Introduction
Trophoblast migration and invasion are proposed to
play a pivotal role in the process of embryo implantation and
placentation, which are critical for embryonic development and
successful pregnancy (1). Numerous
similarities exist between embryo implantation and the growth of
cancer cells; however, the former is stringently controlled both
spatially and temporally (2,3). The
invasion of trophoblasts to a sufficient depth of the uterus is a
key factor in determining the outcome of pregnancy (1). Dysregulation of this invasive
behavior can trigger a wide spectrum of pregnancy-related diseases
which can occur throughout various stages of pregnancy (4). Inadequate invasion can lead to
pathological pregnancies such as infertility (5), miscarriage (6–8),
premature birth (5), fetal growth
restriction (5,7) and preeclampsia (PE) (5,7,9),
while excessive invasion has been implicated in the pathophysiology
of placenta accreta (1,5) and premalignant or malignant
conditions, such as hydatidiform moles and choriocarcinoma (CH)
(10). Despite improvements in
diagnosis and treatment of these conditions, they can still lead to
the illness and mortality of mother and child. Hence, it is
imperative to find effective prevention and treatment measures for
pregnancy-related events from the perspective of trophoblast
invasion.
MicroRNAs (miRNA/miRs) are a class of noncoding RNAs
~22 nucleotides in length that post-transcriptionally regulate
diverse cellular processes by targeting mRNAs for cleavage or
translational repression (11,12).
It has been suggested that miRNAs are involved in placental
development and function (13). In
addition, recent data have also indicated that miRNAs play a
fundamental role in trophoblast proliferation, migration, invasion
and apoptosis (14,15). For example, it has been reported
that miR-184 was highly expressed in recurrent spontaneous abortion
(SA) and targeted zinc finger matrin-type 3 (ZMAT3) to promote the
apoptosis of trophoblasts by upregulating Fas expression levels
(16). miR-616-3p modulated cell
proliferation and migration via targeting tissue factor pathway
inhibitor 2 in PE (17). miR-21
was upregulated in hydatidiform mole tissues and promoted CH cell
proliferation, migration and invasion (18). However, the association between
miRNAs and trophoblast biological function in each of these
previous studies was evaluated for a single event, such as PE or
SA, thereby lacking a comprehensive analysis of such
pregnancy-related events.
In the present study, samples were collected from
multiple conditions with abnormal invasion capacity, and it was
found that miR-181b-5p was highly expressed in events with impaired
trophoblast invasiveness, such as SA (19) and PE (20,21),
but lowly expressed in CH which has excessive invasiveness
(10,22). These findings suggested that
miR-181b-5p may have a pivotal role in multiple abnormal
trophoblast invasion-related events involved in development or
pathogenesis. Moreover, growing evidence has indicated that
miR-181b-5p functions in regulating tumor cell migration and
invasion (23,24). Of note, it has been suggested that
trophoblasts and tumor cells share similar biological
characteristics, including proliferation, migration, invasion and
apoptosis (25). Consequently, the
vital role of miR-181b-5p in tumor cells is suggestive of its
effect on the regulation of trophoblast cell migration and
invasion. In the present study, bioinformatics analysis predicted
that miR-181b-5p potentially targets the 3′-untranslated region
(UTR) of sphingosine-1-phosphate (S1P) receptor 1 (S1PR1) mRNA;
this was investigated via luciferase reporter assays. Therefore,
the functional investigation of miR-181b-5p and its downstream
targets in multiple abnormal trophoblast invasion-related events
was the primary aim of the present study.
Materials and methods
Sample collection
Peripheral blood and tissue samples were obtained
from 100 females at the Department of Obstetrics and Gynecology,
Nanjing Maternity and Child Health Care Hospital from March 2013 to
March 2018. Among these females, there were 40 healthy controls (20
first trimester, mean age, 30.9±5.2 years; 20 third trimester, mean
age, 31.3±4.6 years), and 20 cases of PE (mean age, 30.7±5.8
years), SA (mean age, 32.6±3.6 years) and CH (mean age, 32.4±3.2
years), respectively. A total of 5 ml blood was extracted from
middle vein of the elbow of each participant. Red blood cells were
removed by centrifugation at 1,880 × g for 20 min at 4°C, and
plasma was stored at −20°C prior to use. Villi tissues were
collected from normal early pregnancy (NV) and patients with SA,
placenta samples were collected from normal full-term pregnancy
(NP) and PE, and tumor tissue samples were collected from
individuals with CH. PE was defined as systolic blood pressure ≥140
mm Hg and/or diastolic blood pressure ≥90 mmHg on a minimum of two
occasions at least 4–6 h apart, with urinary protein ≥0.3 g/24 h
after 20 weeks gestation. All women with NV terminated pregnancy
for nonmedical reasons. Samples of SA were excluded if due to other
causes, such as parental chromosomal, fetal chromosome abnormality,
autoimmunity, thrombophilia, reproductive malformation, hormonal
dysfunction (such as polycystic ovarian syndrome, thyroid
abnormalities, diabetes mellitus, hyperprolactinemia or luteal
insufficiency), folate deficiency or infection. All CH samples were
collected after hysterectomy from patients without detectable
infections or other systemic conditions. The clinical
characteristics of these cases are shown in Tables I and II. The present study was approved by the
Ethics Committee of the Nanjing Maternity and Child Health Care
Hospital affiliated to Nanjing Medical University (Nanjing, China),
and all enrolled patients provided written consent for sample
collection and analyses.
 | Table I.Clinical characteristics of PE
pregnancies and CH. |
Table I.
Clinical characteristics of PE
pregnancies and CH.
| Variable | PE (n=20) | NP (n=20) | CH (n=20) | P-value (PE vs.
NP) | P-value (CH vs.
NP) |
|---|
| Maternal age,
years | 30.7±5.8 | 31.3±4.6 | 32.4±3.2 | >0.05 | >0.05 |
| Week of
pregnancy | 35.2±3.9 | 38.5±2.1 | After
pregnancy | >0.05 | – |
| Birth weight,
g | 2,412±872 | 3,312±415 | – | <0.05 | – |
| Systolic blood
pressure, mmHg | 165±20.5 | 111±10.4 | 117±13.5 | <0.01 | >0.05 |
| Diastolic blood
pressure, mmHg | 109±16.1 | 71±8.3 | 76±6.4 | <0.01 | >0.05 |
| Proteinuria,
g/day | 5.23±1.34 | <0.3 | <0.3 | <0.01 | >0.05 |
| Table II.Clinical characteristics of SA and NV
in the first trimester. |
Table II.
Clinical characteristics of SA and NV
in the first trimester.
| Variable | SA (n=20) | NV (n=20) | P-value (SA vs.
NV) |
|---|
| Maternal age,
years | 32.6±3.6 | 30.9±5.2 | >0.05 |
| Week of
pregnancy | 8.4±2.5 | 9.1±1.3 | >0.05 |
Cell culture
HTR-8/SVneo cells, derived from primary first
trimester human villous explant trophoblasts (26) and considered as a representative
model of human extravillous trophoblasts (EVTs) (27), were selected as an in vitro
model for the present study. HTR-8/SVneo cells were purchased from
The Cell Bank of Type Culture Collection of the Chinese Academy of
Sciences and were cultured in RPMI-1640 medium containing 10% FBS
(both Gibco; Thermo Fisher Scientific, Inc.), penicillin (100 U/ml)
and streptomycin (100 µg/ml) at 37°C and 5% CO2. Cells
were passaged when they reached 90% confluence.
