Introduction
Alveolar bones in the mouth-jaw system are unique
and dynamic. They can not only distribute occlusal forces and
regulate tooth eruption, but also connect the external environment
to periodontium and the interior of the body through the
periodontium (1). Accordingly,
external mechanical stimuli, such as tooth extraction, will trigger
cellular and morphological changes of alveolar bones, accompanied
by inflammation (2,3). These properties are vital in the
remodeling process of alveolar bones and the quality and quantity
of alveolar bones are also essential for the success and
maintenance of dental implant treatment for partially or fully
edentulous patients (4,5) and for maintaining the health of
stomatognathic system (6).
Although dental implantation and cures possess a high success rate
at present (5), problems and risks
still exist, for example, 1–2% of patients have primary implant
failure in the first few months due to inadequate bone integration
(7). Therefore, understanding the
molecular mechanism of alveolar bone remodeling may provide a new
way for effective treatment of periodontitis.
Notoginsenoside R1 (NTR1) is the main functional
ingredient derived from Panax notoginseng, a traditional
Chinese herbal medicine (8).
Previous studies have shown that NTR1 has cardio-protective and
neuro-protective effects (9–11).
In addition, NTR1 has the function of promoting osteoblastogenesis
through regulating runt-related transcription factor 2 (RUNX2) and
activating the Smad-independent signaling pathway p38/MAPK
(12). It can also inhibit
osteolysis, osteoclastogenesis and bone absorption by suppressing
the receptor activator of NF-κB ligand (RANKL)-regulated MAPK
signaling pathway and NF-κB signaling pathway (13). NTR1 has an anti-inflammatory effect
on inflammatory bowel disease by elevating the expressions of
xenobiotic-metabolizing enzymes and downregulating the activity of
the NF-κB signaling pathway (14).
Furthermore, NTR1 can mediate inflammation induced by oxidized
low-density lipoprotein and endotoxin and involving the NF-κB
signaling pathway (8,15). However, studies on the association
between NTR1 and alveolar osteoblasts are limited.
The Wnt/β-catenin signaling pathway serves an
essential role in embryo growth and tissue homoeostasis (16). It has been confirmed that the
Wnt/β-catenin signaling pathway not only regulates whole-body
energy metabolism in osteoblasts involving glucose homeostasis,
energy expenditure and fat accumulation (17), but also modulates the viability and
health of the skeleton (18) and
predominantly regulates osteoblast differentiation (19). It has been reported that
parthenolide promotes the differentiation of osteoblasts by
activating the Wnt//b-catenin pathway and inhibiting the NF-kB/p50
and ERK signaling pathways (20).
NTR1 promotes the growth of cortical neurons of neonatal rats via
the Wnt/β-catenin signaling pathway (21), but whether NTR1 has an effect on
the differentiation of human alveolar osteoblasts (HAOBSs)
involving this signaling pathway remains to be elucidated.
The present study hypothesized that NTR1 could
function as a regulator in the differentiation process of HAOBs. To
the best of the authors' knowledge, the present study the first to
investigate the regulatory effects of NTR1 on the differentiation
of HAOBs through regulation of the NF-κB and Wnt/β-catenin
signaling pathways in the inflammatory microenvironment induced by
TNF-α, which may bring new understanding of dental implantation and
periodontal treatment in clinical dentistry, to the benefit of
dental patients.
Materials and methods
Alveolar osteoblasts specimen
collection
The present study enrolled 163 patients with normal
gums who needed tooth extraction due to impacted teeth or
orthodontic treatment between March 2016 and March 2019. X-rays
were used to further screen the patients without periapical tissue
lesions and alveolar bone resorption. In addition, patients
suffering from systemic diseases, infectious diseases, diseases
that affect bone metabolism, or being treated with drugs that
affect bone metabolism were excluded. Finally, a total of 40
patients (19 males and 21 females aged 25–40 years old; median age,
32.5 years) were included. Next, 0.5 cm2 alveolar bone
tissues were surgically collected from the alveolar socket wall of
the individuals. The current study was approved by the Ethics
Committee of the Jingmen Number 1 People's Hospital (Jingmen,
China; approval no. JMPH20160123). All patients signed informed
consent and agreed to their tissues being used for clinical
research.
Cell isolation and cell culture
Alveolar bones samples were cleaned and maintained
in phosphate buffer saline (PBS; Thermo Fisher Scientific, Inc.)
containing 100 units/ml penicillin (TargetMol) and 100 µg/ml
streptomycin (TargetMol). The alveolar bone granules were then
digested in 0.25% of trypsin digestion solutions (Beijing Solarbio
Science & Technology Co., Ltd.) at 37°C for 20 min and further
digested in a mixture of 0.1% collagenase type I (Sigma-Aldrich;
Merck KGaA) and 0.1% collagenase type II (Sigma-Aldrich; Merck
KGaA) at a 1:1 ratio. The digested alveolar bone granules were
cultured in Dulbecco's Modified Eagle Medium (DMEM; Invitrogen;
Thermo Fisher Scientific, Inc.) containing 10% fetal bovine serum
(FBS; Invitrogen; Thermo Fisher Scientific, Inc.), 100 units/ml
penicillin (TargetMol) and 100 µg/ml streptomycin (TargetMol) at
37°C for 12 h. Osteogenic medium (OM) was used for osteoblast
differentiation, which consisted of high-glucose DMEM supplemented
with 10% FBS, 100 units/ml penicillin, 100 µg/ml streptomycin, 10
mM β-glycerophosphate (APeXBIO Technology LLC), 50 mM ascorbic acid
(BioVision, Inc.) and 100 mM dexamethasone (TargetMol),
subsequently, the granules were incubated under saturated humidity
and 5% CO2 at 37°C for 14 days and the medium was
replaced by new medium every three days. Third generation cells
were used for further experiments. Images of cell passages were
captured using an inverted phase contrast microscope IX53 (Olympus
Corporation). The third generation cells were stained using an
alkaline phosphate (ALP) staining assay and Alizarin Red S staining
assay for identification.
ALP staining assay
ALP staining assay was performed using an Alkaline
Phosphatase Stain kit (Shanghai Yeasen Biotechnology Co., Ltd.)
according to the manufacturer's instructions. HAOBs of third
generation were used to make cell slides. Following fixation with
4% formaldehyde for 4 h at room temperature, the cultured cells
were treated with a staining solution containing 3 ml buffer
solution, 10 µl 300XBCIP reagent and 20 µl 150XNBT reagent. The
cells were incubated at room temperature for 5–30 min in the dark.
