Introduction
Bladder cancer is a common human malignancy
worldwide, with an increasing incidence in recent years (1). Currently, the main treatment of
bladder cancer is surgery, however due to high rates of recurrence
and metastasis, the overall survival rate of patients with bladder
cancer is still unsatisfactory (1–3).
Recently, various oncogenes and tumor suppressors have been
reported to have roles in the development and progression of
bladder cancer (4,5). Understanding the molecular mechanisms
may be beneficial for developing effective therapeutic strategies
for bladder cancer.
Long non-coding RNAs (lncRNAs) are a type of
non-coding transcripts containing >200 nucleotides, they have
been shown to serve important regulatory roles in different types
of cancer via interactions with microRNAs (miRNAs/miRs) or proteins
(6,7). Recently, the dysregulation of lncRNA
TINCR ubiquitin domain containing (TINCR) has been observed in
gastric (8), breast (9) and colon cancer (10), and esophageal squamous cell
carcinoma (11). Moreover, it has
been reported that TINCR levels are significantly increased in
bladder cancer, and that the inhibition of TINCR has the ability to
suppress bladder cancer cell proliferation while also promoting
cell apoptosis (12). miRNAs are
also a type of non-coding transcripts containing only 22–25
nucleotides. They can directly interact with the 3′-untranslated
region (UTR) of their target mRNAs, and thus are important
regulators in gene expression. A large number of studies have
demonstrated that miRNAs participate in different cellular
processes, such as cell proliferation, survival, differentiation,
motility and tumorigenesis (13–15).
By mediating the expression of different target genes, miR-7 serves
different roles in various types of human cancer (16–19).
For instance, miR-7 functions as a tumor suppressor in non-small
cell lung cancer by inhibiting the expression of paired box 6
(16). Conversely, miR-7
contributes to colorectal tumorigenesis by targeting YY1
transcription factor (18). mTOR,
a serine/threonine kinase, belongs to the PI3K-associated family.
Previous studies have demonstrated that mTOR is essential for the
survival, proliferation, migration and invasion of tumor cells
(20–22). Therefore, it has been suggested
that mTOR could be a potential therapeutic target in the treatment
of bladder cancer (20–22).
However, until now, the exact association between
TINCR, miR-7 and mTOR in bladder cancer remains unknown. The
present study explored the regulatory mechanism of TINCR underlying
bladder cancer progression involving miR-7 and mTOR. The present
study demonstrated that the upregulation of TINCR was associated
with an aggressive tumor phenotype and poor prognosis in bladder
cancer, and suggested that TINCR promotes the malignant phenotypes
of bladder cancer by regulating the miR-7/mTOR axis.
Materials and methods
Clinical samples
The present study obtained approval from the Ethics
Committee of Shengli Hospital of Shengli Petroleum Administration
and all patients provided written informed consent. Bladder cancer
tissues and their matched adjacent normal tissues were obtained
from 53 patients during surgical treatment between March 2010 and
March 2012 at Shengli Hospital. TNM staging for bladder cancer was
used in the present study (23).
No patients received chemotherapy or radiotherapy prior to surgery.
The clinical characteristics of the patient cohort are summarized
in Table I.
 | Table I.Association between TINCR expression
and clinicopathological characteristics in bladder cancer. |
Table I.
Association between TINCR expression
and clinicopathological characteristics in bladder cancer.
|
|
| TINCR
expression |
|
|---|
|
|
|
|
|
|---|
| Variables | Cases (n=53) | High (n=23) | Low (n=30) | P-value |
|---|
| Age |
|
|
| 0.775 |
|
<55 | 19 | 9 | 10 |
|
|
≥55 | 34 | 14 | 20 |
|
| Sex |
|
|
| 0.779 |
|
Male | 33 | 15 | 18 |
|
|
Female | 20 | 8 | 12 |
|
| Grade |
|
|
| 0.052 |
| Well
and moderately | 40 | 14 | 26 |
|
|
Poor | 13 | 9 | 4 |
|
| Lymph node
metastasis |
|
|
| 0.009a |
|
Negative | 34 | 10 | 24 |
|
|
Positive | 19 | 13 | 6 |
|
| Distant
metastasis |
|
|
| 0.012a |
|
Present | 5 | 5 | 0 |
|
|
Absent | 48 | 18 | 30 |
|
| TNM stage |
|
|
| 0.002a |
|
I–II | 27 | 6 | 21 |
|
|
III–IV | 26 | 17 | 9 |
|
Cell culture and transfection
The normal human urinary tract epithelial SV-HUC-1
cell line and bladder cancer T24, HT-1376 and 5637 cell lines were
obtained from the American Type Culture Collection. These cell
lines were cultured in DMEM (Thermo Fisher Scientific, Inc.) with
10% FBS (Thermo Fisher Scientific, Inc.) at 37°C with 5%
CO2. Cell transfection was conducted in T24 and 5637
cells with Lipofectamine® 2000 (Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. In
brief, T24 and 5637 cells (2×105 cells/well) were seeded
into 6-well plates, and transfected with 100 nM negative control
(NC) small interfering (si)RNA (cat. no. 12935200; Thermo Fisher
Scientific, Inc.), 100 nM TINCR siRNA (cat. no. AM16708; Thermo
Fisher Scientific, Inc.), 100 nM NC inhibitor (cat. no. 4464076;
Thermo Fisher Scientific, Inc.), 100 nM miR-7 inhibitor (cat. no.
4464084; Thermo Fisher Scientific, Inc.), 100 nM miR-NC mimic (cat.
no. 4464058; Thermo Fisher Scientific, Inc.), 100 nM miR-7 mimic
(cat. no. 4464066; Thermo Fisher Scientific, Inc.), 800 ng blank
pcDNA3.1 vector (cat. no. V79020; Thermo Fisher Scientific, Inc.),
or 800 ng mTOR expression plasmid (Yearth Biotech; www.yearthbio.com). After cell transfection for 48 h,
reverse transcription-quantitative (RT-q)PCR was conducted to
examine gene expression levels.
RT-qPCR
Total RNA was isolated from tissues and cell lines
using TRIzol® Reagent (Thermo Fisher Scientific, Inc.),
which was reverse transcribed into cDNA with a PrimeScript RT
reagent kit (Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocols. Reverse transcription was performed at
16°C for 30 min, 42°C for 30 min and 85°C for 5 min.
