Introduction
Acute myocardial infarction (AMI) is a common
cardiovascular disease that is the leading cause of morbidity and
mortality among 240 causes of death between 1990 and 2013 globally
across all ages and both sexes (1). Myocardial ischemia-hypoxia is the
primary cause of AMI (2), and
ischemia-hypoxia generates cellular stresses, such as oxidative
stress, that lead to irreversible myocardial injuries (3). Mitochondria are prone to cellular
stresses, including hypoxia, and mitochondrial dysfunction is
associated with the intrinsic pathway of apoptosis in
cardiomyocytes under hypoxia (3,4).
Thus, lowering oxidative damage may be a promising approach to
salvage ischemic myocardium (5,6).
Non-coding (nc)RNAs, comprising microRNAs (miRNA or
miR) and long ncRNAs (lncRNAs), are promising novel targets for the
treatment of cardiovascular diseases, including AMI (7). Abnormal ncRNA transcriptions have
been identified in ischemic-hypoxic heart and cardiomyocytes
(8,9). Moreover, ncRNAs are associated with
myocardial ischemia-reperfusion (I/R), the primary therapeutic
choice for AMI (10). In addition,
mutual interaction between lncRNAs, miRNAs and mRNA serves a key
role in transcriptional regulation following AMI, and a competing
endogenous RNA (ceRNA) network has been identified in patients with
AMI (11) as well as I/R-mediated
injury (12).
lncRNA FGD5 antisense RNA 1 (FGD5-AS1) is a
cancer-associated gene (13–15)
and is a key lncRNA in patients with AMI, due to its roles in
regulating >50 differentially expressed genes, according to the
GSE48060 dataset (11). miR-195-5p
is a notable oncogene in cancer (16), and has been identified as a cardio-
and stress-responsive miRNA that participates in cardiac remodeling
(17,18). Retinoid acid receptor-related
orphan receptor α (RORA) is an important circadian rhythm gene
(19) and defends against
cardiomyocyte injury in mouse heart cells (20).
The present study aimed to analyze the expression
levels of FGD5-AS1 and RORA in patients with AMI using the GSE48060
dataset, and to confirm their roles in a hypoxia-induced model of
AMI in human cardiomyocyte AC16 cells. The present study also aimed
to identify the association between FGD5-AS1, miR-195 and RORA in
regulating oxidative damage of hypoxic AC16 cells.
Materials and methods
In silico data analysis
The GSE48060 (ncbi.nlm.nih.gov/GSE48060) dataset was downloaded
(11). This cohort of blood
samples was isolated from 31 patients with AMI and 21 normal
controls. The expression levels of FGD5-AS1 and RORA mRNA were
compared between AMI and normal cases and results were plotted as a
scatter diagram.
Cell model of AMI
Human cardiomyocyte cell line AC16 (SCC109) was
purchased from EMD Millipore, and cultured in DMEM with 10% FBS
(both HyClone; Cytiva) at 37°C in normoxic conditions (21%
O2, 5% CO2 and 74% N2). For
hypoxia treatment, AC16 cells were starved in FBS-free DMEM for 12
h, and then incubated in hypoxic conditions (1% O2, 5%
CO2 and 94% N2) for 12 h.
Cell transfection
For overexpression, the full length of FGD5-AS1
(ENSG00000225733) and coding domain sequence of RORA (NM_134261)
were separately inserted into pEGFP-C1 (Clontech Laboratories,
Inc.), generating overexpression vectors oe-FGD5-AS1 and oe-RORA,
respectively; miR-195 (5′-UAGCAGCACAGAAAUAUUGGC-3′) and miR-NC
mimics (5′-GUCCAGUGAAUUCCCAG-3′) were provided by Sangon Biotech
Co., Ltd. For silencing, siRNA against RORA (isoform a) (si-RORA;
5′-TACGTGTGAAGGCTGCAAGGGC-3′) and FGD5-AS1 (si-FGD5-AS1;
5′-UUGGUCGUUGUCAACUUCCCA-3′), and anti-RNA against miR-195
(anti-miR-195; 5′-GCCAAUAUUUCUGUGCUGCUA-3′) were purchased from
Sangon Biotech Co., Ltd., as well as the negative controls (NC)
si-NC (5′-UUCUCCGAACGUGUCACGUTT-3′) and anti-NC
(5′-UUCUCCGAACGUGUCACGUTT-3′). The transfection was performed in
AC16 cells in 6-well plates using Lipofectamine® 3000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer's instructions. Briefly, 2 µg overexpression
vectors and 30 nM oligonucleotides diluted in Opti-MEM™
Medium (Invitrogen; Thermo Fisher Scientific, Inc.) were mixed with
3.5 µl Lipofectamine 3,000 reagent diluted in Opti-MEM Medium. The
transfection efficiency was assessed prior to hypoxia treatment,
following transfection for 36 h.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA in AC16 cells was extracted using RNAiso
Plus (Takara Bio, Inc.) prior to complementary DNA synthesis using
PrimeScript™ RT MasterMix kit (Invitrogen; Thermo Fisher
Scientific, Inc.). Reverse transcription was performed at 25°C for
10 min, 42°C for 15 min and 85°C for 5 min. The RNA expression
levels were analyzed using SYBR-Green (Takara Bio, Inc.) on a
7900HT Fast Real-Time PCR system (Thermo Fisher Scientific, Inc.)
and the following primers: FGD5-AS1 forward,
5′-AGAAGCGGAGGGGTGAAAAT-3′ and reverse, 5′-CCGCCTTATAGTTGGCCCTC-3′;
miR-195 forward, 5′-GTCTAGCAGCACAGAAATA-3′ and reverse,
5′-GTGCAGGGTCCGAGGT-3′; RORA forward, 5′-AAAAACATGGAGTCAGCTCCG-3′
and reverse, 5′-AGTGTTGGCAGCGGTTTCTA-3′; GAPDH forward,
5′-CAGTCACTACTCAGCTGCCA-3′ and reverse, 5′-GAGGGTGCTCCGGTAG-3′; U6
forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′; 18S ribosomal N1 RNA (18S rRNA)
forward, 5′-CTGGATACCGCAGCTAGGAA-3′ and reverse,
5′-GAATTTCACCTCTAGCGGCG-3′. The qPCR thermocycling conditions were
as follows: 95°C for 15 min, 40 cycles at 94°C for 15 sec, 60°C for
30 sec and 72°C for 60 sec. The results were calculated via the
2−∆∆Cq method (21),
and normalized to the expression levels of GAPDH (for FGD5-AS1 and
RORA) or U6 (for miR-195). PCR reactions were repeated four times.
For subcellular distribution of FGD5-AS1, Nuclear/Cytosol Fraction
Kit (BioVision, Inc.) was utilized to obtain RNA in cytoplasm and
nuclear fractions, followed by normal RT-qPCR analysis as
aforementioned.
Western blotting
AC16 cells were lysed in radioimmunoprecipitation
assay buffer (Thermo Fisher Scientific, Inc.), and the supernatant
containing total protein samples was collected following
centrifugation at 12,000 × g at 4°C for 15 min. Following
separation of 20 µg protein samples (determined by Bradford method)
via SDS-PAGE on 12–15% gels, the proteins were electrophoretically
transferred onto PVDF membranes (EMD Millipore). Following blocking
with 3% bovine serum albumin at room temperature for 1 h, the
proteins were incubated with the following primary antibodies at
4°C overnight: RORA (1:500; cat. no. ab60134), superoxide dismutase
2 (SOD2; 1:10,000; cat. no. ab118340), glutathione peroxidase1
(GPx1; 1:500; cat. no. ab50427), Bax (1:1,000; cat. no. ab53154),
cleaved caspase-3 (cleaved casp-3; 1:500; cat. no. ab49822) and
GAPDH (1:2,500; cat. no. ab9485). The HRP-conjugated secondary
antibodies [goat anti-rabbit (1:50,000; cat. no. ab205718) and
rabbit anti-goat (1:20,000; cat. no. ab6741)] were used to make
protein bands immunoreactive at room temperature for 1 h for
further visualization using via chemiluminescence (EMD Millipore).
The gray intensity was quantified using ImageJ version 1.8.0
software (National Institutes of Health). The relative protein
expression levels were normalized to those of GAPDH. All antibodies
were obtained from Abcam.
