Introduction
The cancer-related mortality rate increases annually
on a global scale, making cancer the leading cause of death in
South Korea (1,2). The incidence of skin cancer is rising
faster compared with other types of cancer, concurrent with
increasing outdoor activity and subsequent exposure to ultraviolet
radiation. Comprising only 4% of all types of cancer, but with a
mortality rate approaching 80%, melanoma is considered to be highly
malignant, and it is caused by the malignant alteration of melanin
cells (3,4). Melanoma spreads rapidly through the
lymphatic ducts and blood vessels to internal organs, such as the
liver, lungs and bones, and it is highly resistant to chemotherapy
and radiotherapy. Therefore, surgical excision following early
diagnosis is considered as the only treatment for melanoma. In
addition, melanoma is characterized as an intractable disease, as
there is currently no evidence to support any treatment to prevent
disease recurrence (5). Due to an
increasing melanoma occurrence and high mortality rate, a number of
studies on melanoma have been conducted in the western world;
however, measures should be taken to encourage further studies in
Korea, where the number of studies is limited (6). Previously, several international and
Korean studies have been performed aiming to reduce the side
effects and improve the anticancer effects of chemotherapy using
carcinostatic substances made of natural compounds isolated from
animals and plants (7–11). Widely distributed throughout the
vegetable kingdom, flavonoids are known to exist in ~4,000 types of
water-soluble pigments in various vegetables and fruits, such as
green tea, grapes and onions. Functional studies have reported that
flavonoids exhibit several bioactivities (12–14).
The flavonoid 4′,5,7,-trihydroxyflavone (apigenin) has been
demonstrated to exert an inhibitory effect on cancer cell growth
and several other effects including anti-oxidant (15), anti-inflammatory (16) and anti-tumor (17) effects in vitro and in
vivo.
Apigenin, an aromatic compound, is found in various
fruits and plants, including parsley, onions, oranges, tea,
chamomile, wheat and other seasonings, and has no evident toxicity
(18). Several studies have been
conducted on the antioxidant (19), anti-inflammatory (20) and anti-viral activities (21) of apigenin. Apigenin has been
reported to affect cancer cell growth by promoting apoptosis and
inhibiting the cell cycle of colorectal cancer cells (22), and inducing apoptosis via
inhibiting angiogenesis in breast and prostate cancer cells
(23,24). These findings suggest that apigenin
could be considered as a potential carcinostatic substance. In
addition, apigenin has been demonstrated to have cytotoxic and
anti-proliferative activities by promoting G2/M cell cycle arrest
in melanoma cells (25), as well
as the ability to attenuate tumor invasion and metastatic potential
via inhibiting lung colonization (26). Furthermore, apigenin can impede
metastasis of melanoma cells by impairing interactions between
tumor cells and endothelial cells (27).
Apoptosis is known to inhibit the development and
progression of cancer, and is distinguished from necrosis as a
defense mechanism that voluntarily eliminates cells with
accumulated DNA damage caused by intracellular DNA damage and viral
infection (28). The Bcl-2 family,
which is commonly known to suppress the genesis and progression of
cancer through apoptosis, is classified into pro- and
anti-apoptotic proteins (29). The
pro-apoptotic proteins include Bax, BH3-interacting domain death
agonist (Bid) and Bad, which induce apoptosis via rupturing the
outer mitochondrial membrane, whereas anti-apoptotic Bcl-2, Bcl-xL
and A1 proteins inhibit apoptosis by preserving the outer
mitochondrial membrane (30–32).
The phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) and
mitogen-activated protein kinase (MAPK) pathways are among the
signals associated with cell viability; they activate various
intracellular signaling pathways to regulate cell proliferation and
angiogenesis (33,34). As an enzyme that phosphorylates
serine/threonine residues, Akt is known to be involved in cell
cycle progression and survival-related proliferation, and inhibits
apoptosis via downregulating the expression of the pro-apoptotic
proteins Bcl-2 and caspase-9 via the PI3K/Akt pathway (35). The MAPK pathway includes three main
kinases, namely extracellular signal-regulated protein kinase
(ERK), c-Jun N-terminal kinase (JNK) and p38 MAPK kinase, each with
different activities, that suppress tumor growth and cell
differentiation (36).
The present study aimed to explore the inhibitory
effect of apigenin on human melanoma A375P and A375SM cell
proliferation and its inductive activity on apoptosis, and to
further elucidate whether the Akt and MAPK signaling pathways
mediated the apigenin-induced cell apoptosis.
Materials and methods
Reagents
The human melanoma cell lines, A375P and A375SM,
were obtained from the Korean Cell Line Bank (Korean Cell Line
Research Foundation). Dulbecco's modified Eagle's medium (DMEM),
minimum essential medium (MEM), fetal bovine serum (FBS) and
penicillin-streptomycin were purchased from Welgene, Inc. Apigenin
(Fig. 1), MTT cell lysis buffer,
DAPI and dimethyl sulfoxide (DMSO) were purchased from
Sigma-Aldrich (Merck KGaA). Fluoresceinisothiocyanate (FITC)
Annexin V Apoptosis Detection kit was obtained from BD Pharmingen
(BD Biosciences). Anti-β-actin (cat. no. 4967), anti-Bax (cat. no.
2772), anti-Bcl-2 (cat. no. 2876), anti-cleaved caspase-9 (cat. no.
9501), anti-caspase-9 (cat. no. 9502), anti-poly ADP-ribose
polymerase (PARP; cat. no. 9542), anti-cleaved PARP (cat. no.
5625), anti-p53 (cat. no. 2527), anti-phosphorylated (p)-p38 (cat.
no. 4631), anti-p38 (cat. no. 9212), anti-p-JNK (cat. no. 4668),
anti-JNK (cat. no. 9252), anti-p-ERK (cat. no. 4376), anti-ERK
(cat. no. 4695), anti-p-Akt (cat. no. 4060), anti-Akt (cat. no.
9272) and goat anti-rabbit IgG (cat. no. 7074) antibodies were
obtained from Cell Signaling Technology, Inc.
Cell line and culture
Melanoma cells were maintained in MEM and DMEM
supplemented with 5% FBS and 1% penicillin/streptomycin under
standard culture conditions at 37°C in a humidified atmosphere of
95% air and 5% CO2. The culture medium was replaced
every 2–3 days. For apigenin treatment, melanoma cells were seeded
into a 175 cm2 flask (Nalge Nunc International; Thermo
Fisher Scientific, Inc.) when density reached ~80–90% and allowed
to attach overnight.
Cell viability assay
The anticancer effects of apigenin were assessed
using an MTT assay. A375P and A375SM cells were seeded onto a
96-well plate at a density of 2×104 cells/ml and a
volume of 200 µl/well. Following incubation for 24 h, cells were
treated with apigenin (0, 25, 50, 75 and 100 µM) for 24 h in
triplicate. After apigenin treatment, the medium was discarded and
a total of 40 µl MTT solution (5 mg/ml) was added to each well
followed by incubation for an additional 2 h. Subsequently, the
medium was aspirated and the formazan product generated by viable
cells was solubilized with the addition of 100 µl DMSO. The
absorbance of the solution was determined at 595 nm using a
microplate reader (Bio-Rad Laboratories, Inc.). The percentage of
viable cells in the apigenin treatment group was estimated in
comparison with the untreated control cells.
Wound healing migration assay
A375P and A375SM cells were seeded in 60-mm culture
dishes at a density of 1×105 cells/ml and allowed to
grow for 24 h. A uniform wound was introduced by scraping the
monolayers with a sterile blue-pipette tip. Subsequently, the
culture medium was replaced with non-FBS fresh medium supplemented
with increasing concentrations of apigenin (0, 50 and 100 µM) and
cells were cultured for an additional 24 h. The rate of wound
closure in apigenin-treated and untreated cells was monitored by
images captured with a phase-contrast microscope (magnification,
×100) immediately after wound incision (0 h) and following 24 h.
The percentage of the migrated cells was estimated compared with
the cell density in the wound of the untreated control group.
DAPI staining
Apoptotic cell death was determined by observing
morphological changes using DAPI fluorescent nuclear dye. DAPI
stains cells undergoing apoptosis characterized by chromatin
condensation and nuclear fragmentation. A375P and A375SM cells were
treated with PBS or various concentrations of apigenin (0, 50 and
100 µM) for 24 h, harvested by trypsinization and fixed in 70%
ethanol overnight at 4°C. The following day, cells were stained
with DAPI at room temperature for 1 min, deposited onto slides and
observed under a fluorescence microscope (magnification, ×200) to
detect characteristics of apoptosis. The apoptotic rate was
measured using the following equation: Number of positive
cells/total number of cells in three random fields from each
sample.
Annexin V-propidium iodide (PI)
staining
Apoptosis rate was also determined using a
FITC-Annexin V Apoptosis Detection kit (BD Pharmingen; BD
Biosciences). For the Annexin V-PI staining, A375P and A375SM human
melanoma cells were treated with 0, 50 and 100 µM apigenin for 24
h. Following treatment, cells were washed with PBS, suspended in
trypsin-EDTA solution and centrifuged (200 × g) at 4°C for 5 min to
obtain cell pellet. The harvested cells were diluted in 1X binding
buffer at a density of 1×106 cells/ml. Subsequently,
cells were treated with 5 µl/100 µl FITC-conjugated Annexin V and
phycoerythrin-conjugated PI for 15 min and apoptosis was measured
by flow cytometry (BD FACSVerse™ Flow cytometer, BD Life Sciences).