Oligonucleotide and plasmid
transfection
Hsa-miR-181b-5p mimic and inhibitor, S1PR1
overexpression plasmid and their corresponding negative controls
were purchased from Shanghai GenePharma Co., Ltd. The sequences
were as follows: Has-miR-181b-5p mimics:
5′-AACAUUCAUUGCUGUCGGUGGGU-3′ and 5′-ACCCACCGACAGCAAUGAAUGUU-3′;
Has-miR-181b-5p mimics-NC: 5′-UUUGUACUACACAAAAGUACUG-3′ and
5′-CAGUACUUUUGUGUAGUACAAA-3′; Has-miR-181b-5p inhibitor:
5′-ACCCACCGACAGCAAUGAAUGUU-3′; and Has-miR-181b-5p inhibitor-NC:
5′-CAGUACUUUUGUGUAGUACAAA-3′. HTR-8/SVneo cells were seeded onto
6-well plates at a density of 1×105 cells/well. When
cells were 70–90% confluent, transient transfections were conducted
using Lipofectamine® RNAi-MAX for oligonucleotides
transfections or Lipofectamine 2000 for plasmid transfections
(Thermo Fisher Scientific, Inc.), according to the manufacturer's
instructions. The final concentration of all mimics, inhibitor and
plasmid used for transfection was 100 nM. Transfection medium was
replaced 8 h post-transfection and cells were harvested at 48 h for
further experimentation. Transfection efficiency was examined via
reverse transcription-quantitative PCR (RT-qPCR).
In vitro migration and invasion
assays
An in vitro scratch assay was performed to
evaluate cell migration ability. When cells reached 90% confluence,
wounds were made by scraping confluent cell monolayers with a
100-µl pipette tip, and non-adherent cells and debris were removed
using PBS. In order to minimize cell proliferation, cells were
cultured in serum-free medium. The width of the scratch was
monitored every 12 h, and the wound width was measured using a
Nikon ECLIPSE Ts2 light microscope (Nikon Corporation;
magnification, ×10) at 0 and 48 h, respectively. The percentage of
wound closure areas was calculated using the following formula:
(Wound width at 0 h-wound width at 48 h)/wound width at 0 h ×100%
using ImageJ software version 1.8.0 (National Institutes of
Health).
Invasion assays were performed using Transwell
inserts (Costar; Corning, Inc.) pre-coated with Matrigel (BD
Biosciences). Briefly, a total of 5×104 transfected
cells suspended in 200 µl RPMI-1640 medium without FBS were seeded
in the upper compartment of Transwell inserts. As a
chemoattractant, medium containing 20% FBS was added to the lower
well. After a 24-h incubation at 37°C and 5% CO2, the
cells remaining in the top side of the insert membrane (non-invaded
cells) were gently cleared with cotton swabs and the cells invading
to the bottom side of the insert membrane were fixed using 4%
paraformaldehyde for 30 min at room temperature. After washing with
PBS, the fixed cells were stained using 0.1% crystal violet for 30
min at room temperature, and the number of stained cells was
counted using Image J software version 1.8.0 (National Institutes
of Health) under a light microscope (Olympus Corporation,
magnification, ×10). A total of five random field views were
selected for counting.
RT-qPCR
Total RNA was extracted from plasma, tissue and
cells using TRIzol® reagent (Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions. Then, the RNA
was reverse-transcribed into cDNA using Reverse Transcription kit
(Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. RT-qPCR was performed using a SYBR Premix Ex Taq™ II
(Perfect Real-Time) kit (Takara Bio, Inc.) and
Mastercycler® ep qrealplex thermal cycler (Eppendorf) as
previously described (28,29). The thermocycling conditions were as
follows: Pre-degeneration at 95°C for 15 min, followed by 40 cycles
at 95°C for 5 sec, and annealing at 60°C for 30 sec. The highly
conserved and universally expressed small nuclear RNA U6 was used
as an endogenous control to normalize the miRNA RT-qPCR data.
β-actin expression was used for the normalization of S1PR1
expression. All primers were synthesized by Thermo Fisher
Scientific, Inc. The following primers were used: S1PR1 forward,
5′-CAGCAAATCGGACAATTCCT-3′ and reverse, 5′-GCCAGCGACCAAGTAAAGAG-3′;
β-actin forward, 5′-CCTGGCACCCAGCACAAT-3′ and reverse,
5′-GGGCCGGACTCGTCATAC-3′; CXCL3: Forward, 5′-TCCGTGGTCACTGAACTGC-3′
and reverse, 5′-AGTTGGTGCTCCCCTTGTTC-3′; PPP3R1: Forward,
5′-TGGAAGAGTTCATGTCTCTGCCTGA-3′ and reverse,
5′-TGACACTGAACTGAGAGACGCCCT-3′; miR-181b-5p: Forward,
5′-CCAGCTGGGCTCACTGAACAATGA-3′ and reverse,
5′-CAACTGGTGTCGTGGAGTCGGC-3′; and U6: Forward,
5′-CTCGCTTCGGCAGCACAT-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′.
RT-qPCR data were analyzed using the 2−ΔΔCq method
(30).
Luciferase assays
TargetScan version 7.2 (targetscan.org) and miRanda (version mirSVR;
microrna.org/microrna/home.do) were
used to predict the target gene of miR-181b-5p. The 3′-UTR region
of S1PR1 and a seven-nucleotide mutated sequence located at
1,019-1,026 bp were cloned into a pmiR plasmid (Thermo Fisher
Scientific, Inc.) to generate the recombinant constructs,
pmiR-S1PR1-wild-type (WT) and pmiR-S1PR1-mutated (MUT),
respectively. For the luciferase assay, HTR-8/SVneo cells were
seeded onto 12-well plates at a density of 1×105
cells/well and co-transfected with 1 µg pmiR-S1PR1-WT or
pmiR-S1PR1-MUT and 1 µg miR-181b-5p mimics or mimics control using
Lipofectamine 2000 (Thermo Fisher Scientific, Inc.). Cells were
cultured at 37°C and harvested after 24 h and Renilla and
firefly luciferase activity was analyzed using a Dual-luciferase
Reporter assay system (Promega Corporation). Renilla
luciferase activity was normalized to firefly luciferase
activity.
