Introduction
Neurodegenerative diseases, such as epilepsy,
Alzheimer's disease and Huntington's, are a major global burden to
human health. Currently, there are no effective treatments
available for the majority of neurodegenerative diseases (1,2), and
the existing pharmacotherapy is inconsistent with severe adverse
effects. Cell replacement therapy with neurons may be an
alternative therapy for these diseases (1). Therefore, in vitro generation
and expansion of neurons are critical for the cell therapy
approach, although neurons can be generated by direct conversion
for disease modeling (2,3). Current efforts are focused on using
exogenous cells derived from embryonic stem cells (ESCs) or induced
pluripotent stem cells to generate neurons (4,5).
However, cell transplantation approaches using stem cell-derived
neurons face significant hurdles, such as immunorejection,
tumorigenesis, unstable genetic manipulation and differentiation
uncertainty (6,7). Therefore, there is a need to find a
novel approach for neuronal generation. Small molecular compounds
have been used to directly reprogram human fibroblasts into neurons
due to their advantages, including being non-immunogenic, lower
costs and easy optimization (8,9).
Compared with transcription factor-based reprogramming, small
molecular compound-mediated trans-differentiation has the potential
to evolve into a rapid and direct approach for the generation of
customized cell types, due to the availability of somatic cells
(mainly skin-derived fibroblasts) and the flexibility to generate
autologous target cells (9).
Several studies have reported that a combination of small chemical
compounds efficiently and directly converts human fibroblasts into
functional neurons (10–12). However, these reprogramming
strategies still have some limitations, such as low efficiency and
time-consuming induction (11).
Therefore, it is essential to identify more efficient small
molecule-induced reprogramming strategies for cell therapies of
neurological disorders.
The present study reports a defined combination of
eight small molecular compounds capable of directly reprogramming
human foreskin fibroblasts (HFFs) into functional neurons following
sequential treatment in vitro. Additionally, the present
results revealed the course of morphological cell changes during
the inductive periods.
Materials and methods
Cell culture
HFFs were cultured from the foreskin of a
20-year-old man at The General Hospital of Ningxia Medical
University (Yinchuan, China) in October 2016. The present study was
approved by the Ethics Review Board of Ningxia Medical University
(approval no. 2015–049). Written informed consent was obtained from
the patient, and written and signed informed consent for
publication of the present study was also obtained. Cells were
cultured in DMEM with high glucose (4.5 g/l; HyClone; Cytiva)
containing 100 U penicillin and 0.1 mg/ml streptomycin with 10% FBS
(Clark Bioscience) in a humidified atmosphere containing 5%
CO2 at 37°C. HFFs labeled with green fluorescent protein
(GFP) by lentiviral infection (HFF-GFP). Lentiviral vector and
Polybrene (final concentration 5 µg/ml) were purchased from
Shanghai GeneChem Co., Ltd., and were used following the
manufacturers' protocols.
Cell induction
Plates were coated with Matrigel at room temperature
overnight. Then, HFFs were seeded into Matrigel-coated 24-well
plates (2×104 cells/well) and cultured overnight. The
fibroblast medium was replaced with neuronal induction medium
[DMEM/F12:Neurobasal (1:1) with 0.5% N2 supplement, 1% B27 and 100
mM cAMP], supplemented with 10 µM Forskolin (F), 1 µM RepSox (R),
10 µM SP600125 (S), 3 µM CHIR99021 (C), 5 µM GO6983 (G), 5 µM
Y-27632 (Y), 20 µM ISX-9 (I) and 2 µM I-BET151 (IB), all purchased
from PeproTech, Inc. Subsequently, culture media (neuronal
induction media plus the small molecular compounds) were refreshed
every 3–4 days during the induction period. Following small
molecular compound treatment for 7 days, induction media were
changed to neuronal maturation media [the neuronal induction medium
plus 20 ng/ml basic fibroblast growth factor (bFGF), 1% GlutaMAX,
20 ng/ml brain-derived neurotrophic factor (BDNF), 20 ng/ml glial
cell line-derived neurotrophic factor (GDNF) and 20 ng/ml
neurotrophin-3 (NT3), all purchased from PeproTech, Inc.],
supplemented with C+F+Y+I. Neuronal maturation media were changed
every other day. Small molecular compounds were dissolved and
diluted in dimethyl sulfoxide (Sigma-Aldrich; Merck KGaA) according
to the manufacturer's protocols and then processed to use at the
following final concentrations: 10 µM F, 1 µM R, 10 µM S, 3 µM C, 5
µM G, 5 µM Y, 20 µM I, 2 µM IB, 0.5 mM valproic acid (VPA) and 1 µM
Dorsomorphin (D). The concentrations of small molecules were
selected on the basis of previous literatures (8). All small molecular compounds were
purchased from Stemcell Technologies, Inc.
Immunofluorescent staining
Cells were grown on coverslips and fixed with 4%
formaldehyde for 10 min at room temperature and subsequently washed
with PBS containing 0.1% Tween-20 (PBST). After blocking at room
temperature with 5% normal goat serum (Beijing Solarbio Science
& Technology Co., Ltd.) in PBST for 30 min, cells were
incubated with the primary antibody overnight at 4°C, then washed
with PBST, and finally incubated with a goat anti-mouse IgG Alexa
Fluor 555-conjugated secondary antibody (1:1,000; cat. no. A-21422;
Thermo Fisher Scientific, Inc.) for 1 h at room temperature. Cells
were observed and imaged using laser scanning confocal microscopy
(Fluoview FV10; Olympus Corporation) and an inverted fluorescence
microscope (TS100; Nikon Corporation). The primary antibodies used
in the present study were as follows: mouse anti-class III
β-tubulin (Tuj1; 1:250; cat. no. 480011; Thermo Fisher Scientific,
Inc.), rabbit anti-microtubule-associated protein 2 (MAP2; 1:200;
cat. no. 17490–1-AP; ProteinTech Group, Inc.), rabbit
anti-vesicular glutamate transporter 1 (vGLUT1; 1:1,000; cat. no.
48-2400; Thermo Fisher Scientific, Inc.), rabbit
anti-γ-aminobutyric acid (GABA; 1:500; cat. no. PA5-32241; Thermo
Fisher Scientific, Inc.), rabbit anti-neuronal nuclei (NeuN; 1:200;
cat. no. 26975-1-AP; ProteinTech Group, Inc.), rabbit anti-OCT4
(1:500; cat. no. MA1-104; Thermo Fisher Scientific, Inc.) and
rabbit anti-glial fibrillary acidic protein (GFAP; 1:200; cat. no.
16825-1-AP; ProteinTech Group, Inc.). Semi-quantification was
performed using ImageJ software (version 1.44; National Institutes
of Health).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from cells using the
RNAsimple Total RNA kit (Tiangen Biotech Co., Ltd.) and reverse
transcribed to cDNA using the TIANScript RT kit (Tiangen Biotech
Co., Ltd.) according to the manufacturers' protocols. Thermocycling
conditions for reverse transcription were as follows: 42°C for 60
min and 85°C for 5 min. The mRNA expression levels were quantified
by SuperReal PreMix Plus (SYBR-Green) kit (Tiangen Biotech Co.,
Ltd.) using a Bio-Rad CFX Manager 3.0 (Bio-Rad Laboratories, Inc.).