Finally, the staining solution was removed and the cells were
examined using an Invitrogen EVOS M700 Imaging system (Invitrogen;
Thermo Fisher Scientific, Inc.).
Alizarin Red S staining assay
Alizarin Red S (Sigma-Aldrich; Merck KGaA) was used
to identify and evaluate the mineralization of HAOBs. For cell
identification, HAOBs of third generation were produced as a cell
smear and stained at 37°C for 5 min with 1% Alizarin Red S and
examined under an Invitrogen EVOS M7000 Imaging system after being
rinsed with distilled water.
For mineralization evaluation, HAOBs were divided
into five groups: Control group (the cells treated with DMEM for 14
days at 37°C); OM group (the cells treated with OM for 14 days at
37°C); TNF-α group (the cells stimulated by 10 ng/ml TNF-α for 14
days at 37°C); TNF-α + NTR1 2.5 group (the cells were pre-treated
with 2.5 µmol/l NTR1 and stimulated by 10 ng/ml TNF-α for 14 days
at 37°C) and TNF-α + NTR1 20 group (the cells were pre-treated with
20 µmol/l NTR1 and stimulated by 10 ng/ml TNF-α for 14 days at
37°C). The treated cells were fixed with 4% formaldehyde for 4 h at
room temperature and then stained with 1% Alizarin Red S at room
temperature for 20 min. Microscopic examinations were performed
under an inverted phase contrast microscope IX53 (Olympus
Corporation) after the cells had been washed with distilled
water.
ALP activity assay
Alkaline Phosphatase Activity Detection kit
(Shanghai Yeasen Biotechnology Co., Ltd.) was used to measure the
ALP activity of HAOBs treated with TNF-α and NTR1 according to the
manufacturer's instructions. The cells were pre-treated by OM for
osteoblast differentiation and then stimulated with 10 ng/ml TNF-α
for 7 and 14 days at 37°C. The cells were lysed in 0.2% Triton
X-100 and then centrifuged at 12,000 × g for 10 min at 4°C. The
supernatant of the medium was collected for the detection of ALP
activity under a microplate reader (Bio-Rad Laboratories, Inc.) at
a wavelength of 405 nm.
MTT assay
MTT assay was performed to detect cell viability
during cell culture process. HAOBs were treated with different
concentrations of NTR1 (2.5, 5, 10, 20 and 40 µmol/l) for 1, 3 and
7 days at 37°C. Cytotoxicity was assessed to confirm the influence
of NTR1 on cell viability. The combined effects of NTR1 and TNF-α
on the cells were also tested. A total of 10 µl MTT solution (5
mg/ml, Sigma-Aldrich; Merck KGaA) was added to each well, followed
by incubation with 5% CO2 at 37°C for 4 h. Formazan
crystals were dissolved in 100 µl DMSO at 37°C. Optical density
(OD) value was measured at 495 nm with a microplate reader (Bio-Rad
Laboratories, Inc.).
Reverse transcription-quantitative
(RT-q)PCR
Gene expression changes caused by NTR1 and TNF-α
were analyzed using RT-qPCR. HAOBs (1×106) were
pre-treated with or without 2.5 and 20 µmol/l of NTR1 and then
placed into an inflammatory microenvironment induced by TNF-α.
Total RNAs were extracted from the cells using TRIzol®
(Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocols, and then the concentration of the total RNAs was
determined using UV spectrophotometer DR3900 (Hach). A TaqMan
microRNA reverse transcription kit (Thermo Fisher Scientific, Inc.)
was used according to the manufacturer's instructions to synthesize
first-strand cDNAs. RT-qPCR was then performed with BlazeTaq™ SYBR
Green RT-qPCR Mix 2.0 (BioCat GmbH) according to the manufacturer's
protocols using PCR instrument CFX96 Touch (Bio-Rad Laboratories,
Inc.), and the reaction system with 10 µl total volume was
prepared. The conditions for RT-qPCR were 1 cycle of 95°C for 30
sec, followed by 40 cycles of at 95°C for 10 sec and at 60°C for 30
sec. The comparative cycle threshold method (2−ΔΔCq) was
used to calculate the relative expression of each mRNA (22). The experiment was repeated three
times. The primer sequences of the genes used in the experiment are
listed in Table I.
 | Table I.Primers used in reverse
transcription-quantitative PCR analysis. |
Table I.
Primers used in reverse
transcription-quantitative PCR analysis.
| Gene | Primer sequence |
|---|
| Runt-related
transcription factor 2 | Forward:
5′-TGGTTACTGTCATGGCGGGTA-3′ |
|
| Reverse:
5′-TCTCAGATCGTTGAACCTTGCTA-3′ |
| Osteopontin | Forward:
5′-GGGAATTCATGAGATTGGCAGTG-3′ |
|
| Reverse:
5′-AAGGATCCGTTGACCTCAGAAGA-3′ |
| Osteocalcin | Forward:
5′-GGGGTACCCAGAGCAGTAATTTCCGGCC-3′ |
|
| Reverse:
5′-GAAGATCTCGAACCGAACCGCCTTTATA-3′ |
| GAPDH | Forward:
5′-GGAGCGAGATCCCTCCAAAAT-3′ |
|
| Reverse:
5′-GGCTGTTGTCATACTTCTCATGG-3′ |
| AXIN2 | Forward:
5′-CAACACCAGGCGGAACGAA-3′ |
|
| Reverse:
5′-GCCCAATAAGGAGTGTAAGGACT-3′ |
| Dickkopf-related
protein 1 | Forward:
5′-CCTTGAACTCGGTTCTCAATTCC-3′ |
|
| Reverse:
5′-CAATGGTCTGGTACTTATTCCCG-3′ |
| β-actin | Forward:
5′-GTGACGTTGACATCCGTAAAGA-3′ |
|
| Reverse:
5′-GCCGGACTCATCGTACTCC-3′ |
Western blot assay
HAOBs were cultured in OM or DMEM, pretreated with
2.5 and 20 µmol/l NTR, alone or further stimulated with TNF-α for
14 days at 37°C. Then western blotting was performed to measure the
expression levels of proteins related to NF-κB pathways (at 30- and
60-min culture) and Wnt/β-catenin pathways (at 3- and 7-day
culture). HAOBs were stimulated with TNF-α, with or without NTR1
(2.5 and 20 µmol/l) for 14 days at 37°C. Total proteins were
extracted from HAOBs using Pierce RIPA buffer (Thermo Fisher
Scientific, Inc.) for 30 min on ice. Then centrifugation was
performed for 10 min at 14,000 × g at 4°C. A bicinchoninic acid
protein quantitative kit (Sigma-Aldrich; Merck KGaA) was used to
determine protein concentration according to the manufacturer's
instructions. Protein (10 µg) was separated by 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (Thermo Fisher
Scientific, Inc.), placed in a water bath with boiling water for 3
min and transferred onto nitrocellulose membranes (Thermo Fisher
Scientific, Inc.). The transferred membranes were blocked with 5%
non-fat dried milk for 1 h at room temperature and incubated with
primary antibodies at 4°C overnight. The primary antibodies were
p50 (1:1,000; cat no. 13586; Cell Signaling Technology, Inc.),
phosphorylated (p-)p65 (1:1,000; cat no. 3039; Cell Signaling
Technology, Inc.), p65 (1:1,000; cat no. 8242; Cell Signaling
Technology, Inc.), GADPH (1:1,000; cat no. 4292; Cell Signaling
Technology, Inc.), β-catenin (1:1,000; cat no. 9562; Cell Signaling
Technology, Inc.), Dickkopf-related protein 1 (DKK1) (1:1,000; 29
kDa; cat no. ab61275; Abcam) and AXIN2 (1:1,000, cat no. ab109307,
Abcam); GAPDH served as an internal reference. The membranes were
washed three times and then incubated for 1 h with corresponding
fluorescent secondary antibodies [Goat Anti-Mouse IgG H&L
(HRP); 1:5,000; cat no. ab6789; Abcam]. Finally, the grey values of
the strips were calculated by ImageJ software (version 5.0;
National Institutes of Health).