Flow cytometry (FCM)
Hypoxia-treated AC16 cells were analyzed using
Annexin V-FITC early apoptosis detection kit (Cell Signaling
Technology, Inc.) according to the manufacturer's instructions. The
stained cells were analyzed on a BD Accuri™ C6 Flow Cytometer (BD
Biosciences), and apoptotic cells (%) were the percentage of cells
in Annexin V+/PI+ and Annexin
V+/PI− quadrants analyzed on CellQuest™ Pro
software 5.1 (BD Biosciences).
Reactive oxygen species (ROS),
malonaldehyde (MDA) and SOD assays
Generation of ROS was determined via the
2′,7′-dichlorofluorescin diacetate (DCFH-DA) method. In brief,
hypoxia-treated AC16 cells were exposed to 10 µM DCFH-DA (ROS
probe) for 1 h at 37°C in the dark. Then, the stained cells were
analyzed using a Tecan Infinite M200 fluorescence plate reader
(Tecan Group Ltd.), and fluorescence intensity was measured at 525
nm with an excitation wavelength of 488 nm. MDA and SOD levels were
measured using MDA Assay (Colorimetric/Fluorometric) (cat. no.
ab118970; Abcam) and human SOD2 ELISA kits (cat. no. ab178012;
Abcam) according to the manufacturer's instructions. The absorbance
was read at 532 (for MDA) or 450 nm (for SOD).
Dual-luciferase reporter assay
StarBase software version 2.0 (http://starbase.sysu.edu.cn/index.php)
was used to predict the potential binding site of miRNAs on both
FGD5-AS1 (starbase.sysu.edu.cn/lncRNA) and RORA (starbase.sysu.edu.cn/mRNA). A total of two paired
recombinant reporter vectors psiCHECK-2 (Promega Corporation) were
constructed, carrying wild-type (WT) or mutant (MUT) FGD5-AS1
fragment and the 3′-untranslated region (3′UTR) of RORA. AC16 cells
in 24-well plates were co-transfected with vectors and miR-195 or
miR-NC mimic using Lipofectamine 3000 reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. Following transfection for 48 h, the luciferase
activity of psiCHECK-2 vector was measured via Dual-Glo®
dual-luciferase kit (cat. no. E2920; Promega Corporation). The
ratio of firefly luciferase activity compared to that of
Renilla was calculated.
RNA pull-down assay
miRNA mimics were labelled with bio-16-UTP using T7
High Yield Transcription kit (Thermo Fisher Scientific, Inc.). The
bio-16-UTP-labelled miR-195 (bio-miR-195) or miR-NC mimics
(bio-miR-NC) were conjugated with streptavidin magnetic beads at
4°C for 2 h. After centrifugation at 2,000 × g for 1 min at 4°C,
the RNA-beads were collected and lysed in RNAiso Plus (Takara Bio,
Inc.) supplemented with 50 µg/ml Proteinase K (Thermo Fisher
Scientific, Inc.) at 4°C for 2 h. The supernatant was collected for
total RNA samples after centrifugation for 1 min at 2,000 × g at
4°C, and gene abundance was measured using RT-qPCR as
aforementioned.
Statistical analysis
Data are presented as the mean ± standard deviation
from at least three independent biological repeats. The statistical
analysis was performed using GraphPad Prism 6.0 software (GraphPad
Software, Inc.), and the P-value was calculated using unpaired
Student's t-test (two-tailed) or one-way ANOVA. Tukey's post hoc
test was performed following ANOVA. Pearson rank correlation
analysis was used to evaluate the correlation between gene
expression levels. P<0.05 was considered to indicate a
statistically significant difference.
Results
FGD5-AS1 and RORA are downregulated in
patients with AMI and hypoxia-induced human cardiomyocytes
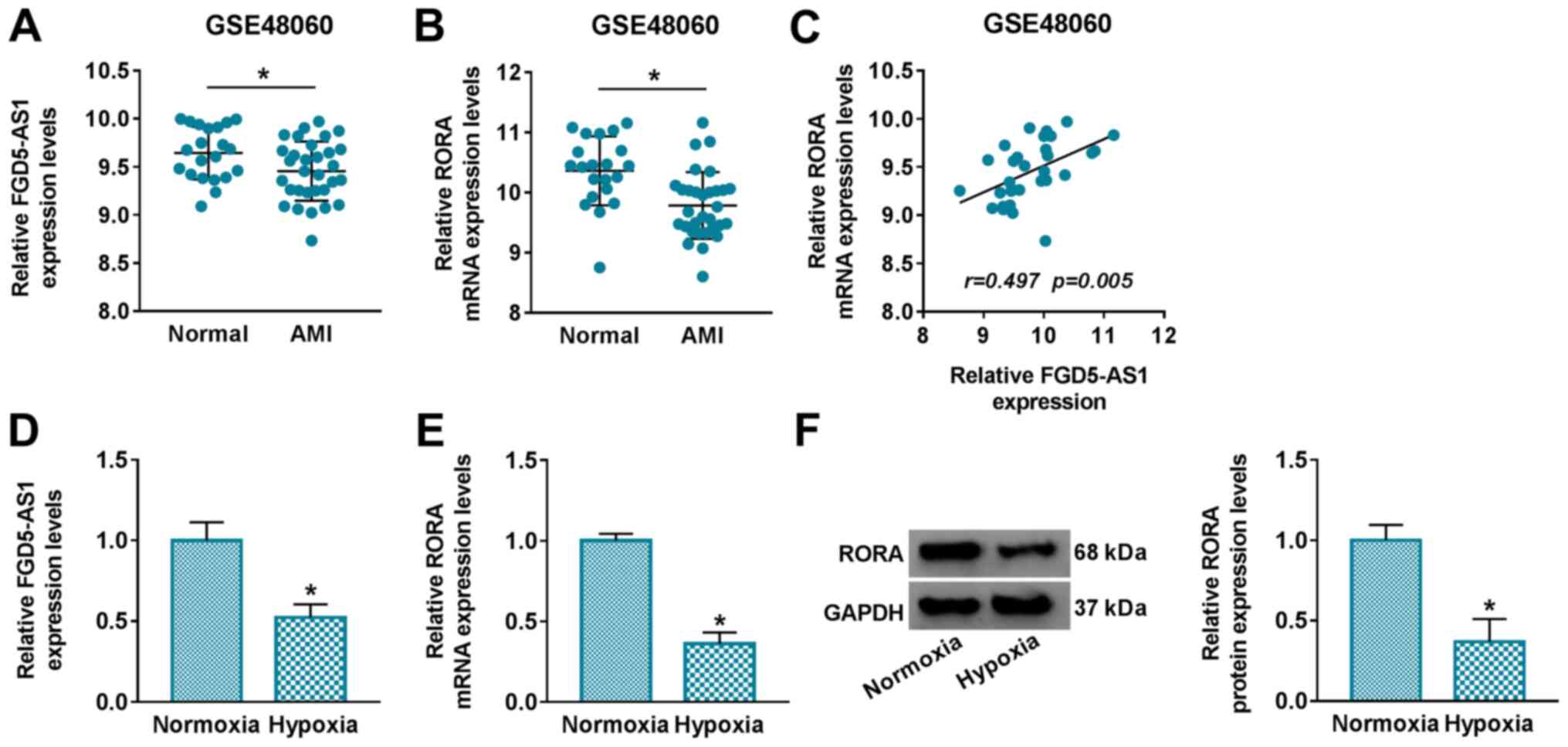
Analysis of the GSE48060 dataset demonstrated that
FGD5-AS1 and RORA were significantly downregulated in the serum of
patients with AMI compared with healthy normal controls (Fig. 1A and B). Moreover, Pearson rank
correlation analysis confirmed a positive linear correlation
between FGD5-AS1 and RORA expression levels in patients with AMI
(r=0.497; P=0.005) (Fig. 1C).
These data indicated a potential role of FGD5-AS1 and RORA in
myocardial damage. In order to investigate their role in myocardial
damage, the expression levels of FGD5-AS1 and RORA in a cell model
of AMI were further evaluated. Relative expression levels of
FGD5-AS1 and RORA were revealed to be significantly decreased in
hypoxia-challenged AC16 cells (Fig. 1D
and F).
Ectopic expression levels of FGD5-AS1
or RORA suppress hypoxia-induced oxidative stress and apoptosis in
cardiomyocytes
Next, the functional role of FGD5-AS1 and RORA
overexpression in an AMI cell model was investigated. The
overexpression of FGD5-AS1 and RORA was induced in AC16 cells via
transfection (Fig. 2A and B). In
response to hypoxic stress, ROS and MDA levels were highly induced
(Fig. 2C and E), but SOD activity
and the expression levels of SOD2 and GPx1 were significantly
inhibited in AC16 cells (Fig. 2D, G
and H), along with promotion of apoptosis and the expression
levels of Bax and cleaved casp-3 (Fig.
