Introduction
It has been reported that 8–34% of patients
experience chronic postsurgical pain (CPSP) following surgery,
which results in a decrease in the quality of postoperative daily
life (1). This is a difficult
clinical problem. The mechanism underlying the transformation of
acute pain to chronic pain following tissue damage remains to be
elucidated. An improved understanding of this mechanism is the key
for clinically preventing CPSP.
Growing evidence has indicated that nerve damage
causes the resting astrocytes of the spinal cord to enter into an
active state, which is necessary for the development and
maintenance of pain (2–4). The astrocytes in the central nervous
system (CNS) form a highly interconnected network via gap junctions
or adherens junctions (5). In some
pain models, there is an increase in the number of gap junction
channels and a series of junction proteins, such as connexin43
(Cx43) and p120, leading to the secretion of various types of
cytokines that are involved in inducing pain (6). The role of Cx43 in chronic pain has
been studied previously (7–9). The
purpose of the present study was to explore the role of p120 in the
maintenance of CPSP by detecting the expression of p120 in the DRG
and spinal cord of rats with CPSP.
Our previous studies have shown that adenosine
triphosphate-sensitive potassium channel (KATP) agonists may
relieve postoperative pain in rats by inhibiting the apoptosis of
vascular endothelial cells (10,11).
KATP agonists can inhibit touch-induced pain in a dose-dependent
manner (11). Notably, nicorandil
is the first clinical KATP opener, which has been reported to
exhibit activities in models of inflammatory and nociceptive pain
(12,13). The present study attempted to
extend the investigation on the antinociceptive activity of
nicorandil by evaluating its effects on the expression of p120 and
also to evaluate the possible mechanisms involved in the
antinociceptive activity of this drug through the p120 pathway.
Materials and methods
Animals
A total of 42 male Sprague-Dawley rats (weight,
200–250 g; age, 8–10 weeks) were provided water and feed ad
libitum until 3 days prior to the experiment. The rats were
house with a l2 h light/dark cycle at 23±1°C and 55–60% humidity.
The rats were provided by the Experimental Animal Center of Nantong
University (Nantong, China) and the study procedures were approved
by the Animal Care and Use Committee of Nantong University
(approval no. 20171015S1051122).
Experimental design and drugs
The rats were randomly divided into seven groups
(n=6/group). Naive group, no treatment was performed; sham group,
the rats received an incision through the skin and muscle;
skin/muscle incision and retraction (SMIR) (14) group, the rats underwent 1-h
retraction following the skin/muscle incision; SMIR + nicorandil
groups, referring to the dose of nicorandil in the relevant
literature (15–17), the rats were intraperitoneally
injected with either low (1.0 mg/kg), middle (1.5 mg/kg) or high
dose nicorandil (2.0 mg/kg) separately 0.5 h prior to the SMIR
procedure; SMIR + nicorandil (1.5 mg/kg) + glibenclamide (10.0
mg/kg) group, 0.5 h before SMIR modeling, a mixture of 1.5 mg/kg
nicorandil (cat. no. N3539; Sigma-Aldrich; Merck KGaA) and 10.0
mg/kg glibenclamide (cat. no. PHR1287; Sigma-Aldrich; Merck KGaA)
was injected intraperitoneally.
Behavioral testing
The mechanical withdrawal threshold (MWT) was
detected prior to, and 1, 3, 7 and 14 days following SMIR surgery.
The rats were habituated to the testing environment for at least 30
min before testing. Mechanical allodynia was assessed using the
Up-Down paradigm (18) with von
Frey filaments (IITC Life Science, Inc.) ranging between 1.4 and 26
g. Shrinking, swinging or paw licking were regarded as positive
reactions. Each filament was presented five times within 30 sec to
determine the response threshold. If the response was not elicited
at least twice, the next ascending von Frey filament was applied
until at least two responses were observed.
Immunofluorescence staining
On day 7 after SMIR, the rats were anesthetized with
isoflurane (induction with 3–4%, maintenance with 1–2%) and
perfused transcardially with PBS followed by 4% paraformaldehyde in
PBS (250 ml; pH 7.0). After perfusion, the L3-L5 spinal cord and
DRG tissues from rats in the SMIR group were extracted and
post-fixed in the same fixative at 4°C overnight, and then placed
in 20% and subsequently in 30% sucrose solution at 4°C overnight.
After freezing sequentially, the tissues were continuously
sectioned at 6 µm and stored at −20°C. These sections were selected
randomly and blocked with 5% serum antibody blocking solution
(Beyotime Institute of Biotechnology) for 2 h. The tissue slices
were then incubated independently with antibodies against p120
(1:300; Santa Cruz Biotechnology, Inc; cat. no. sc-23873), glial
fibrillary acidic protein (GFAP; 1:5,000; EMD Millipore; cat. no.
AG230), calcitonin gene-related peptide (CGRP; 1:800; Cell
Signaling Technology, Inc.; cat. no. 14959), NeuN (1:5,000; EMD
Millipore; cat. no. SAB4300883), ionized calcium-binding adapter
molecule 1 (Iba1; 1:800; FUJIFILM Wako Pure Chemical Corporation;
cat. no. 019-19741) and isolectin B4 (IB4; 1:1,000; Advanced
Targeting Systems, Inc.; cat. no. PR-02) at 4°C overnight, then
co-incubated with Cy3- or FITC-conjugated secondary antibodies
(1:1,000, Jackson ImmunoResearch Laboratories, Inc.; cat. no.
115-165-205, 115-095-205) in the dark for 2 h at room temperature.