Statistical analysis
All data were expressed as mean ± standard deviation
unless otherwise shown. Student's t-test was used to analyze
statistical differences between two groups and one-way ANOVA was
used to analyze statistical differences between >2 groups,
followed by Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
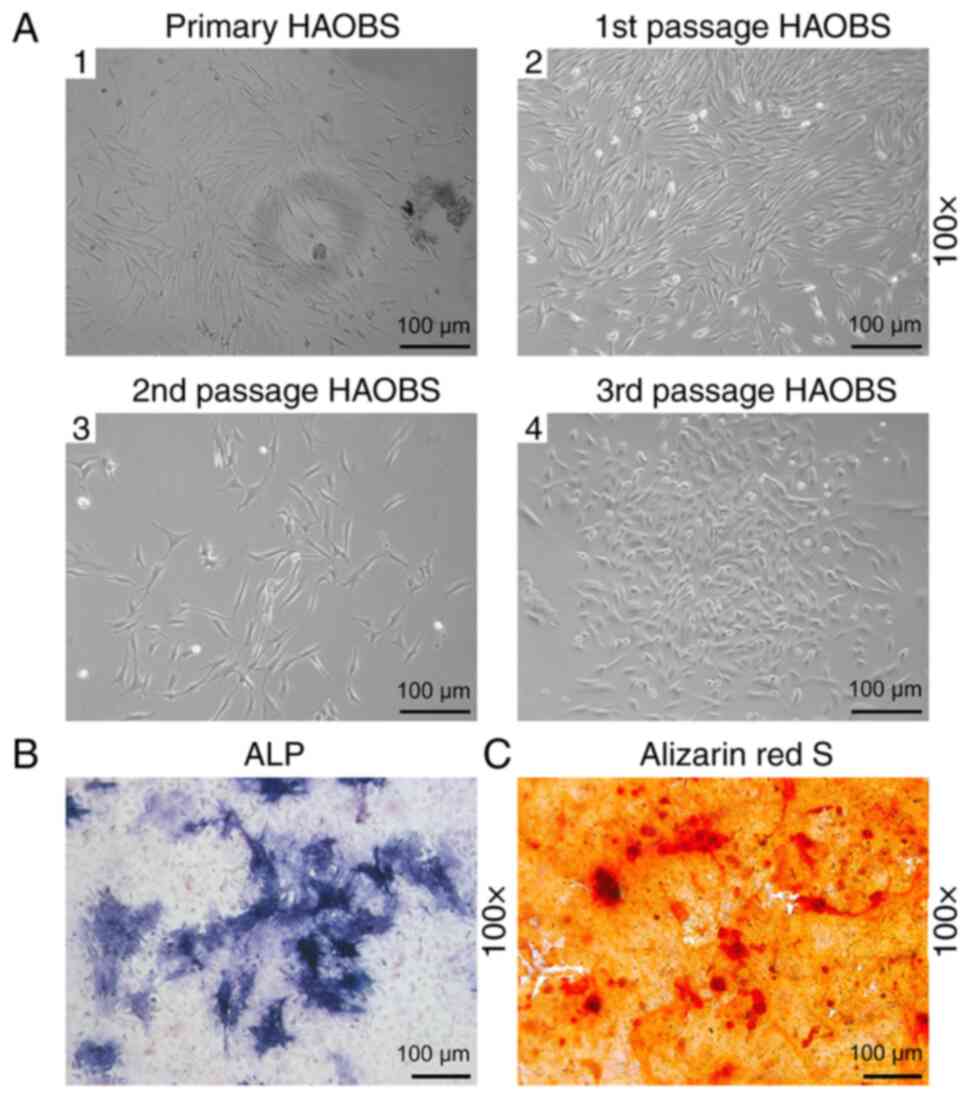
HAOBs passage and identification
Morphological changes of HAOBs during cell passage
were observed under an inverted phase contrast microscope. After 5
days of culture, primary osteoblasts started to extend from the
edge of alveolar bone granule (Fig.
1A1). The first and the second passages of osteoblasts were
usually in irregular shapes similar to triangle, spindle and
polygon (Fig. 1A2-A3). When
osteoblasts passaged to the third generation, most of them appeared
to be triangular and fusiform (Fig.
1A4). At this stage, osteoblasts were large in size, rich in
cytoplasm and had more protuberances compared with the second
passages of osteoblasts with which to connect with each other.
Large oval single nuclei with one to three nucleoli were located in
the center of HAOBs cells. As the osteoblast number increased, the
protuberances radially expanded to the surrounding areas. Then the
osteoblasts fused and overlapped, which further disclosed the white
mineralized nodules to the naked eye. Next, conventionally cultured
osteoblasts were stained with ALP. The positive expression of HAOBs
was identified by a cluster of blue granules in the cytoplasm
(Fig. 1B). The osteoblasts were
authenticated through mineralization assay. After staining with
Alizarin Red S, red calcium deposits of various sizes and shapes
could be observed (Fig. 1C). These
results indicated that HAOBs were successfully isolated and
cultured.