BD FACSuite (v1.0.6; BD Life Sciences) was used for analysis. The
early and late apoptosis rate was indicated by the percentage of
Annexin V positive cells.
Western blot analysis
Cells were treated with various concentrations of
apigenin (0, 50 and 100 µM) for 24 h, total proteins were extracted
using PRO-PREP protein extraction solution (Intron Biotechnology,
Inc., cat. no. 17081), and then the protein concentration was
determined using a Bradford protein assay (Bio-Rad Laboratories,
Inc.). Total proteins (30 µg per lane) in each cell lysate were
resolved via SDS-PAGE on 6–14% gels, and subsequently
electrotransferred onto nitrocellulose membranes. Following
blocking with 5% non-fat dry milk in Tris-buffered saline with 0.5%
Tween-20 (TBST) buffer for 1 h at room temperature, membranes were
incubated with specific primary antibodies diluted in blocking
solution at 4°C overnight. After washing with TBST, membranes were
incubated with horseradish peroxidase (HRP)-conjugated secondary
antibodies for 1 h at room temperature. Following washing, bands
were visualized using an enhanced chemiluminescence detection
reagent (Pierce; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. The following antibodies were used:
β-actin (1:1,000), Bax (1:1,000), Bcl-2 (1:1,000), caspase-9
(1:1,000), cleaved-caspase-9 (1:1,000), PARP (1:1,000),
cleaved-PARP (1:1,000), p38 (1:1,000), p-p38 (1:1,000), JNK
(1:1,000), p-JNK (1:1,000), ERK (1:1,000), p-ERK (1:1,000), Akt
(1:1,000) and p-Akt (1:1,000) as primary antibodies and rabbit IgG
(1:1,000) as the secondary antibody. Protein intensity was
semi-quantified by the ImageJ software (National Institutes of
Health, v1.8.0).
Animals and in vivo xenograft tumor
model
A total of 15 male BALB/c nude (nu/nu) mice (age, 5
weeks, body weight, 17–19 g) were purchased from Orient Bio Inc.
Animal experiments were performed in accordance with the Guidelines
for the Kongju National University Institutional Animal Care and
Use Committee (Chungcheongnam, Korea) and approved by the Ethics
Committee of Kongju National University (approval no. KNU_2018-5).
Equipment was provided by the Laboratory Animal Resource Center
(Gwangju, Korea). Mice were maintained under a 12 h light/dark
cycle and housed under a controlled temperature of 23±3°C and
humidity of 40±10%. Mice were allowed ad libitum access to
laboratory pellet food and water. A375SM cells at 80–90% density
were maintained in DMEM and MEM supplemented with 10% FBS and 1%
penicillin-streptomycin at 37°C in a humidified atmosphere of 5%
CO2. A375SM cells were harvested from cultures using
0.25% trypsin. Trypsinization was stopped using a solution
containing 10% FBS, cells were then rinsed twice and resuspended in
DMEM and MEM. Subsequently, a total of 2×107 cells in
0.2 ml culture medium were injected subcutaneously into the right
flank of donor nude mice. On day 7 following injection, A375SM
cells growing under the skin of nude mice developed tumors. When
the tumors became palpable, mice were assigned randomly into three
groups (n=5), namely the vehicle-treated control group and the
apigenin-treated groups (25 or 50 mg/kg body weight). Apigenin was
orally administrated five times/week for 3 weeks at a dose of 25 or
50 mg/kg body weight, while control mice were treated with the
vehicle only. Oral administration was performed using an oral zonde
needle. Animal health and behavior were monitored daily. Body
weight and tumor volume were monitored twice weekly. The tumor
volumes were calculated using the following equation: Tumor volume
(mm3)=0.5 × length × width2. Then, three
weeks after the start of apigenin injection, the final tumor size
was measured. All mice were sacrificed using CO2 gas
(30% per min, 3 min) and tumors were excised to measure tumor
weight. A section of the tumor tissue was embedded in paraffin and
fixed with 10% formalin at room temperature for 12 h was
subsequently used for TUNEL and immunohistochemistry (IHC) assays.
The criteria used to determine when an animal should be euthanized
were set as follows: i) Mice showed a weight loss of ≥20% of its
normal weight; ii) tumor grew to ≥10% of its normal weight; iii)
mice developed ulcers or infections in the tumor area; or iv)
erosion of surrounding tissues.
TUNEL assay
TUNEL staining was performed in paraffin-embedded
5-µm-thick tumor sections using the DeadEnd™ Colorimetric TUNEL
System (Promega Corporation), according to the manufacturer's
protocol. Briefly, sections were deparaffinized in xylene,
dehydrated via a series of graded alcohol rinses (100, 95, 85, 70
and 50% ethanol (v/v) in double-distilled H2O) and
rehydrated in PBS (pH 7.5). Subsequently, the tissue samples were
permeabilized with a proteinase K solution following refixing in 4%
paraformaldehyde solution at room temperature for 15 min. Slides
were treated with the rTdT reaction mix and incubated at 37°C for 1
h. Reactions were terminated by immersing the slides in 2X SSC
solution for 15 min at room temperature. Following blocking of
endogenous peroxidase activity with 0.3% hydrogen peroxide, slides
were washed with PBS, and then incubated with streptavidin HRP
solution for 30 min at room temperature. After washing, slides were
incubated with a 3,3-diaminobenzidine (DAB; substrate) solution
until a light brown background appeared (10 min) and rinsed several
times in deionized water. Following mounting, slides were observed
under a light microscope. The number of positive cells in three
random fields from each sample was counted indicating the number of
apoptotic cells.
Immunohistochemistry
The paraffin-embedded sections were deparaffinized
and dehydrated by sequential immersion in xylene and graded alcohol
solutions, respectively. Sections were blocked using 1X Animal-Free
Blocking Solution (Cell Signaling Technology, Inc., cat. no. 15019)
at room temperature for 1 h. The sections were incubated with an
antibody against p-ERK (1:100) at 4°C overnight, and subsequently
incubated with a HRP-conjugated goat anti-rabbit antibody for 1 h
at room temperature. The tumor sections were visualized using a DAB
solution, treated with mounting reagent and observed under a
routines light microscope (magnification, ×200). Finally, p-ERK
positive cells were counted in three random fields from each
sample.
Histological examination
The excised liver and kidney specimens were
immediately fixed in 10% neutral buffered formalin at room
temperature for 72 h, embedded in paraffin and cut into 5-µm-thick
sections. Following hematoxylin and eosin (H&E) staining at
room temperature (hematoxylin for 5 min, eosin for 1 min), the
sections were examined under a light microscope (magnification,
×200).
Statistical analysis
Results are presented as the mean ± standard error
of the mean for tumor volume, tumor weight, and body weight in
vivo, while others are presented as the mean ± standard
deviation. Differences in the mean values between control and
apigenin-treated groups were assessed by one-way ANOVA followed by
a Dunnett's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Apigenin inhibits the survival of
melanoma cells
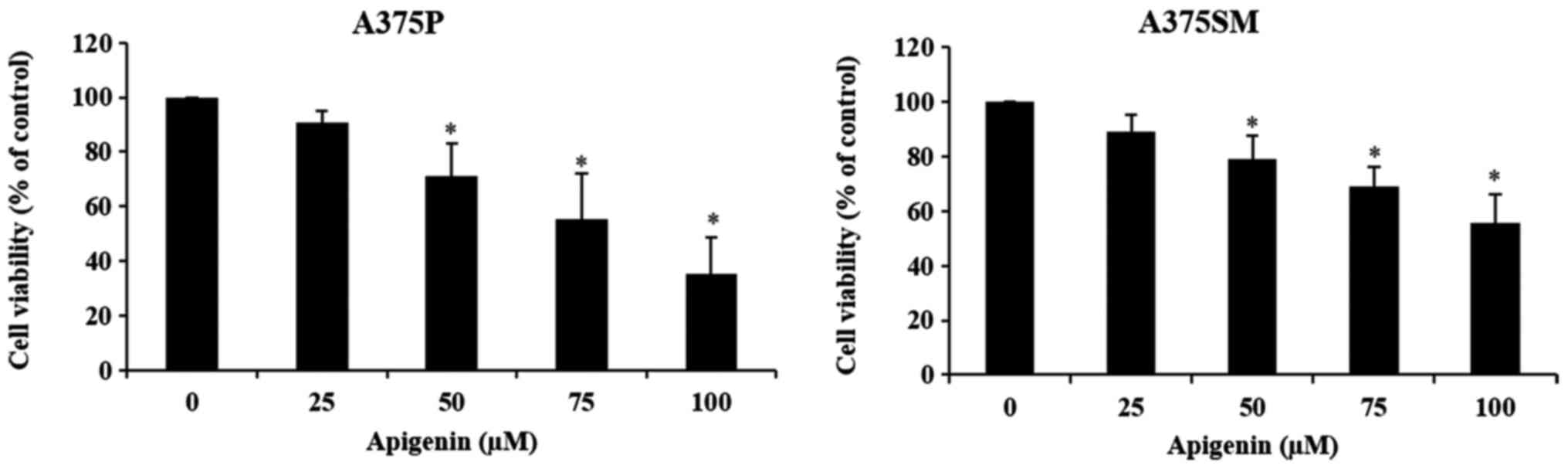
To investigate the effects of apigenin on A375P and
A375SM cell viability, cells were treated with different
concentrations (0, 25, 50, 75 and 100 µM) of apigenin and viability
was assessed using an MTT assay. When A375P and A375SM cells were
treated with 0, 25, 50, 75 and 100 µM of apigenin for 24 h, the
cell survival rates were 100, 90, 71, 55 and 35%, and 100, 89, 78,
69 and 55% for A375P and A375SM cells, respectively (Fig. 2). The decrease in cell viability
was dose-dependent, as it decreased with increasing concentrations
of apigenin. Significant effects were observed at concentrations
≥50 µM.