A total of five sections were randomly selected from the spinal
cord and DRG of each rat. The localizations of p120 in the spinal
cord dorsal horn and DRG were examined under a fluorescence
microscope (Olympus Corporation) in the dark to capture images.
Western blotting
The rats were anesthetized and sacrificed as
previously described, and the L3-L5 spinal cord and DRG tissues
were homogenized in sodium dodecyl sulfate (SDS) sample buffer
containing a mixture of protease and phosphatase inhibitors
(Sigma-Aldrich; Merck KGaA), and measured with a bicinchoninic acid
protein assay kit (Beyotime Institute of Biotechnology). For
separation, 30 µg total protein per gel lane was loaded onto 10%
gels (Beyotime Institute of Biotechnology). The separated proteins
were then transferred onto nitrocellulose membranes. The membranes
were incubated for 2 h at room temperature in tris-buffered saline
and Tween-20 blocking solution containing 5% skimmed milk, followed
by overnight incubation at 4°C in blocking solution containing
primary antibodies against the following proteins: p120 (1:300;
Santa Cruz Biotechnology, Inc.; cat. no. sc-23873) and GAPDH
(1:5,000; Sigma-Aldrich; Merck KGaA; cat. no. G2267). Membranes
were washed three times and incubated with the anti-mouse or
anti-rabbit peroxidase-conjugated secondary antibodies (1:2,000;
Jackson Immuno Research Laboratories, Inc.; cat. nos. 115-035-003,
111-005-003, respectively) at room temperature for 2 h. After
washing, immunolabeling was detected using the Tanon2500 gel
imaging system (Tanon Science and Technology Co., Ltd.) and
hypersensitive ECL chemiluminescence detection kit (Absin). ImageJ
software (version no. 1.8.0, National Institutes of Health) was
used to capture and analyze the intensity of the bands.
Statistical analysis
The data were presented as mean ± standard error of
mean of at least three experimental repeats. Data were analyzed
using one- or two-way analysis of variance followed by Bonferroni's
or Dunnett's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
SMIR induces persistent mechanical
allodynia and nicorandil attenuates mechanical allodynia
To study the mechanism underlying CPSP, an SMIR
model was established according to a previous study (14) (Fig.
1A). As shown in Fig. 1B, the
MWT significantly decreased on postsurgical days 1, 3, 7 and 14 in
the sham, SMIR + 1.5 and SMIR + 2.0 mg/kg nicorandil groups, as
compared with the SMIR group, in a time-dependent manner. Whereas,
no statistical difference was observed in the MWT of the SMIR + 1.0
mg/kg nicorandil group compared with the SMIR group (P>0.05). It
was identified that the anti-nociceptive effects of 1.5 and
2.0-mg/kg nicorandil were more pronounced compared with 1.0 mg/kg
nicorandil (P<0.05), and no significant difference was observed
between 1.5 and 2.0 mg/kg (P>0.05; Fig. 1B). These results suggested that 1.5
and 2.0 mg/kg nicorandil may inhibit SMIR-induced mechanical
hyperalgesia, and that 1.5 mg/kg nicorandil may be the most
effective dose. Therefore, 1.5 mg/kg nicorandil was used to
determine whether the anti-nociceptive effects of a KATP activator
could be relieved by the KATP blocker glibenclamide (10.0 mg/kg).
As expected, injection of glibenclamide together with nicorandil
reversed the anti-mechanical nociceptive effects of nicorandil on
postsurgical days 1, 3, 7 and 14. These findings suggested that
nicorandil may alleviate CPSP through activating KATP.
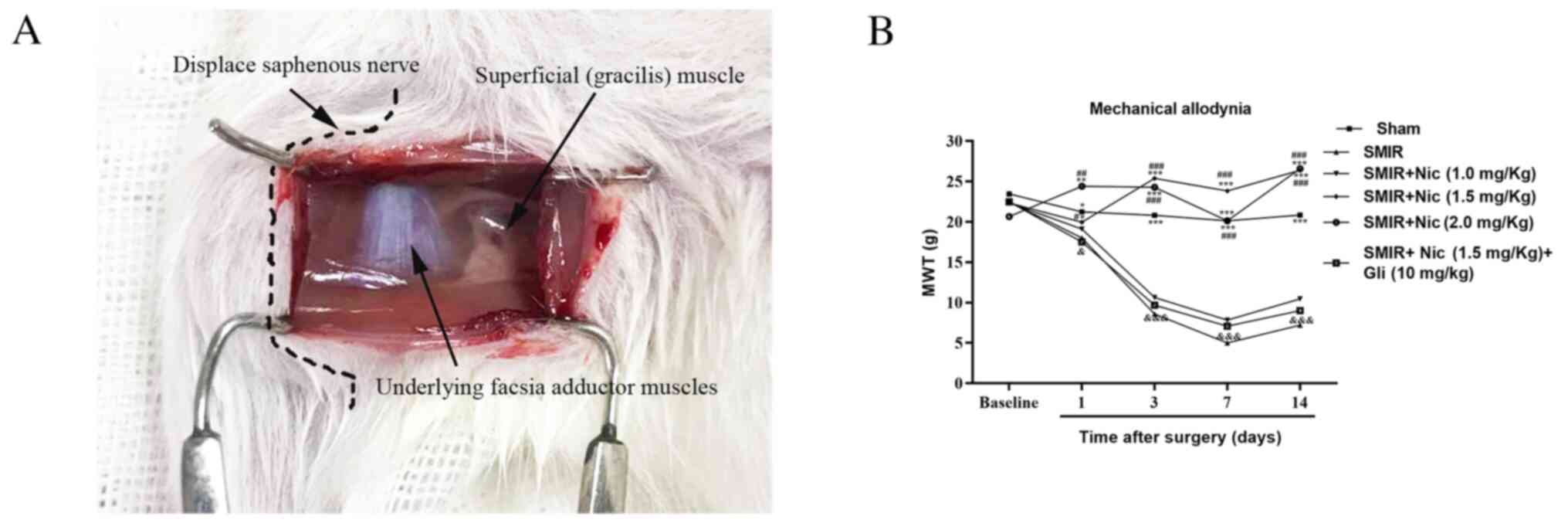 | Figure 1.Establishment of the SMIR model in
rats and detection of MWT. (A) SMIR model was established; the
saphenous nerve was pulled and displaced. (B) Comparison of MWT in
the sham, SMIR, Nic-treated groups and SMIR + Nic (1.5 mg/kg) + Gli
group (n=6). *P<0.05, **P<0.01, ***P<0.001 vs. SMIR group;
#P<0.05, ##P<0.01,
###P<0.001 vs. SMIR + Nic (1.0 mg/kg) group;
&P<0.05, &&&P<0.001 vs.