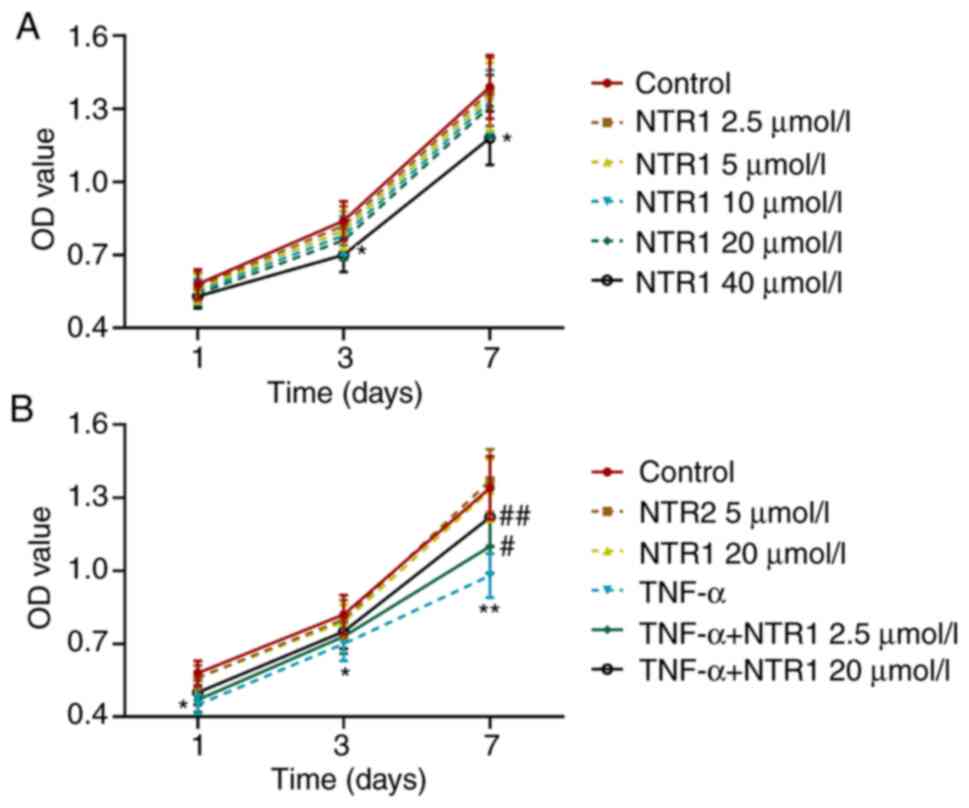
Effects of NTR1 and TNF-α on
HAOBs
HAOBs were pre-treated with different concentrations
of NTR1 in medium and then detected by MTT assay. It was identified
that the viability of the cells remained unchanged with 0–20 µmol/l
of NTR1, but was slightly inhibited by 40 µmol/l NTR1 after 3-day's
culture (P<0.05 vs. Control; Fig.
2A). Next, the cells were treated with NTR1 (2.5 and 20 µmol/l)
alone, or further stimulated by TNF-α. As shown in Fig. 2B, TNF-α could significantly inhibit
the viability of HAOBs (P<0.05, P<0.01 vs. Control or TNF-α),
thus demonstrating that TNF-α could create an inflammatory
microenvironment. NTR1 (2.5 and 20 µmol/l) and TNF-α had no effect
on the proliferation of HAOBs. These results suggested that 40
µmol/l NTR1 affected cell viability.
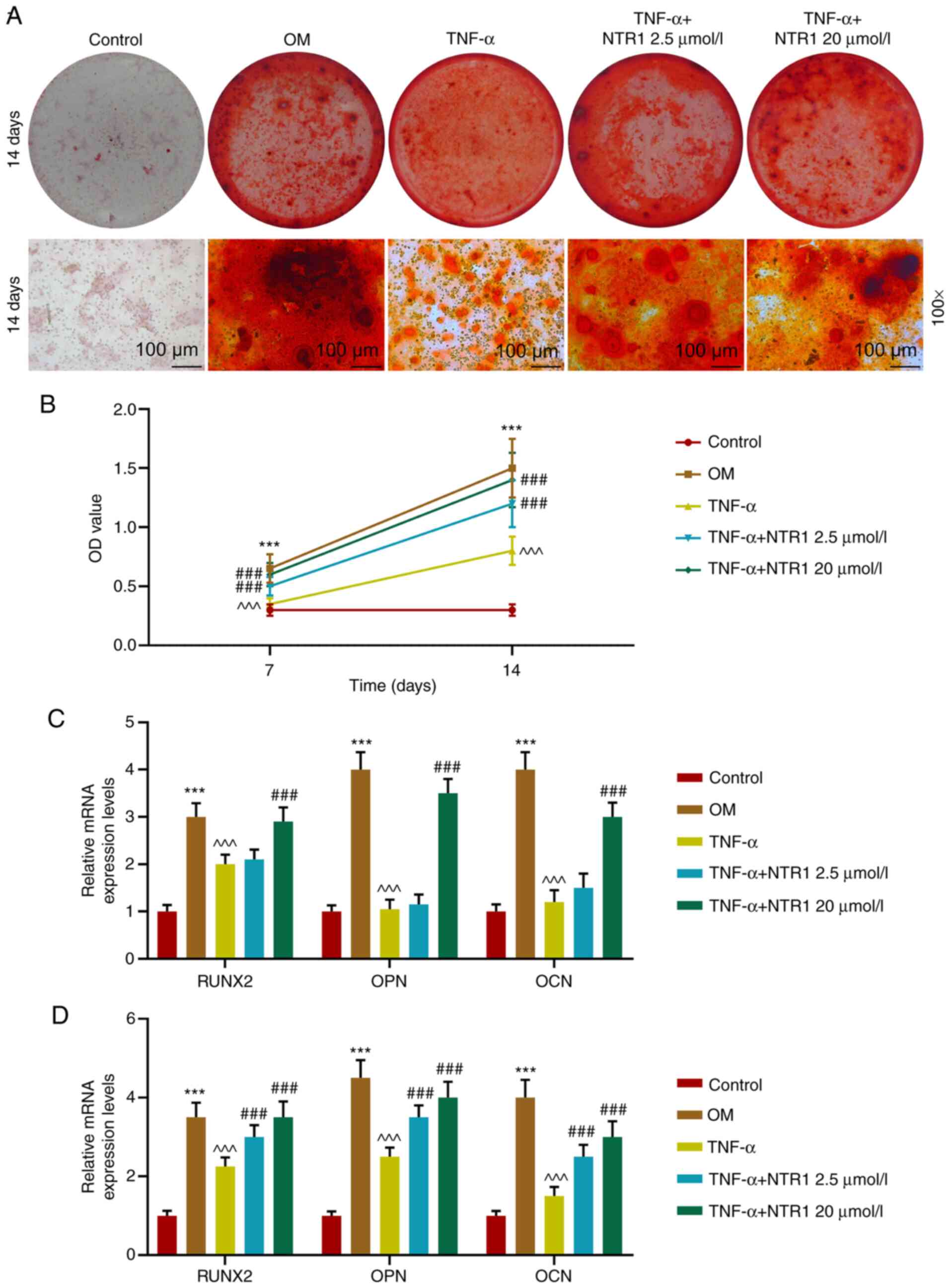
NTR1 promotes the differentiation of
HABOs in the inflammatory microenvironment induced by TNF-α
Using Alizarin Red S staining, red mineralized
nodules could be detected in OM, whereas the calcium deposits were
largely reduced by TNF-α. Nevertheless, NTR1 at an concentration of
≤20 µmol/l could abrogate the function of TNF-α in a dose-dependent
manner; TNF-α plus 20 µmol/l NTR1 resulted in more calcium deposits
compared with TNF-α plus 2.5 µmol/l NTR1 (Fig. 3A). In addition, the ALP activity in
HAOBs co-treated with TNF-α and NTR1 (2.5 and 20 µmol/l) was
significantly higher compared with that in HAOBs stimulated with
TNF-α only on both day 7 and 14 (both P<0.001 vs. Control, OM or