Apigenin affects the migratory ability
of melanoma cells
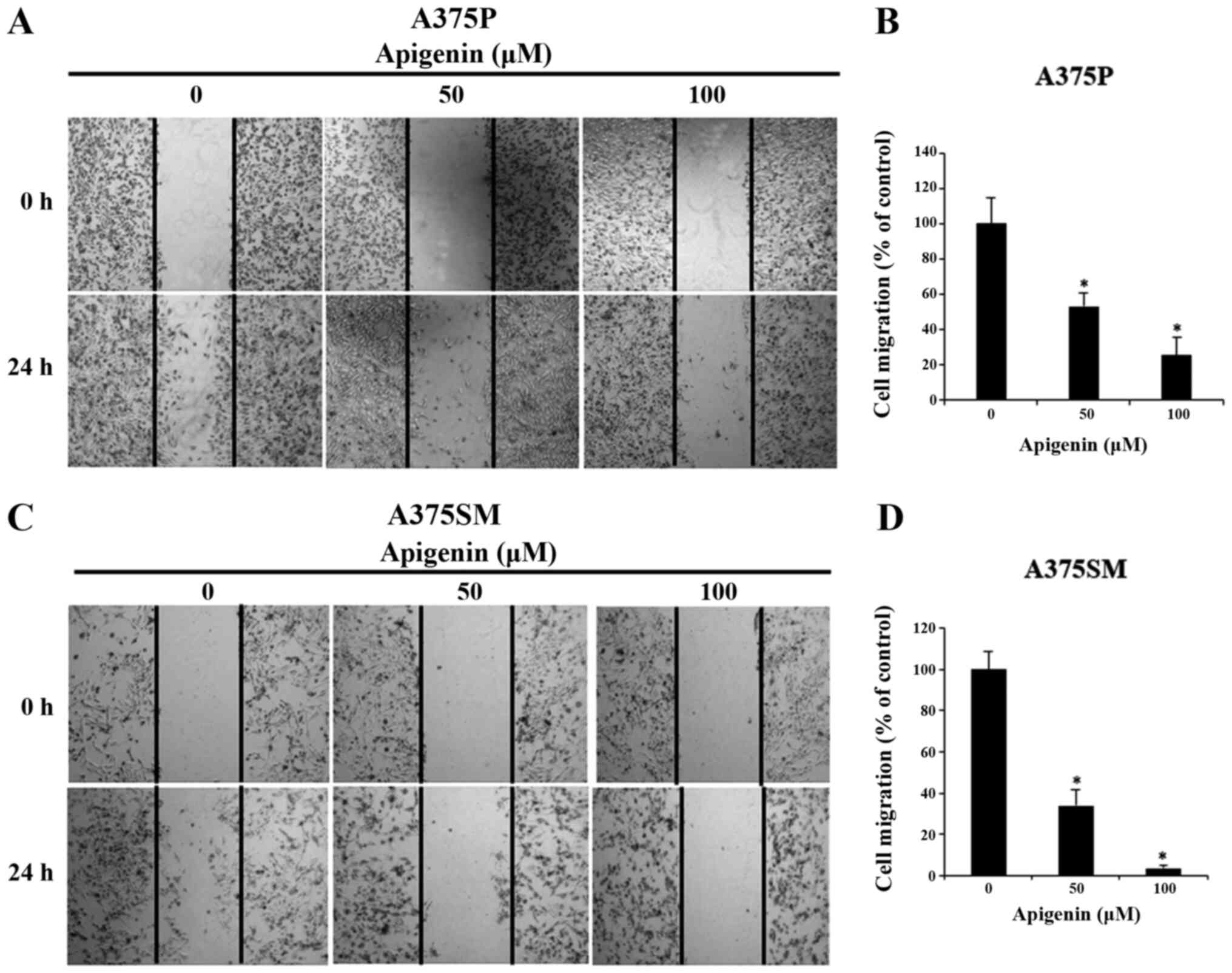
Subsequently, to explore the effect on apigenin on
A375P and A375SM cell migration, melanoma cells were treated with
various concentrations of apigenin (0, 50 and 100 µM) that
significantly affected the viability of A375P and A375SM cells as
demonstated by the MTT assay. The results revealed that cell
migration was attenuated in a dose-dependent manner compared with
that noted in the control group (Fig.
3A and C). The migration rates in A375P and A375SM cells
treated with 0, 50 and 100 µM apigenin were 100, 53 and 25%, and
100, 34 and 4%, respectively (Fig. 3B
and D).
Effect of apigenin on morphological
changes in melanoma cells
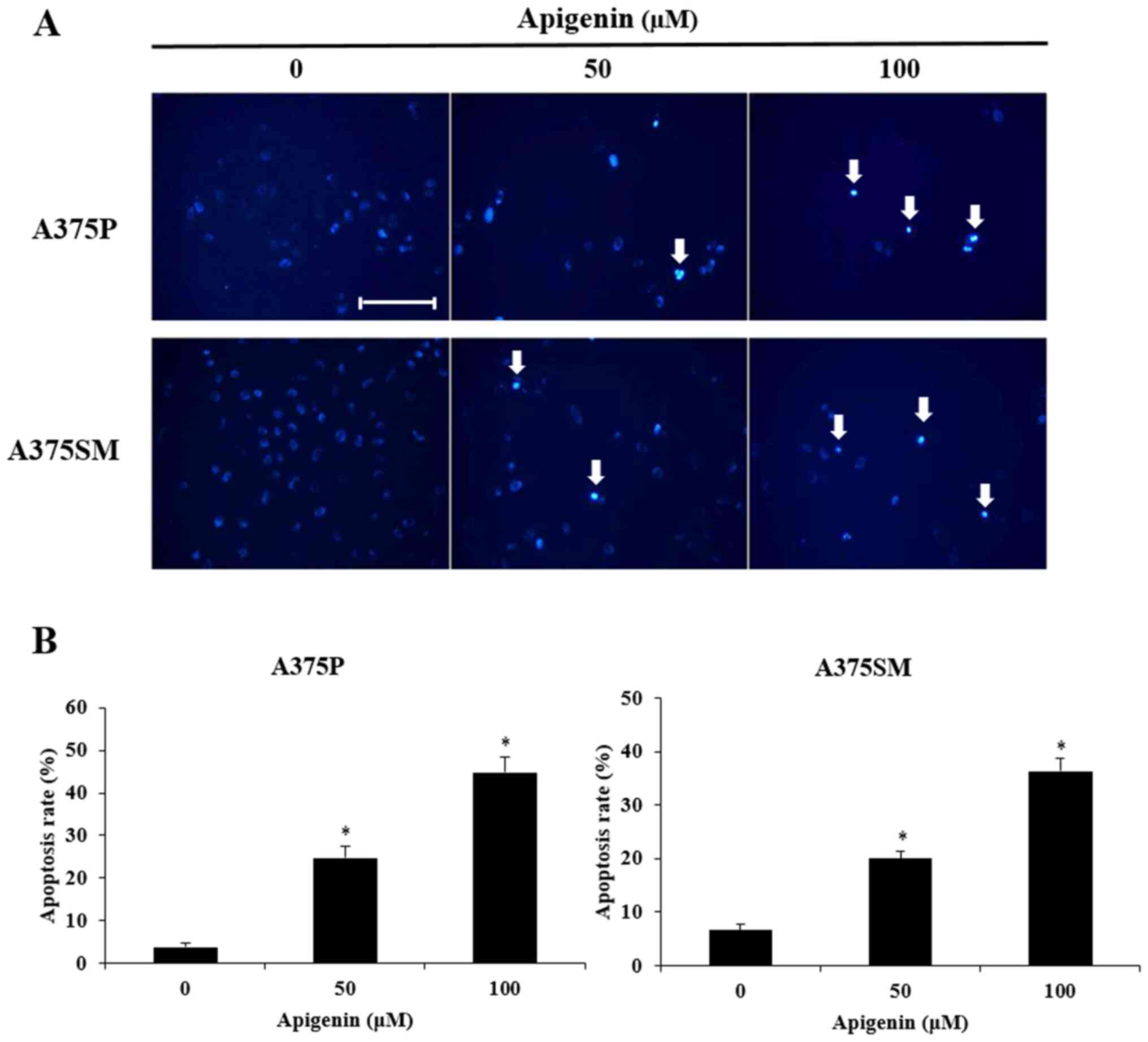
To determine whether the decreased cell viability
observed in apigenin-treated A375P and A375SM cells was mediated by
apoptosis, the morphological changes and chromatin condensation in
the nucleus of apigenin-treated cells were observed following DAPI
staining. Therefore, A375P and A375SM cells were treated with 0, 50
and 100 µM of apigenin for 24 h, co-treated with DAPI staining and
observed under a fluorescence microscope (Fig. 4A). Consistent with the MTT assays,
the results revealed that apigenin exerted inhibitory effects on
cancer cell growth by decreasing total cell count. The inhibitory
effect of apigenin on cancer cell growth was mediated by apoptosis
as confirmed by the presense of morphological features of
apoptosis, such as apoptotic bodies and chromatin condensation. To
quantitatively analyze apoptosis, DAPI-positive cells were counted
(Fig. 4B) and the morphology of
the apoptotic bodies was observed under a fluorescence microscope.