SYBR® Premix Ex Taq™ (Takara Biotechnology Co., Ltd.)
was used to examine the expression of lncRNA, miRNA and mRNA. PCR
was initiated at 95°C for 3 min, followed by 40 cycles at 95°C for
15 sec, 60°C for 30 sec; and 72°C for 15 sec. The relative gene
expression was determined using the 2−ΔΔCq method
(24). U6 was used as an internal
reference for miRNA. GAPDH was used as an internal reference for
lncRNA and mRNA. Primer sequences used were as follow: GAPDH
forward, 5′-GCACCGTCAAGGCTGAGAAC-3′ and reverse,
5′-ATGGTGGTGAAGACGCCAGT-3′; TINCR forward,
5′-TGTGGCCCAAACTCAGGGATACAT-3′ and reverse,
5′-AGATGACAGTGGCTGGAGTTGTCA-3′; U6 forward, 5′-CTCGCTTCGGCAGCACA-3′
and reverse, 5′-AACGCTTCACGAATTTGCGT-3′; and mTOR forward,
5′-ATGCTTGGAACCGGACCTG-3′ and reverse,
5′-TCTTGACTCATCTCTCGGAGTT-3′. Primers for miR-7 (cat. no.
HmiRQP0785) were obtained from FulenGen, Co., Ltd.
Cell proliferation, migration and
invasion assays
Transfected cells (5,000 cells/well) were seeded in
96-well plates, and a Cell Counting Kit-8 (Dojindo Molecular
Technologies, Inc.) was used to detect cell proliferation,
according to the manufacturer's instruction. The abundance at 450
nm was measured. Cell migration and invasion were detected using
Transwell chambers with or without Matrigel (BD Biosciences). The
precoating was conducted at room temperature for 1 h. The
re-suspended transfected cells (1×105 cells) in
serum-free DMEM were added to the upper chamber, and the lower
chamber contained DMEM with 10% FBS. Cells were then incubated at
37°C for 48 h. The migratory and invading cells through the
membrane were fixed using 70% methanol at room temperature for 30
min, stained with 0.1% crystal violet at room temperature for 10
min, and images were captured under a light microscope
(magnification, ×200).
Bioinformatics analysis and luciferase
reporter assay
TargetScan software version 7.1 (www.targetscan.org) was used for bioinformatics
analysis. To clarify the association between TINCR and miR-7, a
QuikChange Site-Directed Mutagenesis kit (Stratagene; Agilent) was
applied to generate the mutated miR-7-binding sites of TINCR. Then,
the wild or mutated TINCR was sub-cloned into a Dual-luciferase
Target Vector (Promega Corporation), generating the wild-type (WT)
or mutated (MT) TINCR luciferase reporter plasmid, respectively. To
clarify the association between miR-7 and mTOR, a QuikChange
Site-Directed Mutagenesis kit (Stratagene; Agilent) was applied to
generate the mutated miR-7-binding sites of mTOR 3′UTR. The WT or
MT mTOR luciferase reporter plasmid was then generated. Cells were
co-transfected as aforementioned with miR-7 mimic or negative
control miR mimic (miR-NC), WT (or MT) TINCR reporter plasmid, WT
(or MT) mTOR 3′UTR reporter plasmid. At 48 h after cell
transfection, a Dual-Luciferase Reporter Assay System (Promega
Corporation) was used to examine the luciferase activities. The
ratio of firefly luciferase activity to Renilla luciferase
activity was determined.
Western blotting
The cells were lysed using RIPA buffer (Beyotime
Institute of Biotechnology). Protein concentration was determined
using a BCA kit (Abcam). Proteins (60 µg per lane) were separated
using 12% SDS-PAGE, and then transferred onto PVDF membranes
(Thermo Fisher Scientific, Inc.). The membranes were blocked with
5% dry milk in Tris-buffered saline with 0.2% Tween-20 at 4°C
overnight, and then incubated with primary antibodies against mTOR
(1:500; cat. no. ab134903) and GAPDH (1:500; cat. no. ab8245) at
room temperature for 3 h, and then incubated with horseradish
peroxidase-conjugated goat anti-rabbit secondary antibody (1:5,000;
cat. no. ab6721) at room temperature for 1 h. All antibodies were
purchased from Abcam. Then, the signals were visualized with an
enhanced chemiluminescence kit (GE Healthcare), and quantified
using ImageJ software v1.46 (National Institutes of Health).
Statistical analysis
Statistical analyses were conducted using SPSS 20.0
software (IBM Corp). Data in this study are expressed as the mean ±
SD. A Student's t-test was used for comparisons between two groups,
and a one-way ANOVA followed by a Turkey's post hoc test was used
for comparisons among >2 groups. A χ2 test was used
to analyze the association between gene expression and clinical
characteristics of patients. A Kaplan-Meier method followed by a
log-rank test was applied for the survival analyses. Spearman's
correlation analysis was also performed. P<0.05 was considered
to indicate a statistically significant difference.
Results
Upregulation of TINCR is associated
with aggressive phenotypes and poor prognosis in bladder
cancer
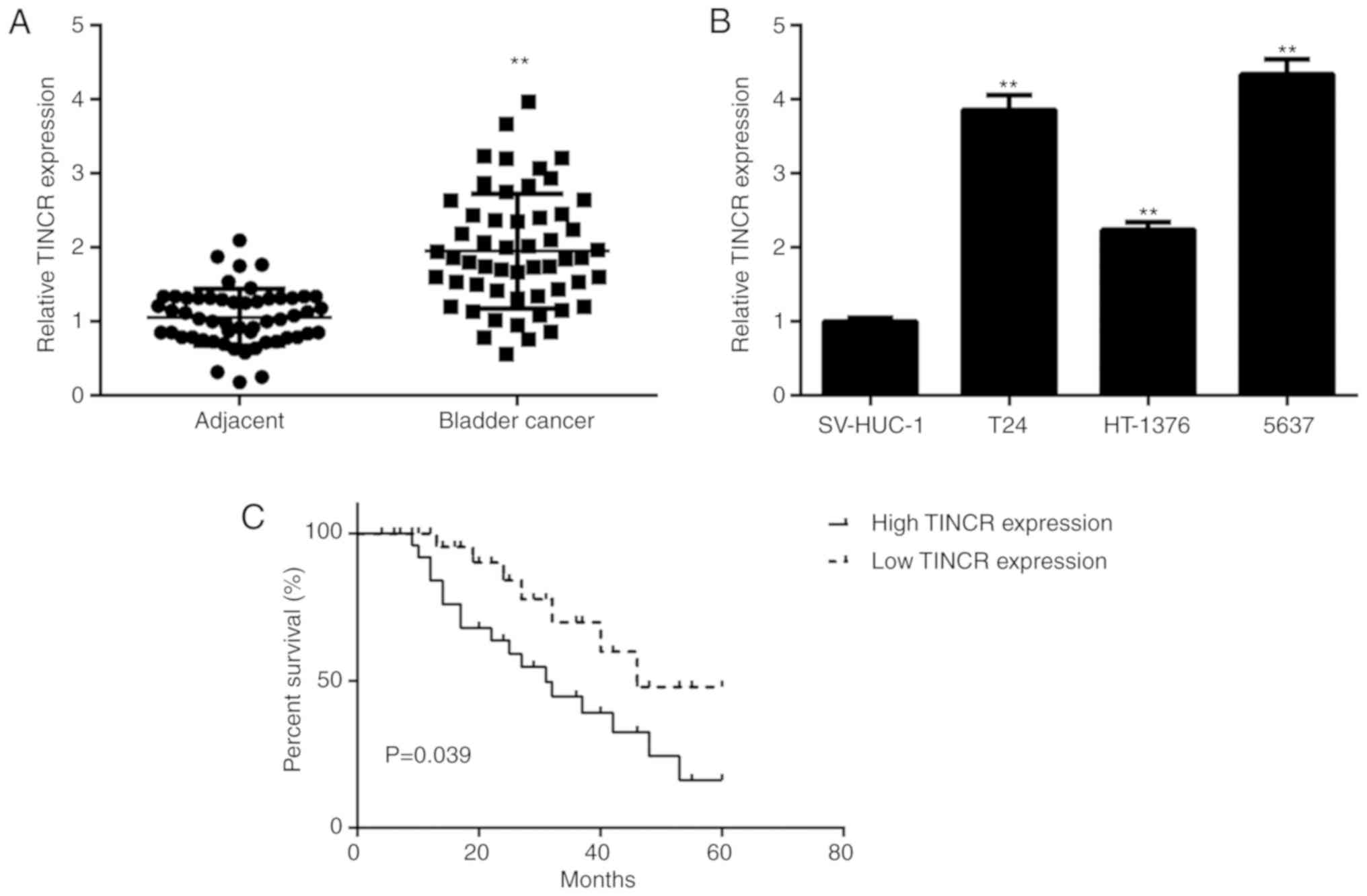
To reveal the function of TINCR in bladder cancer
progression, the expression levels of TINCR in clinical tissue
samples were examined. The results of the RT-qPCR assays showed
that the expression of TINCR was significantly increased in bladder
cancer tissues compared to matched adjacent normal tissues
(Fig. 1A). Moreover, the TINCR
levels were also increased in the bladder cancer cell lines
compared with those in the SV-HUC-1 cells (Fig. 1B). Based on the median expression
value (1.97) of TINCR as a cut-off value, the patients with bladder
cancer were divided into high and low TINCR expression groups.