TNF-α; Fig. 3B). RT-qPCR was used
to measure the expression of gene markers of osteoblasts. After
3-day culture, the relative mRNA expression levels of RUNX2, OPN
and OCN were significantly inhibited by TNF-α compared with those
in the OM group at day 3 and 7 (all P<0.001 vs. Control, OM or
TNF-α; Fig. 3C and D). It was
identified that 2.5 µmol/l NTR1 had no significant effect on
TNF-α-induced results, while 20 µmol/l NTR1 reversed the effect of
TNF-α on HAOBs (P<0.001; Fig.
3C). On the 7th day of osteoblast culture, mRNA expressions
were also inhibited by TNF-α. However, both 2.5 and 20 µmol/l NTR1
could reverse the inhibitory effect of TNF-α to a large extent
(P<0.001 vs. Control, OM or TNF-α; Fig. 3D). These results indicated that
NTR1 promoted differentiation of HABOs in an inflammatory
microenvironment induced by TNF-α.
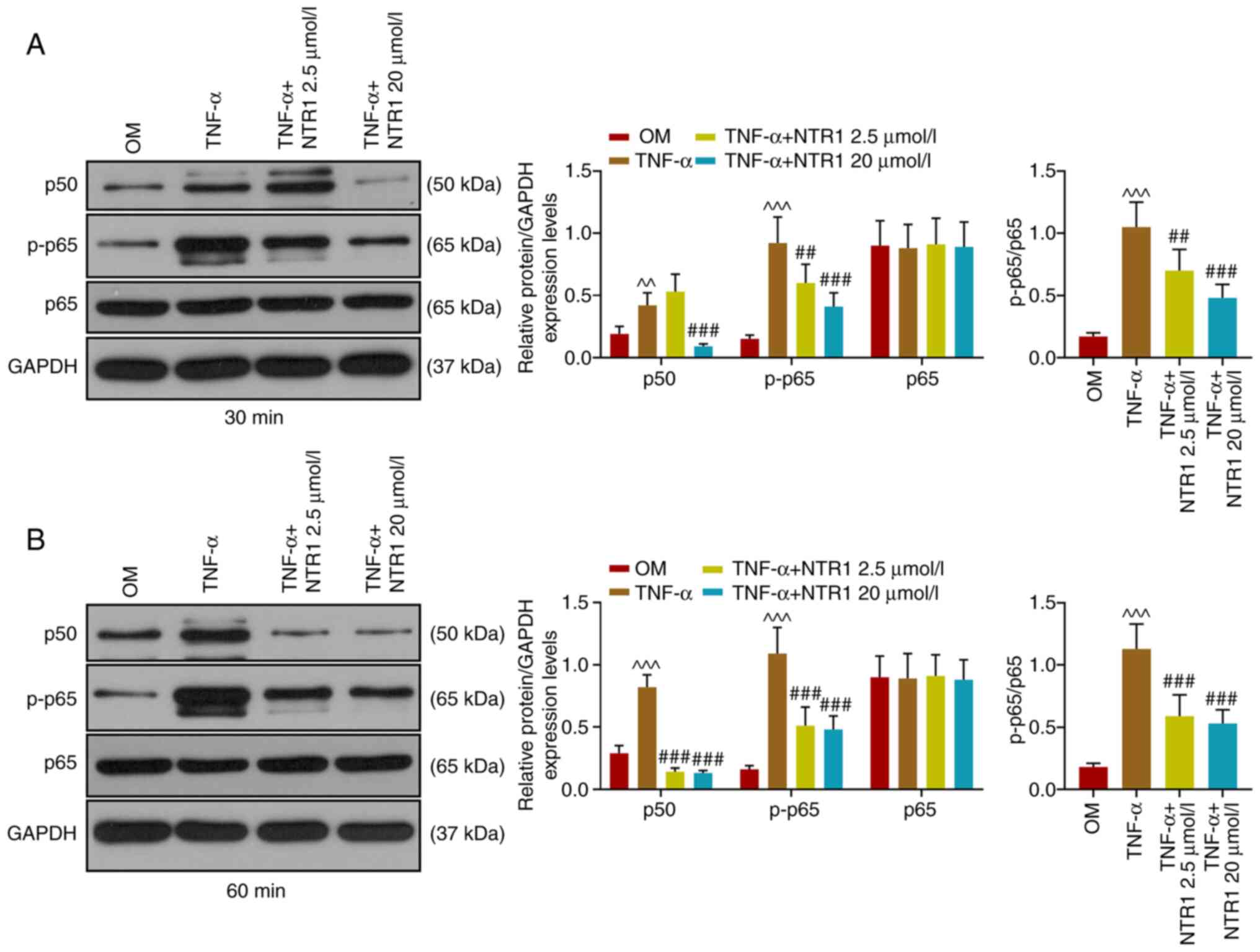
TNF-α-activated NF-κB signaling
pathway is restrained by NTR1
After HAOBs were stimulated with TNF-α for 30 and 60
min, western blotting was performed to determine the relative
protein expression levels of components of the NF-κB signaling
pathway (Fig. 4). The protein
expressions of p50 and p-p65 were both increased by TNF-α, while
that of p65 remained stable. Adding NTR1 (2.5 and 20 µmol/l)
resulted in the downregulation of p50 expression in osteoblasts
after 60-min TNF-α treatment compared with the TNF-α group
(P<0.001 vs. Control, OM or TNF-α). These results indicated that
TNF-α-activated NF-κB signaling is restrained by NTR1.