SMIR + Nic (1.5 mg/kg) group. SMIR, skin/muscle incision and
retraction; MWT, mechanical withdrawal threshold; Nic, nicorandil;
Gli, glibenclamide. |
Effects of CPSP on expression of p120
in the spinal cord and DRG
To explore the role of p120 in central sensitization
of CPSP and its possible mechanism, changes in the expression of
p120 in the spinal cord and DRG were examined on postsurgical days
3, 7 and 14 in the SMIR and sham groups. It was identified that
SMIR-induced CPSP enhanced the expression of p120 in the spinal
cord and DRG of rats compared with in the naive group (Fig. 2A and B). There was also a high
expression of p120 on postsurgical days 7 and 14 in the sham group
(Figs. S1 and S2). As the 7th day after surgery may be
the turning point of the transformation from acute pain to chronic
pain, the present study selectively compared the expression of p120
on postsurgical day 7 in the sham group with that in the SMIR
group. The results demonstrated that the expression of p120 in the
spinal cord of the SMIR group was higher compared with that of the
sham group on postsurgical day 7, whereas no significant difference
was observed in DRG between the two groups (Fig. 2A and B).
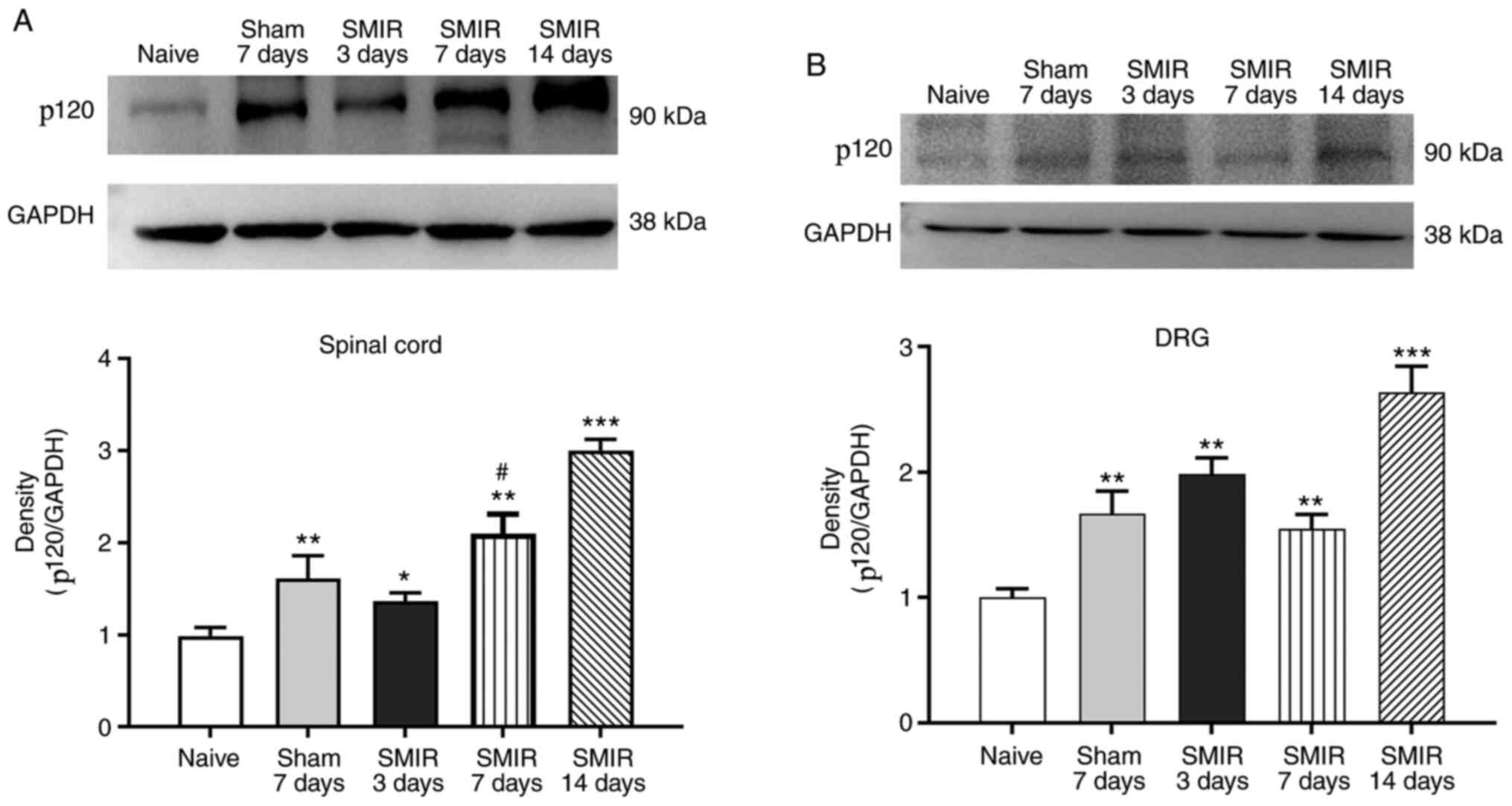 | Figure 2.Expression of p120. (A) In the spinal
