Introduction
Colon cancer, which occurs in digestive system, is
one of the most common types of malignant cancer and is the second
cause of cancer-associated mortality worldwide, causing ~600,000
cases of mortality per year (1).
It is reported that >1.4 million people are diagnosed with this
disease in the USA, and an additional 134,490 cases are diagnosed
annually in the USA (2). Although
great progress has been achieved in multimodality therapy, patients
with colon cancer at an advanced stage still have poor outcomes.
For instance, it has been reported that >90% of patients with
colon cancer at stage I can survival for ≥5 years, but this is
declined to ~10% in patients with stage IV colon cancer (3). Dysregulated genes serve a crucial
role in the progression of colon cancer, but there are numerous
unidentified genes or known genes whose functions are yet to be
elucidated in colon cancer (4).
Ras-GTPase-activating protein SH3 domain-binding
proteins (G3BPs) are a class of RNA-binding proteins and contain
two homologous proteins (G3BP1 and G3BP2) (5). All G3BPs include a RNA recognition
motif, and have been identified to exert both mRNA-stabilizing and
mRNA-degrading roles (6,7). Although both G3BP1 and G3BP2 are
reported to be involved in carcinogenesis (8), G3BP1 appears to have been studied
more extensively. For example, Zheng et al (9) revealed that elevated expression of
G3BP1 predicted poor prognosis in patients with non-small cell lung
cancer after surgical resection. Furthermore, Winslow et al
(10) compared the effects of
G3BP1 and G3BP2 on breast cancer progression, and the results
indicated that G3BP1 to a larger extent than G3BP2 influenced mRNA
expression levels of peripheral myelin protein 22 (PMP22), a gene
regulated by G3BP1 and that potentially mediates G3BP1 effects on
cell proliferation enhancement. G3BP1 has been demonstrated to be
involved in multiple biological processes, including RNA metabolism
(11), cell proliferation
(12), apoptosis (13), motility and invasion (7). Moreover, G3BP1 upregulation has been
frequently observed in several type of cancer, such as breast
cancer (14), gastric cancer
(15), lung cancer (16) and colon cancer (17). Zhang et al (18) reported that knockdown of G3BPs
(G3BP1 and G3BP2) significantly inhibited the proliferation of
colon cancer HCT116 cells and improved sensitivity to cisplatin, as
well as weakened cell tumorigenesis, suggesting that G3BP1 serves a
crucial role in the progression of colon cancer. However, the
mechanisms via which G3BP1 facilitates colon cancer progression and
the clinical values of G3BP1 require further investigation.
Previous studies have revealed that aberration of
the Wnt/β-catenin signaling pathway significantly contributes to
the malignant transformation of colon cancer (19,20).
Inhibition of Wnt/β-catenin is suggested to be an anti-tumor
mechanism in colon cancer (21).
In the absence of Wnt, the phosphoprotein scaffold Dishevelled3
(Dsh3) promotes the destabilization of β-catenin 1 mRNA (22). The β-catenin protein is controlled
by a protein complex that consists of axis inhibitor (Axin),
adenomatous polyposis coil (APC), casein kinase 1α and glycogen
synthase kinase 3β, and is confined in the cytoplasm (22). However, β-catenin is increased in
cytoplasm and eventually translocates into the nucleus once Wnt
signaling is activated, inducing the transcription of target genes,
including c-myc, a cell proliferation-associated gene (21). G3BP1 has been identified to be a
Dsh-associated protein that is methylated in response to Wnt3a and
negatively regulates β-catenin expression (23). However, the role of Wnt/β-catenin
signaling in G3BP1-mediated colon cancer progression remains
unknown.
The aim of the present study was to investigate
whether G3BP1 promoted the progression of colon cancer by
regulating the Wnt/β-catenin signaling pathway, which could further
reveal the role of G3BP1 in colon cancer and help to identify novel
targets for colon cancer treatment.
Materials and methods
Colon tissue specimens
All primary colon cancer tissues and matched healthy
colon tissues (≥3.5 cm from the cancer tissue) were obtained from
92 patients (age range, 38–73 years; 50 male and 42 female
patients) with colon cancer who received a colectomy between
January 2010 and January 2017 in The Affiliated Hospital of
Southwest Medical University. All patients signed the informed
consent forms and received colectomy as the first treatment method.
The fresh tissues were immediately immersed in liquid nitrogen and
stored at −80°C until further analysis.
Experiments involving human samples were performed
in accordance with the Declaration of Helsinki and were approved by
The Ethical Committee of The Affiliated Hospital of Southwest
Medical University.
Immunohistochemistry (IHC)
For IHC, routine three-step procedures were
performed as previously described (24) with a primary antibody against G3BP1
(cat. no. ab56574; Abcam). Tissue samples were fixed in 10% neutral
formalin for 2 days at room temperature and embedded in paraffin,
after which 4-µm sections were cut and mounted onto slides. Slides
were incubated at 56°C, deparaffinized in xylene and dehydrated in
a graded series of alcohol. Heat-induced antigen retrieval was
carried out with sodium citrate (pH 6.7) in a pressure-cooker for
30 min. Following washing with PBS, the slides were blocked with
PBS + 5% goat serum for 1 h at room temperature, then and incubated
overnight at 4°C with primary antibody against G3BP1 at a dilution
of 1:150. Next, a HRP-conjugated secondary antibody (cat. no.
ab5879; Abcam) was added for 30 min at room temperature. The
staining was examined under a light microscope (Olympus
Corporation; magnification, ×100).
The expression of G3BP1 was determined by three
pathologists in a blinded manner by multiplying the staining extent
and intensity scores. The extent of positively stained cells was
scored as: i) 0 for 0–5%; ii) 1 for 6–25%; iii) 2 for 26–50%; iv) 3
for 51–75%; and v) 4 for 76–100%. The staining intensity was scored
as: i) 0 for negative staining; ii) 1 for weak staining; iii) 2 for
moderate staining; iv) and 3 for strong staining. A total score of
lower or equal to the average score calculated from all samples was
defined as low G3BP1expression, and high expression otherwise.
Cell culture
A normal human intestinal epithelial cell (HIEC)
line, and the colon cancer cell lines SW620, LoVo, RKO, COLO 205
and HCT116 were obtained from the Cell Bank of the Chinese Academy
of Sciences. HIECs were cultured in medium containing 90% DMEM-H
(Thermo Fisher Scientific, Inc.) and 10% FBS (HyClone; Cytiva).
LoVo cells were cultured in F-12K medium (Thermo Fisher Scientific,
Inc.) supplemented with 10% FBS. RKO cells were cultured in 90%
Eagle's Minimum Essential Medium (Thermo Fisher Scientific) and 10%
FBS. SW620, HCT116 and COLO 205 cells were cultured in RPMI-1640
(Thermo Fisher Scientific, Inc.) medium containing 10% FBS. All
cells were maintained in a 5% CO2 atmosphere at
37°C.
Alteration of gene expression
To overexpress G3BP1, colon cancer cells were
infected with the lentivirus vector (Vector-G3BP1; Shanghai
GenePharma Co., Ltd.) using 5 µg/ml polybrene (Hanbio Biotechnology
Co., Ltd.) with an MOI of 10, and the infected cells were incubated
with G401 (100 µg/ml) for 14 days at 37°C to establish the stably
infected cells.
