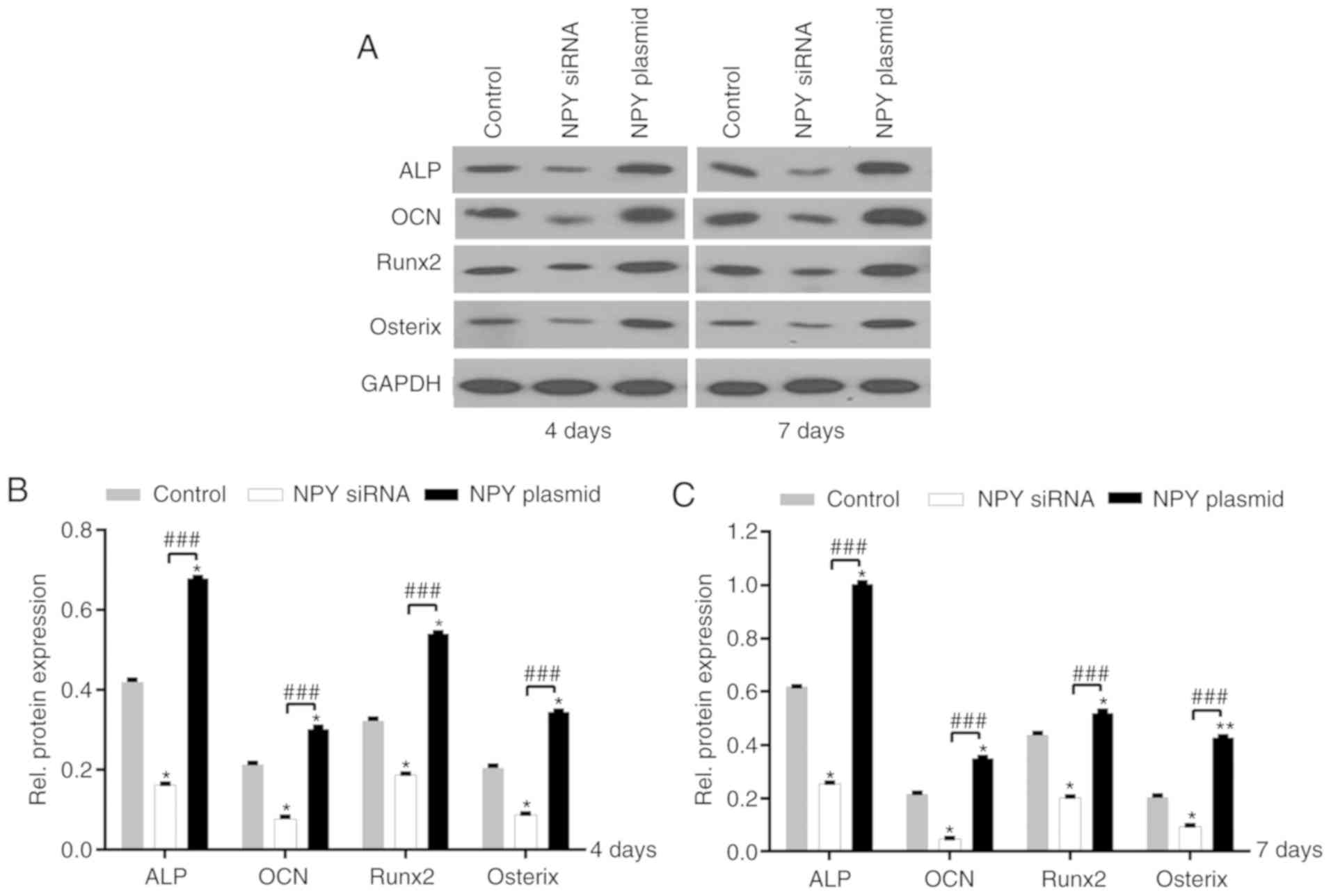 | Figure 3.Expression of ALP, OCN, Runx2 and
osterix protein in MC3T3-E1 cells. (A) Western blotting analysis of
ALP, OCN, Runx2 and osterix protein expression at days 4 and 7.