2F, G and I). These results indicated that hypoxia induced
oxidative stress and apoptosis of AC16 cells. However, transfection
of oe-FGD5-AS1 and oe-RORA separately attenuated hypoxia-induced
oxidative stress and apoptosis (Fig.
2C-I). These results indicated a suppressive role of FGD5-AS1
and RORA in hypoxic injury in AC16 cells.
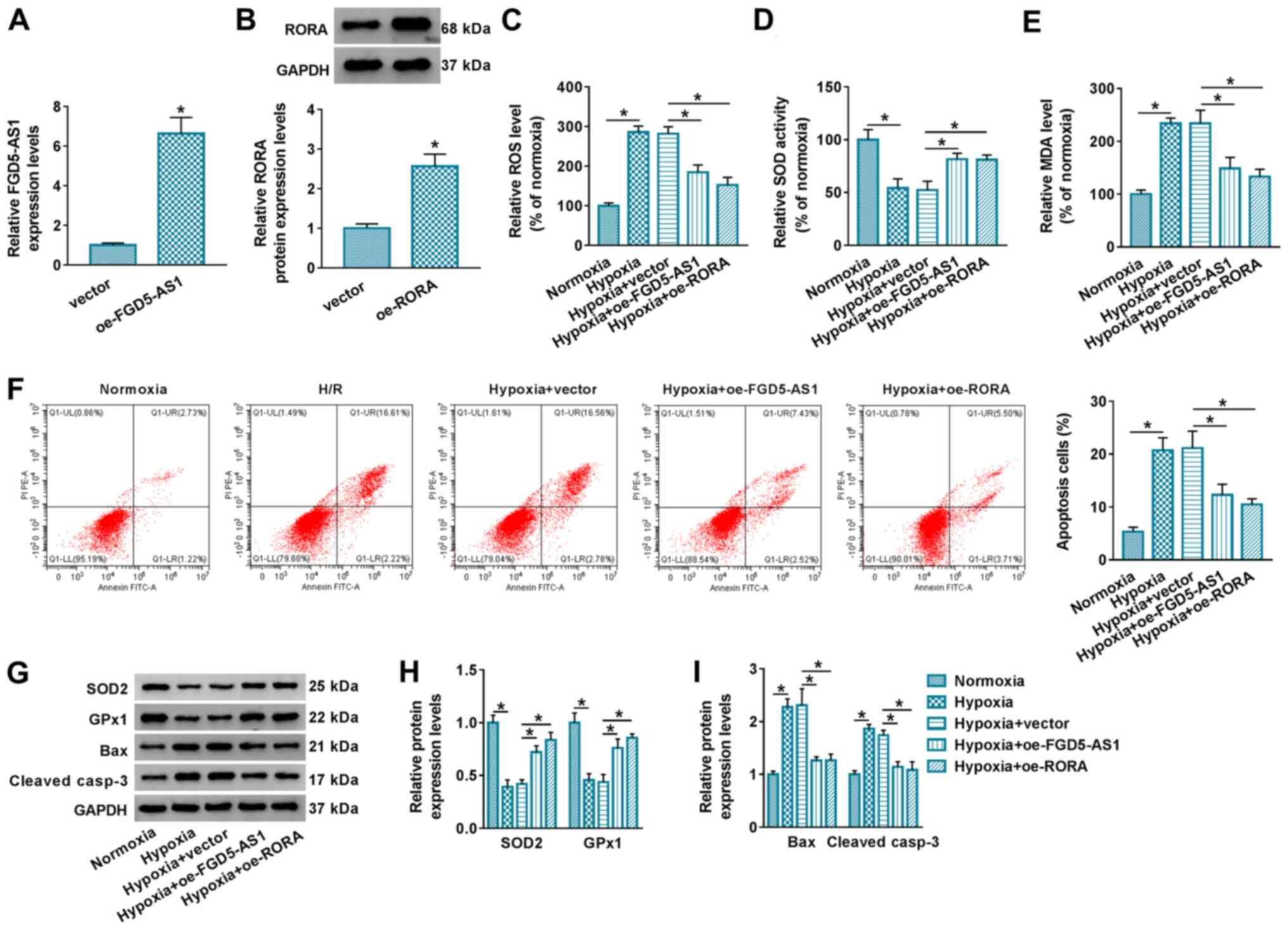 | Figure 2.Effect of FGD5-AS1 and RORA on
hypoxic injury in cardiomyocytes. (A) Reverse
transcription-quantitative PCR determined FGD5-AS1 levels in AC16
cells transfected with oe-FGD5-AS1 or empty vector. (B) Western
blotting determined RORA protein level in AC16 cells transfected
with oe-RORA or empty vector. AC16 cells were transfected with
oe-FGD5-AS1, oe-RORA or vector, and then treated with hypoxia for
12 h. Commercial kits examined the relative levels of (C) ROS, (D)
SOD and (E) MDA. (F) Flow cytometry evaluated apoptotic cells in
Annexin V-FITC+/PI+ and Annexin
V-FITC+/PI− quadrants. (G) Western blotting
assessed the protein expression levels of SOD2, GPx1, Bax and
cleaved casp-3. ImageJ software determined the relative protein
expression levels of (H) SOD2, GPx1 and (I) Bax and cleaved casp-3
by analyzing gray density. *P<0.05 vs. control group or as
indicated. FGD5-AS1, FGD5 antisense 1; RORA, retinoid acid
receptor-related orphan receptor α; oe, overexpression; ROS,
reactive oxygen species; SOD, superoxide dismutase; MDA,
malondialdehyde; GPx1, glutathione peroxidase 1; cleaved casp-3,
cleaved caspase-3. |
Silencing RORA abrogates the
suppressive role of FGD5-AS1 overexpression in hypoxic injury in
cardiomyocytes
It was next determined whether there was an
interaction between FGD5-AS1 and RORA. FGD5-AS1 ectopic expression
levels resulted in RORA upregulation in AC16 cells (Fig. 3A). Therefore, the effect of RORA
knockdown on FGD5-AS1 overexpression was evaluated. RORA knockdown
was performed via siRNA transfection (Fig. 3B). Assays demonstrated that the
lower levels of ROS and MDA in hypoxic AC16 cells transfected with
oe-FGD5-AS1 were restored, and higher SOD activity was diminished
following RORA silencing (Fig.
3C-E), accompanied by decreased SOD2 and GPx1 expression levels
(Fig. 3G and H). FCM demonstrated
that FGD5-AS1-mediated apoptosis inhibition in hypoxic AC16 cells
was relieved by blocking RORA (Fig.
3F), along with increased Bax and cleaved casp-3 expression
levels (Fig. 3G and I). These
results indicated that RORA knockdown partially abolished the
suppressive effect of FGD5-AS1 on oxidative stress and apoptosis in
hypoxic AC16 cells.