To silence β-catenin expression, colon cancer cells
were infected with the lentivirus vector [short hairpin RNA
(sh)-β-catenin; OriGene Technologies, Inc.] and puromycin (100
µg/ml) was applied to select the stably infected cells at 37°C for
14 days.
To knockdown G3BP1 expression, colon cancer cells
were transfected with the small interfering (si)RNAs targeting
G3BP1 (si-G3BP1; OriGene Technologies, Inc.) using
Lipofectamine® 2000 reagent (Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions. The sequences
were as follows: i) si-G3BP1-1, 5′-CCACACCAAGATTCGCCAT-3′; ii)
si-G3BP1-2, 5′-GGAGATTCATGCAAACGTT-3′; iii) si-G3BP1-3:
5′-GGAGGAGTCTGAAGAAGAA-3′; and iv) si-NC:
5′-CCAAACCTTAGCGCACCAT-3′. Vector-NC, sh-NC and si-NC (OriGene
Technologies, Inc.) were used as the negative controls for
Vector-G3BP1, sh-β-catenin and si-G3BP1, respectively. After 48 h
of transfection/infection, the cells were collected for subsequent
experiments.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from snap-frozen tissues or
cultured cells using TRIzol® reagent (Takara
Biotechnology Co., Ltd.) based on the manufacturer's instructions.
The obtained RNA samples were quantitated using a NanoDrop 2000
system (NanoDrop Technologies; Thermo Fisher Scientific, Inc.) and
identified using 2% agarose gel electrophoresis. Next, the
first-strand cDNA was synthesized with random primers (Beijing
Solarbio Science & Technology Co., Ltd.) using the
High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher
Scientific, Inc.), and RT-qPCR was performed with a
SuperScript® III One-Step RT-PCR system with Platinum
Taq Mix (Thermo Fisher Scientific, Inc.), according to the
manufacturer's instructions. Reaction conditions were as follows:
i) 94°C for 5 min; ii) 40 amplification cycles at 94°C for 30 sec,
57°C for 30 sec and 72°C for 30 sec; and iii) 72°C for 5 min. The
mRNA expression of GAPDH was used as the endogenous control. The
mRNA levels were calculated by using the 2−ΔΔCq method
(25). The following primers were
used: G3BP1 forward, 5′-CAGCCGCGTAGGTTGAATTG-3′ and reverse,
5′-AGAGAGAAGCCCCATCACCT-3′; GAPDH forward,
5′-CCACTAGGCGCTCACTGTTCTC-3′ and reverse,
5′-ACTCCGACCTTCACCTTCCC-3′.
Western blotting
Total proteins extracted from cells and colon
tissues were obtained using RIPA lysis buffer (Beijing Solarbio
Science & Technology Co., Ltd.) according to the manufacturer's
instructions. The nuclear and cytoplasmic protein samples were
isolated from cells using a Nuclear and Cytoplasmic Protein
Extraction kit (Beyotime Institute of Biotechnology) following the
manufacturer's instructions. A total of 30 µg proteins from
different groups were subjected to 10% SDS-PAGE and separated at 85
V for 30 min and 120 V for 90 min, which were then transferred onto
PVDF membranes (Thermo Fisher Scientific, Inc.). After being
blocked with 5% non-fat milk for 1 h at room temperature, the
membranes were incubated with the primary antibodies, including
anti-G3BP1 (cat. no. ab56574; Abcam), Axin (cat. no. ab32197;
Abcam), β-catenin (cat. no. MA1-300; Thermo Fisher Scientific,
Inc.), Frizzled (Frz; cat. no. AF1617; R&D Systems, Inc.), Dsh
(cat. no. TA300981; OriGene Technologies, Inc.), TCF/LEF
transcription factor family (TCF; cat. no. 9383; Cell Signaling
Technology, Inc.), c-myc (cat. no. M4439; Sigma-Aldrich; Merck
KGaA), APC (cat. no. ab15270; Abcam), Lamin B1 (cat. no. ab16048;
Abcam) and GAPDH (cat. no. 60004–1-Ig; Wuhan Sanying Biotechnology)
overnight at 4°C. The following day, the membranes were washed four
times (8 min each time) with TBS-Tween-20 [1% (v/v)] and incubated
with horseradish peroxidase (HRP)-conjugated secondary antibodies
(1:10,000 dilution; cat. nos. SA00001-1 and SA00001-2; Wuhan
Sanying Biotechnology) at room temperature for 1 h. The protein
bands were visualized with using a chemiluminescent HRP substrate
(EMD Millipore). ImageJ software (version 1.48; National Institutes
of Health) was used to assess the relative expression levels of
proteins.
Co-immunoprecipitation (Co-IP)
The interaction between G3BP1 and β-catenin proteins
was assessed using a Co-IP assay. SW620 and RKO cells infected with
Vector-G3BP1 or Vector-negative control (NC) were rinsed with cold
PBS and lysed in IP lysis buffer, followed by centrifugation at
10,000 × g at 4°C for 30 min. Then, 200 µg proteins from each
sample were incubated with Dynabeads® protein G (Thermo
Fisher Scientific, Lnc.) for 1 h at room temprature, and incubated
with 2 µg of anti-G3BP1 (cat. no. ab181150; Abcam) or
anti-β-catenin (cat. no. ab16051; Abcam) antibody overnight at 4°C.
Anti-IgG antibody (cat. no. ab182931) was used as a negative
control. This was followed by incubation with Dynabeads®
protein G for another 1 h at room temperature. Subsequently, the
immunocomplex was washed five times with IP lysis buffer (Thermo
Fisher Scientific, Inc.) and then subjected to western blot
analysis with anti-β-catenin (cat. no. MA1-300; Thermo Fisher
Scientific, Inc.) or anti-G3BP1 (cat. no. ab56574; Abcam)
antibodies at 1:2,000 dilution.
Cell proliferation and apoptosis
detection
SW620 and RKO cells in the logarithmic phase with
lentivirus infection, siRNA transfection or without were seeded
into 96-well plates at 3,000 cells/well density and cultured at
37°C. Cell proliferation was tested using the Cell Counting Kit-8
(CCK-8; Dojindo Molecular Technologies, Inc.) and the absorbance at
the wavelength of 450 nm was measured according to the
manufacturer's instructions after 1, 2, 3, 4 or 5 days of cell
inoculation.
Apoptotic rates (the percentage of early- and
late-apoptotic cells) were measured in SW620 and RKO cells with
lentivirus infection, siRNA transfection or without using the
Annexin V-FITC Apoptosis Detection kit I (BD Biosciences) and
assessed by flow cytometry. The cells were suspended in 1X binding
buffer and incubated with 5 µl Annexin V-FITC and 5 µl PI in the
dark for 10 min. Cell apoptotic rates were detected on a BD
FACSCanto II instrument (BD Biosciences) and analyzed using FlowJo
7.6 software (FlowJo LLC).