Moreover, the patients with median expression values were included
in high TINCR expression group. It was identified that a high
expression of TINCR was significantly associated with advanced TNM
stage and metastasis (Table I).
Moreover, the patients with a high TINCR expression exhibited
poorer survival rates compared with those with a low TINCR
expression (Fig. 1C).
miR-7 is negatively regulated by TINCR
in bladder cancer
The function of TINCR in bladder cancer was then
examined, as well as the underlying molecular mechanism.
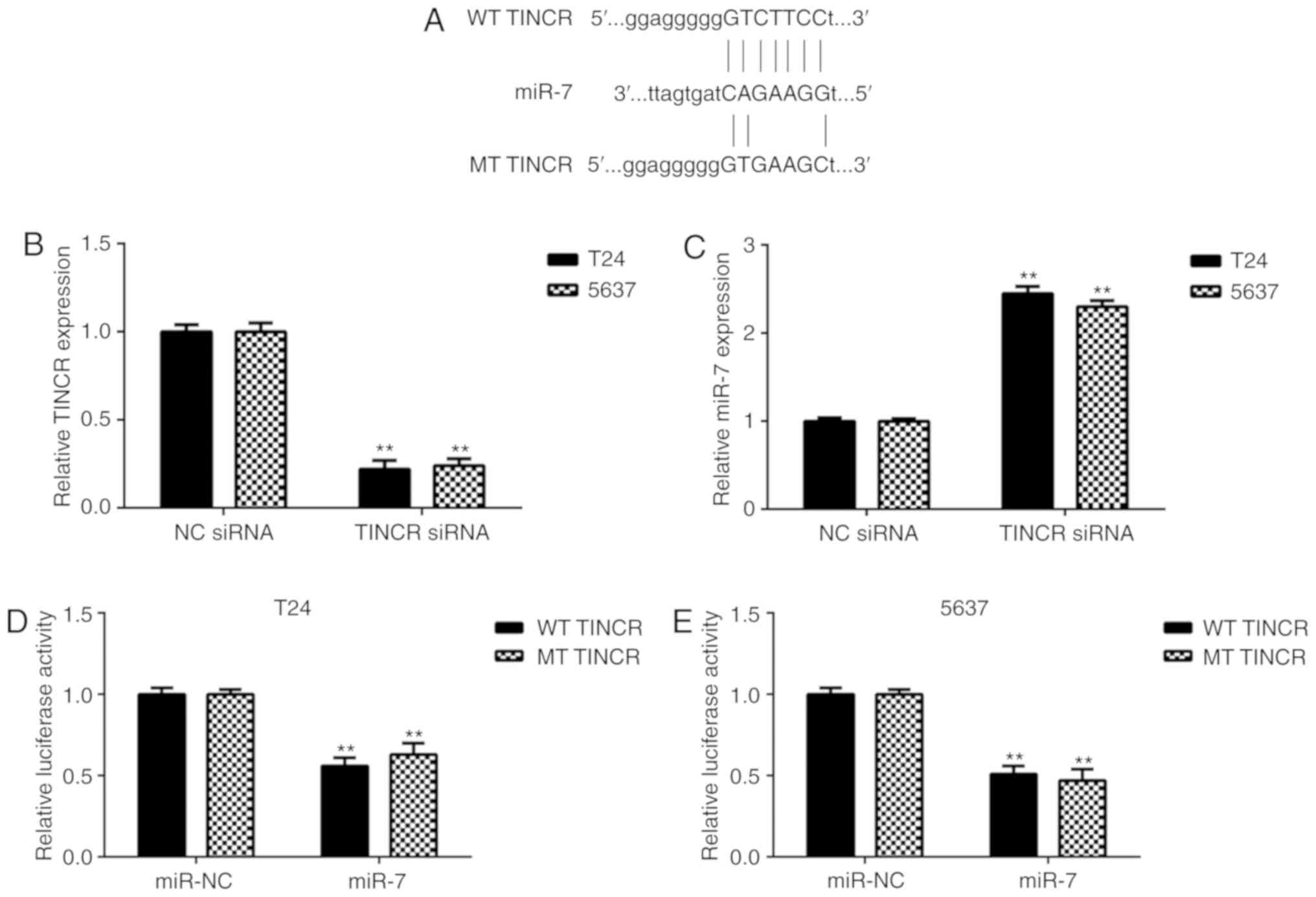
Bioinformatics analysis data showed that miR-7 could potentially
bind to TINCR, suggesting a possible interaction between TINCR and
miR-7 (Fig. 2A). T24 and 5637
cells were used in further in vitro experiments. As TINCR
was upregulated in bladder cancer, a TINCR siRNA was transfected
into T24 and 5637 cells to knock down TINCR. Following
transfection, TINCR expression levels were significantly
downregulated in the TINCR siRNA group compared with the negative
control (NC) siRNA group (Fig.