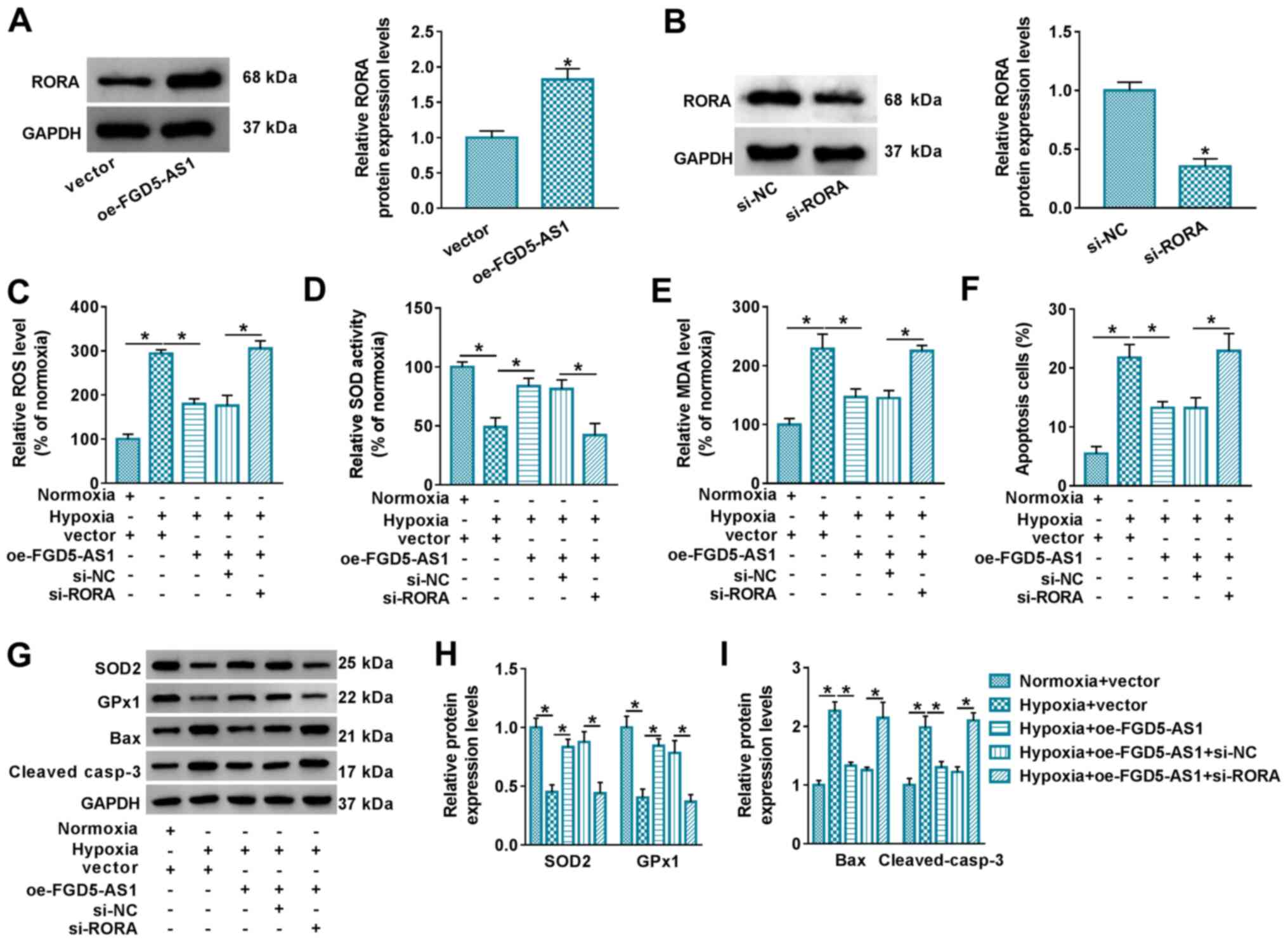 | Figure 3.Interfering RORA abrogates the role
of FGD5-AS1 overexpression in hypoxia-induced cardiomyocyte injury.
Western blotting measured RORA protein expression levels in AC16
cells transfected with (A) oe-FGD5-AS1 or vector and (B) si-RORA or
si-NC. Following hypoxia treatment for 12 h, assays were performed
to assess the relative levels of (C) ROS, (D) SOD and (E) MDA. (F)
Flow cytometry was performed to evaluate apoptotic cells. (G)
Western blotting was performed, and then semi-quantified to assess
the relative protein expression levels of (H) SOD2, GPx1, (I) Bax,
and cleaved casp-3. *P<0.05 vs. control group or as indicated.
RORA, retinoid acid receptor-related orphan receptor α; FGD5-AS1,
FGD5 antisense 1; oe, overexpression; siRNA, small interfering RNA;
NC, negative control; ROS, reactive oxygen species; SOD, superoxide
dismutase; MDA, malondialdehyde; GPx1, glutathione peroxidase 1;
cleaved casp-3, cleaved caspase-3. |
miR-195 is sponged by FGD5-AS1, and
RORA is a target of miR-195
Analysis of the relative expression levels of
FGD5-AS1 in subcellular fractions demonstrated that FGD5-AS1 was
predominantly present in the cytoplasm rather than the nucleus,
which was similar to the distribution of 18S rRNA (Fig. 4A). Thus, a potential ceRNA pathway
between FGD5-AS1, RORA and miRNAs was hypothesized. StarBase
software was utilized to predict the potential binding site of
miRNAs on both FGD5-AS1 and RORA. miR-195, a cardio-miRNA and
stress-responsive miRNA that plays a role in regulating cardiac
remodeling (17,18), was suggested to be complementary to
both FGD5-AS1 and RORA, thus this miRNA was selected for further
investigation (Fig. 4B). The
miR-195 or miR-NC mimic was exogenously administrated in AC16 cells
(Fig. 4C), and dual-luciferase
reporter assays were performed. Relative luciferase activity of
FGD5-AS1 WT was significantly attenuated by miR-195 mimic
transfection compared with miR-NC mimic transfection, whereas
FGD5-AS1 MUT luciferase activity exhibited little difference in the
presence of miR-195 or miR-NC mimic (Fig. 4D and E). FGD5-AS1 and RORA
expression levels were enriched in the bio-miR-195-mediated RNA
complex in AC16 cells (Fig. 4F).
oe-FGD5-AS1 transfection resulted in FGD5-AS1 overexpression and
miR-195 downregulation (Figs. 2A
and 4H); si-FGD5-AS1 transfection
decreased FGD5-AS1 and upregulated miR-195 (Fig. 4G and H). In addition, miR-195
overexpression via mimic transfection downregulated RORA protein
levels in AC16 cells, and this effect was further modulated by
FGD5-AS1 (Fig. 4I). These results
demonstrated that FGD5-AS1 sponged miR-195 to modulate RORA in AC16
cells. miR-195 was highly induced in response to hypoxic stress
(Fig. 4J).
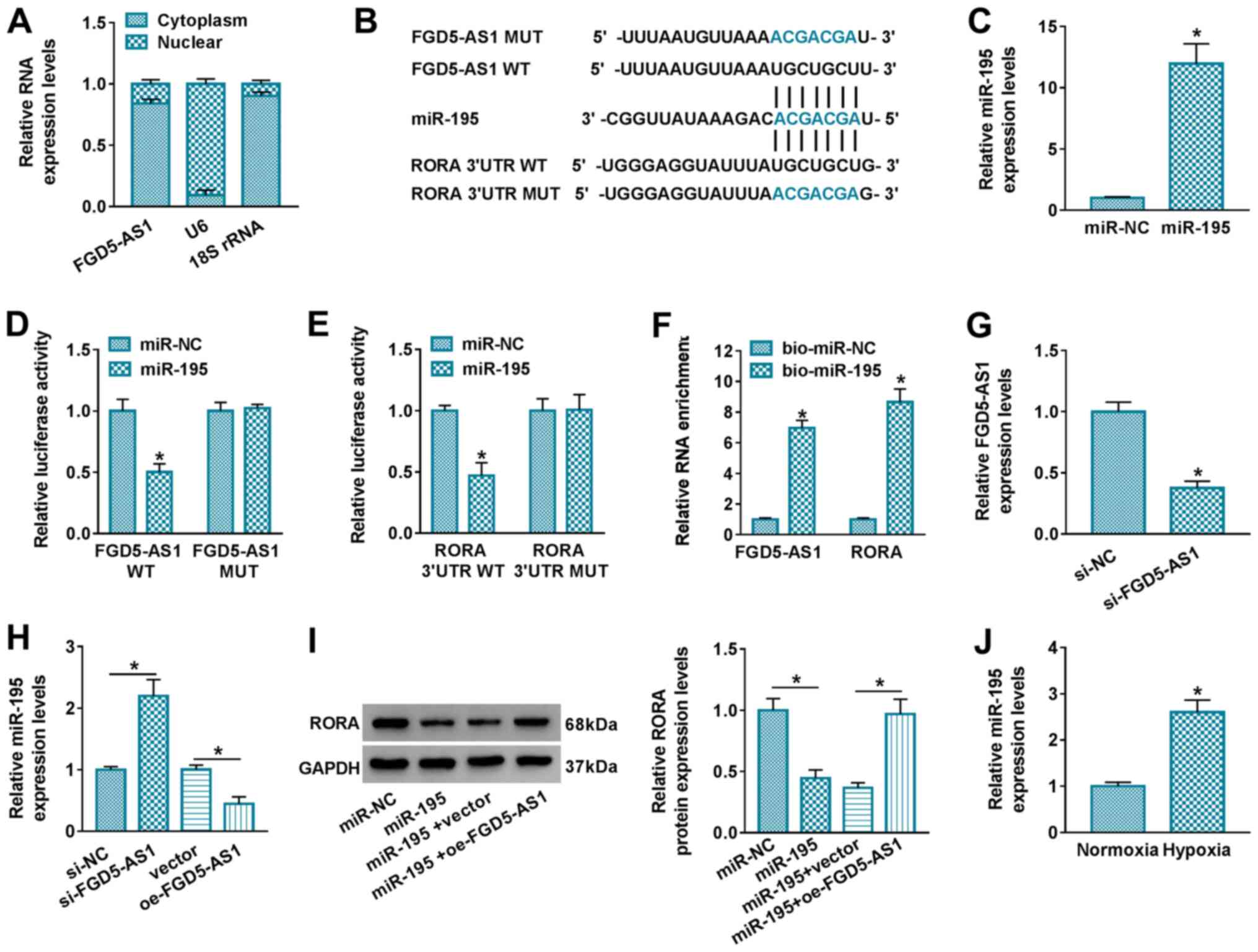 | Figure 4.Interaction between FGD5-AS1, RORA
and miR-195 in cardiomyocytes. (A) RT-qPCR assessed the relative
RNA expression levels in the cytoplasm and nuclear fraction of AC16
cells. (B) StarBase software predicted the putative binding sites
of miR-195 on WT and MUT FGD5-AS1 and RORA 3′UTR. (C) RT-qPCR
determined the relative miR-195 expression levels in AC16 cells
transfected with miR-195 or miR-NC mimic. Dual-luciferase reporter
assay measured the relative luciferase activity of AC16 cells
co-transfected with miR-195/NC and vectors carrying (D) FGD5-AS1
WT/MUT or (E) RORA 3′UTR WT/MUT. (F) RNA pull-down and qPCR assays
determined the relative enrichment of FGD5-AS1 and RORA in AC16
cells transfected with bio-miR-195 or bio-miR-NC. RT-qPCR
determined (G) the relative FGD5-AS1 expression levels in AC16
cells transfected with si-FGD5-AS1 or si-NC and (H) the relative
miR-195 expression levels in AC16 cells transfected with
si-FGD5-AS1, si-NC, oe-FGD5-AS1 or vector. (I) Western blotting
assessed the relative RORA protein expression levels in AC16 cells
transfected with miR-195 or miR-NC, and co-transfected with miR-195