Semi-quantification of protein expression at (B) day 4 and (C) day
7. *P<0.05, **P<0.01 vs. control group;
###P<0.001 vs. siRNA group. NPY, neuropeptide Y;
siRNA, small interfering RNA; ALP, alkaline phosphatase; OCN,
osteocalcin; Runx2, runt-related transcription factor 2. |
Following NPY siRNA interference, ALP and OCN
protein expression was also assessed; the level of ALP at days 4
and 7 was decreased by ~0.60-fold, and the level of OCN protein
expression was decreased by ~0.64-fold at day 4 and ~0.78-fold at
day 7, compared with the control group (P<0.05; Fig. 3A-C).
NPY regulates Runx2 and osterix
expression in MC3T3-E1 cells
To the best of our knowledge, Runx2 is an important
early transcription factor involved in the expression of
osteoblast-specific genes, and the differentiation of mesenchymal
stem cells into osteoblasts (19).
In addition, Runx2 is hypothesized to serve a primary role in the
terminal differentiation of osteoblasts (20).
Therefore in the present study, the effects of NPY
on Runx2 and osterix expression were determined using MC3T3-E1
cells. NPY overexpression was found to significantly increase Runx2
and osterix mRNA expression. Compared with the control group, Runx2
mRNA expression increased by ~5.4-fold at day 4, and ~6-fold at day
7. Additionally, osterix mRNA was increased by ~2.7-fold at day 4
and ~3-fold at day 7, compared with the control group. Of note, the
level of Runx2 mRNA expression was twice that of osterix
(P<0.001; Fig. 2C and D).
To further understand the effects of NPY on Runx2
and osterix mRNA expression, MC3T3-E1 cells were transfected with
NPY siRNA and the expression levels of Runx2 and osterix mRNA were
determined at subsequent time points. At day 4, Runx2 mRNA
expression was decreased by ~0.50-fold compared with the control
group, and ~0.35-fold at day 7. Osterix mRNA expression decreased
by ~0.80-fold at day 4 and 0.55-fold at day 7, compared with the
control group. The differences in osterix mRNA expression were
statistically significant at day 4 (P<0.001; Fig. 2C and D).
Next, the levels of Runx2 and osterix mRNA
expression were compared between the siRNA and overexpression
groups. In the overexpression group, Runx2 mRNA expression was
10-fold higher than that of the siRNA group at days 4 and 7, and
osterix mRNA expression was ~15-fold greater than that observed in
the siRNA group at day 4, and ~8-fold higher at day 7 (P<0.001;
Fig. 2C and D).
Semi-quantitative analysis of Runx2 and osterix
expression revealed that compared with the control group, NPY
overexpression increased Runx2 protein expression by 1.6-fold at
day 4, and 1.2-fold at day 7. Similarly, osterix protein expression
was increased by 1.6-fold at day 4, and 2.1-fold at day 7, compared
with the control group (P<0.05; Fig. 3A-C).
In the siRNA group, Runx2 protein expression was
decreased by ~0.40- and 0.55-fold at days 4 and 7, respectively,
compared with the control group. There was no statistical
difference in Runx2 mRNA expression in the siRNA group in Fig. 2A, whereas the difference in Runx2
protein expression was statistically significant in Fig. 3A-C (P<0.05). Likewise, osterix
protein expression was decreased by ~0.60- and 0.55-fold at days 4
and 7, respectively, compared with the control group (P<0.05;
Fig. 3A-C).
In conclusion, these findings indicated that NPY may
be a positive regulator of osteogenic differentiation in mouse
MC3T3-E1 cells.
Discussion
NPY is a polypeptide of 36 amino acids, belonging to
a larger family of neuropeptides that also includes the YY peptide
and the pancreatic polypeptide (21,22).
Baldock et al (23) found
that Y1 or Y2 receptor knockout in mice resulted in increased
trabecular and cortical bone mass. However, within the bone
microenvironment, the regulatory mechanisms of NPY at the genetic
level remain unclear. As a result, the aim of the present study was
to conduct an in-depth assessment of these hypotheses in the
MC3T3-E1 cell line.