2B). Further investigation indicated that the expression levels
of miR-7 were significantly increased in the TINCR siRNA group
compared with the NC siRNA group, suggesting that the miR-7 levels
were negatively affected by TINCR in bladder cancer cells (Fig. 2C). Moreover, the luciferase
reporter gene assay data showed that co-transfection of miR-7 and
WT TINCR luciferase reporter plasmid significantly decreased
luciferase activity (Fig. 2D and
E). These results indicated the association between TINCR and
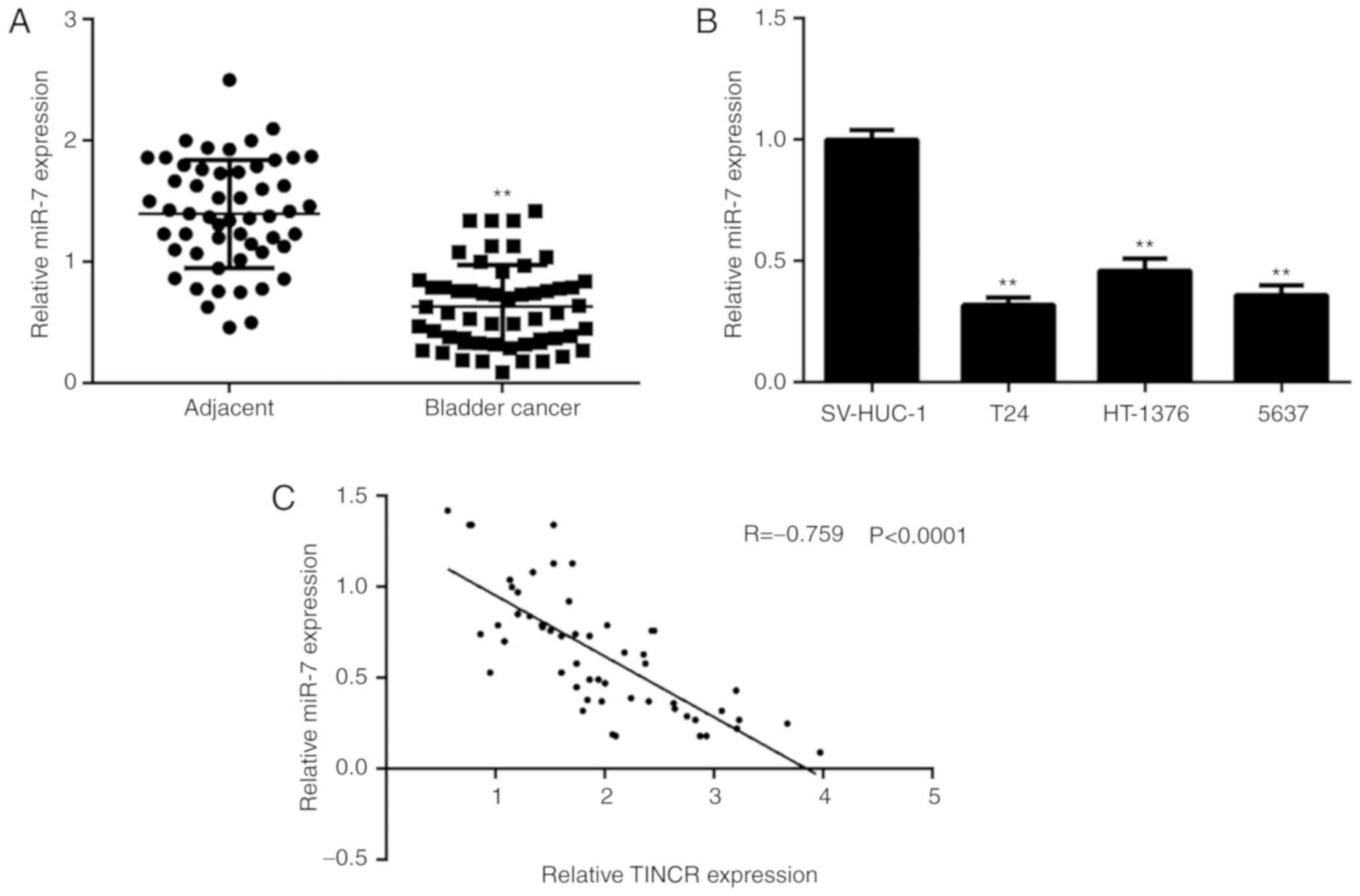
miR-7 in bladder cancer cells. It was then identified that miR-7
was significantly downregulated in bladder cancer tissues and cell
lines (Fig. 3A and B). It was then
demonstrated that there was an inverse correlation between the
expression of miR-7 and TINCR in bladder cancer tissues (Fig. 3C), which further confirms their
targeting relationship. In addition, low miR-7 expression was
associated with advanced TNM stage and lymph node metastasis
(Table II).
| Table II.Association between miR-7 expression
and clinicopathological characteristics in bladder cancer. |
Table II.
Association between miR-7 expression
and clinicopathological characteristics in bladder cancer.
|
|
| miR-7
expression |
|
|---|
|
|
|
|
|
|---|
| Variables | Cases (n=53) | High (n=26) | Low (n=27) | P-value |
|---|
| Age |
|
|
| 0.398 |
|
<55 | 19 | 11 | 8 |
|
|
≥55 | 34 | 15 | 19 |
|
| Sex |
|
|
| 0.779 |
|
Male | 33 | 17 | 16 |
|
|
Female | 20 | 9 | 11 |
|
| Grade |
|
|
| 0.054 |
| Well
and moderately | 40 | 23 | 17 |
|
|
Poor | 13 | 3 | 10 |
|
| Lymph node
metastasis |
|
|
| 0.021a |
|
Negative | 34 | 21 | 13 |
|
|
Positive | 19 | 5 | 14 |
|
| Distant
metastasis |
|
|
| 0.051 |
|
Present | 5 | 0 | 5 |
|
|
Absent | 48 | 26 | 22 |
|
| TNM stage |
|
|
| 0.014a |
|
I–II | 27 | 18 | 9 |
|
|
III–IV | 26 | 8 | 18 |
|
miR-7 is involved in the
TINCR-mediated bladder cancer cell proliferation, migration and
invasion
The regulatory mechanism of TINCR and miR-7
underlying bladder cancer cell proliferation and invasion was
further studied. Firstly, bladder cancer cells were transfected
with an NC inhibitor, miR-7 inhibitor, miR-NC mimic or miR-7 mimic.