and oe-FGD5-AS1 or vector. (J) RT-qPCR detected the relative
miR-195 expression levels in hypoxic and normoxic cells. *P<0.05
vs. control group or as indicated. FGD5-AS1, FGD5 antisense 1;
RORA, retinoid acid receptor-related orphan receptor α; miR,
microRNA; RT-qPCR, reverse transcription-quantitative PCR; WT,
wild-type; MUT, mutant; UTR, untranslated region; NC, negative
control; bio, biotin-labeled; siRNA, small interfering RNA; oe,
overexpression; 18S rRNA, 18S ribosome RNA. |
Upregulating FGD5-AS1 or RORA
abolishes the diminishing effect of miR-195 knockdown on
hypoxia-induced oxidative stress and apoptosis in
cardiomyocytes
The role of miR-195 knockdown was further assessed
in hypoxic AC16 cells via anti-RNA transfection. Silencing
efficiency of anti-miR-195 was determined via RT-qPCR (Fig. 5A). Commercial assays demonstrated
that the promotion of ROS and MDA levels was reversed following
anti-miR-195 transfection in hypoxic AC16 cells, as well as
decreased SOD activity (Fig.
5B-D), and increased SOD2 and GPx1 expression levels (Fig. 5F and G). FCM demonstrated that
hypoxia-mediated apoptosis of AC16 cells was suppressed by miR-195
knockdown, as evidenced by decreased apoptosis rate (Fig. 5E) and the expression levels of Bax
and cleaved casp-3 (Fig. 5F and
H). These results revealed that miR-195 knockdown alleviated
hypoxic injury in AC16 cells. Notably, the presence of either
si-FGD5-AS1 or si-RORA counteracted the biological role of miR-195
knockdown in hypoxic AC16 cells (Fig.
5B-H). These results indicated a novel FGD5-AS1/miR-195/RORA
ceRNA pathway underlying hypoxic injury in cardiomyocytes in
vitro.
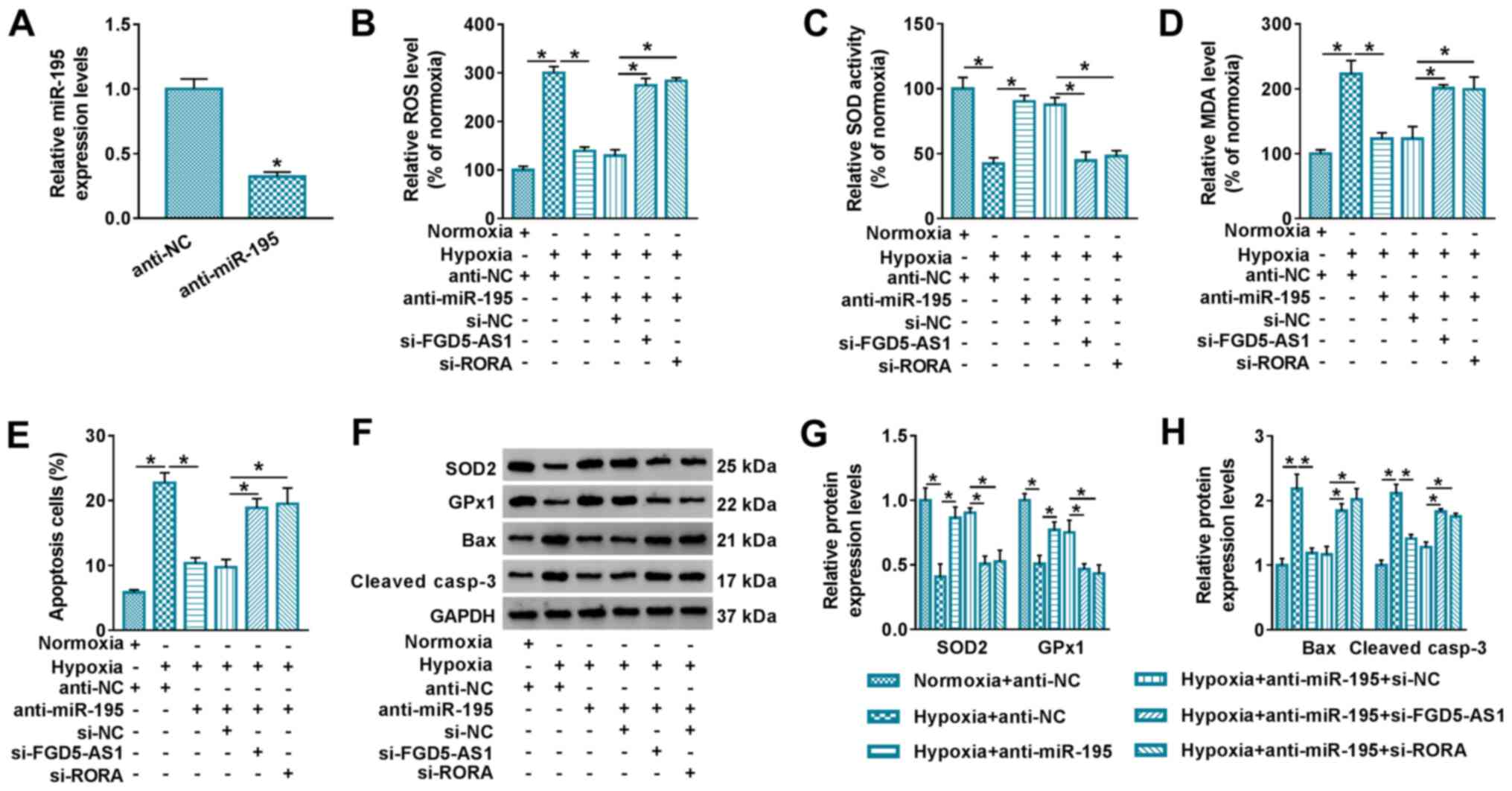 | Figure 5.Interfering FGD5-AS1 or RORA
abolishes the role of miR-195 knockdown in hypoxia-induced
cardiomyocyte injury. (A) Reverse transcription-quantitative PCR
detected the relative miR-195 expression levels in AC16 cells
transfected with anti-miR-195 or anti-NC. AC16 cells were
transfected with anti-miR-195 or anti-NC, or co-transfected with
anti-miR-195 and si-FGD5-AS1, si-RORA or si-NC, followed by hypoxia
treatment for 12 h. Assays were performed to assess the relative
levels of (B) ROS, (C) SOD and (D) MDA. (E) Flow cytometry
evaluated apoptotic cells. (F) Western blotting was performed, and
then semi-quantified to assess the relative protein expression
levels of (G) SOD2, GPx1, (H) Bax and cleaved casp-3. *P<0.05
vs. control group or as indicated. FGD5-AS1, FGD5 antisense 1;
RORA, retinoid acid receptor-related orphan receptor α; miR,
microRNA; NC, negative control; siRNA, small interfering RNA; ROS,
reactive oxygen species; SOD, superoxide dismutase; MDA,
malondialdehyde; GPx1, glutathione peroxidase 1; cleaved casp-3,
cleaved caspase-3. |
Discussion
Oxidative stress occurs in cardiomyocyte injury,
such as acute ischemia-hypoxia and
reperfusion/reoxygenation-mediated injury (22). Previous research has demonstrated
that lncRNAs are abnormally expressed in ischemia-hypoxia injury
and can affect proliferation, autophagy, migration, invasion and
apoptosis of cardiomyocytes (23,24),
as well as the inflammatory response (25). In terms of oxidative stress, Su
et al (26) reported that
lncRNA taurine upregulated 1 knockdown improved viability and
inhibited apoptosis and ROS production in
H2O2-induced hypoxic cardiomyocytes via
targeting the miRNA-132-3p/histone deacetylase 3 axis. Jiao et
al (27) demonstrated that
lncRNA ZNFX antisense RNA 1 induced mitochondrial apoptosis via
cytosolic Ca2+ overload in AMI mice. However, little is
currently known about the association between lncRNAs and oxidative
damage in ischemic/hypoxic cardiomyocytes; further investigation is
therefore required. The present study confirmed a protective effect
of FGD5-AS1 against oxidative stress and apoptosis in a
hypoxia-induced model of AMI in AC16 cells via targeting the
miR-195/RORA axis.