NPY is a neurotransmitter that has dual functions in
the regulation of energy consumption and bone mass (11). A previous study determined that
SNPs within NPY and neuraminidase (NA) were closely associated with
the susceptibility and prognosis of patients with cervical vertigo
(19). Therefore, NPY and NA could
be used as diagnostic markers and therapeutic targets for cervical
vertigo (19). Igwe et al
(15) revealed that mouse
calvarial osteoblasts fostered in the presence of NPY decreased the
expression of differentiation-related markers, including OCN, BSP
and dentin matrix acidic phosphoprotein 1 (DMP1), and also reduced
the extent of mineralization. Their findings indicated that the
levels of intracellular cAMP and differentiation markers (OCN, BSP
and DMP1) in mouse calvarial osteoblasts were reduced by exogenous
NPY treatment (15). In the
present study, NPY was overexpressed in MC3T3-E1 cells to observe
its effects on osteogenic ability. Overexpressing NPY was found to
markedly enhance the osteogenic ability of mouse MC3T3-E1 cells via
an autocrine mechanism, coincident with the upregulation of Runx2
and osterix. In addition, the mRNA expression levels of ALP, OCN,
osterix and Runx2 were increased at different time points. In
contrast to Igwe et al (15), the present study reported that
endogenous NPY enhanced the expression of MC3T3-E1 cell
differentiation markers, including ALP, OCN, Runx2 and osterix.
BMP signaling stimulates osteoblast differentiation
and osteogenic activity by upregulating osterix and Runx2, which
are essential osteogenic transcription factors (20,24).
Runx2 is an important factor for bone growth, and the heterozygous
loss of Runx2 results in clavicular dysplasia in humans, and the
corresponding phenotype in mice (25). It has also been confirmed that
homozygous Runx2-II−/− mice possess lower levels of bone
mass and hypertrophic cartilage (25). Runx2-I is necessary for early bone
growth and intramembranous ossification, whereas Runx2-II is needed
for the complete maturation of osteoblasts and osteogenesis in
cartilage (26). Osterix, a
downstream effector of Runx2 required for osteoblast
differentiation, is significantly reduced in selective
Runx2-II−/− mice (27).
An associated decrease in the expression of bone ALP, OCN,
osteopontin and matrix extracellular phosphoglycoprotein was also
observed. However, the levels of these markers were normal or
slightly decreased in heterozygous mutant mice (25). Despite this, the relationship
between Runx2 and osterix in osteogenic differentiation (in the
MC3T3-E1 cell line specifically) remains to be elucidated.
Nakashima et al (27) found
that osterix was not expressed in Runx2-II−/− mice. In
the present study, it was hypothesized that NPY may regulate
osteoblast differentiation via the upregulation of Runx2 and
osterix, and the role of NPY in promoting the cell differentiation
and maturation of MC3T3-E1 cells via the Runx2 and osterix pathways
was further validated. In a previous study based on the findings of
Xiao et al (26), Runx2 was
revealed to be the upstream regulatory gene of osterix. Runx2-I is
also involved in maintaining 50% of osterix gene expression in
Runx2-II-deficient (Runx2-II−/−) mice; thus, 50% of
osterix mRNA may be regulated by upstream Runx2 mRNA (26). In the present study (Fig. 2C and D), osterix gene expression
was found to be consistent with that of Runx2, and the level of
osterix gene expression was almost half that of Runx2. It was
therefore hypothesized that osterix is a target gene of Runx2, by
which it can be positively regulated.
The RFect siRNA and plasmid DNA transfection
reagents were specifically designed by Changzhou Baidai
Biotechnology Co., Ltd. (Patent application number: 20100022618)
for transfecting siRNA and DNA into eukaryotic cells; this is a
novel method derived from animal-origin free lipid transfection
reagents, and the technique can be used on a wide range of adherent
cell lines. It has numerous useful features; for example, the
efficiency of transfection is >90%, and the effect of gene
knockdown is markedly apparent (28). Furthermore, minimal cytotoxicity
reduces non-specific effects and cellular stress (28,29).
Low concentrations of siRNA and DNA can be used to obtain high
levels of knockdown and overexpression, respectively (28,29).
In the present study, the interference efficiency of NPY mRNA at
day 4 reached ~75 and 80%, respectively. In addition, the
overexpression of NPY mRNA was ~4- and 6-fold at days 4 and 7,
respectively. However, it was also found that Runx2 gene and
protein expression were not synchronized at the same time point.
Following NPY knockdown, the expression of Runx2 mRNA only showed a
slight decrease, whereas the alteration in Runx2 protein expression
was statistically significant. One reason could be that the
expression of Runx2 protein determined the behavior of the mouse
MC3T3-E1 cells, whereas Runx2 mRNA detection had temporal and
spatial specificity, which may mean that the expression of Runx2
mRNA and protein are not in sync at the same time point. This
mechanism was also supported by previous articles (24,25).
Nevertheless, there were numerous limitations of the present study.