As shown in Fig. 4A and B,
transfection with a miR-7 inhibitor significantly decreased miR-7
expression, and transfection with a miR-7 mimic significantly
increased miR-7 expression in bladder cancer cells. Then, bladder
cancer cells were transfected with NC siRNA + NC inhibitor, TINCR
siRNA + NC inhibitor, TINCR siRNA + miR-7 inhibitor. Following
transfection, the expression levels of TINCR were significantly
downregulated in the TINCR siRNA + NC inhibitor and TINCR siRNA +
miR-7 inhibitor groups, compared with the NC siRNA + NC inhibitor
group (Fig. 4C). Whereas, miR-7
was upregulated in the TINCR siRNA + NC inhibitor group compared
with the NC siRNA + NC inhibitor group, which was reversed by
transfection with a miR-7 inhibitor (Fig. 4D). It was identified that
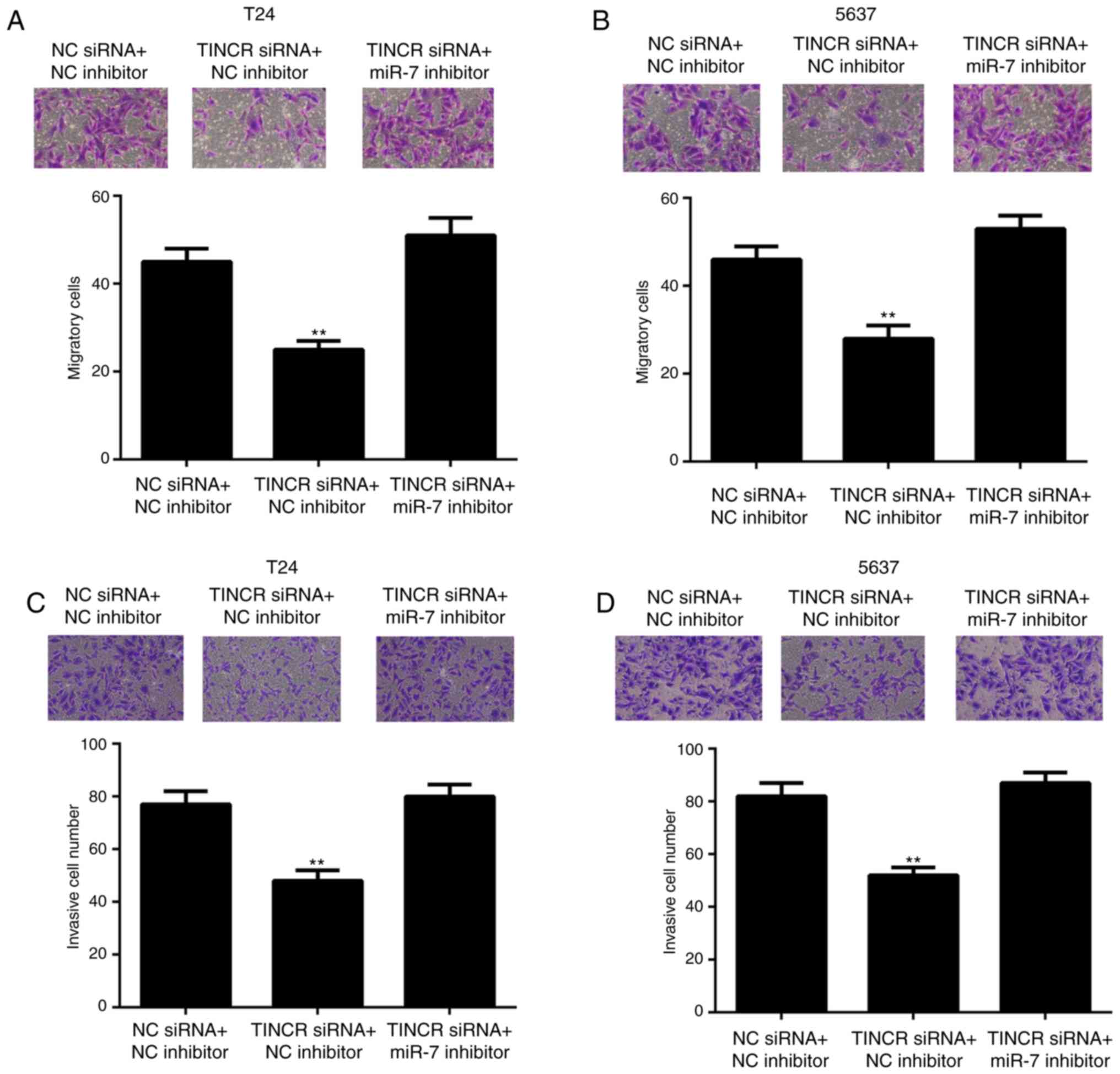
inhibition of TINCR repressed bladder cancer cell proliferation,
which was reversed by miR-7 inhibition (Fig. 4E and F). Moreover, knockdown of
TINCR also suppressed cell migration and invasion, which was
reversed by knockdown of miR-7 (Fig.
5). For cell migration, the number of migratory cells were as
follows: In Fig. 5A, NC siRNA + NC
inhibitor (45.2±3.7), TINCR siRNA + NC inhibitor (23.4±1.8) and
TINCR siRNA + miR-7 inhibitor (49.9±5.1); and in Fig. 5B, NC siRNA + NC inhibitor
(46.3±3.5), TINCR siRNA + NC inhibitor (27.2±3.6) and TINCR siRNA +
miR-7 inhibitor (50.4±3.4). For cell invasion, the number of
invaded cells were as follows: In Fig.
5C, NC siRNA + NC inhibitor (77.2±3.1), TINCR siRNA + NC
inhibitor (45.1±2.6) and TINCR siRNA + miR-7 inhibitor (79.4±2.9);
and in Fig. 5D, NC siRNA + NC
inhibitor (82.4±2.9), TINCR siRNA + NC inhibitor (52.1±1.7) and
TINCR siRNA + miR-7 inhibitor (84.3±2.5). These data suggest that
miR-7 participates in the TINCR-mediated bladder cancer cell
proliferation, migration and invasion.