Thermocycling conditions for qPCR were as follows: 95°C for 15 min,
followed by 40 cycles of 95°C for 10 sec, 60°C for 30 sec. Results
were confirmed in at least three separate analyses. The sequences
of gene primers are shown in Table
I. The relative mRNA levels were calculated using the
2−ΔΔCq method (13) and
normalized to GAPDH.
 | Table I.Primer sets for reverse
transcription-quantitative PCR. |
Table I.
Primer sets for reverse
transcription-quantitative PCR.
| Gene name | Sequence
(5′→3′) |
|---|
| CTGF | F:
GCGAAGCTGACCTGGAAGAGAAC |
|
| R:
GCTCGGTATGTCTTCATGCTGGTG |
| COL1A1 | F:
AGAACGCCAAGGACAAGAAGCAC |
|
| R:
CCATCAGACGCAGGAAGGTAAGC |
| DKK3 | F:
AGCTATCACAATGAGACCAACACAGAC |
|
| R:
GATGATGCACTCGTGGCTCCTTC |
| NeuroD1 | F:
CCTCGAAGCCATGAACGCAGAG |
|
| R:
TCGTCATCCTCCTCTTCCTCTTCTTC |
| ASCL1 | F:
ACAAGAAGATGAGTAAGGTGGAGACAC |
|
| R:
ATGGAGTTCAAGTCGTTGGAGTAGTTG |
| DCX | F:
GGATTGTGTACGCTGTGTCCTCTG |
|
| R:
TTCATCCATGCTTCCGATCTTCCTG |
| MAP2 | F:
TTGGTGCCGAGTGAGAAGAA |
|
| R:
AGGTCTGGCAGTGGTTGGTT |
| GAPDH | F:
CAGGAGGCATTGCTGATGAT |
|
| R:
GAAGGCTGGGGCTCATTT |
Electrophysiological analysis
For whole-cell patch-clamp recordings, the
artificial cerebrospinal fluid extracellular solution contained 140
mM NaCl, 5 mM KCl, 1 mM MgCl2, 1.8 mM CaCl2,
1.25 mM NaH2PO4, 23 mM glucose and 10 mM
HEPES (pH adjusted to 7.4 with NaOH). Patch pipettes (3–5 MΩ) were
filled with an internal solution containing 93 mM K-gluconate, 16
mM KCl, 2 mM MgCl2, 10 mM HEPES, 10 mM
Na-phosphocreatine, 0.3 mM GTP-Na2 and 4 mM ATP-Mg.
Sodium currents were recorded using the voltage clamp method. Step
voltages with an increment of 10 mV were injected to elicit an
inward sodium current. To block the sodium current, 1 µM
tetrodotoxin (TTX) was added into the chamber, and step voltages
were injected again to observe the TTX-sensitive currents 10 min
after applying TTX. Patch pipettes were pulled with a P97
micropipette puller to achieve a 3–6 MΩ tip resistance. Patch-clamp
recordings were taken using an EPC-10 amplifier (HEKA Elektronik
GmbH; Harvard Bioscience, Inc.) with PatchMaster 2.90 (HEKA
Elektronik GmbH; Harvard Bioscience, Inc.).
Western blot analysis
Cells were washed twice in PBS and proteins were
extracted with RIPA lysis buffer (Beyotime Institute of
Biotechnology) supplemented with protease inhibitor cocktail (cat.
no. P1008; Beyotime Institute of Biotechnology) and boiled at 100°C
for 5 min. Protein concentrations were measured by a bicinchoninic
acid assay (cat. no. P0012S; Beyotime Institute of Biotechnology).
Subsequently, 80 µg whole-cell extract proteins were separated on
10% SDS-polyacrylamide gels, and transferred to a polyvinylidine
difluoride membrane (EMD Millipore). Non-specific binding was
blocked by incubation in 5% skimmed milk in PBST for 2 h at room
temperature. Membranes were incubated at 4°C overnight with rabbit
anti-βШ-tubulin (1:1,000; cat. no. ab18207; Abcam), rabbit
anti-Synapsin 1 (1:1,000; cat. no. PA1-4673; Thermo Fisher
Scientific, Inc.) and rabbit anti-NeuN (1:1,000; cat. no.
26975-1-AP; ProteinTech Group, Inc.). Afterwards, the membranes
were washed in PBST and were incubated with appropriate
fluorescently labeled secondary antibodies for 1 h at room
temperature. The secondary antibodies used were: goat anti-rabbit
(IRDye® 680RD; 1:5,000; cat. no. 925-68071; LI-COR
Biosciences) and goat anti-mouse (IRDye® 680RD; 1:6,000;
cat. no. 925-68070; LI-COR Biosciences). The membranes were scanned
using the Odyssey infrared imaging system (LI-COR Biosciences), and
protein band intensity was semi-quantified by computerized
densitometry using ImageJ software 1.46r (National Institutes of
Health) and expressed as arbitrary units after normalization to
GAPDH (1:5,000; cat. no. PA1-988; Thermo Fisher Scientific,
Inc.).
Transmission electron microscopy
Cells were collected and washed briefly with a 0.1 M
sodium cacodylate buffer prior to fixation with 2.5% glutaraldehyde
in 0.1 M sodium cacodylate overnight at 4°C. After being rinsed for
10 min in the same buffer, the cells were fixed at 4°C for 1 h with
1% OsO4 in 0.1 M sodium cacodylate. Following
dehydration with a standard ethanol series and infiltration in
epoxy resin at room temperature overnight, cells were transferred
to beam capsules for polymerization in the oven (35°C for 6 h, 42°C
for 18 h and 60°C for 48 h). The capsules were separated from the
polymerized resin with a razor blade, and embedded cells in
hardened blocks were viewed with an optical microscope so that the
appropriate area was chosen for ultrathin sectioning (75 nm thick).
Subsequently, ultrathin sections were obtained using an
ultramicrotome with a diamond knife. Heavy metal staining was
performed at room temperature for 30 min with 4% uranyl acetate and
lead citrate, and the samples were examined under an electron
microscope (H-7800; Hitachi, Ltd.).
Mitochondrial staining
Cells were grown in confocal Petri dishes filled
with the appropriate culture medium. When cells reached the desired
confluence (~40%), an equal volume of the dye-working solution (AAT
Bioquest, Inc.) was added. The cells were incubated in a 5%
CO2 incubator at 37°C for 1 h. The cell nucleus was
stained with Hoechst 33342 (Sigma-Aldrich; Merck KGaA) at 37°C for
30 min. The cells were observed using a laser scanning confocal
microscope (Fluoview FV10; Olympus Corporation).
Statistical analysis
Data are presented as the mean ± standard deviation,
and experiments were performed at least three times. Statistical
analysis was performed using SPSS 17.0 software (SPSS, Inc.) and
data were assessed for normality and variance. Data were compared
using an unpaired Student's t-test or one-way analysis of variance
with a Bonferroni post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Screening for optimal small molecular
compounds
The present study was designed to refine small
molecular compounds able to reprogram HFFs into functional neurons.