One limitation is that the osteogenic differentiation was performed
with the use of a single cell line for only 7 days, which also
increases the uncertainty underlying the function of NPY in Runx2.
Adding extra cell lines and prolonging the experiment time would
increase the accuracy and validity of the experiment. Furthermore,
the mechanism of action of NPY on Runx2 is not clear, and further
research is required in the future. Additional cell lines and more
time points for osteogenic differentiation are needed in further
experiments. However, overall, the RFect plasmid DNA and siRNA
transfection technique, which was adopted in the present study, was
found to be reliable for use with the MC3T3-E1 cell line.
Based on the results of previous studies (2,30–32),
combining bioactive molecules, such as growth factors, peptides and
small molecules with corresponding bone scaffold may lead to
effective clinical transformation and provide new insights into the
future of bone transplantation. Notably, this study demonstrated
that NPY signaling directly promotes osteogenic differentiation of
MC3T3-E1 cells by upregulating Runx2 and osterix in vitro.
Additionally, NPY serves a positive role in bone formation,
suggesting that NPY may be a therapeutic target for large bone
defects caused by severe trauma, tumor resection, cancer or
congenital diseases in the future.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from The
Second Hospital, Cheeloo College of Medicine, Shandong University,
P.R. China (grant no. 2018YT06).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BZ, XZha and CG designed the study. BZ, XZho and YC
performed the experiments and collected the data. JX performed the
statistical analysis. BZ and XZho wrote the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yang F, Wang J, Hou J, Guo H and Liu C:
Bone regeneration using cell-mediated responsive degradable
PEG-based scaffolds incorporating with rhBMP-2. Biomaterials.
34:1514–1528. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ho-Shui-Ling A, Bolander J, Rustom LE,
Johnson AW, Luyten FP and Picart C: Bone regeneration strategies:
Engineered scaffolds, bioactive molecules and stem cells current
stage and future perspectives. Biomaterials. 180:143–162. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Baraban SC: Neuropeptide Y and limbic
seizures. Rev Neurosci. 9:117–128. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Silva AP, Cavadas C and Grouzmann E:
Neuropeptide Y and its receptors as potential therapeutic drug
targets. Clin Chim Acta. 326:3–25. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Baldock PA, Lin S, Zhang L, Karl T, Shi Y,
Driessler F, Zengin A, Hörmer B, Lee NJ, Wong IP, et al:
Neuropeptide Y attenuates stress-induced bone loss through
suppression of noradrenaline circuits. J Bone Miner Res.
29:2238–2249. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kuo LE, Kitlinska JB, Tilan JU, Li L,
Baker SB, Johnson MD, Lee EW, Burnett MS, Fricke ST, Kvetnansky R,
et al: Neuropeptide Y acts directly in the periphery on fat tissue
and mediates stress-induced obesity and metabolic syndrome. Nat
Med. 13:803–811. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Baldock PA, Allison SJ, Lundberg P, Lee
NJ, Slack K, Lin EJD, Enriquez RF, McDonald MM, Zhang L, During MJ,
et al: Novel role of Y1 receptors in the coordinated regulation of
bone and energy homeostasis. J Biol Chem. 282:19092–19102. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang K, Guan H, Arany E, Hill DJ and Cao
X: Neuropeptide Y is produced in visceral adipose tissue and
promotes proliferation of adipocyte precursor cells via the Y1
receptor. FASEB J. 22:2452–2464. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Khor EC and Baldock P: The NPY system and
its neural and neuroendocrine regulation of bone. Curr Osteoporos
Rep. 10:160–168. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Horsnell H and Baldock PA: Osteoblastic
actions of the neuropeptide Y system to regulate bone and energy
homeostasis. Curr Osteoporos Rep. 14:26–31. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lin S, Boey D and Herzog H: NPY and Y
receptors: Lessons from transgenic and knockout models.
Neuropeptides. 38:189–200. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Allison SJ, Baldock PA and Herzog H: The
control of bone remodeling by neuropeptide Y receptors. Peptides.