NTR1 rescues the impaired
Wnt/β-catenin signaling pathway caused by TNF-α
The expression levels of the Wnt/β-catenin signaling
pathway-related proteins in HAOBs were determined using western
blotting, after which the expressions of corresponding mRNAs were
detected by RT-qPCR. Measurements were taken on day 3 and 7 of
osteoblast treatment. Notably, the gene expression patterns of
AXIN2, DKK1 and β-catenin of both NTR1 groups presented a similar
trend. Compared with the TNF-α only group, NTR1 significantly
upregulated the expression levels of AXIN2 and β-catenin and
downregulated DKK1 expression on the 7th day (P<0.001 vs.
Control, OM or TNF-α; Fig. 5A-D).
These results indicated that NTR1 rescued the impaired
Wnt/β-catenin signaling pathway caused by TNF-α.
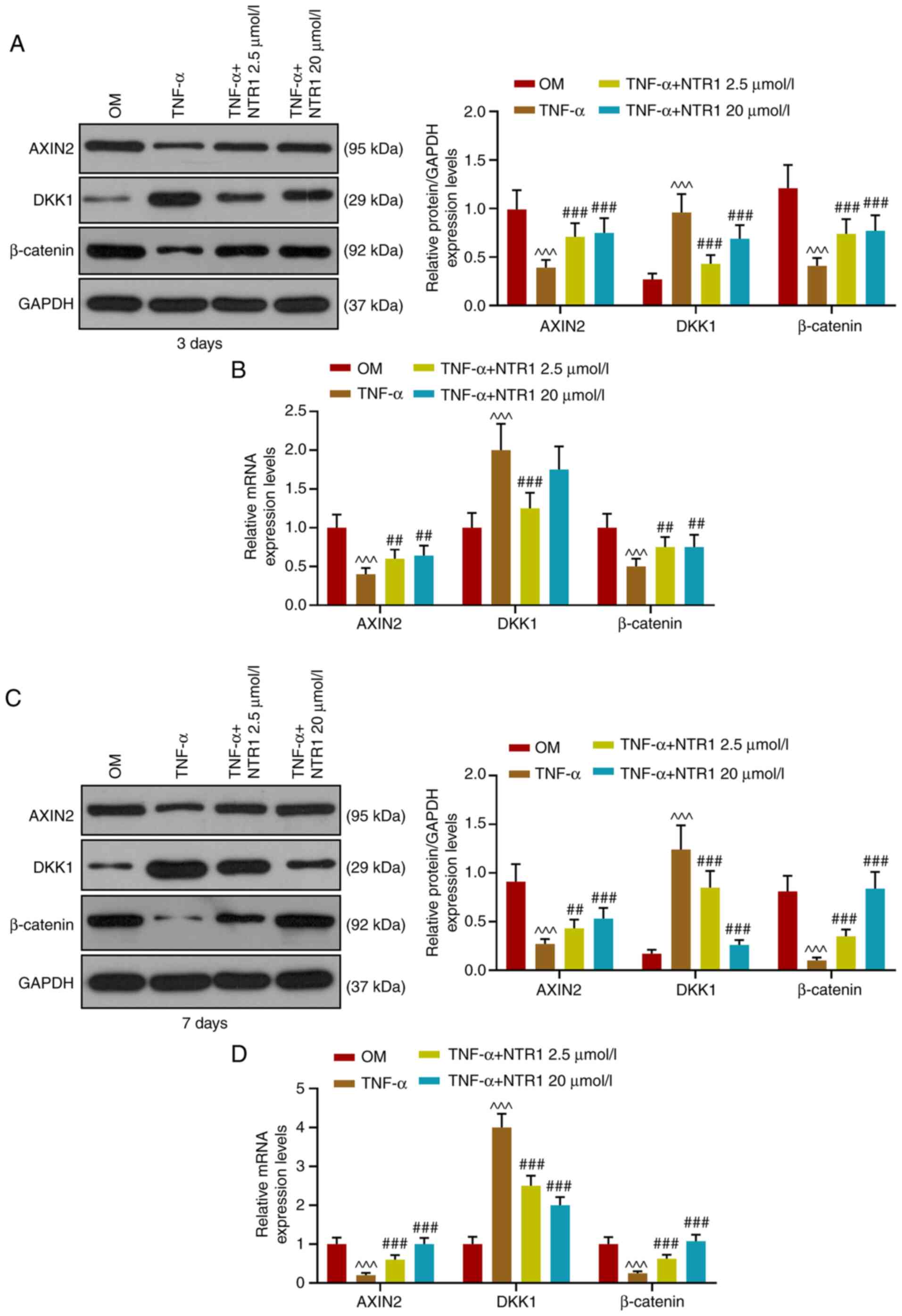 | Figure 5.Regulatory effect of TNF-α on the
Wnt/β-catenin pathway was abrogated by NTR1. After HAOBs had been
treated for 3 days, (A) western blotting and (B) RT-qPCR were used
to detect the expression of AXIN2, DKK1 and β-catenin in HAOBs.
After HAOBs had been treated for 7 days, (C) western blotting and
(D) RT-qPCR assay were used to detect the expressions of AXIN2,
DKK1 and β-catenin in HAOBs. ^^^P<0.001 vs. OM,
##P<0.01, ###P<0.001 vs. TNF-α, n=3.
NTR1, Notoginsenoside R1; HAOBs, human alveolar osteoblasts;
RT-qPCR, reverse transcription-quantitative PCR; OM, osteogenic
medium; DKK1, Dickkopf-related protein 1. |
Discussion
The present study investigated the function of NTR1
in the inflammatory microenvironment induced by TNF-α. The
concentration of NTR1 was controlled at ≤20 µmol/l. NTR1-treated
HAOBs were further stimulated with TNF-α. It was identified that
NTR1 regulated ALP activity, mineralized nodules and expression
levels of osteogenic genes in HAOBs. In addition, NTR1 abrogated
the effects of TNF-α through the NF-κB and Wnt/β-catenin signaling
pathways and served a pivotal role in osteoblast differentiation in
inflammatory microenvironment.