The apoptosis rates of A375P and A375SM cells treated with 0, 50
and 100 µM apigenin were 3.8, 24.8 and 44.8%, and 6.6, 20.0 and
36.4%, respectively. Thus, this indicated that apigenin increased
apoptosis in a dose-dependent manner.
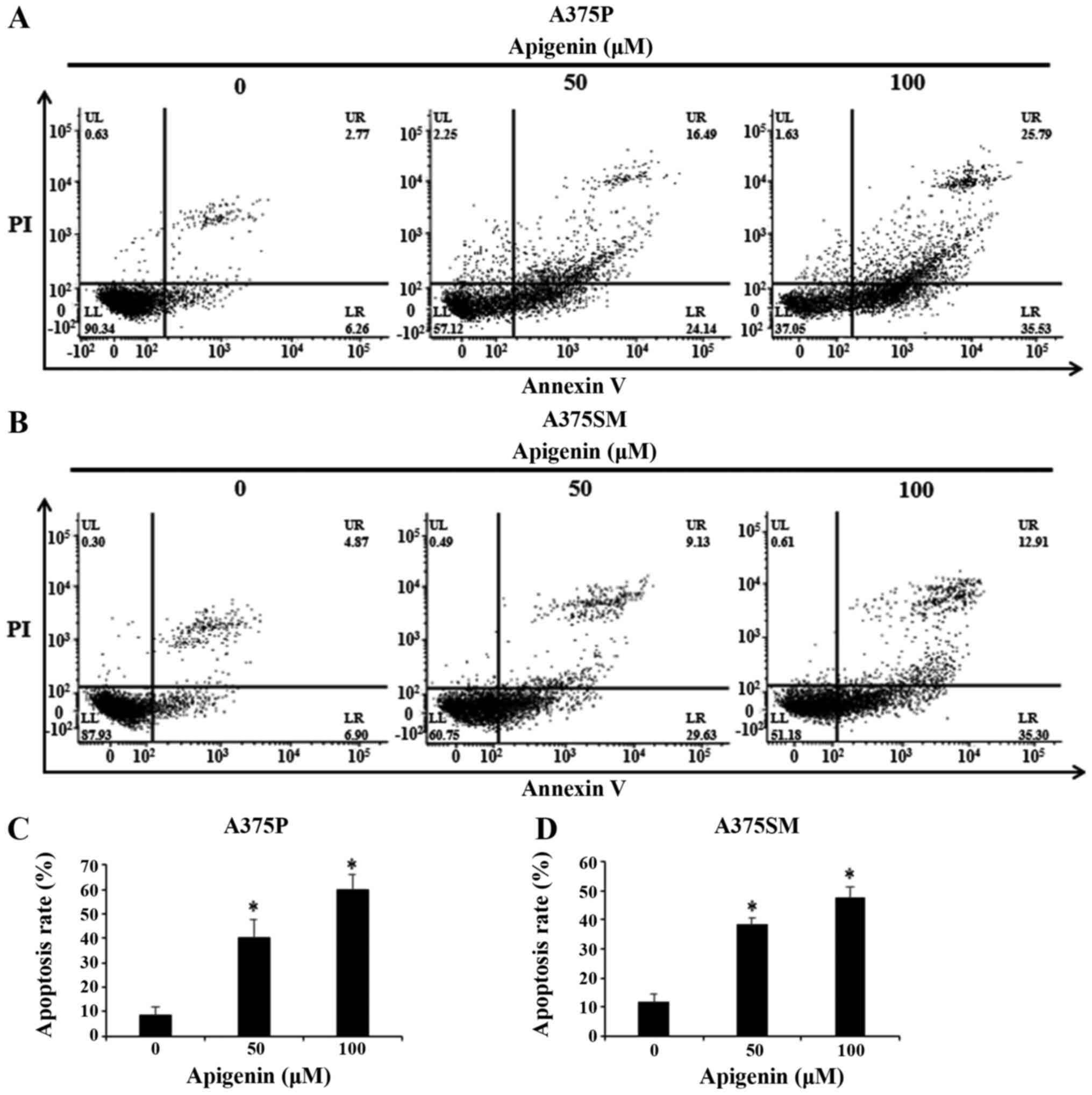
Effect of apigenin on apoptosis in
melanoma cells
To investigate whether the formation of apoptotic
bodies observed with DAPI staining was mediated by apoptosis,
Annexin V-PI staining was used. Therefore, A375P and A375SM cells
were treated with 0, 50 and 100 µM apigenin for 24 h and apoptosis
was confirmed by flow cytometry (Fig.
5A and B). The results showed that the apoptosis rates
(percentage of Annexin V positive cells) of apigenin-treated
melanoma cells increased in a dose-dependent manner. As shown in
Fig. 5C and D, apoptosis rates of
A375P and A375SM cells treated with 0, 50 and 100 µM apigenin were
8.7, 40.3 and 59.6%, and 11.6, 38.5 and 47.5%, respectively.
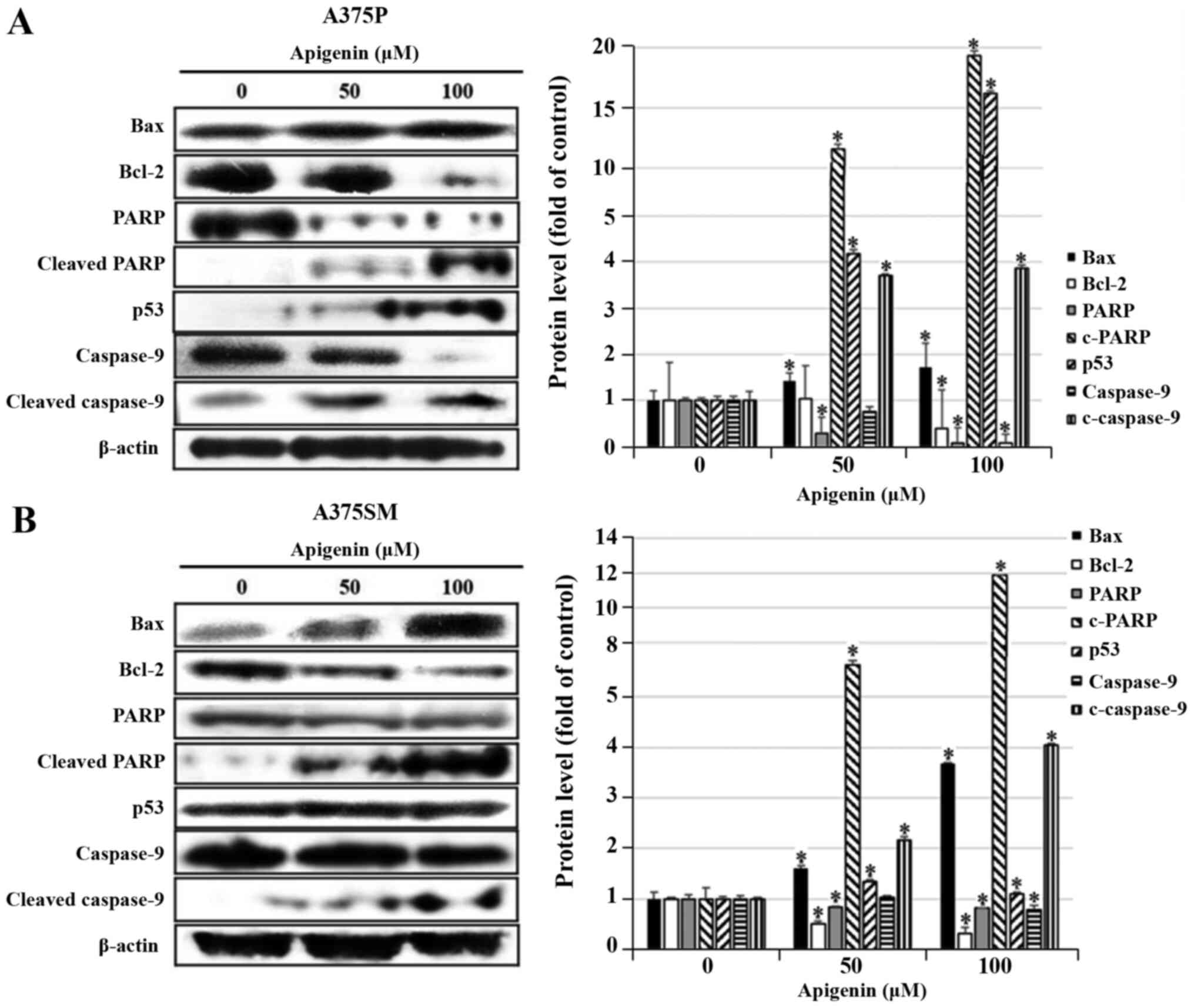
Effect of apigenin on
apoptosis-related protein expression in melanoma cells
DAPI and Annexin V-PI staining showed that apoptosis
was significantly increased in A375P and A375SM cell lines treated
with 50 and 100 µM of apigenin. Therefore, the expression levels of
proteins that regulate apoptosis were measured using western blot
analysis. To detect the expression levels of Bax, Bcl-2, PARP, p53
and caspase-9, which are known to regulate apoptosis, A375P and
A375SM cells were treated with apigenin (0, 50 and 100 µM), and
subsequently western blotting was performed. As the concentration
of apigenin increased, the expression levels of PARP and caspase-9
decreased, and the expression levels of pro-apoptotic proteins Bax,
p53, cleaved PARP and cleaved caspase-9 significantly increased,
whereas the expression of Bcl-2, known to inhibit apoptosis, was
downregulated (Fig. 6A and B).