Western blotting
Total protein was extracted from cells and tissue
using RIPA lysis buffer (Beyotime Institute of Biotechnology)
supplemented with protease inhibitors (Beyotime Institute of
Biotechnology). A BCA kit (Thermo Fisher Scientific, Inc.) was used
to detect the protein concentration. Protein samples (30 µg/sample)
were separated via 10% SDS-PAGE and then transferred to PVDF
membranes. After blocking with 5% non-fat dry milk at room
temperature for 1 h, the membranes were incubated overnight (4°C)
with primary antibodies: S1PR1 (1:1,000; cat. no. ab77076; Abcam),
matrix metalloproteinase (MMP)-2 (1:1,000; cat. no. AF5330;
Affinity Biosciences), MMP-9 (1:1,000; cat. no. AF5228; Affinity
Biosciences), GAPDH (1:10,000; cat. no. ab9485; Abcam) and β-actin
(1:5,000; cat. no. 60008-1-Ig; ProteinTech Group, Inc.). After
three washes with TBS containing 0.1% Tween-20, the membranes were
incubated with horseradish peroxidase-conjugated secondary
antibodies (goat anti-rabbit; 1:5,000; cat. no. ab6721; Abcam) for
1 h at room temperature. Specific immunoreactive protein bands were
visualized using an enhanced chemiluminescence protein detection
kit (EMD Millipore). The quantification of the western blotting
results was performed using ImageJ software version 1.8.0 (National
Institutes of Health).
Statistical analysis
All data were analyzed using SPSS 17.0 (SPSS, Inc.)
and presented as the mean ± SD based on at least three independent
experiments. Unpaired Student's t-test was used to perform
comparisons of two groups, and Dunnett's test was used following
one-way ANOVA to assess multiple comparisons. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-181b-5p is upregulated in low
invasive event samples and downregulated in highly invasive event
samples
To explore the function of miR-181b-5p in the events
caused by abnormal trophoblast invasion, plasma and tissue samples
were collected from multiple conditions with abnormal invasion
capacity, including SA (impaired trophoblast invasiveness), PE
(impaired trophoblast invasiveness) and CH (excessive
invasiveness). The miR-181b-5p expression of the samples was
detected using RT-qPCR. These results demonstrated that circulating
miR-181b-5p was significantly upregulated in the SA and PE groups,
but downregulated in the CH group, compared with the respective
controls (Fig. 1A). The results
using tissue samples were consistent with the plasma samples
(Fig. 1B). These findings
suggested the potential importance of miR-181b-5p in the
pathogenesis of these events caused by abnormal trophoblast
invasion.
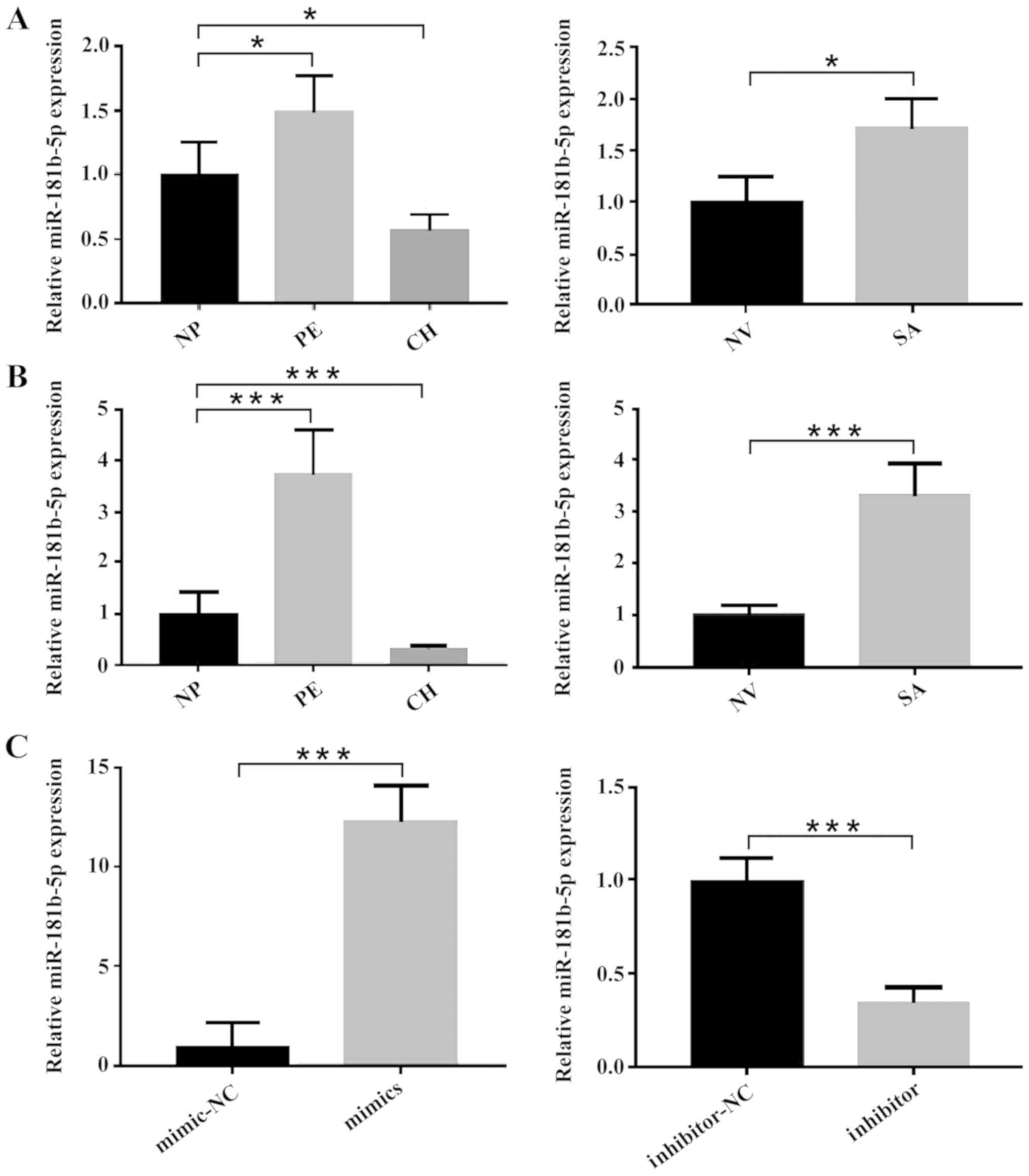 | Figure 1.Expression levels of miR-181b-5p
change with the invasiveness of pregnancy-related events.
Differential miR-181b-5p expression in (A) plasma samples and (B)
tissues samples of NP, PE, CH, NV, and SA were determined by
RT-qPCR. (C) Overexpression and inhibition of miR-181b-5p in
HTR-8/SVneo cells was demonstrated by RT-qPCR. The results are
presented as the mean ± SD of at least three experimental repeats.
*P<0.05, ***P<0.001. NP, normal full-term pregnancy; PE,
preeclampsia; CH, choriocarcinoma; NV, normal early pregnancy; SA,
spontaneous abortion; miR, microRNA; RT-qPCR, reverse
transcription-quantitative PCR. |
To further evaluate the biological functions of
miR-181b-5p in events with abnormal trophoblast invasion, up- or
downregulated miR-181b-5p expression levels were induced in
HTR-8/SVneo cells by transfection with miR-181b-5p mimics or
inhibitors, respectively (Fig.