Mouse xenografts of human colon cancer
cells
Mouse xenografts assays were used to evaluate the
effect of G3BP1 and β-catenin on cell tumorigenesis, and were
performed as previously reported (26). A total of 30 male nude mice
(weight, 12–14 g; age, 4–6 weeks) were purchased from Experimental
Animal Center of The Fourth Military Medical University. The mice
were fed with common feed and sterile water ad libitum, and
housed in 22±1°C with 55±1% humidity and a 12 h light/dark cycles.
Athymic nude mice assays were conducted in accordance with the
Institutional principles for the concern and use of animals, and
the protocol was approved by the ethical committee of The
Affiliated Hospital of Southwest Medical University.
SW620 and RKO cells were stably infected with
Vector-NC, Vector-G3BP1 and Vector-G3BP1 + sh-β-catenin (MOI, 10).
Then, a total of 5×106 cells were resuspended in PBS and
injected subcutaneously into the flanks of nude mice (n=10 in each
group). The animal health and behavior were monitored every 3 days.
Mice were euthanized via cervical dislocation and the tumors were
removed 28 days after injection. However, the mice were also
sacrificed when the tumor diameter was >1.8 cm. The mice were
considered to be dead when the heart and breathing stopped. The
largest tumor size was ~2 cm3 and the largest tumor
diameter was ~1.2 cm.
Statistical analysis
Data from ≥3 separate experiments are presented as
the mean ± SD. P<0.05 was considered to indicate a statistically
significant difference. Data were analyzed using an unpaired or a
paired (Fig. 1A and B) two-tailed
Student's t-test for two groups, or one way ANOVA followed by
Tukey's post hoc test for multiple groups with SPSS 21.0 software
(IBM Corp.). A χ2 test was used to compare the
differences in age, sex, vascular invasion, lymph node invasion,
differentiation status and TNM stage between patients with G3BP1
high expression or low expression (Table I). The relationship between G3BP1
expression levels and patient overall survival was assessed by
using Kaplan-Meier analysis with log rank tests.
 | Table I.Association between
clinicopathological characteristics and G3BP1 expression in 92
patients with colorectal cancer. |
Table I.
Association between
clinicopathological characteristics and G3BP1 expression in 92
patients with colorectal cancer.
|
|
| G3BP1
expression |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | Total no.
patients | Low (n=63) | High (n=29) | P-value |
|---|
| Age, years |
|
|
| 0.541 |
|
<60 | 55 | 39 | 16 |
|
|
≥60 | 37 | 24 | 13 |
|
| Sex |
|
|
| 0.732 |
|
Male | 50 | 35 | 15 |
|
|
Female | 42 | 28 | 14 |
|
| Vascular
invasion |
|
|
| 0.032 |
| No | 70 | 52 | 18 |
|
|
Yes | 22 | 11 | 11 |
|
| Lymph node
invasion |
|
|
| 0.013 |
|
Absent | 61 | 47 | 14 |
|
|
Present | 31 | 16 | 15 |
|
| Differentiation
status |
|
|
| 0.033 |
|
High | 66 | 42 | 24 |
|
|
Low | 26 | 21 | 5 |
|
| TNM stage |
|
|
| 0.044 |
|
I–II | 72 | 53 | 19 |
|
|
III–IV | 20 | 10 | 10 |
|
Results
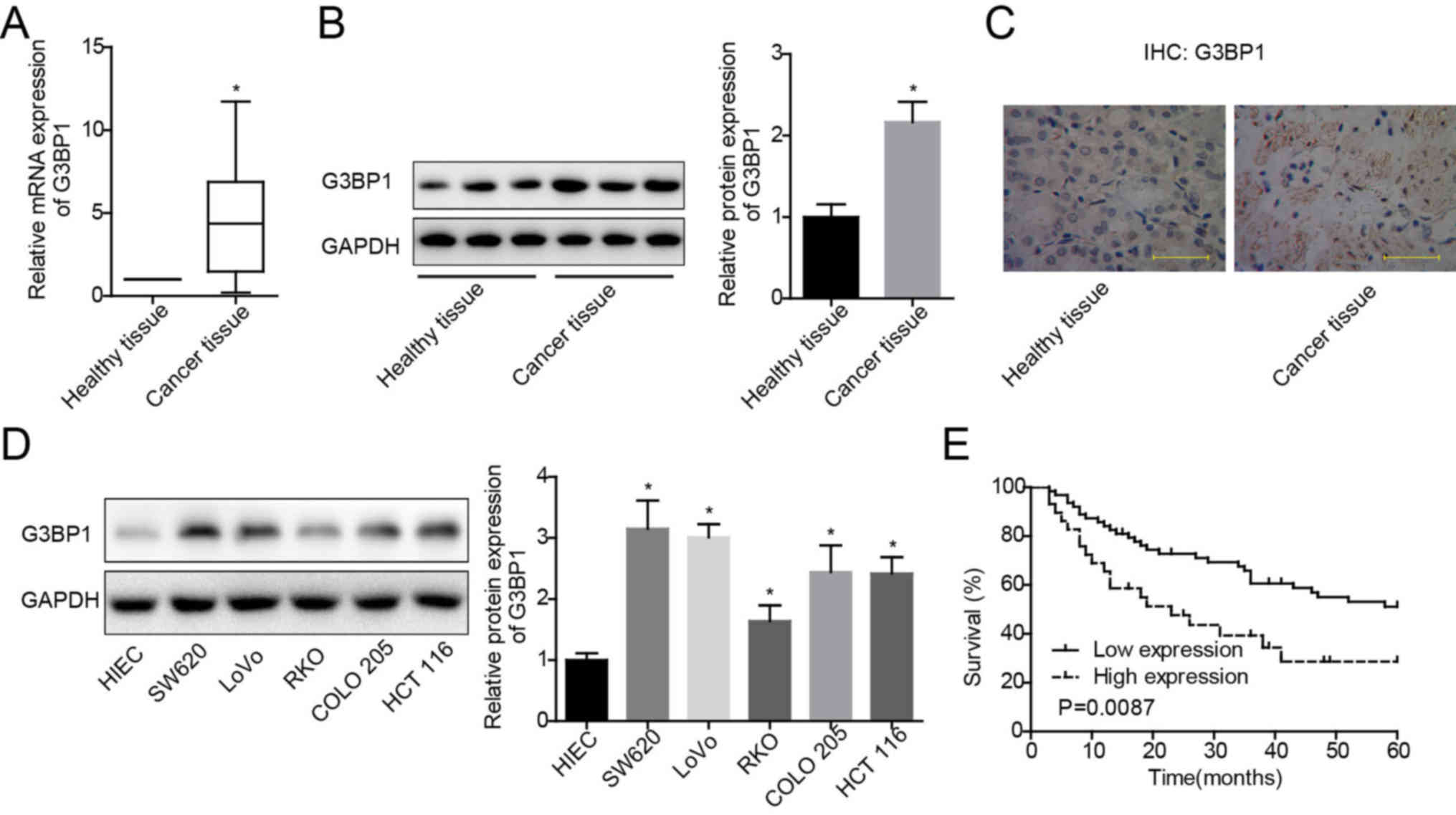
G3BP1 is upregulated in colon cancer
tissue specimens and cells
To further examine the mechanism of G3BP1 in the
carcinogenesis of colon cancer, its expression profile was assessed
in colon cancer tissues and paired para-carcinoma healthy tissues.