The initial HFFs derived from the foreskin of a 20-year-old man
showed no contamination of neuronal cells or neural stem cells
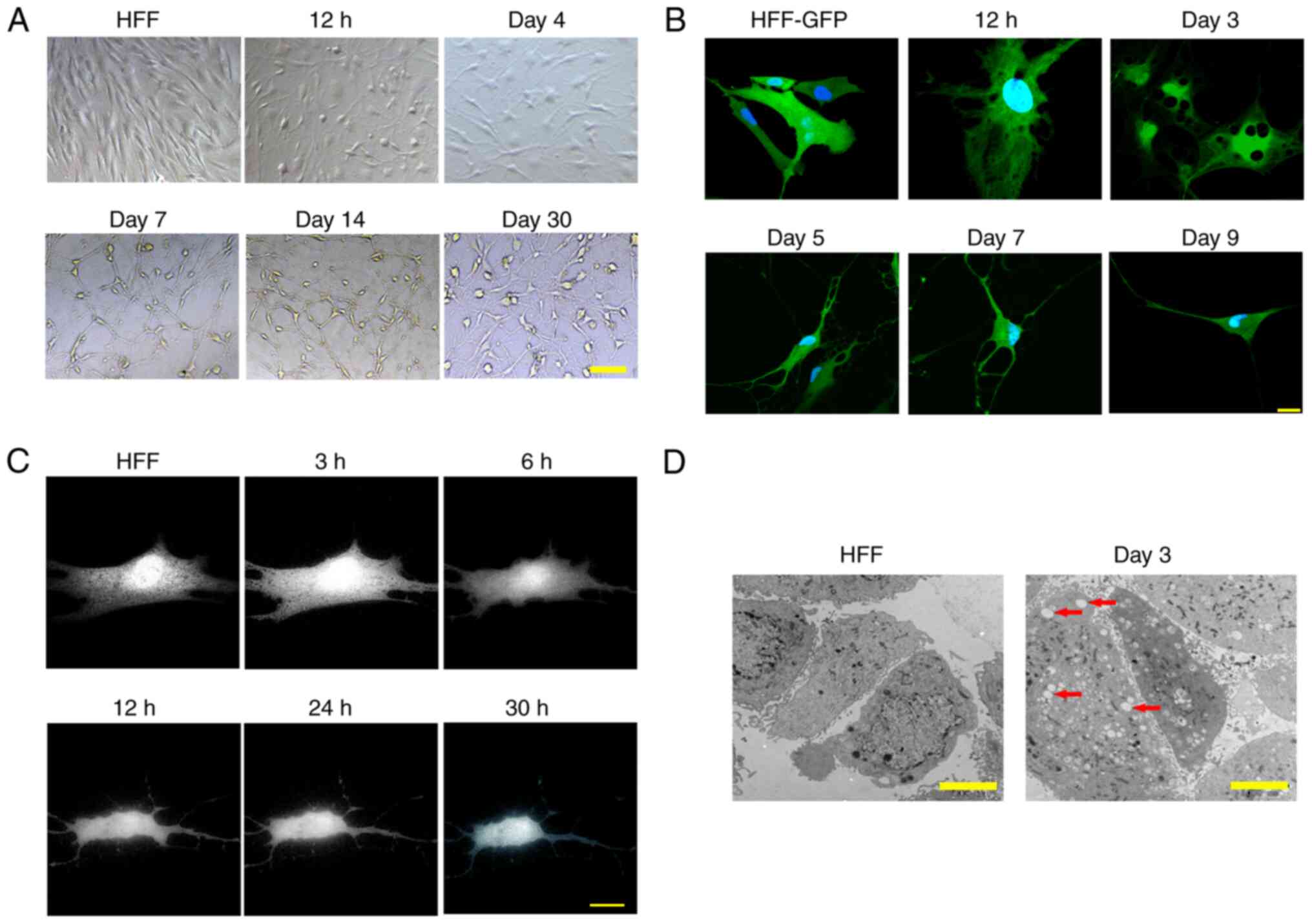
(Fig. 1A). A total of 11 small
molecular compounds were initially included as the starting
candidate pool based on three major criteria of selection that a
compound has: i) the capacity to disrupt fibroblast-specific
programs, such as IB (9); ii) the
ability to activate neural reprogramming related signaling
pathways, to enhance neuronal gene expression, to promote neuronal
conversion of human fibroblasts and reprogramming efficiency, such
as valproic acid (VPA), F, C, G, S, I, purmorphamine (P) and D
(8,9,14–16);
and iii) the potential to promote neuronal survival, such as R and
Y (15,17). The HFFs were treated with a
neuronal induction medium containing F, F+R and F+R+S, and no
neuron-specific marker Tuj1-positive cells were detected on day 4
(Fig. 1B). The HFFs were treated
with F+R+S+C, an agent suggested to improve neuronal conversion of
HFFs (8,18), and 5% of cells expressed Tuj1
(Fig. 1B). In order to improve the
induction efficiency, small molecular compounds G+Y were
supplemented based on the original F+R+S+C. The number of
Tuj1-positive cells was increased to 8% in this medium, but the
cells did not present a typical neuronal morphology (Fig. 1B), which suggested that they were
inefficiently reprogrammed (Fig.
1B). Notably, almost all cells expressed the neuronal maker
Tuj1 following treatment with eight small molecular compounds
(F+R+S+C+G+Y+I+IB) on day 4 (Fig.
1B). Under these conditions, the induced cells exhibited a
bipolar neuron-like cell morphology, particularly on day 7
(Fig. 1C). Notably, there was no
significant difference in the number of both Tuj1-positive and
DAPI-positive cells between those cultured in a medium with or
without V, P and D. Therefore, compounds V, D and P were removed
from the combination of small molecular compounds (Fig. 1D). In order to promote neuronal
maturation following fibroblast-neuron conversion, compounds
R+S+G+IB were also removed from the medium and replaced with
neurotrophic factors (BDNF, GDNF and NT3) on day 7 of culture.
Finally, a cocktail containing eight small molecular compounds (F,
R, S, C, G, Y, I and IB) was used to reprogram human fibroblasts
into neurons, and the compounds were added into the culture medium
in a stepwise manner.