28:320–325. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yahara M, Tei K and Tamura M: Inhibition
of neuropeptide Y Y1 receptor induces osteoblast differentiation in
MC3T3E1 cells. Mol Med Rep. 16:2779–2784. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Karsenty G: Transcriptional control of
skeletogenesis. Annu Rev Genomics Hum Genet. 9:183–196. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Igwe JC, Jiang X, Paic F, Ma L, Adams DJ,
Baldock PA, Pilbeam CC and Kalajzic I: Neuropeptide Y is expressed
by osteocytes and can inhibit osteoblastic activity. J Cell
Biochem. 108:621–630. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sudo H, Kodama HA, Amagai Y, Yamamoto S
and Kasai S: In vitro differentiation and calcification in a new
clonal osteogenic cell line derived from newborn mouse calvaria. J
Cell Biol. 96:191–198. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen J, He G, Wang Y and Cai D:
MicroRNA-223 promotes osteoblast differentiation of MC3T3-E1 cells
by targeting histone deacetylase 2. Int J Mol Med. 43:1513–1521.
2019.PubMed/NCBI
|
|
19
|
Han J, Zuo J, Zhu D and Gao C: The
correlation between SNPs within the gene of adrenergic receptor and
neuropeptide Y and risk of cervical vertigo. J Clin Lab Anal.
32:e223662018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li L, Sapkota M, Gao M, Choi H and Soh Y:
Macrolactin F inhibits RANKL-mediated osteoclastogenesis by
suppressing Akt, MAPK and NFATc1 pathways and promotes
osteoblastogenesis through a BMP-2/smad/Akt/Runx2 signaling
pathway. Eur J Pharmacol. 815:202–209. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tatemoto K, Carlquist M and Mutt V:
Neuropeptide Y-a novel brain peptide with structural similarities
to peptide YY and pancreatic polypeptide. Nature. 296:659–660.
1982. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Allen JM, Adrian TE, Tatemoto K, Polak JM,
Hughes J and Bloom SR: Two novel related peptides, neuropeptide Y
(NPY) and peptide YY (PYY) inhibit the contraction of the
electrically stimulated mouse vas deferens. Neuropeptides. 3:71–77.
1982. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Baldock PA, Sainsbury A, Allison S, Lin
EJ, Couzens M, Boey D, Enriquez R, During M, Herzog H and Gardiner
EM: Hypothalamic control of bone formation: distinct actions of
leptin and y2 receptor pathways. J Bone Miner Res. 20:1851–1857.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Moon JS, Kim SH, Oh SH, Jeong YW, Kang JH,
Park JC, Son HJ, Bae S, Park BI, Kim MS, et al: Relaxin augments
BMP-2-induced osteoblast differentiation and bone formation. J Bone
Miner Res. 29:1586–1596. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hecht J, Seitz V, Urban M, Wagner F,
Robinson PN, Stiege A, Dieterich C, Kornak U, Wilkening U, Brieske
N, et al: Detection of novel skeletogenesis target genes by
comprehensive analysis of a Runx2(−/-) mouse model. Gene Expr
Patterns. 7:102–112. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xiao ZS, Hjelmeland AB and Quarles LD:
Selective deficiency of the ‘bone-related’ Runx2-II unexpectedly
preserves osteoblast-mediated skeletogenesis. J Biol Chem.
279:20307–20313. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nakashima K, Zhou X, Kunkel G, Zhang Z,
Deng JM, Behringer RR and de Crombrugghe B: The novel zinc
finger-containing transcription factor osterix is required for
osteoblast differentiation and bone formation. Cell. 108:17–29.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang X, Yang Y, Sun BF, Chen YS, Xu JW,
Lai WY, Li A, Wang X, Bhattarai DP, Xiaot W, et al:
5-methylcytosine promotes mRNA export-NSUN2 as the
methyltransferase and ALYREF as an m5C reader. Cell Res.
27:606–625. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu J, Fang H, Chi Z, Wu Z, Wei D, Mo D,
Niu K, Balajee AS, Hei TK, Nie L and Zhao Y: XPD localizes in
mitochondria and protects the mitochondrial genome from oxidative
DNA damage. Nucleic Acids Res. 43:5476–5488. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gao C, Peng S, Feng P and Shuai C: Bone
biomaterials and interactions with stem cells. Bone Res.
5:170592017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shuai C, Xu Y, Feng P, Wang G, Xiong S and
Peng S: Antibacterial polymer scaffold based on mesoporous
bioactive glass loaded with in situ grown silver. Chemical
Engineering J. 374:304–315. 2019. View Article : Google Scholar
|
|
32
|
Yang Y, Zan J, Yang W, Qi F, He C, Huang
S, Peng S and Shuai C: Metal organic frameworks as a compatible
reinforcement in a biopolymer bone scaffold. Materials Chem Front.
4:973–984. 2020. View Article : Google Scholar
|