Increased expression of G3BP1 at both mRNA (Fig. 1A) and protein (Fig. 1B and C) levels was observed in
colon cancer tissues compared with healthy tissues. In addition,
G3BP1 expression patterns were evaluated in colon cancer cells and
healthy colon cells using western blotting. The results
demonstrated that G3BP1 protein expression in SW620, RKO, LoVo,
COLO 205 and HCT 116 cells was significantly increased compared
with HIECs, with RKO demonstrating the lowest, and SW620 having the
highest expression levels among these colon cancer cell lines
(Fig. 1D). These findings
indicated that G3BP1 expression was upregulated in colon cancer
tissues and cells.
High expression of G3BP1 is closely
associated with malignant characterization and poor prognosis in
colon cancer
Subsequently, the clinical value of G3BP1 expression
in colon cancer was evaluated using Kaplan-Meier curves to
determine the overall survival. In addition, a χ2 test
was used to analyze the association between G3BP1 expression and
the clinicopathologic features in patients with colon cancer.
Patients with high G3BP1 expression (n=29) had shorter overall
survival time compared with patients with low G3BP1 expression
(n=63) (Fig. 1E). In addition,
G3BP1 expression demonstrated a significant positive association
with the incidence rates of vascular invasion and lymph node
invasion, as well as differentiation status and TNM stage (Table I). These results suggested that
G3BP1 may serve as a marker for the diagnosis and prognosis
prediction in patients with colon cancer.
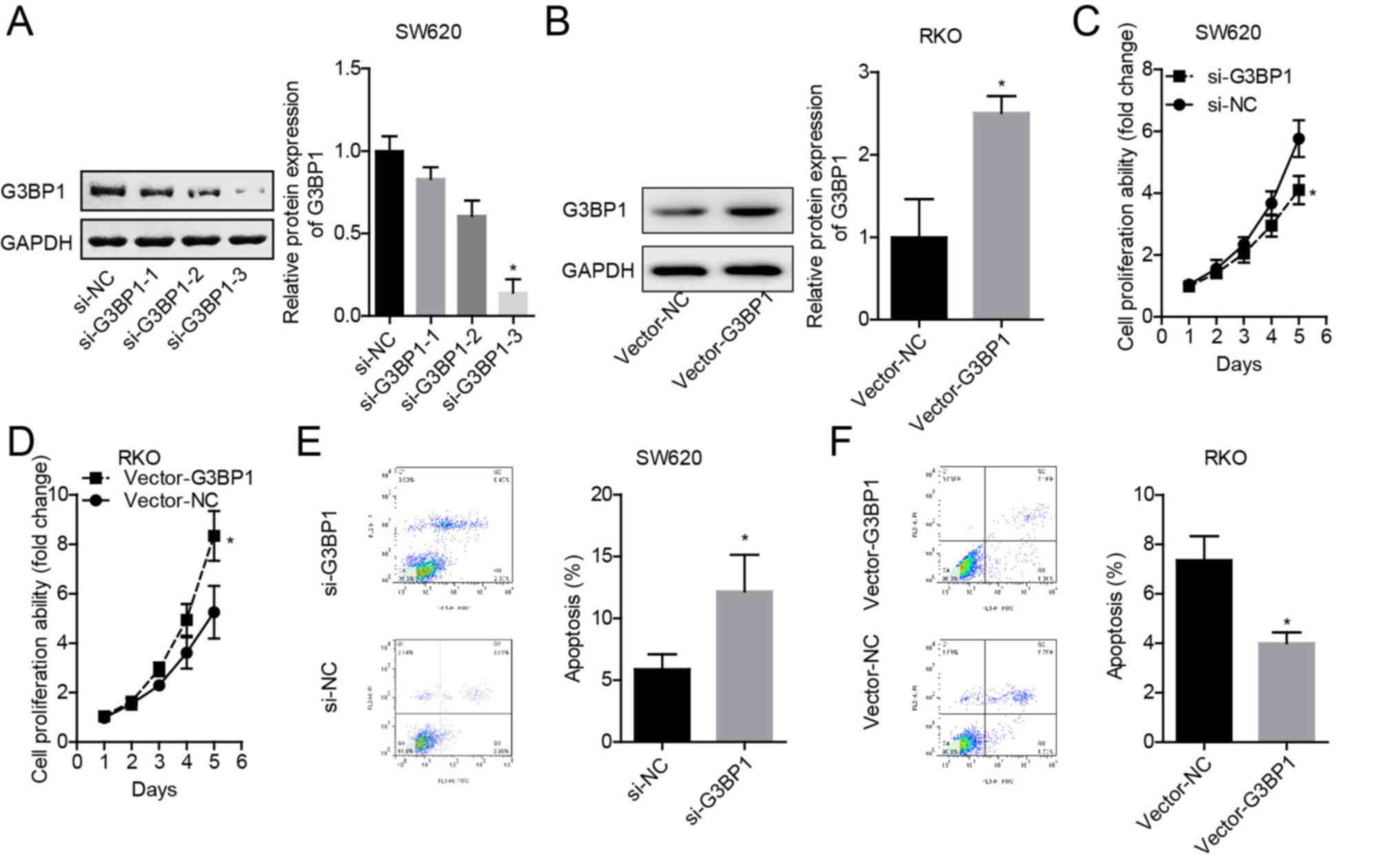
G3BP1 overexpression induces a
malignant phenotype of colon cancer cells
As SW620 cells demonstrated the highest expression
of G3BP1 and RKO cells presented with the lowest, G3BP1 was knocked
down in SW620 cells and overexpressed in RKO cells. Among the three
siRNAs targeting G3BP1 gene, si-3 displayed the highest knockdown
efficiency in SW620 cells (Fig.
2A), and Vector-G3BP1 significantly enhanced G3BP1 expression
in RKO cells (Fig. 2B).
Knockdown of G3BP1 in SW620 cells significantly
inhibited the cell proliferative ability (Fig. 2C), while overexpression of G3BP1 in
RKO cells significantly enhanced proliferation compared with their
corresponding NCs (Fig. 2D). In
addition, knockdown of G3BP1 in SW620 cells induced apoptosis
(Fig. 2E), and overexpression of
G3BP1 in RKO cells decreased apoptosis (Fig. 2F). Thus, these results indicated
that G3BP1 served as an oncogene in colon cancer.
Overexpression of G3BP1 translocates
β-catenin into nucleus of colon cancer cells
To investigate whether Wnt/β-catenin signaling is
involved in G3BP1-mediated colon cancer progression, a series of
assays were conducted to assess the association between
Wnt/β-catenin signaling and G3BP1. The western blotting results
identified that G3BP1 overexpression could significantly increase
the expression levels of β-catenin and c-myc, whereas it had no
notable effect on the expression levels of Frz, Dsh, APC, Axin and
TCF in SW620 (Fig. 3A) and RKO
cells (Fig. 3B). Moreover, G3BP1
overexpression significantly facilitated the translocation of
β-catenin protein from the cytoplasm to the nucleus (Fig. 3C and D). The co-IP assay results
also demonstrated that G3BP1 could directly or indirectly bind to
β-catenin, and the overexpression of G3BP1 markedly enhanced their
interaction (Fig. 3E and F).