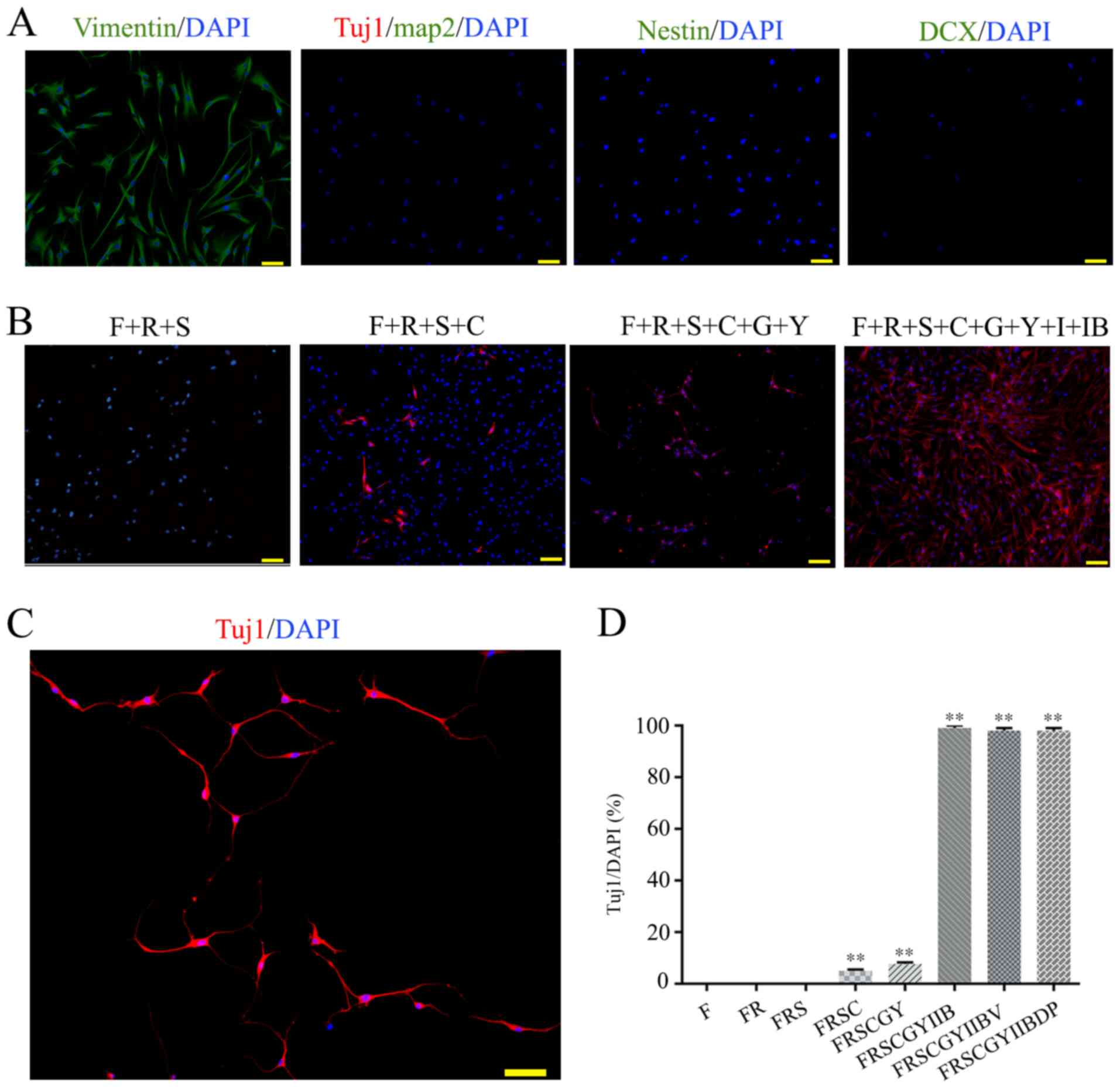 | Figure 1.Characterization of HFFs and optimal
small molecule induction protocol. (A) Immunofluorescence staining
of the HFFs. Fibroblast marker, vimentin; neuronal markers, MAP2,
Tuj1, nestin and DCX. (B) Optimal small molecule combination.
Immunofluorescence staining of HFFs reprogrammed with F, R, S, C,
G, Y, I and IB on day 4. (C) Immunofluorescence staining of Tuj1
(red) on day 7. (D) Percentage of Tuj1+/DAPI after
treatment of HFFs with different small molecular compounds. Cells
from at least five random fields of view in each of the three wells
from three independent experiments were counted. Scale bar, 50 µm.
Cell nuclei were stained with DAPI. Data are presented as the mean
± SD. **P<0.01 vs. F only group. F, forskolin; R, RepSox; S,
SP600125; C, CHIR99021; G, Go6983; Y, Y-27632; I, IXS9; IB,
I-BET151; Tuj1, class III β-tubulin; DCX, doublecortex; MAP2,
microtubule-associated protein 2; HFF, human foreskin
fibroblast. |
Morphological alterations of cells
during induction
When recording the phenotypical changes of cells
between days 1 and 30 of induction, gradual morphological changes
of the HFFs from fibroblasts to typical neurons were observed at 12
h after the addition of small molecular compounds (Fig. 2A). The cells then underwent robust
morphological changes with a decreased cell volume and pervasive
neurite extension during the first 24 h of induction, as determined
by a time course live cell imaging (Fig. 2C). Due to the long-term
reprogramming progression, the induced cells were designated as
induced neurons (iNs) in the present study. The iNs exhibited a
typical neuronal phenotype on day 7 (Fig. 1C). Notably, HFFs labeled with GFP
by lentiviral infection (HFF-GFP) showed small circular blebs in
the cytoplasm at 12 h post induction, which reached the peak on day
3 (Fig. 2B). With the increased
number of blebs, small blebs gradually merged into large blebs in
the cytoplasm before they disappeared on day 9 (Fig. 2B). This finding was further
confirmed by electron microscopy (black arrows; Fig. 2D).
Reprogramming human fibroblasts into
neurons using small molecular compounds
After demonstrating that small molecular compounds
can induce HFFs to express Tuj1, a cocktail of eight small
molecular compounds was demonstrated to induce cells to express
Tuj1 and MAP2 with typical neuron-like morphology by day 7
(Figs. 1C and 3A). Most of these iNs were simultaneously
expressing Tuj1 and MAP2, accompanied by an extensive neurite
outgrowth following 1–2 weeks of induction (Fig. 3A and B). The efficiency of
conversion from fibroblasts into neurons induced by this
combination was at most, >92% and 82.66±2.52% after 7–14 and
14–30 days of induction, respectively (data not shown). This
suggests an efficient conversion acquired by the combination of
small molecular compounds. As the senescence of neurons tended to
occur at maturation at ~7 days, the inductive medium was replaced
with the neuronal maturation medium supplemented with C+F+Y+I and
extra neurotropic factors (BDNF, GDNF and NT3) 9 days after the
initial conversion. This was able to sustain neuronal cell survival
for >30 days, as determined by immunofluorescent staining of
neuronal markers Tuj1, MAP2, NeuN, GABA and vGLUT1 (Fig. 3B). However, the expression levels
of astrocyte marker GFAP and neural stem cell marker OCT4 were
undetectable (data not shown).