cord, the expression of p120 was significantly higher in the SMIR
group compared with in the naive group on postsurgical days 3, 7
and 14. In the sham group, the expression of p120 increased
significantly on postsurgical day 7. Compared with on postsurgical
day 7 in the sham group, the SMIR group exhibited significantly
increased expression of p120 on postsurgical day 7. (B) In the DRG,
compared with the naive group, SMIR increased p120 expression on
postsurgical days 3, 7 and 14. In the sham group, p120 expression
increased significantly on postsurgical day 7. No significant
difference was observed between the sham and SMIR groups on
postsurgical day 7 (P>0.05). *P<0.05, **P<0.01,
***P<0.001 vs. naive group; #P<0.05 vs. sham
group. SMIR, skin/muscle incision and retraction; DRG, dorsal root
ganglion. |
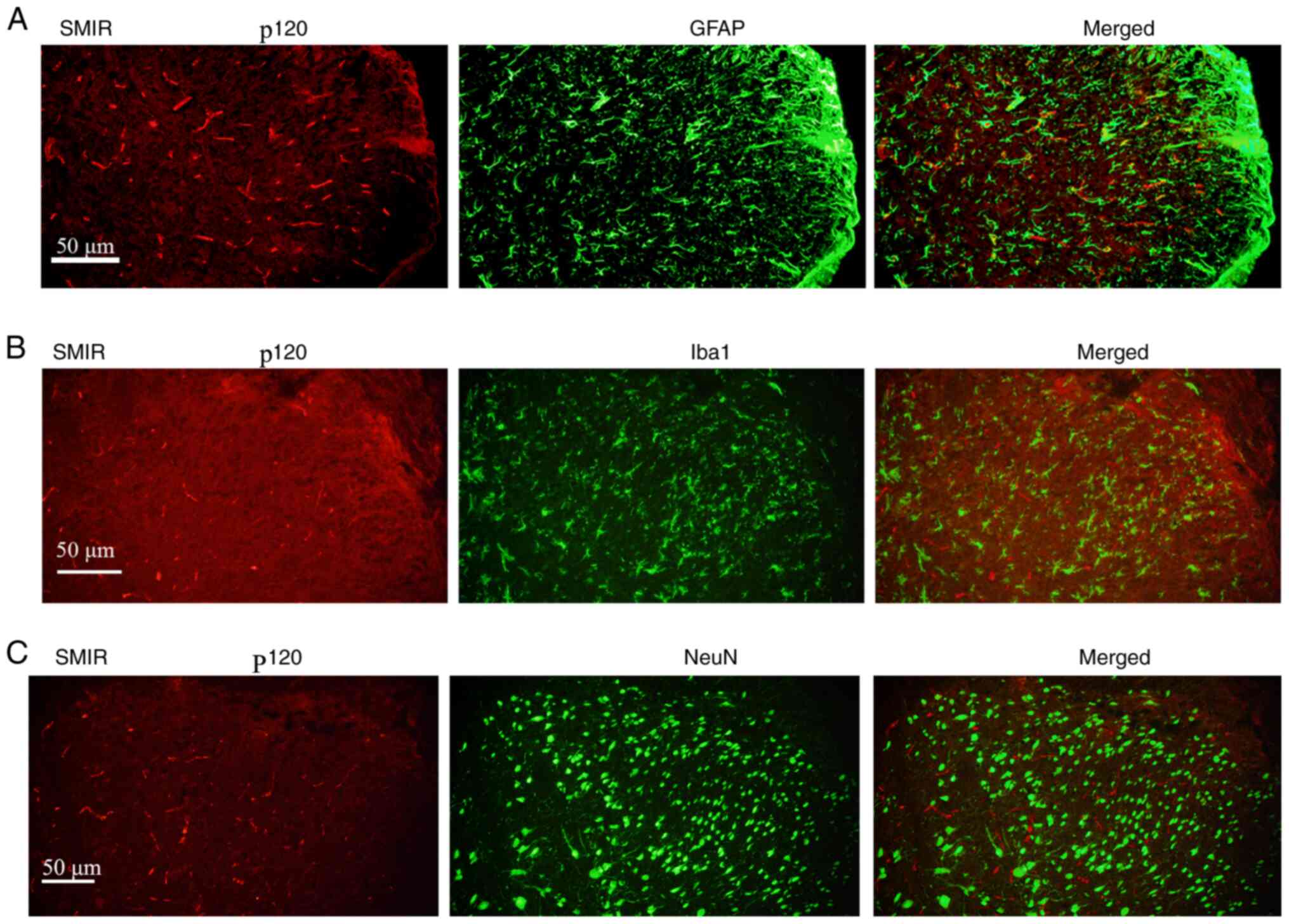
Immunofluorescence staining depicts
localization of p120 in the spinal cord
To study the location of p120, spinal cord sections
of rats in the SMIR group at postsurgical day 7 were stained with
the neuronal marker NeuN, astrocyte marker GFAP and microglia
marker Iba1. Results revealed that p120 was distributed in
GFAP-positive astrocytes. However, p120-positive staining was not
observed in the Iba1- and NeuN-positive neurons (Fig. 3A-C).