1C).
miR-181b-5p reduces migration and
invasion of HTR-8/SVneo cells
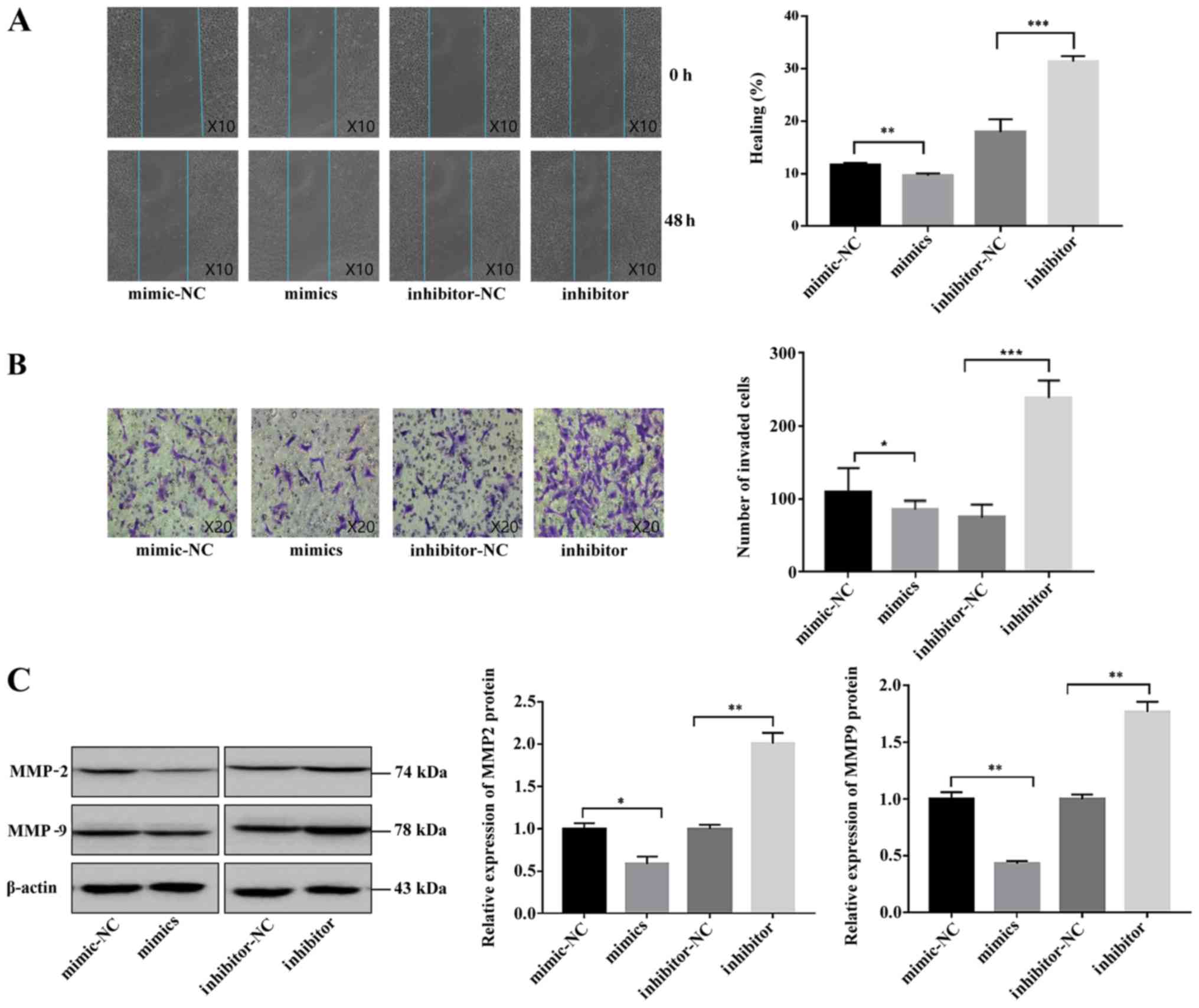
Wound healing and Transwell assays were conducted to
explore the roles of miR-181b-5p in trophoblast cell migration and
invasion. As shown in Fig. 2A,
overexpression of miR-181b-5p suppressed HTR-8/SVneo cell
migration, while downregulation of miR-181b-5p significantly
promoted HTR-8/SVneo cell migration. Similar results were observed
in the Transwell assay experiments (Fig. 2B); as hypothesized, miR-181b-5p
overexpression inhibited, and knockdown of miR-181b-5p
significantly increased, the invasion of HTR-8/SVneo cells.
Furthermore, to investigate the effect of miR-181b-5p on the
invasion-related enzymes, MMP-2 and MMP-9, in HTR-8/SVneo cells,
western blotting was performed. At different stages of embryo
implantation, trophoblast cells secrete different quantities and
types of MMP, among which MMP-2 and MMP-9 are the most important
enzymes involved in trophoblastic invasion (31). It was revealed that the expression
of MMP-2 and MMP-9 was decreased by miR-181b-5p mimics treatment,
whilst MMP-2 and MMP-9 expression was increased by miR-181b-5p
inhibitor treatment (Fig. 2C).
Collectively, these results demonstrated that miR-181b-5p could
suppress the migratory and invasive abilities of trophoblasts.
S1PR1 is a direct target of
miR-181b-5p
To investigate the molecular mechanisms via which
miR-181b-5p suppressed the invasion and migration of trophoblasts,
the potential targets of miR-181b-5p were searched for using the
miRNA target prediction tools TargetScan and miRanda. miR-181b-5p
was found to target the 3′-UTR region of three genes: S1PR1, C-X-C
motif chemokine ligand 3 and protein phosphatase 3 regulatory
subunit B, α. Further RT-qPCR analysis suggested that, of these
three, S1PR1 mRNA was most markedly affected by miR-181b-5p
overexpression in trophoblast cells (data not shown); therefore,
S1PR1 was selected for further experimentation. Fig. 3A showed the S1PR1 sequence
complementary to miR-181b-5p. In order to confirm that S1PR1 might
be a direct target of miR-181b-5p, luciferase assays were performed
by co-transfection of pmiR-S1PR1 luciferase reporter plasmids and
miR-181b-5p mimics. The results showed that transfection with
miR-181b-5p mimics significantly reduced luciferase activity of
S1PR1-WT, but had no effect on the S1PR1-MUT (Fig. 3B). RT-qPCR and western blotting
further validated that S1PR1 was a target of miR-181b-5p: S1PR1
mRNA and protein expression in trophoblasts were both decreased
after administration of miR-181b-5p mimics and upregulated after
transfection with miR-181b-5p inhibitor (Fig. 3C and D). Taken together, these
results strongly indicated that miR-181b-5p directly and
specifically binds to the target site in the 3′-UTR of S1PR1.