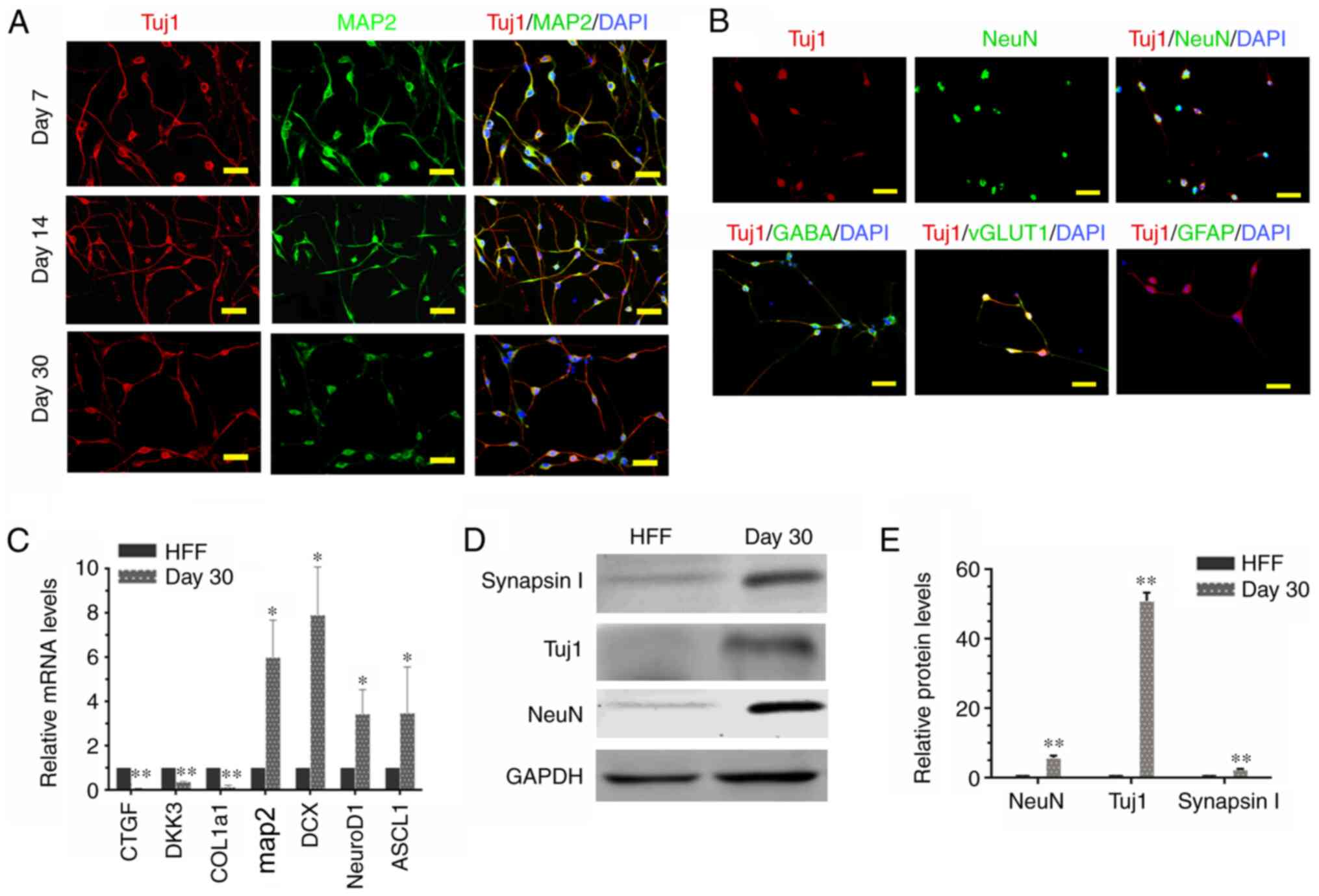 | Figure 3.Characteristics of iNs. (A) iNs
exhibited bipolar neuronal morphologies and expressed tuj1 (red)
and MAP2 (green) at different phases (day 7, 14 and 30). (B) iNs
were stained for NeuN, GABA, vGLUT1 and GFAP (all green) on day 30.
Scale bar, 50 µm. (C) Reverse transcription-quantitative PCR
analysis revealed that in comparison with HFFs, there was a
significant increase in gene expression levels of MAP2, DCX,
NeuroD1 and ASCL1 in iNs, whereas human fibroblast-specific genes
were downregulated in iNs on day 30. Data are presented as the mean
± SD. (D) Western blot analysis of the protein expression levels of
Tuj1, NeuN and GAPDH in HFFs and iNs. (E) Graphs show the
semi-quantification of the western blot analysis. n=3 independent
experiments. Data are presented as the mean ± SD. Unpaired
Student's t-tests were used to compare data. *P<0.05,
**P<0.01 vs. HFF. iNs, induced neurons; Tuj1, class III
β-tubulin; MAP2, microtubule-associated protein 2; HFF, human
foreskin fibroblast; NeuN, neuronal nuclei; GABA, γ-aminobutyric
acid; vGLUT1, vesicular glutamate transporter 1; GFAP, glial
fibrillary acidic protein; DCX, doublecortin; Ascl1, achaete-scute
family bHLH transcription factor 1; CTGF, connective tissue growth
factor; DKK3, dickkopf WNT signaling pathway inhibitor 3; COL1a1,
collagen type 1 α-1 chain. |
To further characterize the iN phenotype, the
expression levels of four neuron-specific genes and three human
fibroblast-specific genes were determined by RT-qPCR in cells
cultured for 30 days following small molecular compound treatment.
Compared with control HFF (treated with DMSO), a significant
increase in the expression levels of several neuron-specific genes,
including MAP2, doublecortin, neuronal differentiation 1 (NeuroD1)
and achaete-scute family bHLH transcription factor 1 (Ascl1), was
observed (Fig. 3C). By contrast,
significantly decreased expression levels of human
fibroblast-specific genes, including dickkopf WNT signaling pathway
inhibitor 3, connective tissue growth factor and collagen type 1
α-1 chain, were also identified (Fig.
3C). Several lines of evidence have demonstrated that
fibroblasts are able to directly transdifferentiate into functional
neurons using transcription factors (19,20).
As expected, 4-fold higher transcription levels of NeuroD1 and
Ascl1 genes were gradually increased with the induction course with
a peak on day 7, as compared with untreated HFFs (Fig. 3C). In addition, the alterations in
neuronal marker expression were also demonstrated at the protein
expression levels by immunoblotting assays in iNs after 30 days of
induction (Fig. 3D and E).
Overall, these results indicated an activation of neuron-specific
genes and inhibition of fibroblast-specific genes in cells exposed
to the cocktail of small molecular compounds, suggesting that a
combination of small molecular compounds (F+R+S+C+G+Y+I+IB) was
able to efficiently convert HFFs into neurons and to bypass the
pluripotent stem stage.