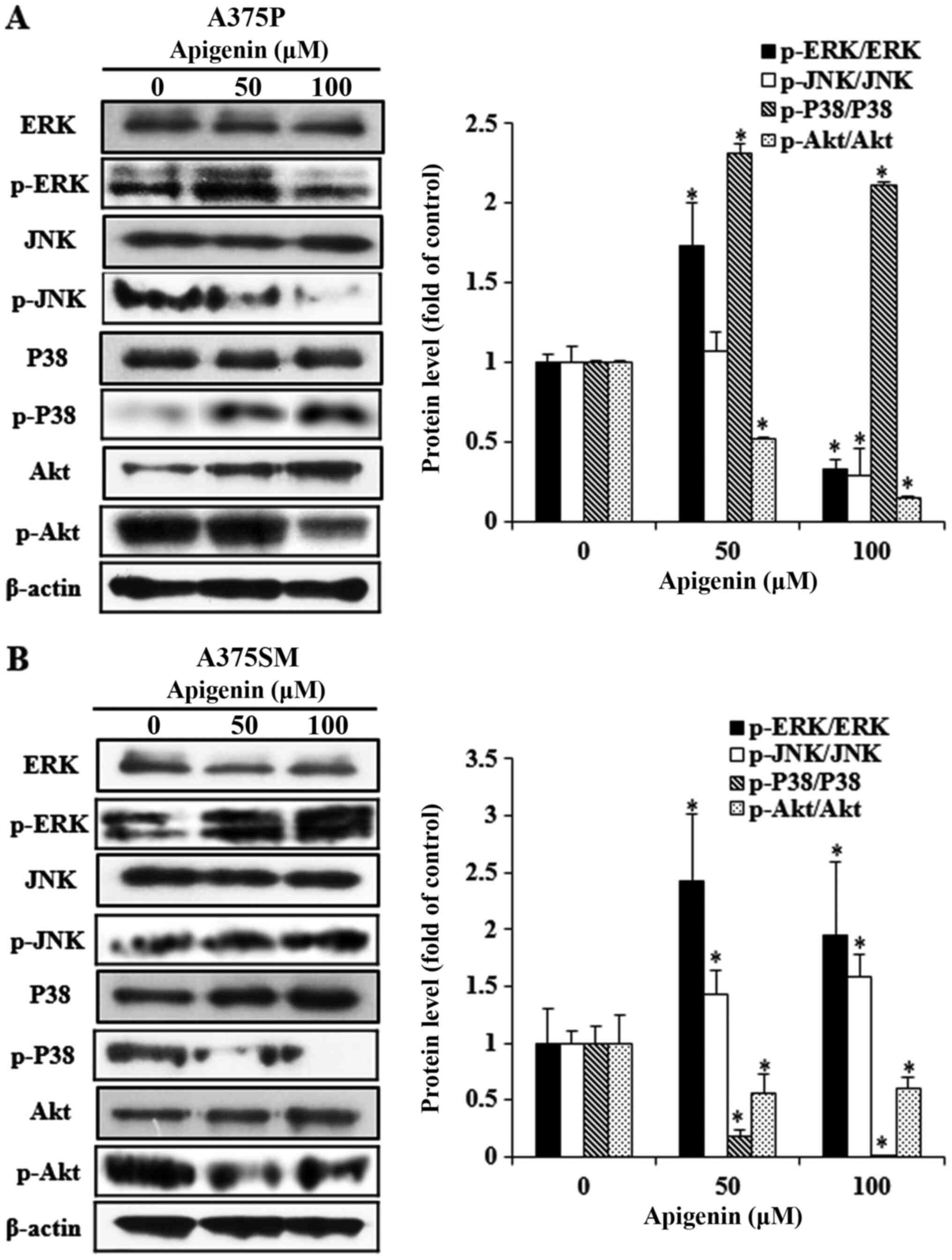
Effect of apigenin on the MAPK and Akt
signaling pathways in melanoma cells
The phosphorylation status of specific proteins in
cancer cells is widely recognized as an important process in
determining cell fate, including apoptosis. Various kinases of the
MAPK pathway, an intracellular signaling system, are involved in
intercellular and intracellular reactions in response to changes in
the intracellular environment (34). ERK, JNK and p38, which belong to
the MAPK pathway, are known to regulate several biological
functions including cell proliferation, differentiation and
apoptosis. Furthermore, Akt plays a central role in cell growth and
proliferation, and is active in the majority of tumor tissues
(35,36). However, Akt expression
significantly differs in response to different active cellular
materials, biological functions and environment. Therefore, western
blot analysis was performed to reveal the effects of apigenin (0,
50 and 100 µM) on the protein expression of members of the Akt and
MAPK pathways in A375P and A375SM cells (Fig. 7A and B). The expression levels of
the MAPK pathway-related proteins, p-JNK and p-p38, significantly
decreased and increased, respectively, in A375P cells. Furthermore,
expression of p-ERK and p-Akt decreased following treatment with
100 µM apigenin. However, the increase in p-p38 expression was not
associated with the concentration of apigenin and p-ERK was not
downregulated in a dose-dependent manner. In A375SM cells, the
expression levels of p-JNK and p-ERK in the MAPK pathway increased,
whereas those of p-p38 and p-Akt decreased, depending on the
concentration of apigenin. However, the increase in p-ERK
expression was not associated with the concentration of apigenin.
The aforementioned results indicated that proteins in the signal
transduction pathways were differentially expressed in the two
melanoma cell lines.
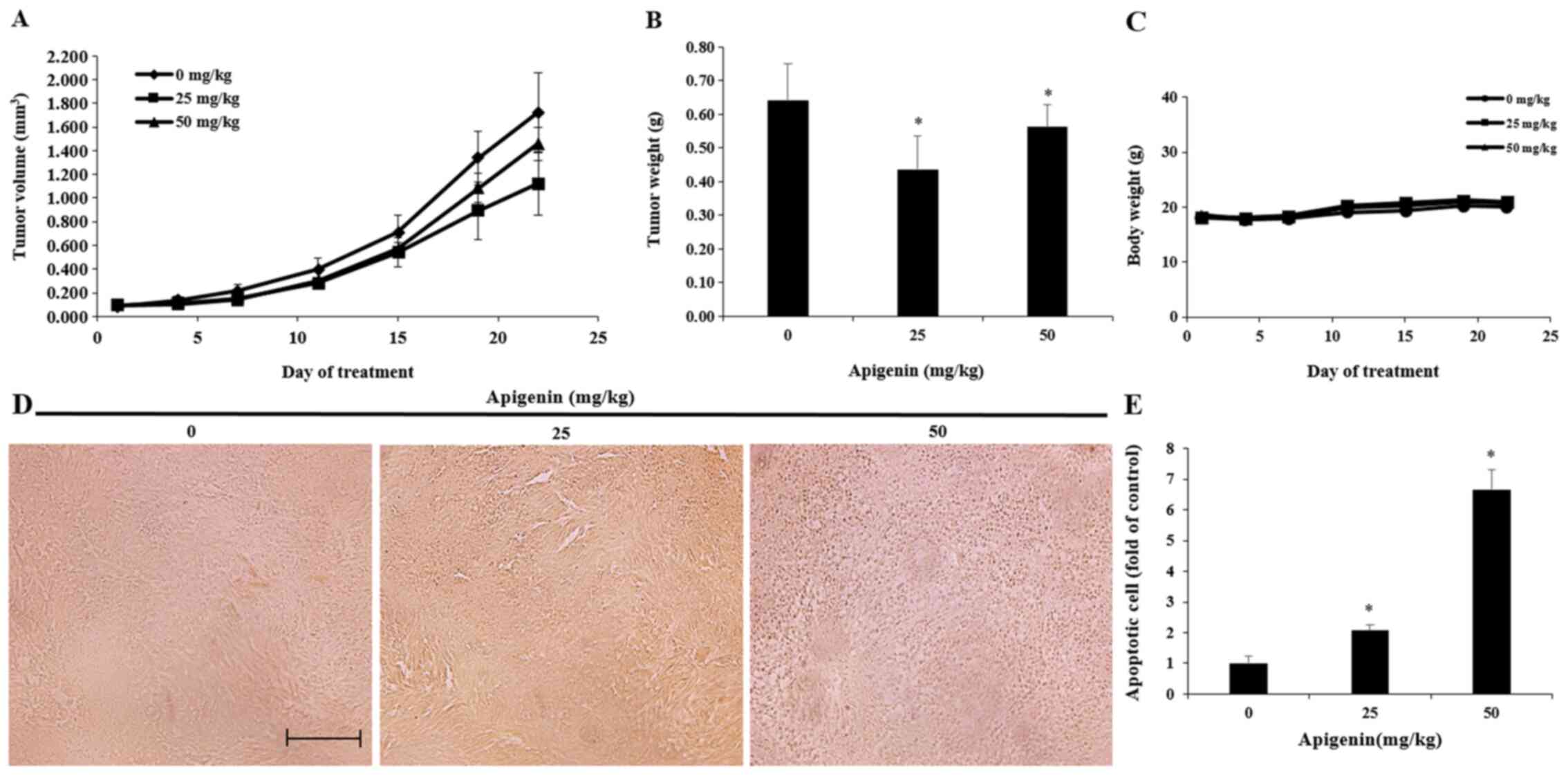
Effect of apigenin on tumor growth in
an in vivo animal model
Based on the in vitro experiments, a A375SM
cell line-derived xenograft mouse model was established. A375SM
melanoma cells were transplanted into nude mice and the effect of
apigenin on tumor growth was investigated. Apigenin was diluted in
PBS and orally administered at concentrations of 25 and 50 mg/kg
five times per week for 3 weeks. Tumor size was measured twice per
week. Tumor size was decreased in the apigenin-treated group after
two weeks of treatment compared with that observed in the control
group (Fig. 8A). The tumor
inhibition rate was 35.0% on the 21st day post-administration in
the low-concentration group (25 mg/kg) and 15.4% in the
high-concentration group (50 mg/kg), as shown in Table I. Tumor weights were 0.64±0.1,
0.46±0.1 and 0.57±0.07 g in the control group, low-concentration
group and high-concentration group, respectively. Therefore, tumor
weight was decreased in both groups following treatment with
apigenin compared with the control group (Fig. 8B). However, there were no
statistically significant changes in body weight following apigenin
administration (Fig. 8C).
 | Table I.Tumor inhibtion rate in mice
implanted with A375SM melanoma cells treated with apigenin. |
Table I.