Therefore, it was suggested that G3BP1 overexpression could induce
the activation of β-catenin signaling.
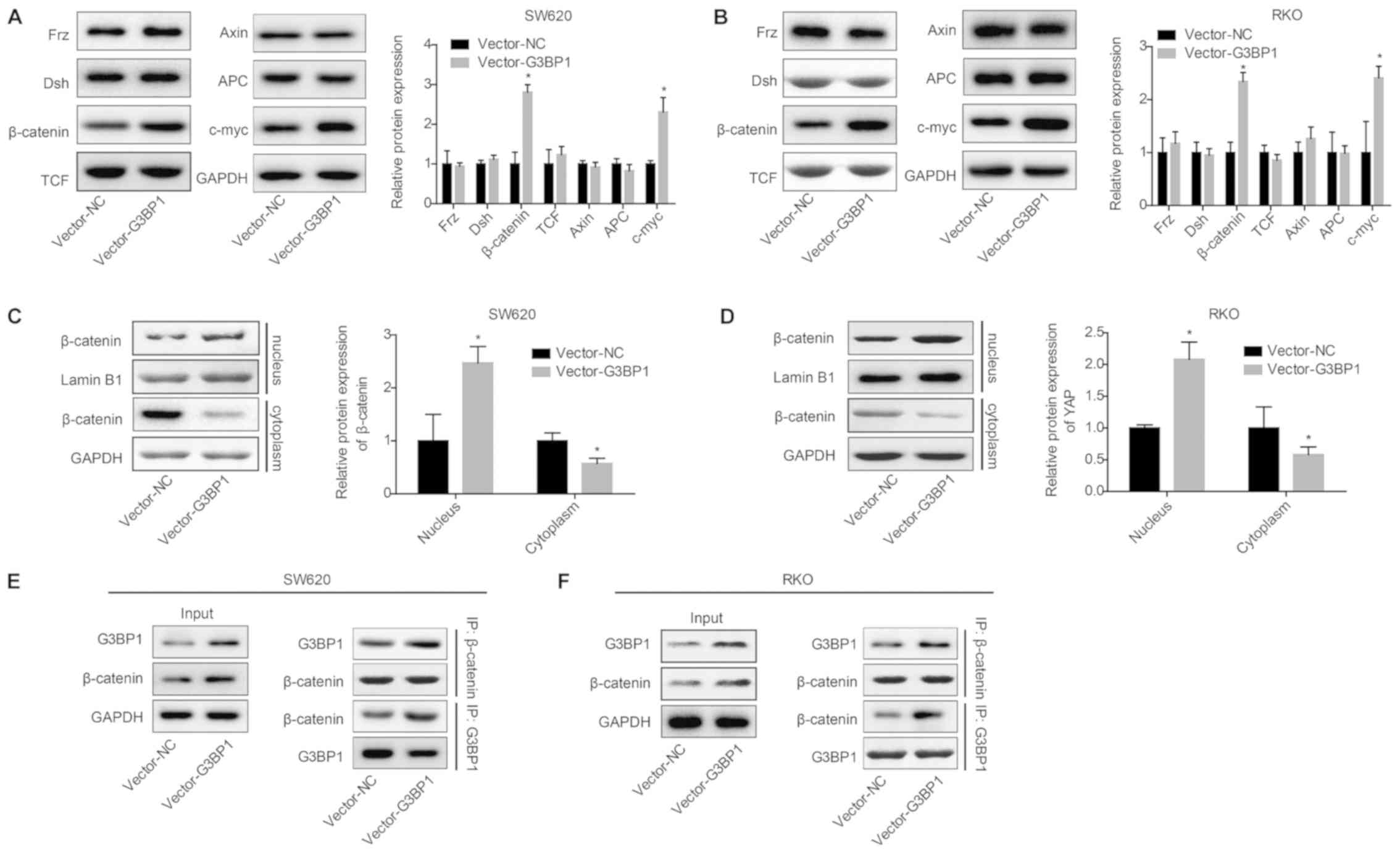 | Figure 3.G3BP1 activates β-catenin in colon
cancer cells. SW620 and RKO cells were transfected with Vector-NC
or Vector-G3BP1 and the following assays were performed. Protein
expression levels of Frz, Dsh, TCF, APC, Axin, c-myc and β-catenin
were detected via western blotting after 48 h of transfection in
(A) SW620 and (B) RKO cells. Cell nuclear and cytoplasm proteins
were extracted for western blot analysis to detect β-catenin levels
in nuclear (Lamin B1 was used as an internal reference for nuclear
protein) and cytoplasmic fractions (GAPDH was used as an internal
reference for cytoplasm protein) in (C) SW620 and (D) RKO cells.
Interaction between G3BP1 and β-catenin was determined using a
co-IP assay in (E) SW620 and (F) RKO cells. Data from three
independent assays are presented as the mean ± SD. *P<0.05 vs.
Vector-NC group. co-IP, co-immunoprecipitation; G3BP1,
Ras-GTPase-activating protein SH3 domain-binding protein 1; NC,
negative control; Frz, Frizzled; Dsh, Dishevelled; TCF,
transcription factor; Axin, axis inhibitor; APC, adenomatous