Downregulation of FGD5-AS1 was observed in the serum
of patients with AMI; this finding was consistent with a previous
study (11) in which FGD5-AS1,
along with other lncRNAs, was identified as key regulator in AMI by
regulating >50 differentially expressed genes, and an
lncRNA-mRNA co-expression network in AMI was identified. The
present study confirmed that FGD5-AS1 was downregulated in
hypoxia-challenged AC16 cells, and that overexpression of FGD5-AS1
attenuated ROS and MDA levels, the apoptosis rate and expression
levels of Bax and cleaved casp-3, but increased SOD activity and
the expression levels of SOD2 and GPx1 in hypoxic AC16 cells. These
outcomes indicated a suppressive effect of FGD5-AS1 in
hypoxia-induced oxidative injury of cardiomyocytes. Chen et
al (28) reported a protective
role of FGD5-AS1 in lipopolysaccharide-induced inflammatory injury
in human periodontal ligament cells, as evidenced by counteraction
of cell apoptosis and pro-inflammatory cytokine secretion via the
NF-κB pathway. The role of FGD5-AS1 in inflammation in AMI requires
further investigation, although ischemia-hypoxia is known to highly
induce an inflammatory response in AMI (29).
FGD5-AS1 has been revealed to sponge a number of
miRNAs, such as miRNA-142-3p, miRNA-125a-3p, miRNA-302e (13,30)
and miR-195, to modulate their cellular concentrations. miR-195 was
revealed to be involved in the cell cycle, apoptosis and
proliferation in multiple diseases, including heart failure
(31). In heart cells, miR-195 may
be a promising therapeutic strategy for myocardial injury, such as
hypoxia- and H2O2-evoked myocardial ischemia
(32,33), I/R injury (34), angiotensin II- and
isoprenaline-induced cardiomyocyte hypertrophy (35,36),
streptozotocin-mimicked diabetic cardiomyopathy (37) and palmitate-activated lipotoxic
cardiomyopathy (38). The present
study detected high expression levels of miR-195 in hypoxic AC16
cells, which was consistent with the results of a previous study
(32). Silencing of miR-195 may
attenuate hypoxia-induced oxidative stress and apoptosis of AC16
cells. Zhang et al (33)
reported that miR-195 overexpression suppressed cell viability, and
promoted cell apoptosis in HCMs under H2O2
stimulation; Hang et al (32) demonstrated that miR-195 deletion
improved cell viability and decreased apoptosis of primary
cardiomyocytes following hypoxia or H2O2
treatment. Furthermore, it was discovered in the present study that
miR-195 expression levels were highly induced by hypoxic stress,
which is consistent with the effects of H2O2
stimulation or high glucose stress on miR-195 regulation (39). By contrast, miR-195 downregulation
was coupled with a diminishing effect on MDA and promoting effect
on SOD; this was consistent with results reported by Liu et
al (40).
Brain-derived neurotrophic factor and Bcl-2 like
protein 2 are downstream target genes of miR-195 (32,33),
and the present study demonstrated that RORA mediated
cardioprotection of miR-195 knockdown against hypoxia-mediated
oxidative injury in AC16 cells by serving as a downstream target of
miR-195. The present study confirmed RORA downregulation in the
serum of patients with AMI and hypoxia-treated AC16 cells,
indicating a potential role of RORA in myocardial anoxia. Moreover,
RORA, along with seven other genes, has been identified as a key
gene in myocardial infarction, according to RNA sequencing data
(41). He et al (20) identified the protective role of
RORA in myocardial I/R injury, and proposed that RORA was
downregulated in a mouse heart following I/R. Deficiency of RORA
exacerbates high fat diet-induced myocardial hypertrophy, fibrosis
and dysfunction (42). Beak et
al (19) also reported
angiotensin II-stimulated cardiac hypertrophy and cardiomyocyte
death were exaggerated following RORA loss, accompanied by
increased inflammatory response and decreased oxidative stress.
Additionally, overexpression of RORA is considered to prevent
oxidative damage in different types of cells, such as neurons
(43) and hepatocytes (44), as well as cardiomyocytes (19,20).
The present study demonstrated that RORA ectopic expression levels
attenuated ROS and MDA levels, the apoptosis rate and the
expression levels of Bax and cleaved casp-3 but increased SOD
activity and the expression levels of SOD2 and GPx1 in hypoxic AC16
cells.
Cathepsins (Cat) are matrix degradation proteases
that are considered to modulate vascular aging and remodeling. For
example, Xin et al (45)
revealed that matrix metalloproteinase (MMP)-2, MMP-9, Cat S and
Cat K were upregulated in response to chronic stress-induced aortic
senescence, accompanied by oxidative and inflammatory responses in
human umbilical vein endothelial cells. The peroxisome
proliferator-activated receptor-γ is upstream of Cat S signaling in
pulmonary vascular remodeling (46). The Cat K/caspase-8 axis is a key
initial step for oxidative stress (47), and Cat K ablation is known to
mitigate high-fat-diet-induced cardiomyocyte apoptosis and cardiac
hypertrophy (48). However, the
role of the FGD5-AS1/miR-195/RORA axis in Cat S/K expression levels
was not determined in the present study, although previous studies
have demonstrated that Cat S and Cat K are downregulated in
intermittent hypoxia-treated and ischemia-induced mice,
respectively (49,50). Moreover, Cat K has been revealed to
promote proliferation, migration and tube formation of human
vascular endothelial cells under hypoxia via acting as target of
miR-185-5p and the Notch 1 signaling pathway (50,51).
Further investigation is required to determine the association
between hypoxia-induced oxidative injury, the FGD5-AS1/miR-195/RORA
axis, cardiomyocytes and Cat S/K expression levels.
FGD5-AS1 (co-expressing with seven other lncRNAs) is
secreted in the blood of patients with AMI (11); to the best of our knowledge,
however, there are no previous studies describing the source and
distribution of FGD5-AS1 in vascular wall cells. miR-195 is be
secreted by endothelial cells of the pulmonary and carotid arteries
into smooth muscle cells (52,53),
indicating endothelial cells are the source of miR-195 in vascular
walls. RORA is present in human vascular endothelial cells of the
umbilical vein and lung microvessels (54,55);
moreover, higher expression levels of RORA isoform 1 have been
detected in human smooth muscle cells of the aorta, compared with
those of endothelial cells, mammary arteries and atherosclerotic
plaques (56). However, smooth
muscle cells cannot be considered as the source of RORA as high
expression of RORA isoform 4 is predominantly found in endothelial
cells (55). Little is known about
the expression levels of FGD5-AS1, miR-195 and RORA in vascular
epithelial cells. RORA and miR-195 are expressed in normal
cardiomyocytes (57,58), and higher levels of FGD5-AS1 and
RORA (isoform 1), as well as lower miR-195 levels were observed in
normoxic cardiomyocytes than in hypoxic cells. To the best of our
knowledge, the present study is the first to determine FGD5-AS1
expression levels in cardiomyocytes.