Tumor inhibtion rate in mice
implanted with A375SM melanoma cells treated with apigenin.
| Apigenin
(mg/kg) | Pre-experiment
size, mm3 | Post-experiment
size, mm3 | Inhibition
rateb, % |
|---|
| 0a | 83.3 | 1,724.6 |
|
| 25 | 93.6 | 1,120.8 | 35.0 |
| 50 | 95.1 | 1,458.3 | 15.4 |
Effect of apigenin on melanoma tumor
cell apoptosis in vivo
A TUNEL assay was performed to determine whether the
inhibitory effect of apigenin on the growth of A375SM melanoma
cells isolated from a melanoma xenograft model was mediated by
apoptosis. The results revealed that the number of TUNEL-positive
cells was increased in the apigenin-treated group compared with the
control group (Fig. 8D and E).
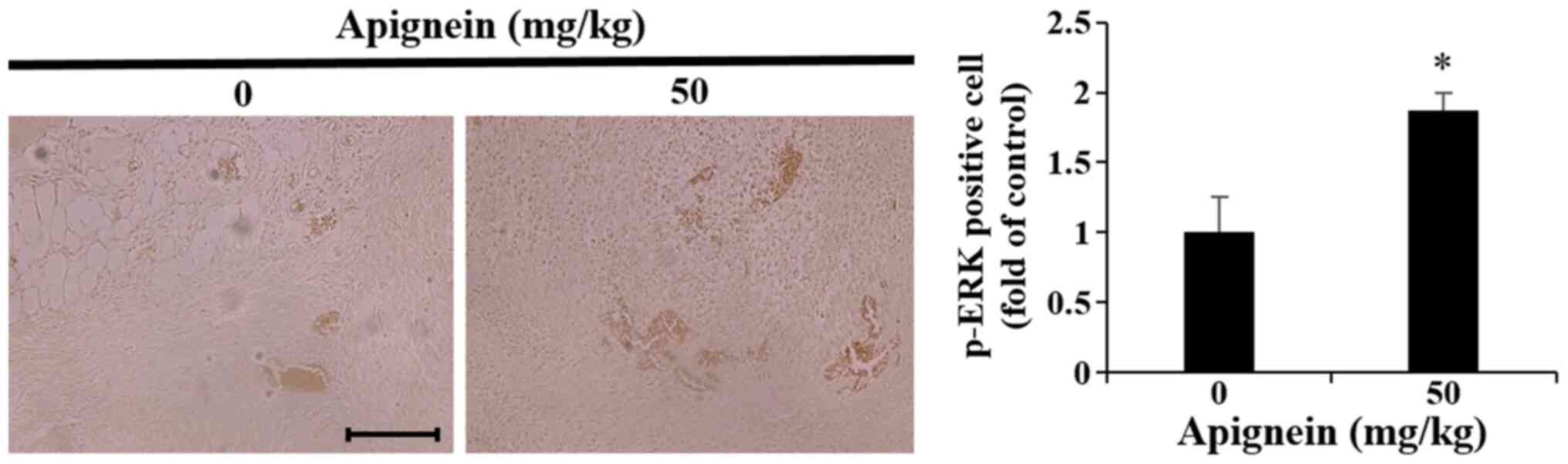
Effect of apigenin on p-ERK expression
in melanoma tumor tissue
The in vitro experiments demonstrated that
p-ERK was upregulated in apigenin-treated melanoma cells.
Therefore, melanoma tumor samples were isolated from the melanoma
mouse xenograft model 3 weeks following apigenin administration,
and IHC revealed an increase in p-ERK expression compared with the
control group (Fig. 9).
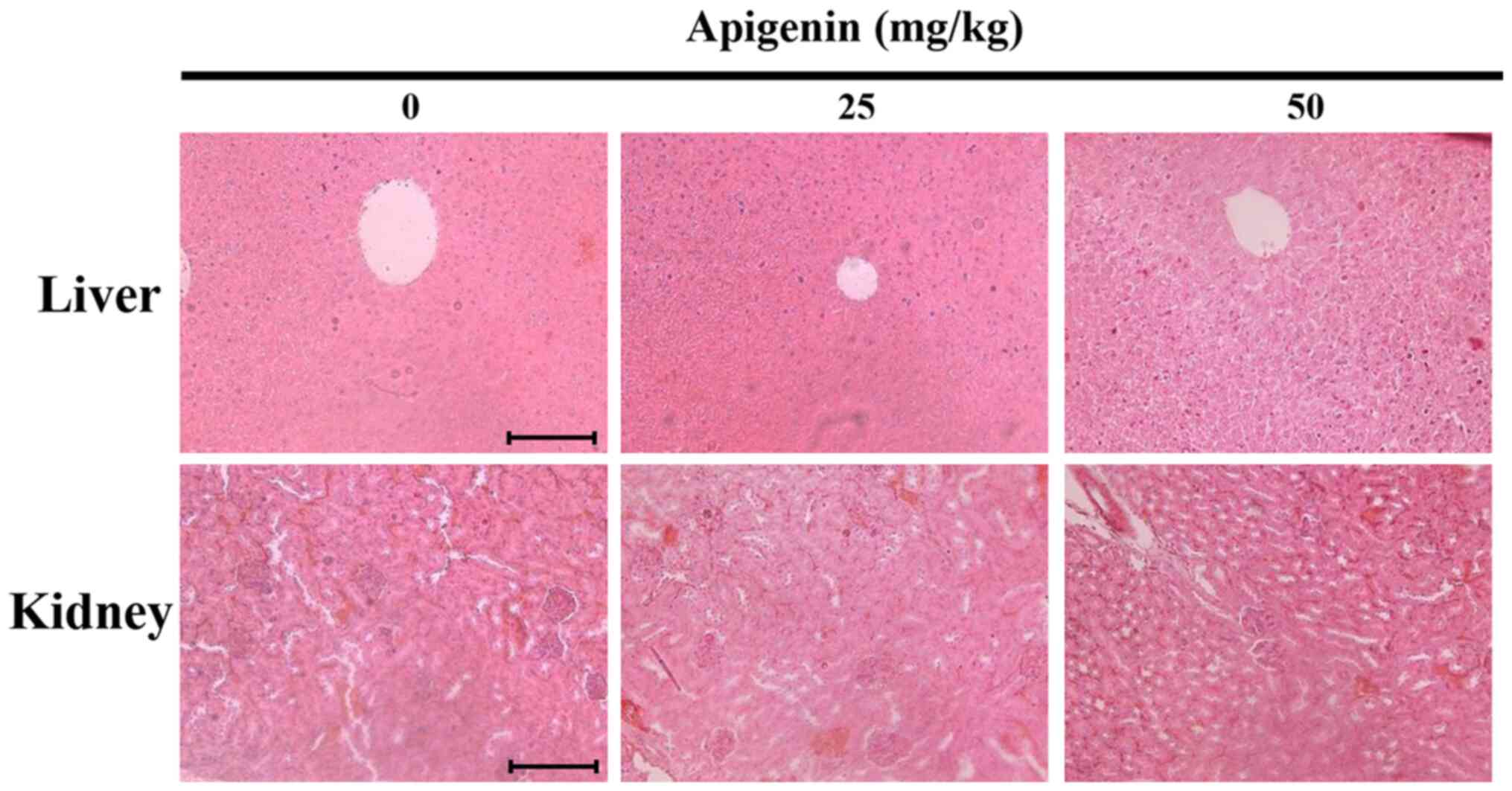
Apigenin induces histopathological
changes in melanoma tumor tissues
Subsequently, to assess apigenin-induced organ
toxicity, liver and kidney tissues derived from tumor-xenografted
mice were histologically examined by H&E staining under a light
microscope (Fig. 10). However, no
histopathological abnormalities were observed, indicating that
apigenin had no detectable toxic effects.
Discussion
The present study suggested a mechanism for the
inhibitory effect of apigenin on A375P and A375SM melanoma cell
growth and proliferation. The two melanoma cell lines differ in
their invasive and metastatic properties, with A375P cells
exhibiting decreased properties compared with A375SM cells
(37). Cancer cells induce
tumorigenesis via invasion to healthy tissues from blood and
lymphatic ducts after their proliferation into the adjacent tissues
(38). To investigate the
inhibitory effects of apigenin in vitro, A375P and A375SM
cells were treated with different concentrations of apigenin, and
MTT and wound healing assays were performed. The results of the MTT
assay showed reduced cell viability of A375P and A375SM melanoma
cells treated with apigenin in a dose-dependent manner. These
results were consistent with previous studies, indicating that
apigenin decreased tumor growth in KB oral cavity cancer cells
(39), HCT-16 colorectal cancer
cells (22) and HL-60 human
myeloma cancer cells (40) in a
dose-dependent manner. The wound healing assay in the present study
demonstrated that apigenin attenuated the migratory ability of
melanoma cells. Previous studies revealed that apigenin inhibits
cell migration of CD44+ prostate cancer cells (41), and DLD1 and SW480 colorectal cancer
cells (42). These previous
findings were consistent with the results from the present study,
demonstrating the inhibitory effects of apigenin on A375P and
A375SM melanoma cell migration when applied at different
concentrations.