polyposis coil. |
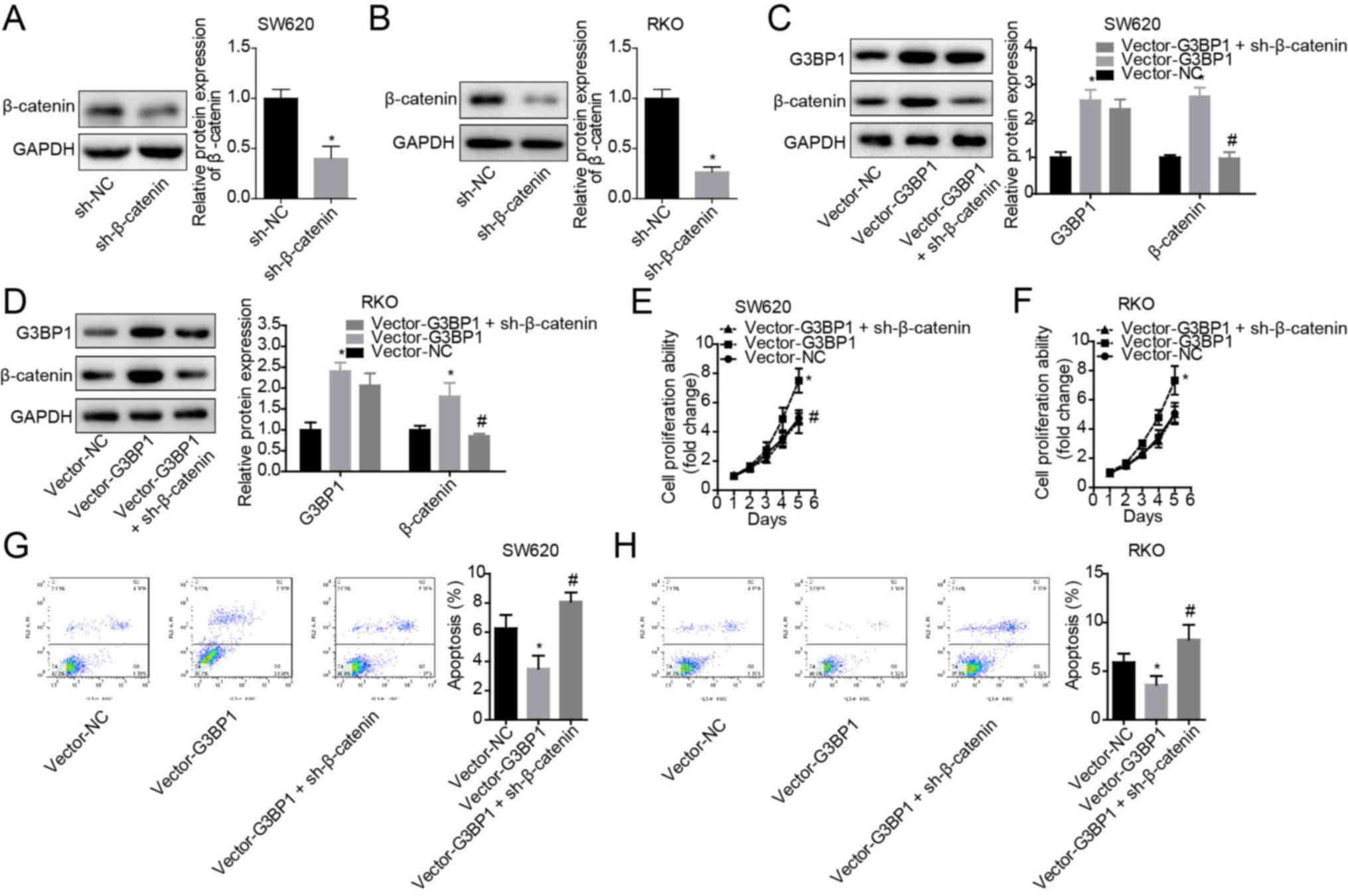
G3BP1 facilitates the progression of
colon cancer by upregulating β-catenin
Finally, the role of β-catenin in G3BP1-mediated
cell proliferation enhancement and apoptosis suppression in colon
cancer was assessed. The expression of β-catenin was significantly
decreased following cell transfection with si-β-catenin in both
SW620 and RKO cells compared with the si-NC group (Fig. 4A and B). It was found that
overexpression of G3BP1 significantly enhanced β-catenin
expression, but this tendency was significantly rescued by
sh-β-catenin infection in SW620 and RKO cells (Fig. 4C and D). Moreover, knockdown of
β-catenin weakened the role of G3BP1 in cell proliferation
enhancement (Fig. 4E and F) and
cell apoptosis inhibition (Fig. 4G and
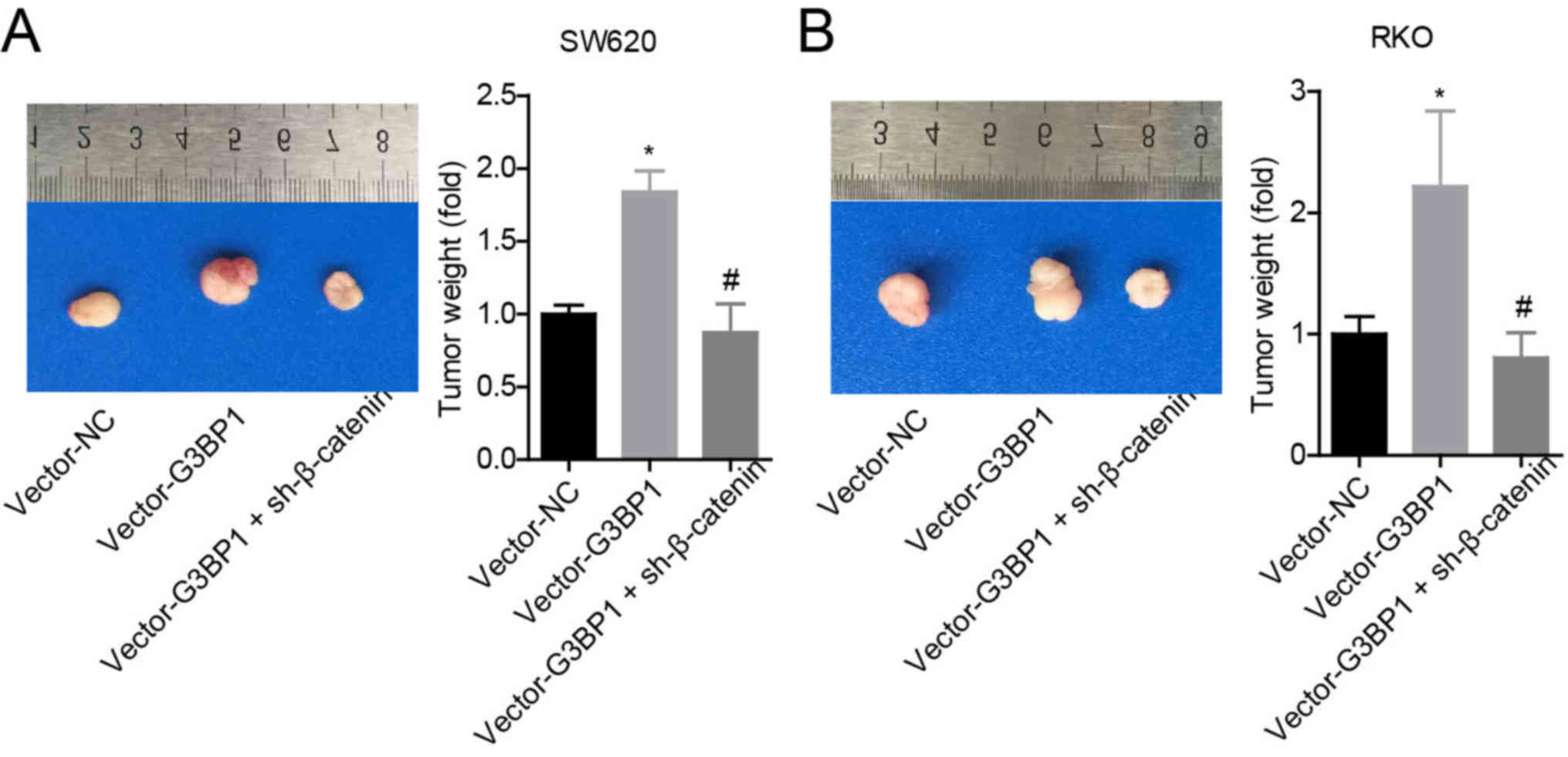
H). The results also demonstrated that G3BP1 overexpression
significantly enhanced the tumorigenesis of SW620 and RKO cells,
whereas this effect was abolished when β-catenin was stably knocked
down (Fig. 5A and B).
Collectively, the results indicated that G3BP1 facilitated the
progression of colon cancer in a β-catenin-dependent manner.