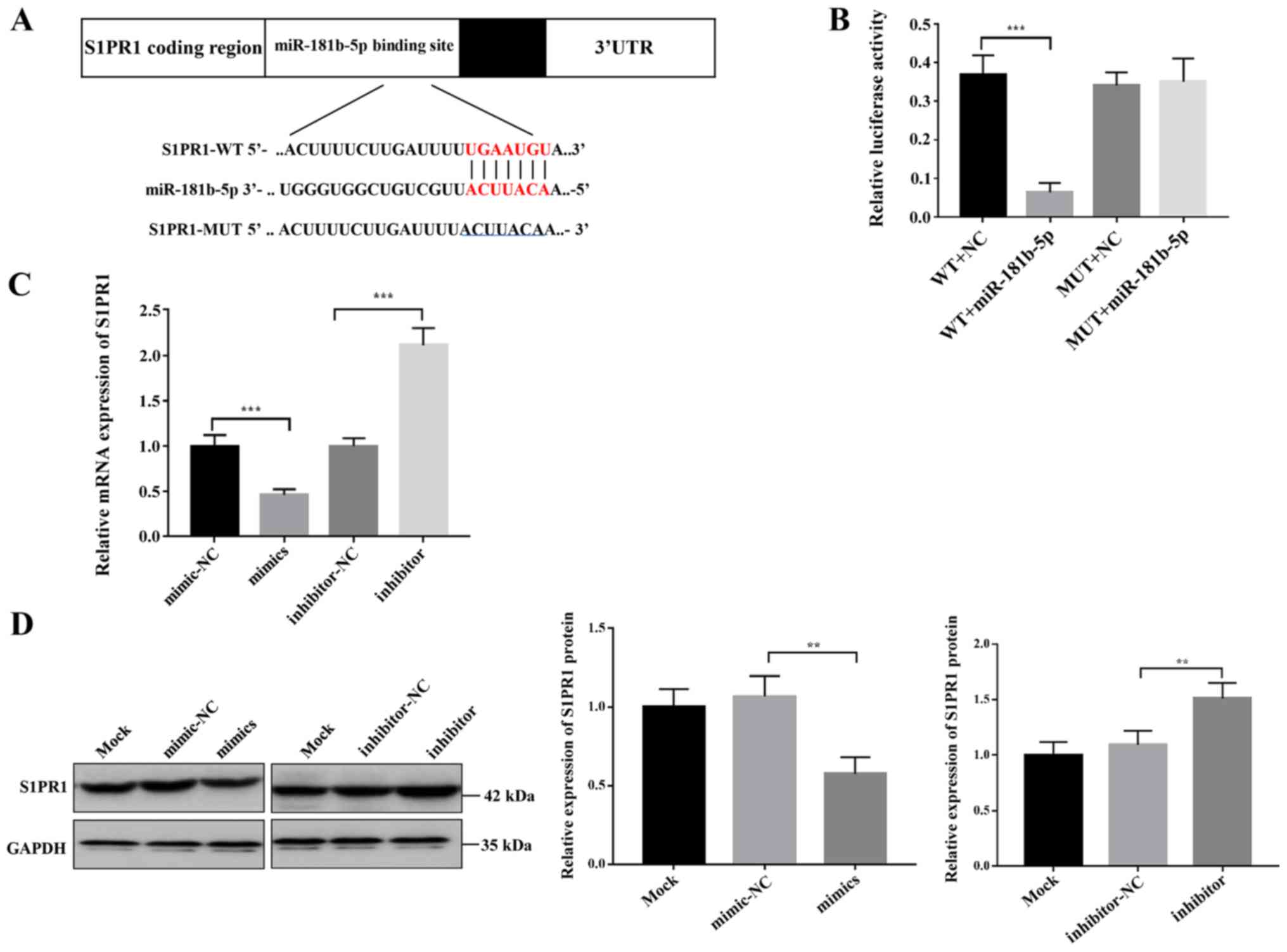 | Figure 3.S1PR1 is a direct target of
miR-181b-5p. (A) Sequence alignment via bioinformatics analysis
between miR-181b-5p and its putative binding site in the S1PR1
3′-UTR. (B) Luciferase activities of reporter vectors containing
either the WT or the MUT S1PR1 3′-UTR were measured in the presence
of miR-181b-5p mimic or mimic-NC. ***P<0.001. (C) Relative S1PR1
mRNA expression in HTR-8/SVneo cells after being transfected with
miR-181b-5p mimics, mimic-NC, miR-181b-5p inhibitor or
inhibitor-NC. (D) S1PR1 relative protein expression in HTR-8/SVneo
cells after being transfected with miR-181b-5p mimics, mimic-NC,
miR-181b-5p inhibitor or inhibitor-NC. The results are presented as
the mean ± SD of at least three experimental repeats. **P<0.01,
***P<0.001. WT, wild-type; MUT, mutated; NC, negative control;
UTR, untranslated region; miR, microRNA; S1PR1,
sphingosine-1-phosphate receptor 1. |
Expression of S1PR1 in abnormal
trophoblast invasion-related events
To evaluate whether the expression of S1PR1 is
associated with miR-181b-5p expression in diseased states, mRNA and
protein expression levels of S1PR1 were examined by RT-qPCR and
western blotting in tissues from multiple conditions with abnormal
trophoblast invasion. As compared with normal placenta or normal
villus tissues, S1PR1 mRNA and protein expression was found to be
significantly downregulated in the PE and SA groups, but
upregulated in the CH group (Fig. 4A
and B). This suggested that miR-181b-5p expression was
negatively associated with S1PR1 expression in tissue samples from
normal pregnancies and abnormal trophoblast invasion-related
events.
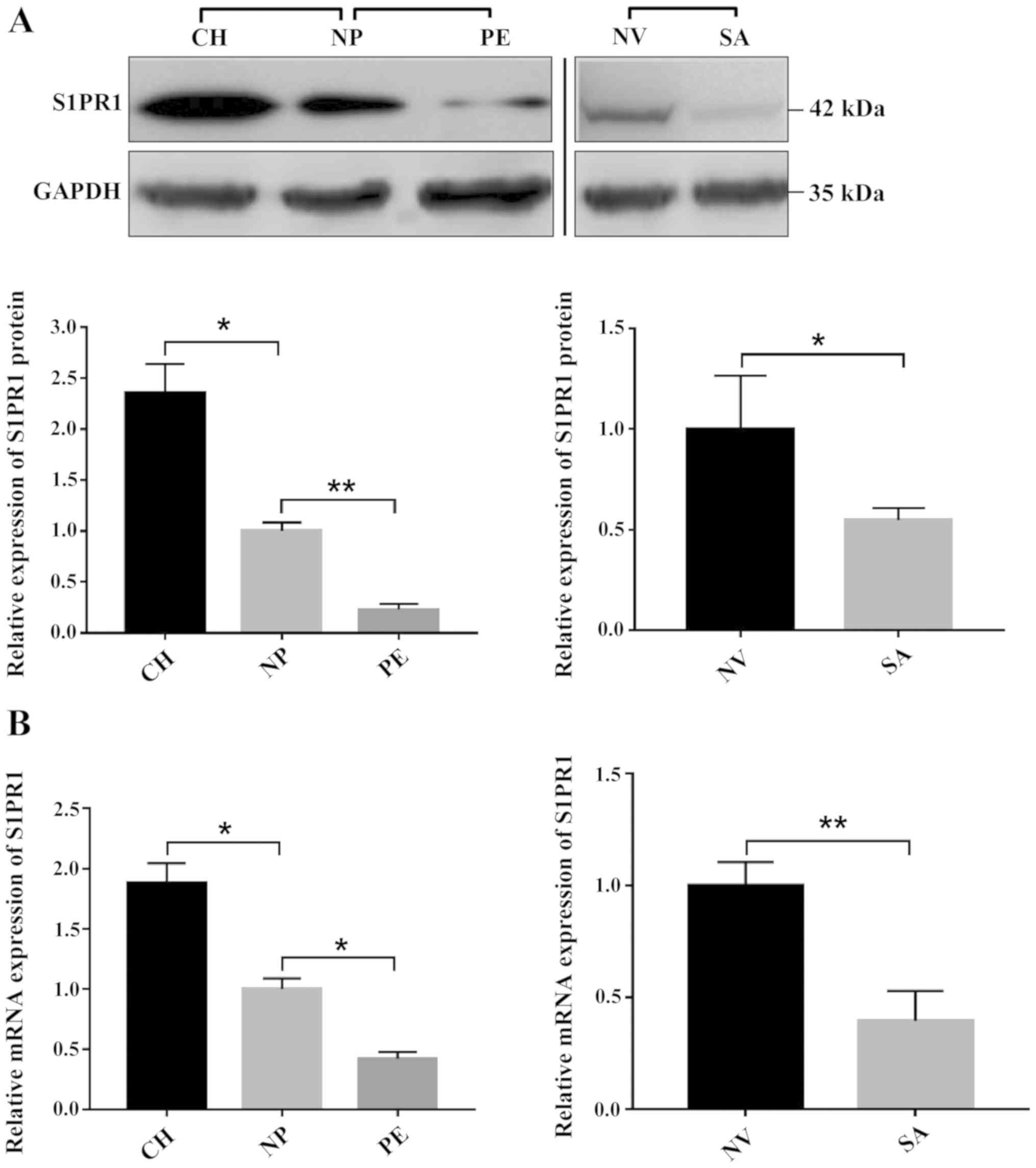 | Figure 4.mRNA and protein expression levels of