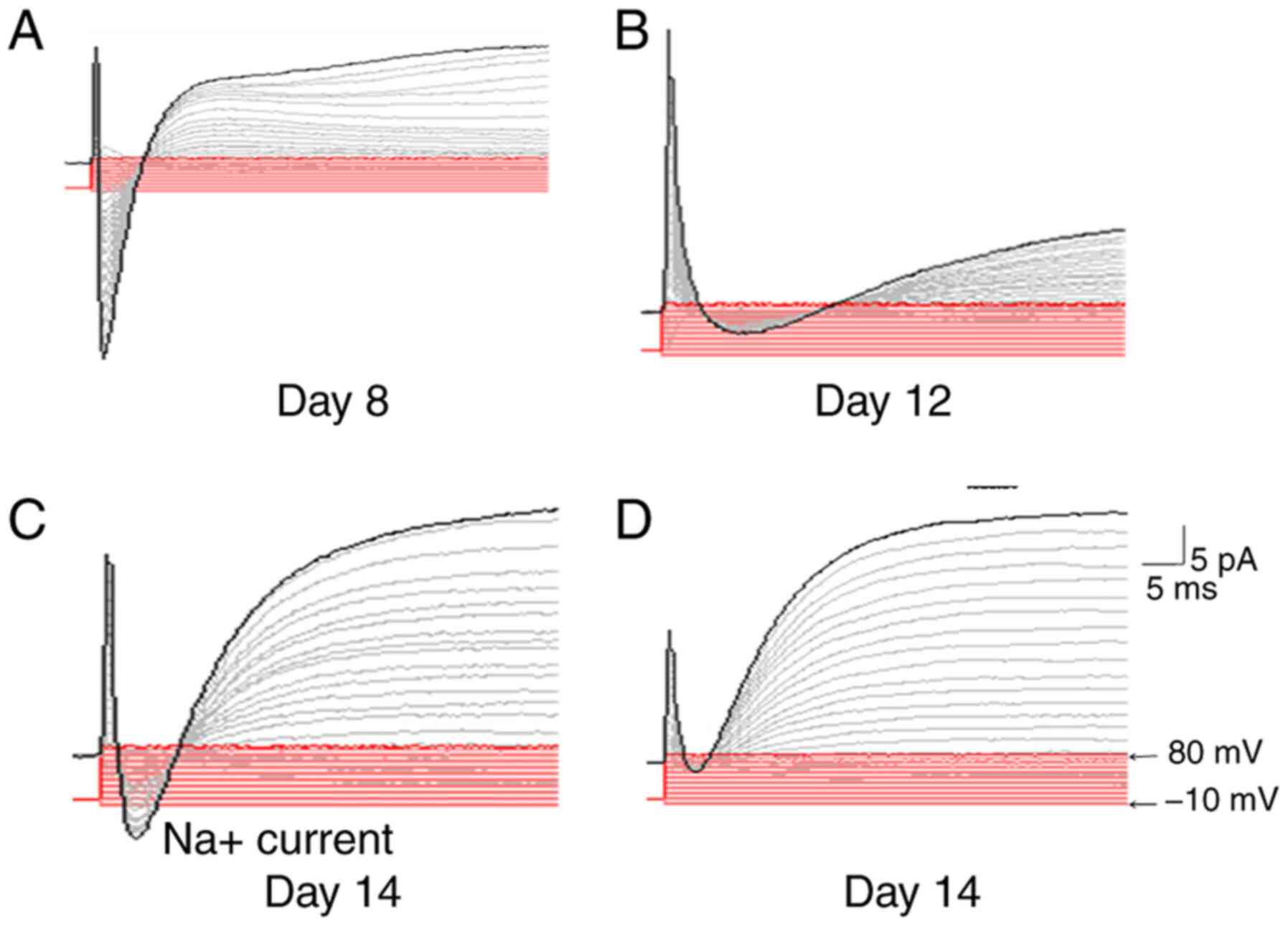
Physiological properties of iNs
To assess whether the iNs from HFFs possess
electrophysiological properties, such as the induction of a
membrane current, electrophysiological recordings of neurons at
different induction stages were performed. None of the cells
exposed to the treatment compounds exhibited functional membrane
properties after 8 days of induction (Fig. 4A). However, the inward and outward
currents could be recorded in cells after 12 days of induction,
although the recorded curve was not a typical sodium current and
potassium current curve (Fig. 4B).
Typical curves of fast inward currents and outward currents were
recorded in cells at day 14 post-induction, which might correspond
to the opening of voltage-dependent K+ and
Na+ channels (Fig. 4C).
Furthermore, the inward current could be completely blocked by the
sodium channel-specific inhibitor TTX (Fig. 4D), which indicated that the iNs had
functional membrane properties.
Discussion
Previous studies have demonstrated that fibroblasts
can be directly converted into functional neuronal stem cells,
neurons, neural progenitors and astrocytes by upregulation of
specific transcription factors or microRNAs (21–24).
However, inefficient induction and integration of exogenous genes
into the genome limit their translation into clinical applications.
Small molecular compounds that target signaling pathways,
epigenetic modifications or metabolic processes exhibit abilities
to regulate cell development, cell fate and function (9,10,25).
Therefore, small molecular compounds (chemical reprogramming) that
induce cell lineage reprogramming have been widely favored due to
their non-immunogenicity, low cost and easy optimization (8). Several studies have reported that
small molecular compounds induce fibroblasts to differentiate into
neurons (8–10). However, the induction efficiency
and cell morphological changes during induction remain unclear. It
is well known that human fibroblasts are bulky, mostly fusiform or
star-shaped flat structures, while neurons are relatively small in
volume and have long axons (10).
The transformation from a relatively large fibroblast into a long
axon neuron needs to be observed. The present study first used a
cocktail of small molecular compounds that are able to efficiently
induce a direct lineage reprogramming from human fibroblasts into
neurons. These iNs possessed neuronal properties in terms of gene
expression pattern and electrophysiological functional
capabilities. Furthermore, the course of morphological changes of
cells was recorded during the induction by live cell imaging under
a fluorescent microscope.
The cAMP signaling pathway may reduce the barriers
of reprogramming by altering downstream epithelial gene expression,
decreasing mesenchymal gene expression and increasing cell
proliferation (26). It has been
reported that activation of the cAMP signaling pathway by forskolin
sufficiently generates OCT4-positive colonies in the absence of the
OCT4 transgene and replaces OCT4 to induce pluripotency (26,27).
R can be used to replace Sox2 and c-Myc via inhibition of the TGF-β
signaling pathway (28).
CHIR99021, an agonist of WNT-β-catenin signaling, promotes ESC
self-renewal by potentially upregulating transcription of the
protein signal transducer and activator of transcription-3
(29), and promotes the conversion
of fibroblasts into neurons (8,9). A
combination of F, C, R, S and G has been demonstrated to
successfully differentiate fibroblasts into neurons (8). In addition, IB disrupts the
fibroblast-specific program, while the neurogenesis inducer I is
indispensable to activate neuron-specific genes (9). Y is able to increase primary
keratinocyte proliferation and immortalization without detectable
cell senescence (30). Based on
previous knowledge regarding the transformation of human
fibroblasts into neurons, a cocktail containing eight small
molecular compounds (F+R+S+C+G+Y+I+IB) was used for the induction
of HFFs into functional neurons. VPA has been demonstrated to
enhance the efficiency of the OCT4, Sox2, Kruppel-like factor 4
(Klf4) and c-Myc four-factor reprogramming mouse fibroblasts
(31,32). Despite this, VPA did not appear to
serve a notable role in the present study.
In addition, the morphological changes of cells
during direct conversion induced by small molecular compounds were
recorded in the present study. The morphology of the cells changed
markedly; in particular, the cell bodies gradually shrank and the
axons gradually formed with time. Additionally, there were
differences in the number and structural distribution of
mitochondria, in comparison with fibroblasts. The major functions
of mitochondria are aerobic energy production, reactive oxygen
species production, calcium homeostasis, cellular signaling
pathways, and synthesis and/or assembly of cellular metabolites
(33). Several studies have
reported that mitochondria serve an important role in cell
reprogramming (34,35). The significantly increased number
and dynamic changes of the mitochondria suggest an increased
requirement of energy support during cell differentiation, which
may have an important role in the differentiation of human
fibroblasts into neurons by small molecular compounds. The
underlying mechanism requires further investigation.
It has been documented that the transcriptional
program underlying embryonic neurogenesis is regulated by the
spatiotemporal expression pattern of critical transcription factors
(19,36). Direct conversion of fibroblasts
into functional neurons has been demonstrated using transcription
factors (18,37). NeuroD1 is a basic Helix-Loop-Helix
(bHLH) transcription factor that serves an important role during
neuronal differentiation, is essential for eliciting the neuronal
development program and possesses the ability to reprogram other
cell types into neurons (38,39).