Discussion
Identification and investigation of dysregulated
genes are important to understand the pathogenesis of cancer types
and for evaluating novel therapeutic targets. Although G3BP1 has
been reported to be upregulated in colon cancer, and it has been
shown that knockdown of G3BP1 significantly inhibits cell
proliferation and improves the chemosensitivity of HCT116 cells
(18), the clinical significance
and mechanisms of G3BP1 in colon cancer development remain largely
unknown.
In the present study, it was identified that G3BP1
was significantly upregulated in colon cancer tissues and cells,
and that high expression of G3BP1 was closely associated with
vascular invasion, lymph node invasion, the differentiation status
and TNM stage of colon cancer, as well as predicted a poor
prognosis. These findings indicated a crucial value of G3BP1 in
predicting the clinic process and outcome of patients with colon
cancer. In line with the current results, Dou et al
(27) revealed that G3BP1 was
upregulated in hepatocellular carcinoma (HCC), and its high
expression was significantly associated with the poor prognosis of
patients with HCC. Furthermore, Min et al (15) observed that G3BP1 was upregulated
in gastric cancer at the protein level, and that the G3BP1
expression pattern was positively correlated with tumor size, TNM
stage, vascular invasion and lymph node metastasis, but negatively
associated with the overall survival of patients. However, it has
also been reported that G3BP1 expression is decreased in gastric
cancer, and is significantly associated with age and disease stage,
but demonstrates no significant correlation with cancer metastasis
in gastric cancer (28).
Therefore, different sources of tissue samples may influence the
varying roles of G3BP1 in carcinogenesis.
G3BP1 is an important marker of stress granules (SG)
and is induced by several stress factors, such as chemotherapy,
heat shock and hypoxia (29). The
formation of SG can protect cells from apoptosis by inhibiting the
stress-responsive MAPK pathways (30). Moreover, G3BP1 serves an important
role in the regulation of a variety of growth-related signaling
pathways, including NF-κB, Ras and PI3K signaling pathways and the
ubiquitin proteasome system (31–34).
G3BP1 also negatively modulates p53 expression (35), a tumor suppressive gene found in
multiple tumor types, including colon cancer (36). Thus, G3BP1 may be implicated in
carcinogenesis. Previous studies have reported that G3BP1
significantly promotes the progression of several types of cancer.
For instance, Winslow et al (10) revealed that G3BP1 was an important
factor for breast cancer cell proliferation, and knockdown of G3BP1
significantly inhibited cell proliferation by upregulating PMP22.
In addition, knockdown of G3BP1 prevents SG formation and tumor
invasion, as well as lung metastasis in mouse sarcoma xenografts
models (37). It has also been
shown that G3BP1 knockdown significantly inhibits the metastasis
capacity of HCC cells in vitro and in vivo (27). In the present study, it was
identified that G3BP1 overexpression could significantly enhanced
the proliferation and tumorigenesis of colon cancer cells and
decreased cell apoptosis, indicating that G3BP1 functioned as an
oncogene in colon cancer progression.
While it has been reported that the Wnt/β-catenin
signaling pathway serves an important role in the progression of
colon cancer (19,20), the mechanism underlying its
hyperactivation is not fully understood. In the current study, it
was demonstrated that the increased expression levels of β-catenin
and c-myc proteins, together with the nuclear accumulation of
β-catenin protein, could be induced by G3BP1 overexpression, with
no notable change in the expression levels of Frz, Dsh, TCF, Axin
and APC. In addition, it was identified that the G3BP1 protein
combined with β-catenin protein. These results suggest that G3BP1
can activate β-catenin signaling. However, by contrast, G3BP1 has
been revealed to exert an mRNA-degrading role in regulating
β-catenin in response to Wnt3a in totipotent mouse embryonic F9
cells (23), which may be caused
by the different cell contents. Guitard et al (17) reported that the expression patterns
of G3BPs and c-myc proteins (c-myc1 and c-myc2) were similar in
human head and neck tumors, which further indicates that G3BPs are
involved in c-myc mRNA stabilization, thereby facilitating cell
proliferation (32). Similarly,
the present results suggested that G3BP1 positively regulated the
expression of c-myc protein, and this may be associated with the
stability of c-myc mRNA. It was identified that the enhancements in
cell proliferation and tumorigenesis, as well as the decreases in
cell apoptosis induced by G3BP1 overexpression were significantly
rescued by β-catenin knockdown, suggesting that G3BP1 facilitated
colon cancer via activating β-catenin signaling.
In conclusion, the present study demonstrated that
G3BP1 was upregulated in colon cancer, and the high expression of
G3BP1 predicted malignant clinical process and poor outcome in
patients with colon cancer. Mechanically, overexpression of G3BP1
promoted the progression of colon cancer by activating β-catenin
signaling. Collectively, the present study provides a novel
understanding of the roles of G3BP1 in colon cancer
progression.
Acknowledgements
Not applicable.
Funding
The current study was supported by a research grant
for Doctors of the Affiliated Hospital of Southwest Medical
University and Luzhou People's Government-Southwest Medical
University Cooperative Scientific Research Project (grant no.
2019LZXNDJ26).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YL and JW conceived the idea of the study and
performed the experiments, as well as wrote the manuscript. SZ and
JL performed the data analyses. WD performed parts of the data
analyses. YL revised the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
Experiments involving human samples were performed
in accordance with the Helsinki Declaration and were approved by
the ethical committee of The Affiliated Hospital of Southwest
Medical University. All patients signed the informed consent.
Athymic nude mice assays were performed in accordance with the
Institutional principles for the concern and use of animals, and
the protocol was approved by the ethical committee of The
Affiliated Hospital of Southwest Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA CAncer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Miller KD, Siegel RL, Lin CC, Mariotto AB,
Kramer JL, Rowland JH, Stein KD, Alteri R and Jemal A: Cancer
treatment and survivorship statistics, 2016. CA Cancer J Clin.
66:271–289. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pothuraju R, Rachagani S, Krishn SR,
Chaudhary S, Nimmakayala RK, Siddiqui JA, Ganguly K, Lakshmanan I,
Cox JL, Mallya K, et al: Molecular implications of MUC5AC-CD44 axis
in colorectal cancer progression and chemoresistance. Mol Cancer.
19:372020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Briggs SD, Bryant SS, Jove R, Sanderson SD
and Smithgall TE: The Ras GTPase-activating protein (GAP) is an SH3
domain-binding protein and substrate for the Src-related tyrosine
kinase, Hck. J Biol Chem. 270:14718–14724. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Atlas R, Behar L, Elliott E and Ginzburg
I: The insulin-like growth factor mRNA binding-protein IMP-1 and
the Ras-regulatory protein G3BP associate with tau mRNA and HuD
protein in differentiated P19 neuronal cells. J Neurochem.
89:613–626. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Taniuchi K, Nishimori I and Hollingsworth
MA: The N-terminal domain of G3BP enhances cell motility and
invasion by posttranscriptional regulation of BART. Mol Cancer Res.
9:856–866. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
French J, Stirling R, Walsh M and Kennedy
HD: The expression of Ras-GTPase activating protein SH3
domain-binding proteins, G3BPs, in human breast cancers. Histochem
J. 34:223–231. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zheng H, Zhan Y, Zhang Y, Liu S, Lu J,
Yang Y, Wen Q and Fan S: Elevated expression of G3BP1 associates
with YB1 and p-AKT and predicts poor prognosis in nonsmall cell
lung cancer patients after surgical resection. Cancer Med.