S1PR1 in tissue samples. (A) Composite image shows gels with tracks
from different exposures. Compared with NP, S1PR1 protein levels in
the PE group were found to be significantly downregulated, but
those in the CH group were upregulated. Compared with NV tissues,
S1PR1 protein levels in the SA group were found to be significantly
downregulated. Bands were semi-quantitatively compared between
groups. (B) Relative mRNA expression of S1PR1 in the peripheral
blood samples of normal pregnant women and women with abnormal
trophoblast invasion-related events. The results are presented as
the mean ± SD of at least three experimental repeats. *P<0.05,
**P<0.01. NP, normal full-term pregnancy; PE, preeclampsia; CH,
choriocarcinoma; NV, normal early pregnancy; SA, spontaneous
abortion; S1PR1, sphingosine-1-phosphate receptor 1. |
Overexpression of S1PR1 attenuates the
effects induced by miR-181b-5p in trophoblast cells
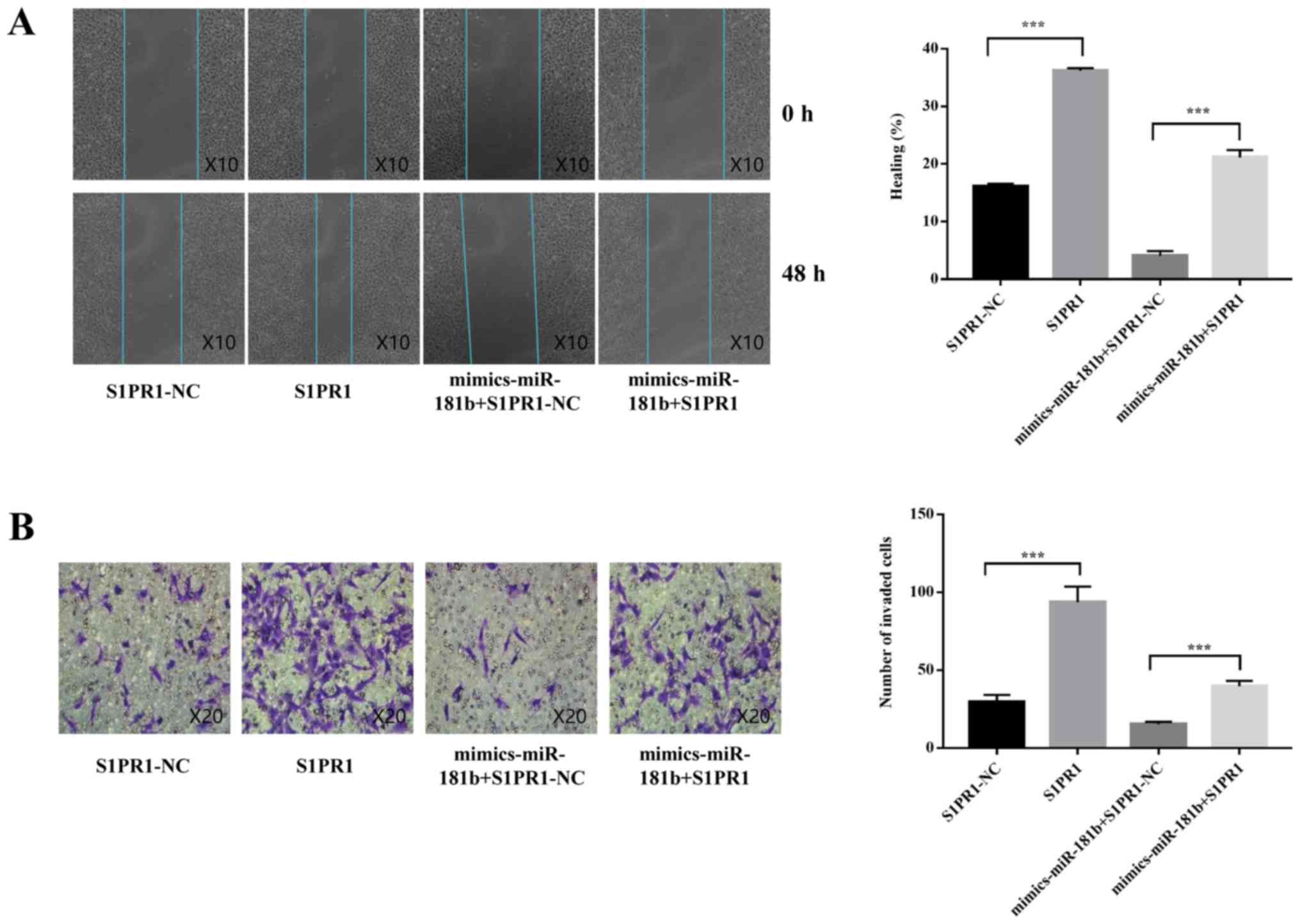
To determine whether the suppressive effect of
miR-181b-5p on trophoblast invasion was mediated by S1PR1
repression, S1PR1 was overexpressed in HTR-8/SVneo cells. As shown
in Fig. S1, the expression of
S1PR1 mRNA was significantly upregulated after plasmid
transfection. Subsequently, miR-181b-5p was overexpressed together
with S1PR1 in HTR-8/SVneo cells to perform a rescue experiment. Of
note, the migration and invasion assay results showed that
restoring expression of S1PR1 counteracted the inhibitory effects
of miR-181b-5p overexpression on the migration and invasion of the
trophoblast cells (Fig. 5A and B).
These data indicated that miR-181b-5p suppressed trophoblast
migration and invasion at least partially by inhibiting S1PR1.