Inflammation is an innate immune response associated
with TNF-α, a key factor in the cytokine network (23). Essentially, inflammatory and bone
healing processes interact with each other, but prolonged
inflammatory process will one way or another hamper bone
regeneration (24). For example,
ATF3 functions as a regulator to aid TNF-α-mediated inhibition of
osteoblast differentiation, while the TNF-α-activated JNK pathway
further induces ATF3 expression (25). Another study demonstrated that p53
activity-mediated TNF-α induces apoptosis and inhibits
differentiation in cementoblasts (26). In addition, p38, Erk1/2, JNK,
PI3K-Akt and NF-κB pathways are activated to limit the role of
TNF-α in inducing apoptosis of cementoblasts (26). As aforementioned, NTR1 is verified
to have anti-inflammatory effects. According to a previous study,
NTR1 can increase cell viability, suppress the expression of
inflammatory cytokines and reverse lipopolysaccharide-induced
activation of the NF-κB signaling pathway in myocardial cells
(15). It has also been shown that
NTR1 inhibits the expression levels of TNF-α and IL-1β to protect
endothelial cells from oxLDL-induced injury through the suppression
of the MAPK and NF-κB signaling pathways (8). Nevertheless, whether NTR1 could
abrogate the negative effects of TNF-α and promote cell
differentiation in HABOs remains to be elucidated.
As a consequence, the present study first performed
cytotoxicity tests to confirm the influence of NTR1 on cell
viability. The concentration of NTR1 influenced the effect of NTR1
on cell viability. NTR1 at ≤20 µmol/l had no effect on HAOBs,
whereas 40 µmol/l NTR1 slightly decreased the viability of HAOBs.
Therefore, the NTR1 dose in subsequent experiments was limited to
2.5 and 20 µmol/l. Pre-treated osteoblasts were stimulated by
TNF-α, which mimicked the inflammatory microenvironment in
vitro. Osteoblast differentiation and mineralization were
detected using an ALP activity assay and Alizarin Red S staining,
respectively (26,27). RUNX2 has been demonstrated to
induce osteoblast differentiation and, simultaneously, OPN and OCN
are defined as mineralization-related genes (28). In the present study, suppression of
mineralization caused by TNF-α was largely overcome by NTR1 in
HAOBs. Furthermore, under inflammatory conditions, the ALP activity
was higher, and the expression levels of osteogenic genes RUNX2,
OPN and OCN were higher, in cells treated with NTR1 compared with
the TNF-α stimulation group. All these results indicated that NTR1
might regulate osteoblast differentiation by reversing the negative
effects of TNF-α.
Previous studies have shown that the NF-κB signaling
pathway serves an important role in the transcription process of
inflammatory genes p50, p-p65 and p65 and the increased p65/p50
dimerization can induce gene transcription of pro-inflammatory
factors (TNF-α) (29). NTR1 has
anti-inflammatory functions by activating the NF-κB signaling
pathway in H9c2 cardiomyocytes (15) and thus the present study
investigated whether it has a similar effect in human alveolar bone
osteoblasts and the results were encouraging. It was identified
that the expression levels of p50, p-p65 and p-p65/p65 in
osteoblasts pre-treated with NTR1 were decreased in a
time-dependent manner compared with the TNF-α stimulation group.
However, no marked influence of NTR1 on HAOBs was observed. The
experimental results demonstrated that the TNF-α-activated NF-κB
signaling pathway was abrogated by NTR1.
The Wnt/β-catenin signaling pathway is of great
significance in bone development and regeneration and changes in
the pathway involves various genes (16,18,19,30).
DKK1 inhibits the Wnt pathway, while β-catenin affects the
initiation of target gene transcription and AXIN2 regulates the
intensity and duration of Wnt/β-catenin pathway (31). In the present study, TNF-α hampered
the Wnt/β-catenin signaling pathway in alveolar osteoblasts by
increasing DKK1 and decreasing AXIN2 and β-catenin, while NTR1
markedly recovered this pathway by downregulating the expression of
DKK1 and upregulating the expressions of AXIN2 and β-catenin in the
inflammatory environment created by TNF-α. Therefore, it can be
inferred that NTR1 potentially promoted alveolar osteoblast
differentiation involving the regulation towards the Wnt/β-catenin
signaling pathway.
Previous studies show that the fundamental part of
dental implantation is osseointegration where osteoblast growth is
of vital importance (4,5). Inflammation is one of the necessary
stages in dental implantation (32). The present study identified that
NTR1 was able to reverse inflammatory responses and could promote
alveolar osteoblast differentiation in vitro, which lays the
foundation for experiments in vivo.
In conclusion, NTR1 demonstrated the ability to
promote the differentiation of HAOBs by antagonizing the activation
of NF-κB signaling pathway and the inhibition of Wnt/β-catenin
signaling pathway, both of which were induced by TNF-α. Thus, NTR1
has the potential to be used to promote bone generation in the
treatment of dental implantation or other dental diseases,
highlighting the practical significance of NTR1 in clinic. For
example, NTR1 could be used as an ingredient in toothpaste for
daily care, managing inflammation and activating alveolar bone
regeneration.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LH and QL conceptualized and designed the study,
collected, analyzed and interpreted data, and drafted and
critically revised the article for important intellectual content.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All procedures performed in studies involving human
participants were in accordance with the ethical standards of the
institutional and/or national research committee and with the 1964
Helsinki declaration and its later amendments or comparable ethical
standards. The current study was approved by the Ethics Committee
of the Jingmen Number 1 People's Hospital (Jingmen, China; approval
no. JMPH20160123). All patients signed informed consent and agreed
that their tissues would be used for clinical research.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Akintoye SO: The distinctive jaw and
alveolar bone regeneration. Oral Dis. 24:49–51. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Monje A, Chan HL, Galindo-Moreno P,
Elnayef B, Suarez-Lopez del Amo F, Wang F and Wang HL: Alveolar
bone architecture: A systematic review and meta-analysis. J
Periodontol. 86:1231–1248. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Araujo MG, Silva CO, Misawa M and Sukekava
F: Alveolar socket healing: What can we learn? Periodontology 2000.