In conclusion, the present study demonstrated that
FGD5-AS1, which is positively correlated to RORA, suppressed
oxidative stress and apoptosis in AC16 cells under hypoxia via
acting as a miR-195 sponge. The results indicated a novel
FGD5-AS1/miR-195/RORA axis in hypoxic injury of cardiomyocytes
in vitro, which may be a potential therapeutic target
against AMI by lowering oxidative damage.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Jiangxi
Provincal Natural Science Foundation (grant no.
20171BAB215005).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XC, SW and LH conceptualized and designed the
experiments, performed the data collection and analysis, and
prepared the original draft of the manuscript. PZ, SY, BL and HZ
performed data validation and checked the manuscript. XY and LS
collected and analyzed the data, and revised the paper critically
for important intellectual content prior to reviewing and editing.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics Review
Committee of Jiangxi Provincial People's Hospital Affiliated to
Nanchang University (Nanchang, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
GBD 2013 Mortality and Causes of Death
Collaborators: Global, regional, and national age-sex specific
all-cause and cause-specific mortality for 240 causes of death,
1990–2013: A systematic analysis for the global burden of disease
study 2013. Lancet. 385:117–171. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lim GB: Acute coronary syndromes:
Supplemental oxygen in myocardial infarction. Nat Rev Cardiol.
14:6322017. View Article : Google Scholar
|
|
3
|
Ham PB III and Raju R: Mitochondrial
function in hypoxic ischemic injury and influence of aging. Prog
Neurobiol. 157:92–116. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yu B, Meng F, Yang Y, Liu D and Shi K:
NOX2 antisense attenuates hypoxia-induced oxidative stress and
apoptosis in cardiomyocyte. Int J Med Sci. 13:646–652. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hou L, Guo J, Xu F, Weng X, Yue W and Ge
J: Cardiomyocyte dimethylarginine dimethylaminohydrolase1
attenuates left-ventricular remodeling after acute myocardial
infarction: Involvement in oxidative stress and apoptosis. Basic
Res Cardiol. 113:282018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang Y, Li C, Meng H, Guo D, Zhang Q, Lu
W, Wang Q, Wang Y and Tu P: BYD ameliorates oxidative
stress-induced myocardial apoptosis in heart failure post-acute
myocardial infarction via the P38 MAPK-CRYAB signaling pathway.
Front Physiol. 9:5052018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lucas T, Bonauer A and Dimmeler S: RNA
therapeutics in cardiovascular disease. Circ Res. 123:205–220.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kaikkonen MU, Halonen P, Liu OH, Turunen
TA, Pajula J, Moreau P, Selvarajan I, Tuomainen T, Aavik E, Tavi P
and Ylä-Herttuala S: Genome-wide dynamics of nascent noncoding RNA
transcription in porcine heart after myocardial infarction. Circ
Cardiovasc Genet. 10:e0017022017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li H, Cheng Z, Tang Y, Feng M, Yin A,
Zhang H, Xu J, Zhang Q, Zhang J and Qian L: Expression profile of
long noncoding RNAs in cardiomyocytes exposed to acute ischemic
hypoxia. Mol Med Rep. 19:302–308. 2019.PubMed/NCBI
|
|
10
|
Ong SB, Katwadi K, Kwek XY, Ismail NI,
Chinda K, Ong SG and Hausenloy DJ: Non-coding RNAs as therapeutic
targets for preventing myocardial ischemia-reperfusion injury.
Expert Opin Ther Targets. 22:247–261. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shen LS, Hu XF, Chen T, Shen GL and Cheng
D: Integrated network analysis to explore the key mRNAs and lncRNAs
in acute myocardial infarction. Math Biosci Eng. 16:6426–6437.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu H, Xu D, Zhong X, Xu D, Chen G, Ge J
and Li H: LncRNA-mRNA competing endogenous RNA network depicts
transcriptional regulation in ischaemia reperfusion injury. J Cell
Mol Med. 23:2272–2276. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li D, Jiang X, Zhang X, Cao G, Wang D and
Chen Z: Long noncoding RNA FGD5-AS1 promotes colorectal cancer cell
proliferation, migration, and invasion through upregulating CDCA7
via sponging miR-302e. In Vitro Cell Dev Biol Anim. 55:577–585.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lei Y, Shi Y, Duan J, Liu Y, Lv G, Shi R,
Zhang F, Yang Q and Zhao W: Identification of alternative splicing
and lncRNA genes in pathogenesis of small cell lung cancer based on
their RNA sequencing. Adv Clin Exp Med. 28:1043–1050. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhu H, Lu J, Zhao H, Chen Z, Cui Q, Lin Z,
Wang X, Wang J, Dong H, Wang S and Tan J: Functional long noncoding
RNAs (lncRNAs) in clear cell kidney carcinoma revealed by
reconstruction and comprehensive analysis of the lncRNA-miRNA-mRNA
regulatory network. Med Sci Monit. 24:8250–8263. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yu W, Liang X, Li X, Zhang Y, Sun Z, Liu Y
and Wang J: MicroRNA-195: A review of its role in cancers. Onco
Targets Ther. 11:7109–7123. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Katoh M: Cardio-miRNAs and onco-miRNAs:
Circulating miRNA-based diagnostics for non-cancerous and cancerous
diseases. Front Cell Dev Biol. 2:612014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
van Rooij E, Sutherland LB, Liu N,
Williams AH, McAnally J, Gerard RD, Richardson JA and Olson EN: A
signature pattern of stress-responsive microRNAs that can evoke
cardiac hypertrophy and heart failure. Proc Natl Acad Sci USA.
103:18255–18260. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Beak JY, Kang HS, Huang W, Myers PH,
Bowles DE, Jetten AM and Jensen BC: The nuclear receptor RORα
protects against angiotensin II-induced cardiac hypertrophy and
heart failure. Am J Physiol Heart Circ Physiol. 316:H186–H200.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
He B, Zhao Y, Xu L, Gao L, Su Y, Lin N and
Pu J: The nuclear melatonin receptor RORα is a novel endogenous
defender against myocardial ischemia/reperfusion injury. J Pineal
Res. 60:313–326. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bar-Or D, Bar-Or R, Rael LT and Brody EN:
Oxidative stress in severe acute illness. Redox Biol. 4:340–345.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wei Q, Zhou HY, Shi XD, Cao HY and Qin L:
Long noncoding RNA NEAT1 promotes myocardiocyte apoptosis and
suppresses proliferation through regulation of miR-129-5p. J
Cardiovasc Pharmacol. 74:535–541. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hu H, Wu J, Yu X, Zhou J, Yu H and Ma L:
Long non-coding RNA MALAT1 enhances the apoptosis of cardiomyocytes
through autophagy inhibition by regulating TSC2-mTOR signaling.
Biol Res. 52:582019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lu W, Zhu L, Ruan ZB, Wang MX, Ren Y and
Li W: HOTAIR promotes inflammatory response after acute myocardium
infarction by upregulating RAGE. Eur Rev Med Pharmacol Sci.
22:7423–7430. 2018.PubMed/NCBI
|
|
26
|
Su Q, Liu Y, Lv XW, Dai RX, Yang XH and
Kong BH: LncRNA TUG1 mediates ischemic myocardial injury by
targeting miR-132-3p/HDAC3 axis. Am J Physiol Heart Circ Physiol.
318:H332–H344. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jiao L, Li M, Shao Y, Zhang Y, Gong M,
Yang X, Wang Y, Tan Z, Sun L, Xuan L, et al: lncRNA-ZFAS1 induces
mitochondria-mediated apoptosis by causing cytosolic
Ca2+ overload in myocardial infarction mice model. Cell
Death Dis. 10:9422019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen H, Lan Z, Li Q and Li Y: Abnormal
expression of long noncoding RNA FGD5-AS1 affects the development
of periodontitis through regulating miR-142-3p/SOCS6/NF-κB pathway.