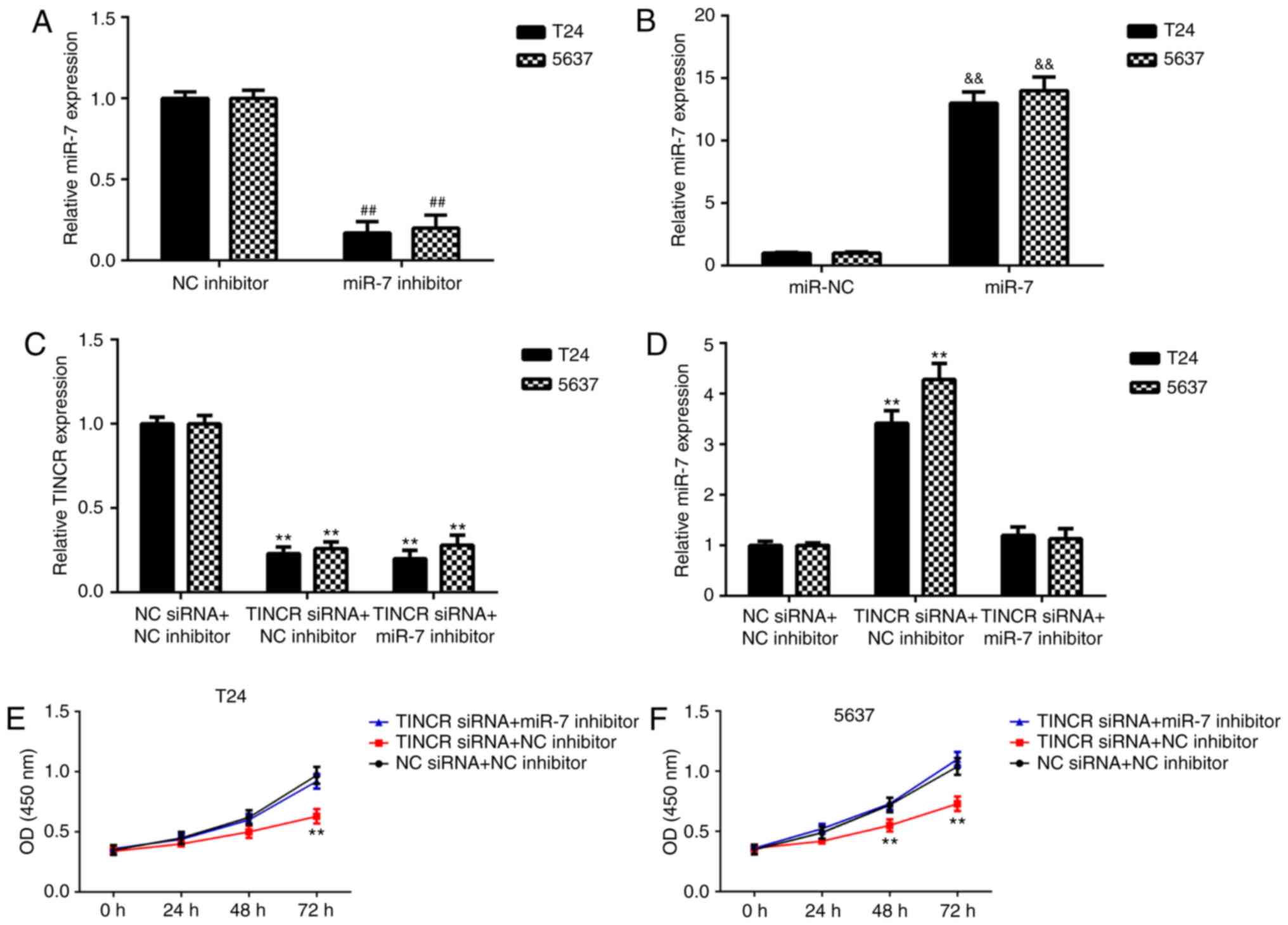 | Figure 4.Knockdown of miR-7 attenuates the
inhibitory effects of TINCR knockdown on bladder cancer cell
proliferation. (A) T24 and (B) 5637 cells were transfected with NC
inhibitor, miR-7 inhibitor, miR-NC mimic or miR-7 mimic. Following
transfection, the expression of miR-7 was examined.
##P<0.01 vs. NC inhibitor;
&&P<0.01 vs. miR-NC. (C and D) T24 and 5637
cells were transfected with NC siRNA + NC inhibitor, TINCR siRNA +
NC inhibitor, TINCR siRNA + miR-7 inhibitor, following which the
expression of (C) TINCR and (D) miR-7 was measured. (E and F) A
CKK-8 assay was conducted to study cell proliferation in (E) T24
and (F) 5637 cells. **P<0.01 vs. NC siRNA + NC inhibitor. TINCR,
TINCR ubiquitin domain containing; miR, microRNA; NC, negative
control; siRNA, small interfering RNA. |
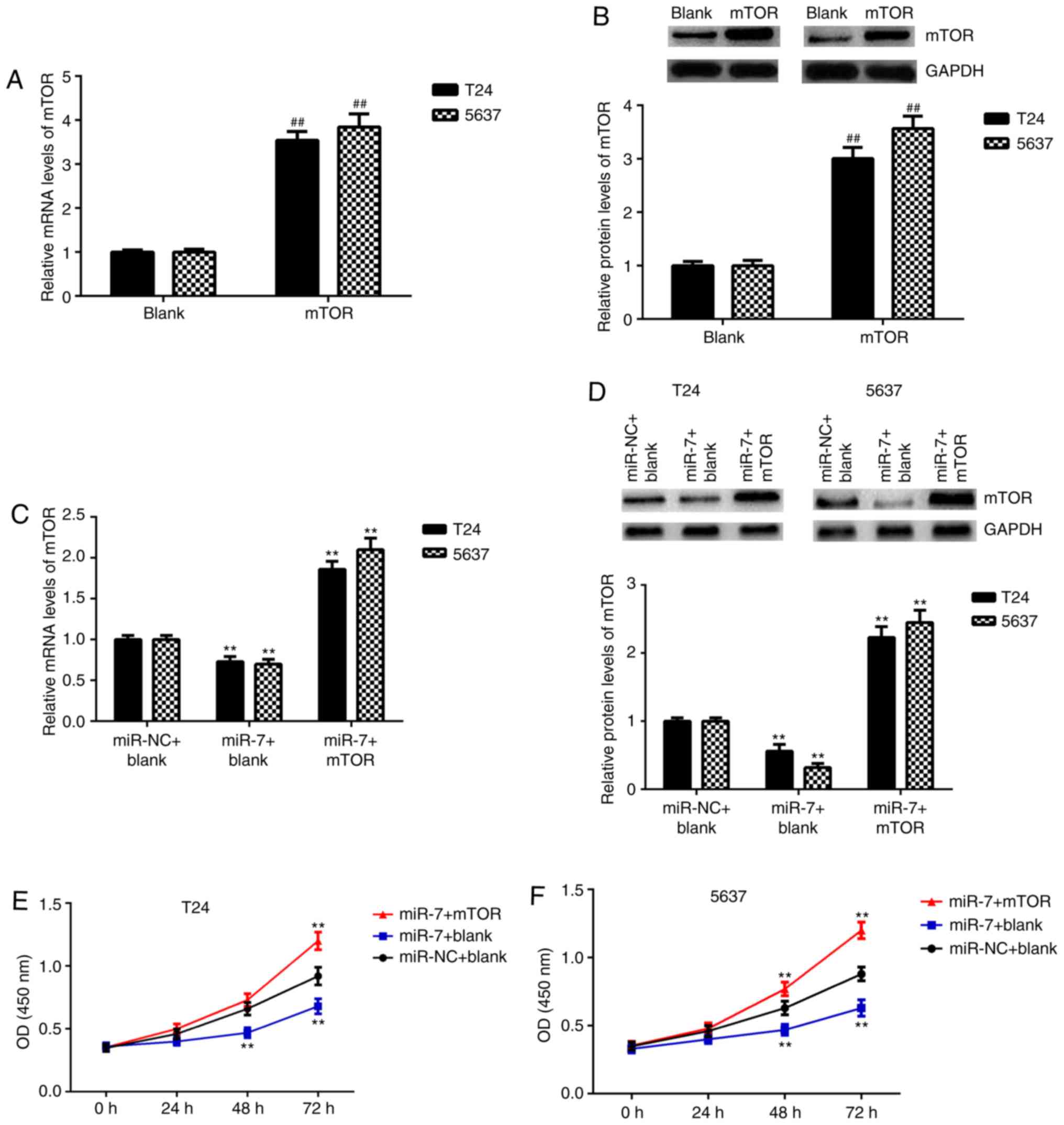
mTOR is a target gene of miR-7 in
bladder cancer cells
Recently, an increasing number of lncRNAs have been
reported to competitively inhibit the expression levels of miRs by
acting as ceRNAs, which could further mediate the expression of the
downstream target genes (7).