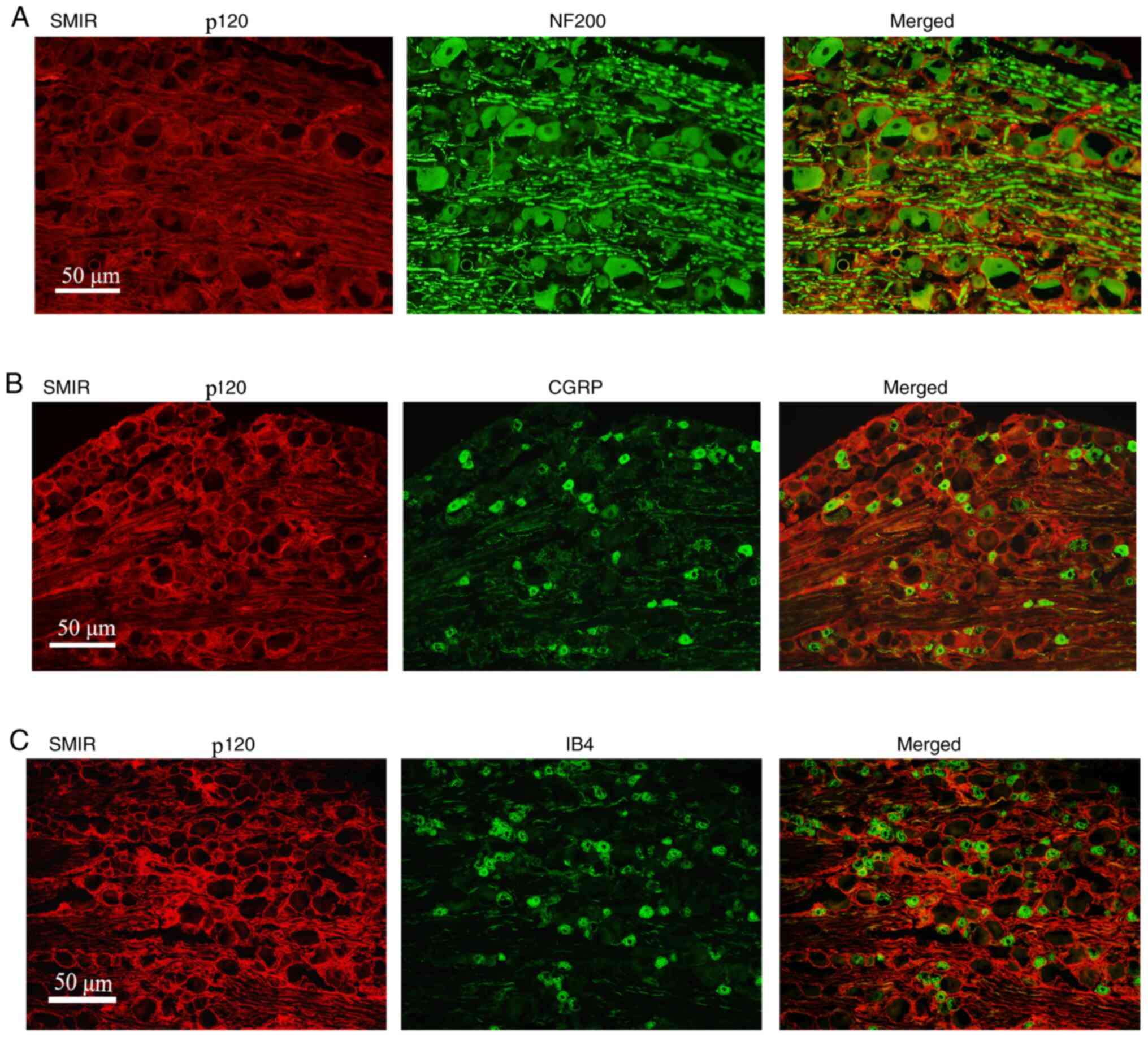
Immunofluorescence staining
demonstrates the localization of p120 in the DRG
At postsurgical day 7, DRG tissue sections were
stained with p120, and the medium and large neuronal marker NF200,
or the medium and small neuronal markers CGRP and IB4. The results
revealed that p120 was mainly distributed in the NF200-positive
medium and large neurons, but not in the CGRP- and IB4-positive
medium and small neurons (Fig.
4A-C).