68:122–134. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Branemark PI: Osseointegration and its
experimental background. J Prosthet Dent. 50:399–410. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Smeets R, Stadlinger B, Schwarz F,
Beck-Broichsitter B, Jung O, Precht C, Kloss F, Grobe A, Heiland M
and Ebker T: Impact of dental implant surface modifications on
osseointegration. Biomed Res Int. 2016:62856202016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen FM: Periodontal tissue engineering
and regeneration. Zhonghua Kou Qiang Yi Xue Za Zhi. 52:610–614.
2017.(In Chinese). PubMed/NCBI
|
|
7
|
Kalsi H, Rodriguez JM, Darbar U and
Bavisha K: The emergency dental appointment: Restorative
emergencies part 2-dental implant related problems. Prim Dent J.
6:62–70. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Su P, Du S, Li H, Li Z, Xin W and Zhang W:
Notoginsenoside R1 inhibits oxidized low-density lipoprotein
induced inflammatory cytokines production in human endothelial
EA.hy926 cells. Eur J Pharmacol. 770:9–15. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu Z, Wang H, Hou G, Cao H, Zhao Y and
Yang B: Notoginsenoside R1 protects oxygen and glucose
deprivation-induced injury by upregulation of miR-21 in
cardiomyocytes. J Cell Biochem. 120:9181–9192. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yu Y, Sun G, Luo Y, Wang M, Chen R, Zhang
J, Ai Q, Xing N and Sun X: Cardioprotective effects of
Notoginsenoside R1 against ischemia/reperfusion injuries by
regulating oxidative stress- and endoplasmic reticulum
stress-related signaling pathways. Sci Rep. 6:217302016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang B, Zhang J, Zhang C, Zhang X, Ye J,
Kuang S, Sun G and Sun X: Notoginsenoside R1 protects against
diabetic cardiomyopathy through activating estrogen receptor alpha
and its downstream signaling. Front Pharmacol. 9:12272018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu Y, Lin Z, Guo J, Xu G, Li Y, Xu T, Lv
H, Chen J and Wu G: Notoginsenoside R1 significantly promotes in
vitro osteoblastogenesis. Int J Mol Med. 38:537–544. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao S, Yan L, Li X, Zhang Z, Sun Y and
Wang J: Notoginsenoside R1 suppresses wear particle-induced
osteolysis and RANKL mediated osteoclastogenesis in vivo and in
vitro. Int Immunopharmacol. 47:118–125. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang J, Ding L, Wang B, Ren G, Sun A,
Deng C, Wei X, Mani S, Wang Z and Dou W: Notoginsenoside R1
attenuates experimental inflammatory bowel disease via pregnane X
receptor activation. J Pharmacol Exp Ther. 352:315–324. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhong L, Zhou XL, Liu YS, Wang YM, Ma F,
Guo BL, Yan ZQ and Zhang QY: Estrogen receptor α mediates the
effects of notoginsenoside R1 on endotoxin-induced inflammatory and
apoptotic responses in H9c2 cardiomyocytes. Mol Med Rep.
12:119–126. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim W, Kim M and Jho EH: Wnt/β-catenin
signalling: From plasma membrane to nucleus. Biochem J. 450:9–21.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yao Q, Yu C, Zhang X, Zhang K, Guo J and
Song L: Wnt/β-catenin signaling in osteoblasts regulates global
energy metabolism. Bone. 97:175–183. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Duan P and Bonewald LF: The role of the
wnt/β-catenin signaling pathway in formation and maintenance of
bone and teeth. Int J Biochem Cell Biol. 77:23–29. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rossini M, Gatti D and Adami S:
Involvement of WNT/β-catenin signaling in the treatment of
osteoporosis. Calcif Tissue Int. 93:121–132. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang X, Chen Q, Liu J, Fan C, Wei Q, Chen
Z and Mao X: Parthenolide promotes differentiation of osteoblasts
through the Wnt/β-catenin signaling pathway in inflammatory
environments. J Interferon Cytokine Res. 37:406–414. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li XT, Ma W, Wang XB, Liang Z, Yang JW, He
Y, Dai YF, Zhang T, Wang JW, Liu KP, et al: Notoginsenoside R1
promotes the growth of neonatal rat cortical neurons via the
Wnt/β-catenin signaling pathway. CNS Neurol Disord Drug Targets.
17:547–556. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Coates DE, Zafar S and Milne TJ:
Quantitative real-time gene profiling of human alveolar
osteoblasts. Methods Mol Biol. 1537:447–459. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zelova H and Hosek J: TNF-α signalling and
inflammation: interactions between old acquaintances. Inflamm Res.
62:641–651. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Giannoudis PV, Hak D, Sanders D, Donohoe
E, Tosounidis T and Bahney C: Inflammation, bone healing, and
anti-inflammatory drugs: An Update. J Orthop Trauma. 29 (Suppl
12):S6–S9. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jeong BC: ATF3 mediates the inhibitory
action of TNF-alpha on osteoblast differentiation through the JNK
signaling pathway. Biochem Biophys Res Commun. 499:696–701. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang YL, He H, Liu ZJ, Cao ZG, Wang XY,
Yang K, Fang Y, Han M, Zhang C and Huo FY: Effects of TNF-α on
cementoblast differentiation, mineralization, and apoptosis. J Dent
Res. 94:1225–1232. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Goseki M, Oida S, Takeda K, Ogata Y,
Iimura T, Maruoka Y and Sasaki S: Identification of bone-type
alkaline phosphatase mRNA from human periodontal ligament cells. J
Dent Res. 74:319–322. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Komori T: Regulation of osteoblast
differentiation by transcription factors. J Cell Biochem.
99:1233–1239. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Janus P, Pakuła-Cis M, Kalinowska-Herok M,
Kashchak N, Szołtysek K, Pigłowski W, Widlak W, Kimmel M and Widlak
P: NF-κB signaling pathway is inhibited by heat shock independently
of active transcription factor HSF1 and increased levels of
inducible heat shock proteins. Genes Cells. 16:1168–1175. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ikehata M, Yamada A, Morimura N, Itose M,
Suzawa T, Shirota T, Chikazu D and Kamijo R: Wnt/β-catenin
signaling activates nephronectin expression in osteoblasts. Biochem
Biophys Res Commun. 484:231–234. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tortelote GG, Reis RR, de Almeida Mendes F
and Abreu JG: Complexity of the Wnt/β-catenin pathway: Searching
for an activation model. Cell Signal. 40:30–43. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liaw K, Delfini RH and Abrahams JJ: Dental
implant complications. Semin Ultrasound CT MR. 36:427–433. 2015.
View Article : Google Scholar : PubMed/NCBI
|