Bioinformatics analyses data have shown that the 3′UTR of mTOR mRNA
contained putative binding sites of miR-7 (Fig. 6A). The results of the luciferase
reporter gene assays showed that transfection of miR-7 mimics
significantly inhibited the luciferase activity of WT of 3′UTR of
mTOR, without affecting the luciferase activity of MT of 3′UTR of
mTOR (Fig. 6B and C). Accordingly,
mTOR is a direct target gene of miR-7 in bladder cancer cells.
Consistently, miR-7 overexpression led to the downregulation of
mTOR expression in bladder cancer cells, while silencing of miR-7
expression increased mTOR expression (Fig. 6D-G).
mTOR participates in the
miR-7-mediated malignant phenotypes of bladder cancer cells
It was then studied whether mTOR was a downstream
effecter in the miR-7-mediated proliferation and invasion of
bladder cells. Firstly, bladder cancer cells were transfected with
a blank vector or mTOR expression plasmid. As shown in Fig. 7A and B, transfection with an mTOR
expression plasmid significantly increased the mRNA and protein
levels of mTOR in bladder cancer cells compared with the blank
group. Bladder cancer cells were co-transfected with miR-NC + blank
vector, miR-7 mimic + blank vector, or miR-7 mimic + mTOR plasmid.
As shown in Fig. 7C and D, the
expression of mTOR was significantly reduced after miR-7
overexpression, which was reversed by transfection with the mTOR
plasmid. Following this, it was identified that cell proliferation
was decreased by miR-7 overexpression, which was reversed by mTOR
overexpression (Fig. 7E and F).
Moreover, overexpression of mTOR also reversed the miR-7-induced
inhibition of migration and invasion of bladder cancer cells
(Fig. 8). For cell migration, the
number of migratory cells were as follows: In Fig. 8A, miR-NC + blank vector (40.1±3.0),
miR-7 mimic + blank vector (23.3±1.9) and miR-7 mimic + mTOR
plasmid (56.6±2.9); and in Fig.
8B, miR-NC + blank vector (45.3±3.9), miR-7 mimic + blank
vector (23.8±1.8) and miR-7 mimic + mTOR plasmid (49.6±3.8). For
cell invasion, the number of invaded cells were as follows: In
Fig. 8C, miR-NC + blank vector
(55.2±2.2), miR-7 mimic + blank vector (37.6±1.1) and miR-7 mimic +
mTOR plasmid (83.4±4.4); and in Fig.
8D, miR-NC + blank vector (62.1±3.2), miR-7 mimic + blank
vector (39.5±2.1) and miR-7 mimic + mTOR plasmid (87.4±4.5).
Therefore, the present findings suggest that miR-7 plays an
inhibitory role in the malignant phenotypes of bladder cancer cells
by directly targeting mTOR.
Discussion
The regulatory mechanism of TINCR during bladder
cancer progression is complicated. In the present study, it was
identified that the expression of TINCR was significantly increased
in bladder cancer tissues and cell lines, when compared with that
in adjacent normal tissues and normal urinary tract epithelial cell
line SV-HUC-1, respectively. Moreover, the high expression of TINCR
was associated with tumor metastasis and advanced TNM stage, as
well as decreased survival rates of patients with bladder cancer.
Further investigation revealed that miR-7 was negatively mediated
by TINCR in bladder cancer cells. Silencing of TINCR expression
significantly increased the expression of miR-7 and reduced bladder
cancer cell proliferation, migration and invasion, while knockdown
of miR-7 expression eliminated the inhibitory effects of TINCR
downregulation on bladder cancer cells. mTOR was then identified as
a target gene of miR-7 in bladder cancer, and it was demonstrated
that overexpression of mTOR abolished the inhibitory effects of
miR-7 on bladder cancer cells.
In recent years, an increasing number of lncRNAs,
including TINCR, have been reported to serve critical roles in
different types of human cancer (25,26).
For example, SP1-induced upregulation of TINCR regulates gastric
cancer cell proliferation and apoptosis (27). Loss of TINCR expression promotes
colorectal cancer cell proliferation and metastasis (28). The present study identified that
TINCR was upregulated in bladder cancer, and that the increased
expression of TINCR was correlated with metastasis and advanced TNM
stage, consistent with the results from a previous study (12). Therefore, the upregulation of TINCR
may contribute to bladder cancer progression. Moreover, it was
found that the patients with bladder cancer who had higher TINCR
expression showed shorter survival time, when compared with those
with low TINCR expression, suggesting that TINCR may be used as a
potential prognostic maker for bladder cancer. Functionally, it was
demonstrated that knockdown of TINCR inhibited the malignant
phenotypes of bladder cancer cells, which indicates that TINCR has
an important role in bladder cancer. In future experiments it would
be useful to study the function of TINCR in bladder cancer in
vivo using animal experiments. A limitation of this study is
that only samples of the tumor tissue and adjacent tissue from the
same patient were collected during surgical resection. Thus,
further studies comparing the expression levels of TINCR in the
normal population with tumor tissue from patients would be
beneficial.
It has been well-established that lncRNAs can
interact with miRNAs and thus affect their function (6,7). In
the present study, it was demonstrated that TINCR sponges miR-7 in
order to negatively regulate its expression in bladder cancer
cells. Moreover, miR-7 was identified to be significantly
downregulated in bladder cancer, which may be due to the
upregulation of TINCR, as it was observed that TINCR expression was
inversely correlated to the miR-7 expression in bladder cancer
tissues. Further investigation indicated that the inhibition of
miR-7 reversed the inhibitory effects of TINCR knockdown on bladder
cancer cells, suggesting that TINCR serves a role in promoting the
malignant phenotypes of bladder cancer cells by negatively
regulating the expression of miR-7. In addition to miR-7, miR-375
was also recently identified to interact with TINCR in gastric
cancer (8).
In addition, it was identified that mTOR was a
direct target gene of miR-7 in bladder cancer cells, and that the
overexpression of mTOR abolished the miR-7-induced inhibition of
bladder cancer cell proliferation, migration and invasion. In fact,
this targeting relationship has also been reported in several other
types of human cancer (29,30).