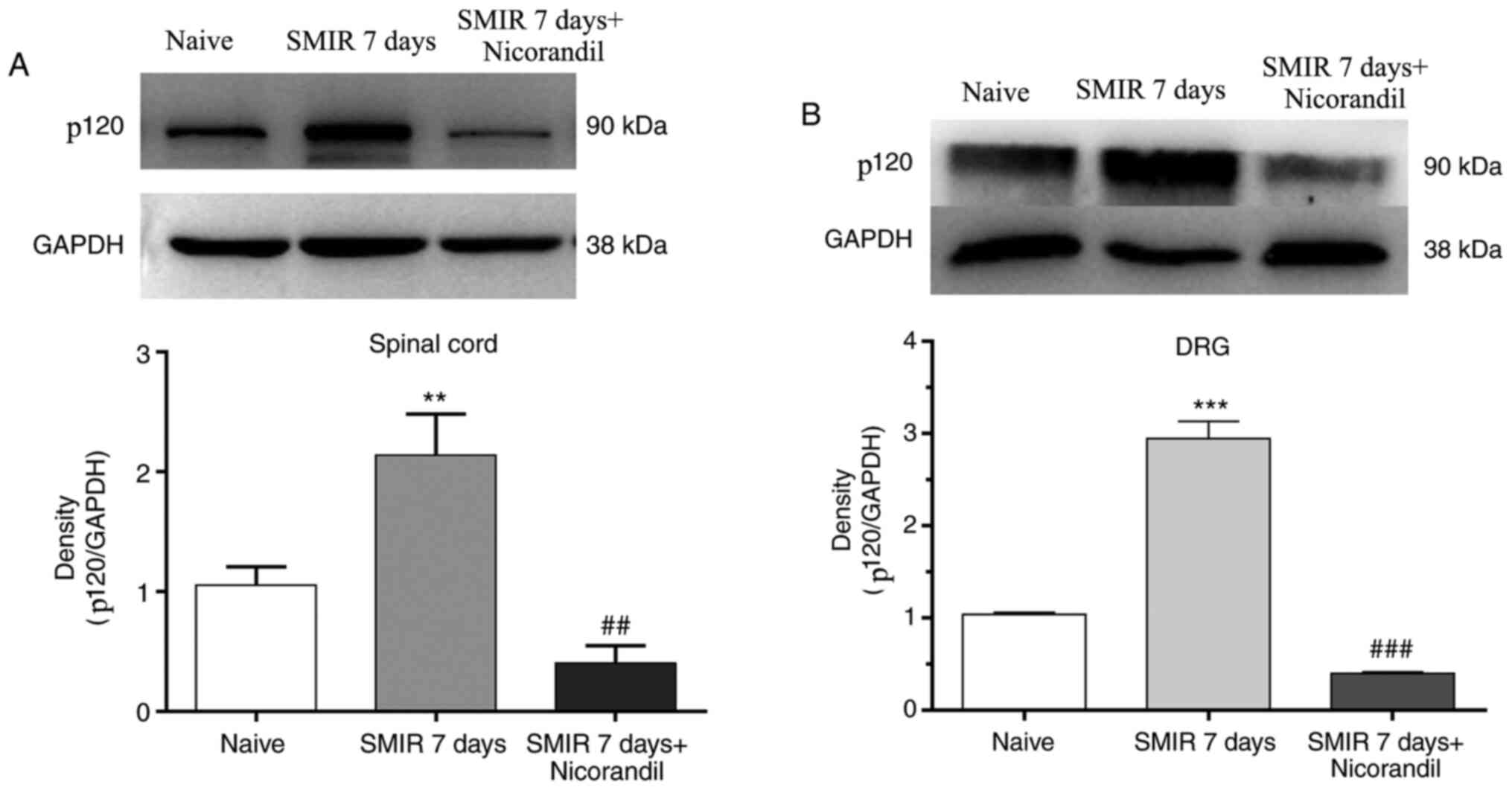
Effect of nicorandil on changes in
expression of p120 in the spinal cord and DRG
To study the mechanism underlying the analgesic
effect of KATP agonists, the present study detected the changes in
the expression of p120 in the spinal cord and DRG following
intraperitoneal injection of 1.5 mg/kg nicorandil 0.5 h before
SMIR. The changes in the expression of p120 were observed on
postsurgical day 7. The results suggested that SMIR promoted the
expression of p120 in the spinal cord and DRG, whereas nicorandil
treatment decreased the expression of p120 (Fig. 5A and B).
Discussion
In 2008, Flatters et al (14) developed the SMIR model to study
CPSP. In the current study, MWT was significantly decreased
following SMIR in a time-dependent manner compared with the sham
group, indicating that the pain model was effective. Recently, the
prevalence of CPSP has been increasingly regarded as a public
health problem; thus, the present study established SMIR,
simulating the state of CPSP in rats, to explore the potential
mechanism of action of nicorandil in relieving pain.
The astrocytes in the spinal cord are considered to
be the key point for central sensitization (19,20),
which is the main mechanism of pain induction (21–23).
Astrocytes in the CNS form a highly interconnected network via
complexes formed by connexins, such as p120 and Cx43 (5,24).
These complexes are involved in the regulation of cell adhesion,
migration, invasion and proliferation (25). In some pain models, the increased
expression of a series of connexins, especially Cx43, leads to the
secretion of various types of cytokines, and participates in the
occurrence and development of pain (26,27).
A previous study examined the role of Cx43 in chronic pain
(28); however, to the best of our
knowledge, the relationship between p120 and chronic pain remains
to be elucidated. p120 is known to exhibit a role in adhesion among
cells and in signal transduction in cells (29–32).
It has also been reported that p120 protects against post-traumatic
inflammation by regulating the adherens and tight gap junctions
(33,34). When the body is subjected to
peripheral stimulation, the DRG first senses this change and
activates the DRG neurons, which then transmit to the dorsal horn
of the spinal cord, causing abnormal neuronal excitability, which
leads to the sensation of pain in the brain (35). Therefore, the present study
selectively detected changes in the expression of p120 in the
spinal cord and DRG of rats with CPSP, and examined the
localization of p120 in the two tissues. Results revealed that SMIR
increased the expression of p120 in the DRG and spinal cord
compared with in the naive group. Furthermore, p120 was mainly
identified to be distributed in GFAP-positive astrocytes in the
spinal cord, and in the NF200-positive medium and large neurons in
the DRG. The interaction between the astrocytes and neurons may
regulate synaptic transmission through p120, which contributes to
central sensitization of spinal cord pain signals (36), indicating that p120 may be involved
in the maintenance of CPSP through astrocyte-neuron
interaction.
It has been demonstrated that KATP channels serve
critical roles in regulating membrane excitability and
neurotransmitter release, and in providing neuroprotection
(37,38). Our previous studies demonstrated
that KATP agonists can significantly reduce postoperative pain in
rats by inhibiting the apoptosis of vascular endothelial cells
around the incision (10,11). The KATP activator nicorandil is a
nitric oxide donor approved for the treatment of patients with
angina pectoris, which also exhibits activities in models of
inflammatory and nociceptive pain (12,13).