Discussion
Trophoblast invasion has been one of the major foci
of placental research due to its pivotal role in the process of
embryo implantation and placentation. A number of similarities of
invasion behavior exist between trophoblasts and cancer cells
(25). However, trophoblast
invasion is transient with rigorous spatial and temporal confines
(32). Multiple studies have
verified that disorders of trophoblast invasion could trigger a
series of pregnancy-related events at various stages of pregnancy:
Abnormalities in vascular invasion of trophoblast in non-arterial
vessels, such as veins and lymphatic vessels, were observed in the
early miscarriage (6); PE was
found to derive from a defect in trophoblast invasion that resulted
in impaired uterine spiral artery remodeling in the second or third
trimester (33,34); and abnormally extensive invasion of
trophoblasts was suggested to lead to premalignant or malignant
conditions, such as CH after pregnancy (10,22).
miRNAs have emerged as crucial regulators that mediate cellular
activities, including invasion (35). Accumulating evidence has suggested
that miRNAs are associated with placental formation and embryonic
development by affecting the biological function of trophoblasts
(36). However, most prior studies
evaluated the association between miRNAs and trophoblasts'
biological function only in a single event. In the present study,
samples were collected from multiple events with abnormal invasion
capacity and miR-181b-5p expression was examined in each group of
events. The results showed that miR-181b-5p was decreased in both
plasma and tissue samples of CH (excessive invasiveness), but
elevated in SA and PE (impaired invasiveness), which suggested an
important role for miR-181b-5p in trophoblast invasion-related
adverse pregnancy outcomes.
miR-181b-5p is a member of the miR-181 family
(miR-181a-d). Functional studies have indicated that miR-181b-5p
may affect malignant cell migration and invasion (23,37).
For example, miR-181b suppresses invasion and migration of glioma
cells by targeting NOVA alternative splicing regulator 1 (23). Furthermore, in the present study,
miR-181b-5p expression was observed in the trophoblast cell line
HTR-8/SVneo, which is similar to EVT cells (27). Wound healing and Transwell assays
demonstrated that pretreatment with miR-181b-5p mimic or inhibitor
suppressed or promoted, respectively, the invasion and migration of
HTR-8/SVneo cells. These results were further verified by examining
invasion-related proteins MMP-2 and MMP-9. Based on these results,
it was indicated that miR-181b-5p may suppress trophoblast invasion
by inhibiting the levels of MMP-2 and MMP-9.
In the present study, S1PR1 was identified as a
candidate target gene of miR-181b-5p through two publicly available
algorithms, TargetScan and miRanda. Luciferase reporter assays
indicated that miR-181b-5p could inhibit S1PR1 expression by
directly binding to the 3′-UTR of S1PR1, which was validated by
RT-qPCR and western blotting. In addition, further rescue
experiments suggested that overexpression of S1PR1 weakened the
inhibitory effects of miR-181b-5p on trophoblast migration and
invasion. S1PR1, also known as endothelial differentiation gene 1,
is a G protein-coupled receptor implicated in the regulation of
vascular remodeling, endothelial barrier integrity and vascular
tone (38). Uterine spiral artery
remodeling and trophoblast invasion are important for increasing
uteroplacental blood flow and lowering maternal vascular resistance
in pregnancy (39). It is widely
accepted that successful placentation depends on the proper
invasion of EVT cells into maternal tissues. Previous studies
demonstrated that S1PR1 is expressed in human EVT cells (40), and that S1P could regulate
migration and invasion of the trophoblast cells via S1P receptors
(41,42). Evidence suggests that S1P/S1PR1
axis activation causes induction of MMP-2 and MMP-9, and thus
enhances invasion and migration (42,43).
In the present study, it was reported for the first time that the
invasion-inhibiting effect of miR-181b-5p is mediated at least
partially by suppressing S1PR1. HTR-8/SVneo cells were selected due
to their similarity to EVT cells. In future studies, the in
vitro functional role of miR-181b-5p should be validated in
primary cells, which are a closer model of the physiological
conditions. In addition, the data presented in the present study
were mainly obtained from human trophoblast cell lines in
vitro; therefore, in future experiments, in vivo models
should be used to explore miR-181b-5p function and its underlying
mechanism in the pathogenesis of multiple abnormal trophoblast
invasion-associated events.
In summary, to the authors' best knowledge, the
present study provides the first evidence that dysregulated
miR-181b-5p contributes to pregnancy-related events by affecting
the migration and invasion of trophoblast cells via direct
targeting of S1PR1.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81501252), the Natural
Science Foundation of Jiangsu Province (grant no. BK20140084) and
the Medical Science and Technology Development Foundation of
Nanjing Department of Health (Grant No. YKK15162).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JM, YZ and LX conceived the study and performed the
experiments. JM drafted the manuscript and performed the analysis.
XH and XZ contributed to drafting the manuscript, interpreting data
and coordinating the study. XZ provided the funding. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Nanjing Maternity and Child Health Care Hospital
affiliated to Nanjing Medical University. All enrolled patients
provided written consent for participation in the study, sample
collection and analyses.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
SA
|
spontaneous abortion
|
|
CH
|
choriocarcinoma
|
|
NV
|
normal early pregnancy
|
|
NP
|
normal full-term pregnancy
|
References
|
1
|
Norwitz ER, Schust DJ and Fisher SJ:
Implantation and the survival of early pregnancy. N Engl J Med.
345:1400–1408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Knöfler M and Pollheimer J: Human
placental trophoblast invasion and differentiation: A particular
focus on Wnt signaling. Front Genet. 4:1902013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Soundararajan R and Rao AJ: Trophoblast
‘pseudo-tumorigenesis’: Significance and contributory factors.
Reprod Biol Endocrinol. 2:152004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Okae H, Toh H, Sato T, Hiura H, Takahashi
S, Shirane K, Kabayama Y, Suyama M, Sasaki H and Arima T:
Derivation of human trophoblast stem cells. Cell Stem Cell.
22:50–63.e56. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Norwitz ER: Defective implantation and
placentation: Laying the blueprint for pregnancy complications.
Reprod Biomed Online. 13:591–9. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Windsperger K, Dekan S, Pils S, Golletz C,
Kunihs V, Fiala C, Kristiansen G, Knöfler M and Pollheimer J:
Extravillous trophoblast invasion of venous as well as lymphatic
vessels is altered in idiopathic, recurrent, spontaneous abortions.
Hum Reprod. 32:1208–1217. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ye Y, Vattai A, Zhang X, Zhu J, Thaler CJ,
Mahner S, Jeschke U and von Schönfeldt V: Role of Plasminogen
activator inhibitor type 1 in pathologies of female reproductive
diseases. Int J Mol Sci. 18:16512017. View Article : Google Scholar
|
|
8
|
Ding J, Cheng Y, Zhang Y, Liao S, Yin T
and Yang J: The miR-27a-3p/USP25 axis participates in the
pathogenesis of recurrent miscarriage by inhibiting trophoblast
migration and invasion. J Cell Physiol. 234:19951–19963. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cudihy D and Lee RV: The pathophysiology
of pre-eclampsia: Current clinical concepts. J Obstet Gynaecol.