Apoptosis may occur in response to normal cell
injury or pathological factors (28) and is accompanied by various
features, such as cytoplasmic or chromatin condensation, cell
membrane alteration and DNA fragmentation. These features play
important roles in biological generation and maintenance of
homeostasis. Therefore, DAPI staining was performed to clarify
whether the inhibitory effect of apigenin on melanoma cell growth
and proliferation was mediated by apoptosis. Cancer cell growth and
count were decreased following apigenin treatment. In addition,
characteristics of apoptosis were observed, including chromatin
condensation and the presence of apoptotic bodies. A previous study
also demonstrated that apigenin induces apoptosis in KB oral cavity
cancer cells in a dose-dependent manner (43). Apoptosis was triggered by DNA
fragmentation and subsequent apoptosis in the nucleosome of A375P
and A375SM melanoma cells. DAPI staining showed a significant
increase in apoptotic cell count. Annexin V is a representative
staining agent for apoptosis. Annexin V positive cells are early
and late apoptotic cells, and Annexin V negative PI positive cells
are necrotic cells. Apoptosis rate was measured by Annexin V-PI
staining and supported the results of DAPI staining. Therefore,
these findings suggested that apigenin could induce apoptosis in
A375P and A375SM melanoma cells.
It has been established that apoptosis inhibits
tumor development and progression. Pro-apoptotic proteins, such as
Bax, Bid and Bad, promote apoptosis by rupturing the outer
mitochondrial membrane, whereas anti-apoptotic proteins, including
Bcl-2, Bcl-xL and A1, inhibit apoptosis by preserving the ruprure
of the outer mitochondrial membrane (30–32).
Akt is known to be involved in cell cycle progression and
survival-related proliferation. In addition, Akt inhibits apoptosis
via downregulating the expression of the pro-apoptotic proteins
Bcl-2 and caspase-9 via the PI3K/Akt pathway (35). Three types of MAPKs have been
identified, each with different activities, namely ERK,
stress-activated JNK and p38 kinase MAPK, that attenuate the growth
and differentiation of cancer cells (36).
In the present study, western blot analysis was
performed to detect the expression levels of the apoptosis-related
proteins. The reuslts demonstrated that the expression of Bax, p53,
cleaved PARP and cleaved caspase-9 increased in a dose-dependent
manner following treatment of A375P and A375SM cells with apigenin.
However, Bcl-2 was downregulated. It has been reported that
apigenin upregulates the expression of various proteins in
different ypes of cancer cells in a dose-dependent manner,
including p53 and Bax in SK-BR-3 breast cancer cells (44), and cleaved PARP and cleaved
caspase-9 in PC-3 cancer cells (24). Apigenin has also been demonstrated
to increase the expression of Bax, cleaved PARP and cleaved
caspase-9, and decrease that of Bcl-2 in DU-145 prostate cancer
cells in a dose-dependent manner (45). These findings suggested that
apigenin induced apoptosis via regulating the expression of the
apoptosis-related proteins, Bax, Bcl-2, PARP, p53 and caspase-9 in
A375P and A375SM melanoma cells.
Subsequently, western blot analysis was also
performed to assess the effects of apigenin on the MAPK and Akt
pathway in A375P and A375SM cells. It was demonstrated that
treatment with 100 µM apigenin led to a significant downregulation
of p-ERK, p-JNK and p-Akt expression in A375P cells, and a
downregulation of p-p38 and p-Akt expression in A375SM cells.
Results in A375SM were consistent with those observed in previous
studies demonstrating that apigenin increased the expression of
p-ERK and p-JNK and decreased that of p-p38 in PC-3 prostate
(46) and MDA-MB-231 breast cancer
cells (47). Although p-ERK has
been known to affect cell proliferation and migration in previous
studies, the cellular signaling pathways are complex and the
mechanisms of synergy or antagonism between intracellular signaling
pathways are still unclear (48–50).
Therefore, the change in p-ERK in A375P following apigenin
treatment appeared to be due to complex interactions of
intracellular signaling pathways, and further studies are needed to
clarify the role of signaling pathways in cellular processes.
Apigenin presumably inhibited the proliferation of A375P and A375SM
cells in vitro via promoting apoptosis mediated by the MAPK
and Akt signaling pathways. However, the expression levels of MAPKs
were different between the two melanoma cell lines. It has been
reported that A375P and A375SM melanoma cell lines exert different
metastatic and invasive properties (37). Therefore, consistent with previous
studies, apigenin induced apoptosis in vitro in A375SM
melanoma cells via the Akt and MAPK signal transduction proteins.
However, the protein expression of A375P under the same conditions
was consistent with that observed in previous studies showing that
apigenin decreased expression of p-ERK and p-AKT in A375, C8161
human melanoma cells (51), U266,
RPMI8226 human multiple myeloma cells (52) and decreased expression of p-Akt in
MDA-MB-231 human breast cancer cell (53). The aforementioned findings
suggested that the different effects of apigenin on A375P and
A375SM cells should be further investigated.
Based on the in vitro results, A375SM cells
were transplanted into nude mice, which were in turn treated with
apigenin to determine its effect on tumor growth. Tumor size and
weight decreased in a dose-dependent manner in the apigenin-treated
groups compared with the control group. Previous studies also
demonstrated that intraperitoneally administrated apigenin at a
concentration of 50 and 5.25 mg/kg, attenuated tumor growth of MIA
PaCa-2 pancreatic (54) and
MDA-MB-231 breast cancer cells (23), respectively, when transplanted into
mice compared with non-treated control mice. These results were in
agreement with those of the present study revealing the inhibitory
effect of apigenin on A375SM tumor growth.
DNA fragmentation is a hallmark of apoptosis
(55). In the present study a
TUNEL assay was performed to determine whether the inhibitory
effect of apigenin on A375SM tumor growth was mediated by
apoptosis. TUNEL-positive cells were elevated in the
apigenin-treated group compared with the control group.
Furthermore, the apoptosis rate was increased in a
concentration-dependent manner in the low-concentration and
high-concentration groups, respectively. It has been also reported
that apigenin increases apoptosis in KB oral cavity cancer cells in
a dose-dependent manner (39).
Therefore, these results indicated that apigenin inhibited the
growth of A375SM cells via inducing apoptosis.
Currently, three MAPK pathways have been identified,
namely ERK1/2, p38 MAPK and JNK/stress-activated protein kinase.
Among them, p38 and JNK pathways are activated in response to
stress-like stimuli to induce apoptosis. The ERK1/2 pathway
functions as an anti- or pro-apoptotic factor depending on cellular
characteristics (56–58). In the present study IHC was
performed to explore the effect of apigenin on p-ERK expression.
Consistent with the in vitro western blotting results, p-ERK
expression was increased in A375SM tumor tissues. It was also
previously revleaed that apigenin upregulated p-ERK expression in a
dose-dependent manner in PC-3 prostate cancer cells (46). These results indicated that
apigenin inhibited A375SM tumor growth and proliferation via
regulating p-ERK expression.
In vivo results showed that apigenin did not
induce concentration-dependent inhibition of tumor growth. However,
tumor weight was attenuated in both concentration groups compared
with the control group. Subsequent experiments demostrated that the
apigenin-treated group exhibited a significant increase in
apoptosis compared with the control group. In view of the
differences between tumor volume and weight, and the results
obtained by TUNEL assays, the aforementioned findings suggested
that tumor volume and weight were not proportional to the induction
of apoptosis. Therefore, further studies on the associasion between
tumor weight and apoptosis-related tumor suppression are
necessary.
In conclusion, the results of the in vitro
and in vivo experiments suggested that apigenin induced
apoptosis via regulating the Akt and MAPK signaling pathways, thus
inhibiting the growth and proliferation of A375SM melanoma cells.
Therefore, apigenin may be considered as an alternative therapeutic
target to modulate the Akt and MAPK pathways in A375SM melanoma
cells.
Acknowledgements
Not applicable.
Funding
This research was supported by a research grant of
the Kongju National University in 2017 and Basic Science Research
Program through the National Research Foundation of Korea (NRF)
funded by the Ministry of Education, Science and Technology (grant
no. NRF 2017R1A2B4005516).
Availability of data and materials
The datasets used and/or analysed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JSW, GSC, SDC, JHC, JSN, YSP, CSC, SKK and JYJ
conceived and designed the experiments. JSW, ESY, SHK, JHL, SHH,
HJK, SHJ and BSK performed the experiments. JSW and BKP analyzed
the data. JSW and GSC wrote the paper. JYJ supervised the entire
project. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The animal experiments were conducted in accordance
with the regulations of the Kongju National University
Institutional Animal Care and Use Committee with the approval of
the Ethics Committee of Kongju National University (approval no.
KNU_2018-5).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Doll SR: The lessons of life: Keynote
address to the nutrition and cancer conference. J Cancer Res. 52
(Suppl 7):S2024–S2029. 1992.
|
|
2
|
Jung KW, Won YJ, Kong HJ, Oh CM, Lee DH
and Lee JS: Cancer statistics in korea: Incidence, mortality,
survival, and prevalence in 2011. J Cancer Res. 46:109–123.