Regarding the cardiovascular actions of nicorandil, those that have
been investigated more extensively, it has been concluded that this
drug exhibits important differences when compared to the
traditional organic nitrates, possibly due to its ability to open
potassium channels (39). The
present study provided an additional demonstration of the
antinociceptive activity of nicorandil by demonstrating its
activity in an SMIR model. It was identified that the
antinociceptive effect induced by nicorandil was markedly
attenuated by the KATP blocker glibenclamide, providing solid
evidence of an important mechanism mediating the activity of
nicorandil. In addition, the present study identified that
nicorandil could decrease the expression of p120 in the DRG and
spinal cord. Combined with the effects of nicorandil on the
expression of p120, it was hypothesized that the antinociceptive
activity of nicorandil may depend on the p120 pathway, thus
expanding the scope of the study of the analgesic effects of
nicorandil.
In summary, high expression of p120 may be the key
link in central sensitization of CPSP. The regulation of p120
through targeted intervention by KATP agonists may be an effective
strategy to inhibit central sensitization and may form the basis of
potential therapeutic approaches for the treatment of CPSP.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81701106).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
SSH and JPY designed the study. SSH, SC and CEL
acquired and interpreted the data. YBQ analyzed the data and
assisted JPY in revising the manuscript. SSH prepared the
manuscript and supervised the study. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
All experiments in the current study were approved
by the Experimental Animal Protection and Care Committee of the
Nantong University (approval no. 20171015S1051122).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kim DH, Pearson-Chauhan KM, McCarthy RJ
and Buvanendran A: Predictive factors for developing chronic pain
after total knee arthroplasty. J Arthroplasty. 33:3372–3378. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Christensen RK, Delgado-Lezama R, Russo
RE, Lind BL, Alcocer EL, Rath MF, Fabbiani G, Schmitt N, Lauritzen
M, Petersen AV, et al: Spinal dorsal horn astrocytes release GABA
in response to synaptic activation. J Physol. 596:4983–4994.
2018.
|
|
3
|
Wang S, Deng J, Fu H, Guo Z, Zhang L and
Tang P: Astrocytes directly clear myelin debris through endocytosis
pathways and followed by excessive gliosis after spinal cord
injury. Biochem Biophys Res Commun. Feb 15–2020.doi:
10.1016/j.bbrc.2020.02.069 (Epub ahead of print).
|
|
4
|
Nakagawa T and Kaneko S: Spinal astrocytes
as herapeutic targets for pathological pain. J Pharmacol Sci.
114:347–353. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xing L, Yang T, Cui S and Chen G: Connexin
hemichannels in astrocytes: Role in CNS disorders. Front Mol
Neurosci. 12:232019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang A and Xu C: The role of connexin43 in
neuropathic pain induced by spinal cord injury. Acta Biochim
Biophys Sin (Shanghai). 51:555–561. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang H, Yan H, Li X, Liu J, Cao S, Huang
B, Huang D and Wu L: Inhibition of Connexin 43 and phosphorylated
NR2B in spinal astrocytes attenuates bone cancer pain in mice. J
Neurotrauma. 12:1292018.
|
|
8
|
Komiya H, Shimizu K, Ishii K, Kudo H,
Okamura T, Kanno K, Shinoda M, Ogiso B and Iwata K: Connexin 43
expression in satellite glial cells contributes to ectopic
tooth-pulp pain. J Oral Sci. 60:493–499. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Choi SR, Roh DH, Yoon SY, Kwon SG, Choi
HS, Han HJ, Beitz AJ and Lee JH: Astrocyte sigma-1 receptors
modulate connexin 43 expression leading to the induction of
below-level mechanical allodynia in spinal cord injured mice.
Neuropharmacology. 111:34–46. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shen S, Cao S, Huang S and Chen J: Effect
of adenosine triphosphate-sensitive potassium activation on
peripheral and central pain sensitization. J Surg Res. 195:481–487.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cao S, Qin Y, Chen J and Shen SR: Effects
of pinacidil on changes to the microenvironment around the incision
site, of a skin/muscle incision and retraction, in a rat model of
postoperative pain. Mol Med Rep. 12:829–836. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dutra MM, Godin AM, César IC, Nascimento
EB Jr, Menezes RR, Ferreira WC, Soares DG, Seniuk JG, Araújo DP,
Bastos LF, et al: Activity of nicorandil, a nicotinamide derivative
with a nitrate group, in the experimental model of pain induced by
formaldehyde in mice. Pharmacol Biochem Behav. 106:85–90. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dutra MM, Nascimento Júnior EB, Godin AM,
Brito AM, Melo IS, Augusto PS, Rodrigues FF, Araújo DP, de Fátima
Â, Coelho MM and Machado RR: Opioid pathways activation mediates
the activity of nicorandil in experimental models of nociceptive
and inflammatory pain. Eur J Pharmacol. 768:160–164. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Flatters SJ: Characterization of a model
of persistent postoperative pain evoked by skin/muscle incision and
retraction (SMIR). Pain. 135:119–130. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
He WK, Su Q, Liang JB, Wang XT, Sun YH and
Li L: Nicorandil pretreatment inhibits myocardial apoptosis and
improves cardiac function after coronary microembolization in rats.