Artif Cells Nanomed Biotechnol. 47:2098–2106. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cheng WP, Lo HM, Wang BW, Chua SK, Lu MJ
and Shyu KG: Atorvastatin alleviates cardiomyocyte apoptosis by
suppressing TRB3 induced by acute myocardial infarction and
hypoxia. J Formos Med Assoc. 116:388–397. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li S, Liu X, Li H, Pan H, Acharya A, Deng
Y, Yu Y, Haak R, Schmidt J, Schmalz G and Ziebolz D: Integrated
analysis of long noncoding RNA-associated competing endogenous RNA
network in periodontitis. J Periodontal Res. 53:495–505. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
He JF, Luo YM, Wan XH and Jiang D:
Biogenesis of MiRNA-195 and its role in biogenesis, the cell cycle,
and apoptosis. J Biochem Mol Toxicol. 25:404–408. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hang P, Sun C, Guo J, Zhao J and Du Z:
BDNF-mediates Down-regulation of MicroRNA-195 inhibits ischemic
cardiac apoptosis in rats. Int J Biol Sci. 12:979–989. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang N, Meng X, Mei L, Hu J, Zhao C and
Chen W: The long non-coding RNA SNHG1 attenuates cell apoptosis by
regulating miR-195 and BCL2-like protein 2 in human cardiomyocytes.
Cell Physiol Biochem. 50:1029–1040. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gao CK, Liu H, Cui CJ, Liang ZG, Yao H and
Tian Y: Roles of MicroRNA-195 in cardiomyocyte apoptosis induced by
myocardial ischemia-reperfusion injury. J Genet. 95:99–108. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
You XY, Huang JH, Liu B, Liu SJ, Zhong Y
and Liu SM: HMGA1 is a new target of miR-195 involving
isoprenaline-induced cardiomyocyte hypertrophy. Biochemistry
(Mosc). 79:538–544. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang L, Qin D, Shi H, Zhang Y, Li H and
Han Q: MiR-195-5p promotes cardiomyocyte hypertrophy by targeting
MFN2 and FBXW7. Biomed Res Int. 2019:15809822019.PubMed/NCBI
|
|
37
|
Zheng D, Ma J, Yu Y, Li M, Ni R, Wang G,
Chen R, Li J, Fan GC, Lacefield JC and Peng T: Silencing of miR-195
reduces diabetic cardiomyopathy in C57BL/6 mice. Diabetologia.
58:1949–1958. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhu H, Yang Y, Wang Y, Li J, Schiller PW
and Peng T: MicroRNA-195 promotes palmitate-induced apoptosis in
cardiomyocytes by down-regulating Sirt1. Cardiovasc Res. 92:75–84.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang R, Garrett Q, Zhou H, Wu X, Mao Y,
Cui X, Xie B, Liu Z, Cui D, Jiang L, et al: Upregulation of miR-195
accelerates oxidative stress-induced retinal endothelial cell
injury by targeting mitofusin 2 in diabetic rats. Mol Cell
Endocrinol. 452:33–43. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu P, Peng QH, Tong P and Li WJ:
Astragalus polysaccharides suppresses high glucose-induced
metabolic memory in retinal pigment epithelial cells through
inhibiting mitochondrial dysfunction-induced apoptosis by
regulating miR-195. Mol Med. 25:212019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhao Q, Wu K, Li N, Li Z and Jin F:
Identification of potentially relevant genes for myocardial
infarction using RNA sequencing data analysis. Exp Ther Med.
15:1456–1464. 2018.PubMed/NCBI
|
|
42
|
Zhao YC, Xu LW, Ding S, Ji QQ, Lin N, He
Q, Gao LC, Su YY, Pu J and He B: Nuclear receptor retinoid-related
orphan receptor alpha deficiency exacerbates high-fat diet-induced
cardiac dysfunction despite improving metabolic abnormality.
Biochim Biophys Acta Mol Basis Dis. 1863:1991–2000. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Boukhtouche F, Vodjdani G, Jarvis CI,
Bakouche J, Staels B, Mallet J, Mariani J, Lemaigre-Dubreuil Y and
Brugg B: Human retinoic acid receptor-related orphan receptor
alpha1 overexpression protects neurones against oxidative
stress-induced apoptosis. J Neurochem. 96:1778–1789. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Han YH, Kim HJ, Kim EJ, Kim KS, Hong S,
Park HG and Lee MO: RORα decreases oxidative stress through the
induction of SOD2 and GPx1 expression and thereby protects against
nonalcoholic steatohepatitis in mice. Antioxid Redox Signal.
21:2083–2094. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xin M, Jin X, Cui X, Jin C, Piao L, Wan Y,
Xu S, Zhang S, Yue X, Wang H, et al: Dipeptidyl peptidase-4
inhibition prevents vascular aging in mice under chronic stress:
Modulation of oxidative stress and inflammation. Chem Biol
Interact. 314:1088422019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chang CJ, Hsu HC, Ho WJ, Chang GJ, Pang
JS, Chen WJ, Huang CC and Lai YJ: Cathepsin S promotes the
development of pulmonary arterial hypertension. Am J Physiol Lung
Cell Mol Physiol. 317:L1–L13. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hu L, Huang Z, Ishii H, Wu H, Suzuki S,
Inoue A, Kim W, Jiang H, Li X, Zhu E, et al: PLF-1 (Proliferin-1)
modulates smooth muscle cell proliferation and development of
experimental intimal hyperplasia. J Am Heart Assoc. 8:e0058862019.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hua Y, Zhang Y, Dolence J, Shi GP, Ren J
and Nair S: Cathepsin K knockout mitigates high-fat diet-induced
cardiac hypertrophy and contractile dysfunction. Diabetes.
62:498–509. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yang YY, Li LY, Jiao XL, Jia LX, Zhang XP,
Wang YL, Yang S, Li J, Du J, Wei YX and Qin YW: Intermittent
hypoxia alleviates β-aminopropionitrile monofumarate induced
thoracic aortic dissection in C57BL/6 mice. Eur J Vasc Endovasc
Surg. 59:1000–1010. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Jiang H, Cheng XW, Shi GP, Hu L, Inoue A,
Yamamura Y, Wu H, Takeshita K, Li X, Huang Z, et al: Cathepsin
K-mediated Notch1 activation contributes to neovascularization in
response to hypoxia. Nat Commun. 5:38382014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Li CC, Qiu XT, Sun Q, Zhou JP, Yang HJ, Wu
WZ, He LF, Tang CE, Zhang GG and Bai YP: Endogenous reduction of
miR-185 accelerates cardiac function recovery in mice following
myocardial infarction via targeting of cathepsin K. J Cell Mol Med.
23:1164–1173. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zeng Z, Yao J, Li Y, Xue Y, Zou Y, Shu Z
and Jiao Z: Anti-apoptosis endothelial cell-secreted
microRNA-195-5p promotes pulmonary arterial smooth muscle cell
proliferation and migration in pulmonary arterial hypertension. J
Cell Biochem. 119:2144–2155. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Gu J, Zhang H, Ji B, Jiang H, Zhao T,
Jiang R, Zhang Z, Tan S, Ahmed A and Gu Y: Vesicle miR-195 derived
from endothelial cells inhibits expression of serotonin transporter
in vessel smooth muscle cells. Sci Rep. 7:435462017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Gulec C, Coban N, Ozsait-Selcuk B,
Sirma-Ekmekci S, Yildirim O and Erginel-Unaltuna N: Identification
of potential target genes of ROR-alpha in THP1 and HUVEC cell
lines. Exp Cell Res. 353:6–15. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kim SY, Kim HJ, Park MK, Huh JW, Park HY,
Ha SY, Shin JH and Lee YS: Mitochondrial E3 ubiquitin protein
ligase 1 mediates cigarette smoke-induced endothelial cell death
and dysfunction. Am J Respir Cell Mol Biol. 54:284–296. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Besnard S, Heymes C, Merval R, Rodriguez
M, Galizzi JP, Boutin JA, Mariani J and Tedgui A: Expression and
regulation of the nuclear receptor RORalpha in human vascular
cells. FEBS Lett. 511:36–40. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Xu L, Su Y, Zhao Y, Sheng X, Tong R, Ying
X, Gao L, Ji Q, Gao Y, Yan Y, et al: Melatonin differentially
regulates pathological and physiological cardiac hypertrophy:
Crucial role of circadian nuclear receptor RORα signaling. J Pineal
Res. 67:e125792019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Porrello ER, Johnson BA, Aurora AB,
Simpson E, Nam YJ, Matkovich SJ, Dorn GW II, van Rooij E and Olson
EN: MiR-15 family regulates postnatal mitotic arrest of
cardiomyocytes. Circ Res. 109:670–679. 2011. View Article : Google Scholar : PubMed/NCBI
|