Therefore, it may serve an important role in reprogramming
fibroblasts into neurons. The expression levels of NeuroD1
transcript were only increased 3-fold in iNs compared with human
fibroblasts, and this was not significantly different after 30 days
of induction. It was reasoned that the neurons tend to mature after
30 days of induction, and the expression levels of transcription
factors might decrease as well. This notion was supported by a
previous report demonstrating that in cultured cortical neurons,
expression levels of neuroD/BETA2 were decreased with increasing
days in culture, as neurons became mature (40). neuroD/BETA2 is expressed highly in
immature neurons (40). Indeed,
the transcription of NeuroD1 was increased gradually as induction
progressed, and reached a peak on day 7, increasing as much as
4-fold compared with human fibroblasts, before it began to decline
by 4-fold on day 10 compared with human fibroblasts. In addition,
NeuroD1 is able to override the presence of stimuli for
pluripotency, inducing neuronal gene expression and repressing core
pluripotency genes (OCT4, Klf4 and Nanog) (38). Therefore, the present study tested
whether induction of iNs from HFFs using the cocktail of eight
small molecular compounds bypassed the pluripotent stem stage.
Results indicated that the pluripotency gene OCT4 was not induced
in the iNs at any induction stage, which suggested that a direct
conversion of HFFs to differentiate into iNs occurred, bypassing
the pluripotent stage. In this regard, NeuroD1 may inhibit the
expression of pluripotent genes and thus achieve direct
trans-differentiation. Ascl1 is another bHLH factor and is a
crucial regulator of neurogenesis during normal development
(41). Ascl1 is a pioneering
transcription factor that initiates the iN-reprogramming process by
subsequently aiding other factors to secure the reprogramming route
toward neuronal direction (42).
In this respect, it is also a central and essential component of
direct reprogramming of mouse or human fibroblasts to iNs (37,43).
In the present experiments, a similar trend in Ascl1 expression was
determined. The peak expression was observed on day 7 and
expression gradually decreased by 2-fold compared with that in
fibroblasts on day 30 of induction. In addition to the
aforementioned findings, both bHLH factors NeuroD1 and Ascl1 were
significantly elevated on day 7 of induction in the present study,
which suggested that activation of bHLH factors may be a key to
alter cell fate induced by the cocktail of eight small molecular
compounds.
The present study has demonstrated the potential of
the cocktail of eight small molecular compounds in the induction of
HFF trans-differentiation into neurons. The morphology of cells was
markedly altered during induction. Mechanistically, the induction
of differentiation was at least in part accompanied by increased
expression of endogenous neuronal transcription factors, such as
the bHLH factors NeuroD1 and Ascl1, and decreased expression of
fibroblast-specific genes. Further studies are required to
replicate these results in other types of fibroblasts, such as skin
and lung fibroblasts. Collectively, the present study suggested
that the activation of specific endogenous transcription factors
and cell morphology intervention by small molecular compounds
further increases the induction efficiency of HFF differentiation
into neurons, which may provide a novel approach for the generation
of neuronal resources in cell replacement therapy of neurological
disorders.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Scientific
Research Project of Ningxia Medical University (grant nos.
XT2019024 and XY201712), the West China Top Class Discipline
Project in Basic Medical Sciences, Ningxia Medical University
(grant no. NXYLXK2017B07), the National Natural Science Foundation
of China (grant nos. 31460300 and 81860132) and the Major Science
and Technology Projects for the 13th Five-Year Plan of Ningxia Hui
Autonomous Region (grant no. 2016BZ07).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding authors on reasonable
request.
Authors' contributions
JY, HC, SC and QH conceived and designed the study.
JY, HC, SG and HZ designed the concept and performed the
experiments. JY, HC, HT, LZ and ZC conducted statistical analyses,
interpreted the results and searched the literature. TS collected
the foreskin sample from the patient and performed cell culture. JY
and QH wrote the manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics Review
Board of Ningxia Medical University (approval no. 2015-049).
Written informed consent was obtained from the participant enrolled
in the present study.
Patient consent for publication
Written informed consent for publication of the
study was obtained from the patient.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ESCs
|
embryonic stem cells
|
|
HFFs
|
human foreskin fibroblasts
|
|
PBST
|
PBS with 0.1% Tween-20
|
|
TTX
|
tetrodotoxin
|
References
|
1
|
Kumar A, Narayanan K, Chaudhary RK, Mishra
S, Kumar S, Vinoth KJ, Padmanabhan P and Gulyás B: Current
perspective of stem cell therapy in neurodegenerative and metabolic
diseases. Mol Neurobiol. 54:7276–7296. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chanda S, Marro S, Wernig M and Sudhof TC:
Neurons generated by direct conversion of fibroblasts reproduce
synaptic phenotype caused by autism-associated neuroligin-3
mutation. Proc Natl Acad Sci USA. 110:16622–16627. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu ML, Zang T and Zhang CL: Direct
lineage reprogramming reveals disease-specific phenotypes of motor
neurons from human ALS patients. Cell Rep. 14:115–128. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sahni V and Kessler JA: Stem cell
therapies for spinal cord injury. Nat Rev Neurol. 6:363–372. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Takahashi K and Yamanaka S: Induction of
pluripotent stem cells from mouse embryonic and adult fibroblast
cultures by defined factors. Cell. 126:663–676. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hyun I, Lindvall O, Ahrlund-Richter L,
Cattaneo E, Cavazzana-Calvo M, Cossu G, De Luca M, Fox IJ, Gerstle
C, Goldstein RA, et al: New ISSCR guidelines underscore major
principles for responsible translational stem cell research. Cell
Stem Cell. 3:607–609. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rosemann A: Stem cell treatments for
neurodegenerative diseases: Challenges from a science, business and
healthcare perspective. Neurodegener Dis Manag. 5:85–87. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu W, Qiu B, Guan W, Wang Q, Wang M, Li W,
Gao L, Shen L, Huang Y, Xie G, et al: Direct conversion of normal
and Alzheimer's disease human fibroblasts into neuronal cells by
small molecules. Cell Stem Cell. 17:204–212. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li X, Zuo X, Jing J, Ma Y, Wang J, Liu D,
Zhu J, Du X, Xiong L, Du Y, et al: Small-molecule-driven direct
reprogramming of mouse fibroblasts into functional neurons. Cell
Stem Cell. 17:195–203. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pfisterer U, Ek F, Lang S, Soneji S,
Olsson R and Parmar M: Small molecules increase direct neural
conversion of human fibroblasts. Sci Rep. 6:382902016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qin H, Zhao A, Ma K and Fu X: Chemical
conversion of human and mouse fibroblasts into motor neurons. Sci
China Life Sci. 61:1151–1167. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yang Y, Chen R, Wu X, Zhao Y, Fan Y, Xiao
Z, Han J, Sun L, Wang X and Dai J: Rapid and efficient conversion
of human fibroblasts into functional neurons by small molecules.