8:6894–6903. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Winslow S, Leandersson K and Larsson C:
Regulation of PMP22 mRNA by G3BP1 affects cell proliferation in
breast cancer cells. Mol Cancer. 12:1562013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Irvine K, Stirling R, Hume D and Kennedy
D: Rasputin, more promiscuous than ever: A review of G3BP. Int J
Dev Biol. 48:1065–1077. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kociok N, Esser P, Unfried K, Parker F,
Schraermeyer U, Grisanti S, Toqué B and Heimann K: Upregulation of
the RAS-GTPase activating protein (GAP)-binding protein (G3BP) in
proliferating RPE cells. J Cell Biochem. 74:194–201. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Oi N, Yuan J, Malakhova M, Luo K, Li Y,
Ryu J, Zhang L, Bode AM, Xu Z, Li Y, et al: Resveratrol induces
apoptosis by directly targeting Ras-GTPase-activating protein SH3
domain-binding protein 1. Oncogene. 34:2660–2671. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang H, Ma Y, Zhang S, Liu H, He H, Li N,
Gong Y, Zhao S, Jiang JD and Shao RG: Involvement of Ras
GTPase-activating protein SH3 domain-binding protein 1 in the
epithelial-to-mesenchymal transition-induced metastasis of breast
cancer cells via the Smad signaling pathway. Oncotarget.
6:17039–17053. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Min L, Ruan Y, Shen Z, Jia D, Wang X, Zhao
J, Sun Y and Gu J: Overexpression of Ras-GTPase-activating protein
SH3 domain-binding protein 1 correlates with poor prognosis in
gastric cancer patients. Histopathology. 67:677–688. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shim JH, Su ZY, Chae JI, Kim DJ, Zhu F, Ma
WY, Bode AM, Yang CS and Dong Z: Epigallocatechin gallate
suppresses lung cancer cell growth through Ras-GTPase-activating
protein SH3 domain-binding protein 1. Cancer Prev Res (Phila).
3:670–679. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guitard E, Parker F, Millon R, Abecassis J
and Tocque B: G3BP is overexpressed in human tumors and promotes S
phase entry. Cancer Lett. 162:213–221. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang H, Zhang S, He H, Zhao W, Chen J and
Shao RG: GAP161 targets and downregulates G3BP to suppress cell
growth and potentiate cisplaitin-mediated cytotoxicity to colon
carcinoma HCT116 cells. Cancer Sci. 103:1848–1856. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang J, Li Q, Xue B and He R: MALAT1
inhibits the Wnt/β-catenin signaling pathway in colon cancer cells
and affects cell proliferation and apoptosis. Bosn J Basic Med Sci.
20:357–364. 2019.
|
|
20
|
Shiizaki K, Kido K and Mizuta Y: Insight
into the relationship between aryl-hydrocarbon receptor and
β-catenin in human colon cancer cells. PLoS One. 14:e02246132019.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim JY, Park G, Krishnan M, Ha E and Chun
KS: Selective Wnt/β-catenin Small-molecule Inhibitor CWP232228
Impairs Tumor Growth of Colon Cancer. Anticancer Res. 39:3661–3667.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sebio A, Kahn M and Lenz HJ: The potential
of targeting Wnt/β-catenin in colon cancer. Expert Opin Ther
Targets. 18:611–615. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bikkavilli RK and Malbon CC: Arginine
methylation of G3BP1 in response to Wnt3a regulates β-catenin mRNA.
J Cell Sci. 124:2310–2320. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xin B, He X, Wang J, Cai J, Wei W, Zhang T
and Shen X: Nerve growth factor regulates CD133 function to promote
tumor cell migration and invasion via activating ERK1/2 signaling
in pancreatic cancer. Pancreatology. 16:1005–1014. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Duquet A, Melotti A, Mishra S, Malerba M,
Seth C, Conod A, Ruiz I and Altaba A: A novel genome-wide in vivo
screen for metastatic suppressors in human colon cancer identifies
the positive WNT-TCF pathway modulators TMED3 and SOX12. EMBO Mol
Med. 6:882–901. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dou N, Chen J, Yu S, Gao Y and Li Y: G3BP1
contributes to tumor metastasis via upregulation of Slug expression
in hepatocellular carcinoma. Am J Cancer Res. 6:2641–2650.
2016.PubMed/NCBI
|
|
28
|
Beheshtizadeh M and Moslemi E: Analysis of
G3BP1 and VEZT expression in gastric cancer and their possible
correlation with tumor clinicopathological Factors. J Gastric
Cancer. 17:43–51. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tourriere H, Chebli K, Zekri L, Courselaud
B, Blanchard JM, Bertrand E and Tazi J: The RasGAP-associated
endoribonuclease G3BP assembles stress granules. J Cell Biol.
160:823–831. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Arimoto K, Fukuda H, Imajoh-Ohmi S, Saito
H and Takekawa M: Formation of stress granules inhibits apoptosis
by suppressing stress-responsive MAPK pathways. Nat Cell Biol.
10:1324–1332. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Barnes CJ, Li F, Mandal M, Yang Z, Sahin
AA and Kumar R: Heregulin induces expression, ATPase activity, and
nuclear localization of G3BP, a Ras signaling component, in human
breast tumors. Cancer Res. 62:1251–1255. 2002.PubMed/NCBI
|
|
32
|
Gallouzi IE, Parker F, Chebli K, Maurier
F, Labourier E, Barlat I, Capony JP, Tocque B and Tazi J: A novel
phosphorylation-dependent RNase activity of GAP-SH3 binding
protein: A potential link between signal transduction and RNA
stability. Mol Cell Biol. 18:3956–3965. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Soncini C, Berdo I and Draetta G: Ras-GAP
SH3 domain binding protein (G3BP) is a modulator of USP10, a novel
human ubiquitin specific protease. Oncogene. 20:3869–3879. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang Y, Wernyj RP, Norton DD, Precht P,
Seminario MC and Wange RL: Modulation of specific protein
expression levels by PTEN: Identification of AKAP121, DHFR, G3BP,
Rap1, and RCC1 as potential targets of PTEN. Oncogene.
24:3819–3829. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim MM, Wiederschain D, Kennedy D, Hansen
E and Yuan ZM: Modulation of p53 and MDM2 activity by novel
interaction with Ras-GAP binding proteins (G3BP). Oncogene.
26:4209–4215. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen G, Zhou T, Li Y, Yu Z and Sun L: p53
target miR-29c-3p suppresses colon cancer cell invasion and
migration through inhibition of PHLDB2. Biochem Biophys Res Commun.
487:90–95. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Somasekharan SP, El-Naggar A, Leprivier G,
Cheng H, Hajee S, Grunewald TG, Zhang F, Ng T, Delattre O,
Evdokimova V, et al: YB-1 regulates stress granule formation and
tumor progression by translationally activating G3BP1. J Cell Biol.
208:913–929. 2015. View Article : Google Scholar : PubMed/NCBI
|