J Geriatr Cardiol. 15:591–597. 2018.PubMed/NCBI
|
|
16
|
Ahmed LA, Salem HA, Attia AS and Agha AM:
Pharmacological preconditioning with nicorandil and pioglitazone
attenuates myocardial ischemia/reperfusion injury in rats. Eur J
Pharmacol. 663:51–58. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
He W, Su Q, Liang J, Sun Y, Wang X and Li
L: The protective effect of nicorandil on cardiomyocyte apoptosis
after coronary microembolization by activating Nrf2/HO-1 signaling
pathway in rats. Biochem Biophys Res Commun. 496:1296–1301. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dixon WJ: Staircase bioassay: The
up-and-down method. Neurosci Biobehav Rev. 15:47–50. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rhudy JL, Lannon EW, Kuhn BL, Palit S,
Payne MF, Sturycz CA, Hellman N, Güereca YM, Toledo TA, Huber F, et
al: Assessing peripheral fibers, pain sensitivity, central
sensitization and descending inhibition in native Americans: Main
findings from the oklahoma study of native American pain risk.
Pain. 161:388–404. 2020.PubMed/NCBI
|
|
20
|
Filbrich L, van den Broeke EN, Legrain V
and Mouraux A: The focus of spatial attention during the induction
of central sensitization can modulate the subsequent development of
secondary hyperalgesia. Cortex. 124:193–203. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ikeda H, Kiritoshi T and Murase K:
Contribution of microglia and astrocytes to the central
sensitization, inflammatory and neuropathic pain in the juvenile
rat. Mol Pain. 8:432012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gao YJ, Zhang L, Samad OA, Suter MR,
Yasuhiko K, Xu ZZ, Park JY, Lind AL, Ma Q and Ji RR: JNK-induced
MCP-1 production in spinal cord astrocytes contributes to central
sensitization and neuropathic pain. J Neurosci. 29:4096–4108. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zheng P, Jia S, Guo D, Chen S, Zhang W,
Cheng A, Xie W, Sun G, Leng J and Lang J: Central
sensitization-related changes in brain function activity in a rat
endometriosis-associated pain model. J Pain Res. 13:95–107. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mao Y, Tonkin RS, Nguyen T, O'Carroll SJ,
Nicholson LF, Green CR, Moalem-Taylor G and Gorrie CA: Systemic
administration of Connexin43 mimetic peptide improves functional
recovery after traumatic spinal cord injury in adult rats. J
Neurotrauma. 34:707–719. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu X, Li WE, Huang GY, Meyer R, Chen T,
Luo Y, Thomas MP, Radice GL and Lo CW: Modulation of mouse neural
crest cell motility by N-cadherin and connexin 43 gap junctions. J
Cell Biol. 154:217–230. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhou L, Ao L, Yan Y, Li C, Li W, Ye A, Liu
J, Hu Y, Fang W and Li Y: Levo-corydalmine attenuates
vincristine-induced neuropathic pain in mice by upregulating the
Nrf2/HO-1/CO pathway to inhibit Connexin 43 expression.
Neurotherapeutics. 17:340–355. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vicario N, Pasquinucci L, Spitale FM,
Chiechio S, Turnaturi R, Caraci F, Tibullo D, Avola R, Gulino R,
Parenti R and Parenti G: Simultaneous activation of mu and delta
opioid receptors reduces allodynia and astrocytic Connexin 43 in an
animal model of neuropathic pain. Mol Neurobiol. 56:7338–7354.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu J and Huang D: Progress in gap
junction protein 43 in chronic pain. Zhong Nan Da Xue Xue Bao Yi
Xue Ban. 43:95–99. 2018.(In Chinese). PubMed/NCBI
|
|
29
|
Zhang Y, Jiao H, Wu Y and Sun X:
P120-catenin regulates pulmonary fibrosis and TGF-β induced lung
fibroblast differentiation. Life Sci. 230:35–44. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Garrett JP, Lowery AM, Adam AP, Kowalczyk
AP and Vincent PA: Regulation of endothelial barrier function by
p120-catenin-VE-cadherin interaction. Mol Biol Cell. 28:85–97.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xie Z, Tang Y, Man MQ, Shrestha C and
Bikle DD: p120-catenin is required for regulating epidermal
proliferation, differentiation and barrier function. J Cell
Physiol. 234:427–432. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wehrendt DP, Carmona F, González Wusener
AE, González Á, Martínez JM and Arregui CO: P120-catenin regulates
early trafficking stages of the N-cadherin precursor complex. PLoS
One. 11:e01567582016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gritsenko PG, Atlasy N, Dieteren CEJ,
Navis AC, Venhuizen JH, Veelken C, Schubert D, Acker-Palmer A,
Westerman BA, Wurdinger T, et al: p120-catenin-dependent collective
brain infiltration by glioma cell networks. Nat Cell Biol.
22:97–107. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yuan L and Arikkath J: Functional roles of
p120ctn family of proteins in central neurons. Semin Cell Dev Biol.
69:70–82. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Malcangio M: Role of the immune system in
neuropathic pain. Scand J Pain. 20:33–37. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chauvet N, Privat A and Prieto M:
Differential expression of p120 catenin in glial cells of the adult
rat brain. J Comp Neurol. 479:15–29. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Roper J and Ashcroft FM: Metabolic
inhibition and low internal ATP activate K-ATP channels in rat
dopaminergic substantia nigra neurones. Pflügers Arch. 430:44–54.
1995. View Article : Google Scholar
|
|
38
|
Yamada K and Inagaki N: Neuroprotection by
KATP channels. J Mol Cell Cardiol. 38:945–949. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ahmed LA: Nicorandil: A drug with ongoing
benefits and different mechanisms in various diseased conditions.
Indian J Pharmacol. 51:296–301. 2019. View Article : Google Scholar : PubMed/NCBI
|