For example, miR-7 increases cisplatin sensitivity of gastric
cancer cells by suppressing the expression of mTOR (29). Glover et al (30) reported that miR-7 functioned as a
tumor suppressor and novel therapeutic agent for adrenocortical
carcinoma by targeting mTOR and RAF1. Therefore, the results of the
present study improved the understanding of the function of
miR-7/mTOR axis in human cancer.
In summary, the present study demonstrated that the
upregulation of TINCR was associated with an aggressive tumor
phenotype and poor prognosis in bladder cancer, and suggested that
TINCR promotes the malignant phenotypes of bladder cancer by
regulating the miR-7/mTOR axis. Therefore, the TINCR/miR-7/mTOR
signaling pathway may be a potential therapeutic target for bladder
cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
GX, HY and LS designed the study and wrote the
manuscript. GX, HY, ML, JN and XT collected tissue samples and
performed the experiments. WC analyzed the data. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
Shengli Hospital of Shengli Petroleum Administration. All patients
provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yoshino H, Seki N, Itesako T, Chiyomaru T,
Nakagawa M and Enokida H: Aberrant expression of microRNAs in
bladder cancer. Nat Rev Urol. 10:396–404. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jiang Z, Zhang Y, Cao R, Li L, Zhong K,
Chen Q and Xiao J: miR-5195-3p inhibits proliferation and invasion
of human bladder cancer cells by directly targeting oncogene KLF5.
Oncol Res. 25:1081–1087. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Long Y, Wu Z, Yang X, Chen L, Han Z, Zhang
Y, Liu J, Liu W and Liu X: MicroRNA-101 inhibits the proliferation
and invasion of bladder cancer cells via targeting c-FOS. Mol Med
Rep. 14:2651–2656. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang Y, Dai Q, Zeng F and Liu H: MALAT1
promotes the proliferation and metastasis of osteosarcoma cells by
activating the Rac1/JNK pathway via targeting MiR-509. Oncol Res.
Apr 27–2018.(Epub ahead of print). View Article : Google Scholar
|
|
7
|
Zhang JJ, Wang DD, Du CX and Wang Y: Long
Noncoding RNA ANRIL promotes cervical cancer development by acting
as a sponge of miR-186. Oncol Res. 26:345–352. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen Z, Liu H, Yang H, Gao Y, Zhang G and
Hu J: The long noncoding RNA, TINCR, functions as a competing
endogenous RNA to regulate PDK1 expression by sponging miR-375 in
gastric cancer. Onco Targets Ther. 10:3353–3362. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xu S, Kong D, Chen Q, Ping Y and Pang D:
Oncogenic long noncoding RNA landscape in breast cancer. Mol
Cancer. 16:1292017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zheng Y, Yang C, Tong S, Ding Y, Deng W,
Song D and Xiao K: Genetic variation of long non-coding RNA TINCR
contribute to the susceptibility and progression of colorectal
cancer. Oncotarget. 8:33536–33543. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xu Y, Qiu M, Chen Y, Wang J, Xia W, Mao Q,
Yang L, Li M, Jiang F, Xu L and Yin R: Long noncoding RNA, tissue
differentiation-inducing nonprotein coding RNA is upregulated and
promotes development of esophageal squamous cell carcinoma. Dis
Esophagus. 29:950–958. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen Z, Liu Y, He A, Li J, Chen M, Zhan Y,
Lin J, Zhuang C, Liu L, Zhao G, et al: Theophylline controllable
RNAi-based genetic switches regulate expression of lncRNA TINCR and
malignant phenotypes in bladder cancer cells. Sci Rep. 6:307982016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhou Y, Yang C, Wang K, Liu X and Liu Q:
MicroRNA-33b inhibits the proliferation and migration of
osteosarcoma cells via targeting hypoxia-inducible factor-1α. Oncol
Res. 25:397–405. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Luo J, Li H and Zhang C: MicroRNA-7
inhibits the malignant phenotypes of nonsmall cell lung cancer
in vitro by targeting Pax6. Mol Med Rep. 12:5443–5448. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu K, Chen Z, Qin C and Song X: miR-7
inhibits colorectal cancer cell proliferation and induces apoptosis
by targeting XRCC2. Onco Targets Ther. 7:325–332. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang N, Li X, Wu CW, Dong Y, Cai M, Mok
MT, Wang H, Chen J, Ng SS, Chen M, et al: microRNA-7 is a novel
inhibitor of YY1 contributing to colorectal tumorigenesis.
Oncogene. 32:5078–5088. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xiong S, Zheng Y, Jiang P, Liu R, Liu X
and Chu Y: MicroRNA-7 inhibits the growth of human non-small cell
lung cancer A549 cells through targeting BCL-2. Int J Biol Sci.
7:805–814. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Thiel A and Ristimäki A: Targeted therapy
in gastric cancer. APMIS. 123:365–372. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Latacz A, Russell JA, Ocłon E, Zubel-łojek
J and Pierzchała-Koziec K: mTOR pathway-novel modulator of
astrocyte activity. Folia Biol (Krakow). 63:95–105. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sathe A and Nawroth R: Targeting the
PI3K/AKT/mTOR pathway in bladder cancer. Methods Mol Biol.
1655:335–350. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Magers MJ, Lopez-Beltran A, Montironi R,
Williamson SR, Kaimakliotis HZ and Cheng L: Staging of bladder
cancer. Histopathology. 74:112–134. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu L, Chen X, Zhang Y, Hu Y, Shen X and
Zhu W: Long non-coding RNA TUG1 promotes endometrial cancer
development via inhibiting miR-299 and miR-34a-5p. Oncotarget.
8:31386–31394. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hua F, Li CH, Chen XG and Liu XP: Long
noncoding RNA CCAT2 knockdown suppresses tumorous progression by
sponging miR-424 in epithelial ovarian cancer. Oncol Res.
26:241–247. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu TP, Liu XX, Xia R, Yin L, Kong R, Chen
WM, Huang MD and Shu YQ: SP1-induced upregulation of the long
noncoding RNA TINCR regulates cell proliferation and apoptosis by
affecting KLF2 mRNA stability in gastric cancer. Oncogene.
34:5648–5661. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang ZY, Lu YX, Chang YY, Chang YY, Zheng
L, Yuan L, Zhang F, Hu YH, Zhang WJ and Li XN: Loss of TINCR
expression promotes proliferation, metastasis through activating
EpCAM cleavage in colorectal cancer. Oncotarget. 7:22639–22649.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xu N, Lian YJ, Dai X and Wang YJ: miR-7
increases cisplatin sensitivity of gastric cancer cells through
suppressing mTOR. Technol Cancer Res Treat. 16:1022–1030. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Glover AR, Zhao JT, Gill AJ, Weiss J,
Mugridge N, Kim E, Feeney AL, Ip JC, Reid G, Clarke S, et al:
MicroRNA-7 as a tumor suppressor and novel therapeutic for
adrenocortical carcinoma. Oncotarget. 6:36675–36688. 2015.
View Article : Google Scholar : PubMed/NCBI
|