29:576–582. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li HW, Tsao SW and Cheung AN: Current
understandings of the molecular genetics of gestational
trophoblastic diseases. Placenta. 23:20–31. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun W, Julie Li YS, Huang HD, Shyy JY and
Chien S: microRNA: A master regulator of cellular processes for
bioengineering systems. Annu Rev Biomed Eng. 12:1–27. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sadovsky Y, Mouillet JF, Ouyang Y, Bayer A
and Coyne CB: The function of trophomirs and other MicroRNAs in the
human placenta. Cold Spring Harb Perspect Med. 5:a0230362015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vaiman D: Genes, epigenetics and miRNA
regulation in the placenta. Placenta. 52:127–133. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cai M, Kolluru GK and Ahmed A: Small
molecule, big prospects: MicroRNA in pregnancy and its
complications. J Pregnancy. 2017:69727322017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang Y, Zhou J, Li MQ, Xu J, Zhang JP and
Jin LP: MicroRNA-184 promotes apoptosis of trophoblast cells via
targeting WIG1 and induces early spontaneous abortion. Cell Death
Dis. 10:2232019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu Y, Wu D, Jiang Z, Zhang Y, Wang S, Ma
Z, Hui B, Wang J, Qian W, Ge Z and Sun L: MiR-616-3p modulates cell
proliferation and migration through targeting tissue factor pathway
inhibitor 2 in preeclampsia. Cell Prolif. 51:e124902018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang YX, Zhao JR, Xu YY, Wu WB and Zhang
HJ: miR-21 is overexpressed in hydatidiform mole tissues and
promotes proliferation, migration, and invasion in choriocarcinoma
cells. Int J Gynecol Cancer. 27:364–374. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ball E, Bulmer JN, Ayis S, Lyall F and
Robson SC: Late sporadic miscarriage is associated with
abnormalities in spiral artery transformation and trophoblast
invasion. J Pathol. 208:535–542. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fisher SJ: Why is placentation abnormal in
preeclampsia? Am J Obstet Gynecol. 213 (Suppl 4):S115–S122. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Goldman-Wohl D and Yagel S: Regulation of
trophoblast invasion: From normal implantation to pre-eclampsia.
Mol Cell Endocrinol. 187:233–238. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hiden U, Bilban M, Knöfler M and Desoye G:
Kisspeptins and the placenta: Regulation of trophoblast invasion.
Rev Endocr Metab Disord. 8:31–39. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhi F, Wang Q, Deng D, Shao N, Wang R, Xue
L, Wang S, Xia X and Yang Y: MiR-181b-5p downregulates NOVA1 to
suppress proliferation, migration and invasion and promote
apoptosis in astrocytoma. PLoS One. 9:e1091242014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang X, Chen X, Meng Q, Jing H, Lu H, Yang
Y, Cai L and Zhao Y: MiR-181b regulates cisplatin chemosensitivity
and metastasis by targeting TGFβR1/Smad signaling pathway in NSCLC.
Sci Rep. 5:176182015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lala PK, Lee BP, Xu G and Chakraborty C:
Human placental trophoblast as an in vitro model for tumor
progression. Can J Physiol Pharmacol. 80:142–149. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Irving JA, Lysiak JJ, Graham CH, Hearn S,
Han VK and Lala PK: Characteristics of trophoblast cells migrating
from first trimester chorionic villus explants and propagated in
culture. Placenta. 16:413–433. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Graham CH, Hawley TS, Hawley RG,
MacDougall JR, Kerbel RS, Khoo N and Lala PK: Establishment and
characterization of first trimester human trophoblast cells with
extended lifespan. Exp Cell Res. 206:204–211. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen C, Ridzon DA, Broomer AJ, Zhou Z, Lee
DH, Nguyen JT, Barbisin M, Xu NL, Mahuvakar VR, Andersen MR, et al:
Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic
Acids Res. 33:e1792005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Varkonyi-Gasic E, Wu R, Wood M, Walton EF
and Hellens RP: Protocol: A highly sensitive RT-PCR method for
detection and quantification of microRNAs. Plant Methods. 3:122007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen L, Nakai M, Belton RJ Jr and Nowak
RA: Expression of extracellular matrix metalloproteinase inducer
and matrix metalloproteinases during mouse embryonic development.
Reproduction. 133:405–414. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
van den Brûle F, Berndt S, Simon N, Coulon
C, Le Goarant J, Munaut C, Noël A, Frankenne F and Foidart JM:
Trophoblast invasion and placentation: Molecular mechanisms and
regulation. Chem Immunol Allergy. 88:163–180. 2005.PubMed/NCBI
|
|
33
|
Kaufmann P, Black S and Huppertz B:
Endovascular trophoblast invasion: Implications for the
pathogenesis of intrauterine growth retardation and preeclampsia.
Biol Reprod. 69:1–7. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Powe CE, Levine RJ and Karumanchi SA:
Preeclampsia, a disease of the maternal endothelium: The role of
antiangiogenic factors and implications for later cardiovascular
disease. Circulation. 123:2856–2869. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kotlabova K, Doucha J and Hromadnikova I:
Placental-specific microRNA in maternal circulation-identification
of appropriate pregnancy-associated microRNAs with diagnostic
potential. J Reprod Immunol. 89:185–191. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Navarro A and Monzo M: MicroRNAs in human
embryonic and cancer stem cells. Yonsei Med J. 51:622–632. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
He L, Yao H, Fan LH, Liu L, Qiu S, Li X,
Gao JP and Hao CQ: MicroRNA-181b expression in prostate cancer
tissues and its influence on the biological behavior of the
prostate cancer cell line PC-3. Genet Mol Res. 12:1012–1021. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kitano T, Usui S, Takashima SI, Inoue O,
Goten C, Nomura A, Yoshioka K, Okajima M, Kaneko S, Takuwa Y and
Takamura M: Sphigosine-1-phosphate receptor 1 promotes neointimal
hyperplasia in a mouse model of carotid artery injury. Biochem
Biophys Res Commun. 511:179–184. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cui Y, Wang W, Dong N, Lou J, Srinivasan
DK, Cheng W, Huang X, Liu M, Fang C, Peng J, et al: Role of corin
in trophoblast invasion and uterine spiral artery remodelling in
pregnancy. Nature. 484:246–250. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Goyal P, Brünnert D, Ehrhardt J, Bredow M,
Piccenini S and Zygmunt M: Cytokine IL-6 secretion by trophoblasts
regulated via sphingosine-1-phosphate receptor 2 involving
Rho/Rho-kinase and Rac1 signaling pathways. Mol Hum Reprod.
19:528–538. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Westwood M, Al-Saghir K, Finn-Sell S, Tan
C, Cowley E, Berneau S, Adlam D and Johnstone ED: Vitamin D
attenuates sphingosine-1-phosphate (S1P)-mediated inhibition of
extravillous trophoblast migration. Placenta. 60:1–8. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yang W, Li Q and Pan Z:
Sphingosine-1-phosphate promotes extravillous trophoblast cell
invasion by activating MEK/ERK/MMP-2 signaling pathways via
S1P/S1PR1 axis activation. PLoS One. 9:e1067252014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kim ES, Kim JS, Kim SG, Hwang S, Lee CH
and Moon A: Sphingosine 1-phosphate regulates matrix
metalloproteinase-9 expression and breast cell invasion through
S1P3-Gαq coupling. J Cell Sci. 124:2220–2230. 2011. View Article : Google Scholar : PubMed/NCBI
|