Stem Cell Reports. 13:862–876. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huangfu D, Maehr R, Guo W, Eijkelenboom A,
Snitow M, Chen AE and Melton DA: Induction of pluripotent stem
cells by defined factors is greatly improved by small-molecule
compounds. Nat Biotechnol. 26:795–797. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ladewig J, Mertens J, Kesavan J, Doerr J,
Poppe D, Glaue F, Herms S, Wernet P, Kögler G, Müller FJ, et al:
Small molecules enable highly efficient neuronal conversion of
human fibroblasts. Nat Methods. 9:575–578. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Madhu V, Dighe AS, Cui Q and Deal DN: Dual
inhibition of activin/Nodal/TGF-β and BMP signaling pathways by
SB431542 and dorsomorphin induces neuronal differentiation of human
adipose derived stem cells. Stem Cells Int. 2016:10353742016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lamas NJ, Johnson-Kerner B, Roybon L, Kim
YA, Garcia-Diaz A, Wichterle H and Henderson CE: Neurotrophic
requirements of human motor neurons defined using amplified and
purified stem cell-derived cultures. PLoS One. 9:e1103242014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li W, Zhou H, Abujarour R, Zhu S, Young
Joo J, Lin T, Hao E, Schöler HR, Hayek A and Ding S: Generation of
human-induced pluripotent stem cells in the absence of exogenous
Sox2. Stem Cells. 27:2992–3000. 2009.PubMed/NCBI
|
|
19
|
Ambasudhan R, Talantova M, Coleman R, Yuan
X, Zhu S, Lipton SA and Ding S: Direct reprogramming of adult human
fibroblasts to functional neurons under defined conditions. Cell
Stem Cell. 9:113–118. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Guillemot F: Spatial and temporal
specification of neural fates by transcription factor codes.
Development. 134:3771–3780. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pang ZP, Yang N, Vierbuchen T, Ostermeier
A, Fuentes DR, Yang TQ, Citri A, Sebastiano V, Marro S, Südhof TC
and Wernig M: Induction of human neuronal cells by defined
transcription factors. Nature. 476:220–223. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hou PS, Chuang CY, Yeh CH, Chiang W, Liu
HJ, Lin TN and Kuo HC: Direct conversion of human fibroblasts into
neural progenitors using transcription factors enriched in human
ESC-Derived neural progenitors. Stem Cell Reports. 8:54–68. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim SM, Flasskamp H, Hermann A,
Araúzo-Bravo MJ, Lee SC, Lee SH, Seo EH, Lee SH, Storch A, Lee HT,
et al: Direct conversion of mouse fibroblasts into induced neural
stem cells. Nat Protoc. 9:871–881. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Caiazzo M, Giannelli S, Valente P, Lignani
G, Carissimo A, Sessa A, Colasante G, Bartolomeo R, Massimino L,
Ferroni S, et al: Direct conversion of fibroblasts into functional
astrocytes by defined transcription factors. Stem Cell Reports.
4:25–36. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Biswas D and Jiang P: Chemically induced
reprogramming of somatic cells to pluripotent stem cells and neural
cells. Int J Mol Sci. 17:2262016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fritz AL, Adil MM, Mao SR and Schaffer DV:
cAMP and EPAC signaling functionally replace OCT4 during induced
pluripotent stem cell reprogramming. Mol Ther. 23:952–963. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hou P, Li Y, Zhang X, Liu C, Guan J, Li H,
Zhao T, Ye J, Yang W, Liu K, et al: Pluripotent stem cells induced
from mouse somatic cells by small-molecule compounds. Science.
341:651–654. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ichida JK, Blanchard J, Lam K, Son EY,
Chung JE, Egli D, Loh KM, Carter AC, Di Giorgio FP, Koszka K, et
al: A small-molecule inhibitor of tgf-Beta signaling replaces sox2
in reprogramming by inducing nanog. Cell Stem Cell. 5:491–503.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sato N, Meijer L, Skaltsounis L, Greengard
P and Brivanlou AH: Maintenance of pluripotency in human and mouse
embryonic stem cells through activation of Wnt signaling by a
pharmacological GSK-3-specific inhibitor. Nat Med. 10:55–63. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chapman S, Liu X, Meyers C, Schlegel R and
McBride AA: Human keratinocytes are efficiently immortalized by a
Rho kinase inhibitor. J Clin Invest. 120:2619–2626. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huangfu D, Osafune K, Maehr R, Guo W,
Eijkelenboom A, Chen S, Muhlestein W and Melton DA: Induction of
pluripotent stem cells from primary human fibroblasts with only
Oct4 and Sox2. Nat Biotechnol. 26:1269–1275. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yoshida Y, Takahashi K, Okita K, Ichisaka
T and Yamanaka S: Hypoxia enhances the generation of induced
pluripotent stem cells. Cell Stem Cell. 5:237–241. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dimmer KS and Scorrano L: (De)constructing
mitochondria: What for? Physiology (Bethesda). 21:233–241.
2006.PubMed/NCBI
|
|
34
|
Prieto J, Leon M, Ponsoda X, García-García
F, Bort R, Serna E, Barneo-Muñoz M, Palau F, Dopazo J, López-García
C and Torres J: Dysfunctional mitochondrial fission impairs cell
reprogramming. Cell Cycle. 15:3240–3250. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Prieto J, Leon M, Ponsoda X, Sendra R,
Bort R, Ferrer-Lorente R, Raya A, López-García C and Torres J:
Early ERK1/2 activation promotes DRP1-dependent mitochondrial
fission necessary for cell reprogramming. Nat Commun. 7:111242016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Spitz F and Furlong EE: Transcription
factors: From enhancer binding to developmental control. Nat Rev
Genet. 13:613–626. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Vierbuchen T, Ostermeier A, Pang ZP,
Kokubu Y, Sudhof TC and Wernig M: Direct conversion of fibroblasts
to functional neurons by defined factors. Nature. 463:1035–1041.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pataskar A, Jung J, Smialowski P, Noack F,
Calegari F, Straub T and Tiwari VK: NeuroD1 reprograms chromatin
and transcription factor landscapes to induce the neuronal program.
EMBO J. 35:24–45. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Aprea J, Nonaka-Kinoshita M and Calegari
F: Generation and characterization of Neurod1-CreER(T2) mouse lines
for the study of embryonic and adult neurogenesis. Genesis.
52:870–878. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lee JK, Cho JH, Hwang WS, Lee YD, Reu DS
and Suh-Kim H: Expression of neuroD/BETA2 in mitotic and
postmitotic neuronal cells during the development of nervous
system. Dev Dyn. 217:361–367. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Imayoshi I and Kageyama R: bHLH factors in
self-renewal, multipotency, and fate choice of neural progenitor
cells. Neuron. 82:9–23. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wapinski OL, Vierbuchen T, Qu K, Lee QY,
Chanda S, Fuentes DR, Giresi PG, Ng YH, Marro S, Neff NF, et al:
Hierarchical mechanisms for direct reprogramming of fibroblasts to
neurons. Cell. 155:621–635. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shi Z, Zhang J, Chen S, Li Y, Lei X, Qiao
H, Zhu Q, Hu B, Zhou Q and Jiao J: Conversion of fibroblasts to
parvalbumin neurons by one transcription factor, Ascl1, and the
chemical compound forskolin. J Biol Chem. 291:13560–13570. 2016.
View Article : Google Scholar : PubMed/NCBI
|