2014.
|
|
3
|
Jemal A, Siegel R, Ward E, Murray T, Xu J,
Smigal C and Thun MJ: Cancer statistics, 2006. CA Cancer J Clin.
56:106–130. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Megahed M, Schön M, Selimovic D and Schön
MP: Reliability of diagnosis of melanoma in situ. Lancet.
359:1921–1922. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen J, He X, Peng H and Yang XO: Research
on the antitumor effect of ginsenoside Rg3 in B16 melanoma cells.
Melanoma Res. 18:322–329. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Meier F, Satyamoorthy K, Nesbit M, Hsu MY,
Schittek B, Garbe C and Herlyn M: Molecular events in melanoma
development and progression. Front Bio Sci. 3:D1005–D1010. 1998.
View Article : Google Scholar
|
|
7
|
Changmin K and Bonglee K: Anti-cancer
natural products and their bioactive compounds inducing ER
stress-mediated apoptosis: A review. Nutrients. 10:10212018.
View Article : Google Scholar
|
|
8
|
Simone F: Modulation of apoptosis by
natural products for cancer therapy. Planta Med. 76:1075–1079.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Aung TN, Qu Z, Kortschak RD and Adelson
DL: Understanding the effectiveness of natural compound mixtures in
cancer through their molecular mode of action. Int J Mol Sci.
18:6562017. View Article : Google Scholar
|
|
10
|
Lin YJ, Liang WM, Chen CJ, Tsang H, Chiou
JS, Liu X, Cheng CF, Lin TH, Liao CC, Huang SM, et al: Network
analysis and mechanisms of action of Chinese herb-related natural
compounds in lung cancer cells. Phytomedicine. 58:1528932019.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Anna L and Krzysztof G: Anticancer
activity of natural compounds from plant and marine environment.
Int J Mol Sci. 19:35332018. View Article : Google Scholar
|
|
12
|
Perez-Vizcaino F and Fraga CG: Research
trends in flavonoids and health. Arch Biochem Biophys. 646:107–112.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fraga CG, Croft KD, Kennedy DO and
Tomás-Barberán FA: The effects of polyphenols and other bioactives
on human health. Food Funct. 10:514–528. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang M, Firrman J, Liu L and Yam K: A
review on flavonoid apigenin: Dietary intake, ADME, antimicrobial
effects, and interactions with human gut microbiota. Biomed Res
Int. 2019:70104672019.PubMed/NCBI
|
|
15
|
Dixon RA and Steele CL: Flavonoids and
isoflavonoids-a gold mine for metabolic engineering. Trends Plant
Sci. 4:394–400. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yin Y, Gong FY, Wu XX, Sun Y, Li YH, Chen
T and Xu Q: Antiinflammatory and immunosuppressive effect of
flavones isolated from artemisia vestita. J Ethnopharmacol.
120:1–6. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chuang CM, Monie A, Wu A and Hung CF:
Combination of apigenin treatment with therapeutic HPV DNA
vaccination generates enhanced therapeutic antitumor effects. J
Biomed Sci. 16:492009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Patel D, Shukla S and Gupta S: Apigenin
and cancer chemoprevention: Progress, potential and promise. Int J
Oncol. 30:233–245. 2007.PubMed/NCBI
|
|
19
|
Romanová D, Vachálková A, Cipák L, Ovesná
Z and Rauko P: Study of antioxidant effect of apigenin, luteolin
and quercetin by DNA protective method. Neoplasma. 48:104–107.
2001.PubMed/NCBI
|
|
20
|
Fuchs J and Milbradt R: Skin
anti-inflammatory activity of apigenin-7-glucoside in rats.
Arzneimittelforschung. 43:370–372. 1993.PubMed/NCBI
|
|
21
|
Zhang W, Qiao H, Lv Y, Wang J, Chen X, Hou
Y, Tan R and Li E: Apigenin inhibits enterovirus-71 infection by
disrupting viral RNA association with trans-acting factors. PLoS
One. 9:e1104292014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Moon TS, Cui LG and Yang H: Involvement of
early growth response gene 1 (EGR-1) in growth suppression of the
human colonic tumor cells by apigenin and its derivative
isovitexin. J Life Sci. 17:100–115. 2007. View Article : Google Scholar
|
|
23
|
Tseng YH, Chien MH, Lin WL, Wen YC, Chow
JM, Chen CK, Kuo TC and Lee WJ: Inhibition of MDA-MB-231 breast
cancer cell proliferation and tumor growth by apigenin through
induction of G2/M arrest and histone H3 acetylation-mediated
p21WAF1/CIP1 expression. Environ Toxicol. 32:434–444.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kaur P, Shukla S and Gupta S: Plant
flavonoid apigenin inactivates Akt to trigger apoptosis in human
prostate cancer: An in vitro and in vivo study. Carcinogenesis.
29:2210–2217. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ghițu A, Schwiebs A, Radeke HH, Avram S,
Zupko I, Bor A, Pavel IZ, Dehelean CA, Oprean C, Bojin F, et al: A
comprehensive assessment of apigenin as an antiproliferative,
proapoptotic, antiangiogenic and immunomodulatory phytocompound.
Nutrients. 11:8582019. View Article : Google Scholar
|
|
26
|
Caltagirone S, Rossi C, Poggi A,
Ranelletti FO, Natali PG, Brunetti M, Aiello FB and Piantelli M:
Flavonoids apigenin and quercetin inhibit melanoma growth and
metastatic potential. Int J Cancer. 87:595–600. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Piantelli M, Rossi C, Iezzi M, La Sorda R,
Iacobelli S, Alberti S and Natali PG: Flavonoids inhibit melanoma
lung metastasis by impairing tumor cells endothelium interactions.
J Cell Physiol. 207:23–29. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Han SI, Kim YS and Kim TH: Role of
apoptotic and necrotic cell death under physiologic conditions. BMB
Rep. 41:1–10. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Evans VG: Multiple pathways to apoptosis.
Cell Biol Int Rep. 17:461–476. 1993. View Article : Google Scholar
|
|
30
|
Adams JM and Cory S: The Bcl-2 protein
family: Arbiters of cell survival. Science. 281:1322–1326. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Danial NN and Korsmeyer SJ: Cell death:
Critical control points. Cell. 116:205–219. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Song Q, Kuang Y, Dixit VM and Vincenz C:
Boo, a novel negative regulator of cell death, interacts with Apaf
1. EMBO J. 18:167–178. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ichijo H: From receptors to
stress-activated MAP kinases. Oncogene. 18:6087–6093. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Carnero A: The PKB/AKT pathway in cancer.
Curr Pharm Des. 16:34–44. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Osaki M, Oshimura M and Ito H: PI3K-Akt
pathway: Its functions and alterations in human cancer. Apoptosis.
9:667–676. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dhillon AS, Hangan S, Rath O and Kolch W:
MAP kinase signaling pathways in cancer. Oncogene. 26:3279–3290.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ham SA, Yoo T, Hwang JS, Kang ES, Lee WJ,
Paek KS, Park C, Kim JH, Do JT, Lim DS and Seo HG: Ligand-activated
PPARδ modulates the migration and invasion of melanoma cells by
regulating Snail expression. Am. J Cancer Res. 4:674–682. 2014.
|
|
38
|
Rao KM: MAP kinase activation in
macrophages. J Leukoc Biol. 69:3–10. 2001.PubMed/NCBI
|
|
39
|
Lee JS, Seo HS, Kim SJ, Kim HJ, Kim J, Lee
SH, Park YS, Park BK, Kim BS, Kim SK and Jung JY: Studies on the
anticancer effect of apigenin in KB cell xenograft nude mouse
model. J Life Sci. 20:1519–1524. 2010. View Article : Google Scholar
|
|
40
|
Wang IK, Lin-Shiau SY and Lin JK:
Induction of apoptosis by apigenin and related flavonoids through
cytochrome c release and activation of caspase-9 and caspase-3 in
leukaemia HL-60 cells. Eur J Cancer. 35:1517–1525. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Erdogan S, Doganlar O, Doganlar ZB,
Serttas R, Turkekul K, Dibirdik I and Bilir A: The flavonoid
apigenin reduces prostate cancer CD44(+) stem cell survival and
migration through PI3K/Akt/NF-κB signaling. Life Sci. 162:77–86.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Toker A and Yoeli-Lerner M: Akt signaling
and cancer: Surviving but not moving on. Cancer Res. 66:3963–3966.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Choi EJ and Kim GH: Apigenin causes G(2)/M
arrest associated with the modulation of p21(Cip1) and Cdc2 and
activates p53-dependent apoptosis pathway in human breast cancer
SK-BR-3 cells. J Nutr Biochem. 20:285–290. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Shukla S and Gupta S: Molecular mechanisms
for apigenin-induced cell-cycle arrest and apoptosis of hormone
refractory human prostate carcinoma DU145 cells. Mol Carcinog.
39:114–126. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Shukla S and Gupta S: Apigenin-induced
cell cycle arrest is mediated by modulation of MAPK, PI3K-Akt, and
loss of cyclin D1 associated retinoblastoma dephosphorylation in
human prostate cancer cells. Cell Cycle. 6:1102–1114. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lee WJ, Chen WK, Wang CJ, Lin WL and Tseng
TH: Apigenin inhibits HGF-promoted invasive growth and metastasis
involving blocking PI3K/Akt pathway and beta 4 integrin function in
MDA-MB-231 breast cancer cells. Toxicol Appl Pharmacol.
226:178–191. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Guo YJ, Pan WW, Liu SB, Shen ZF, Xu Y and
Hu LL: ERK/MAPK signalling pathway and tumorigenesis. Exp Ther Med.
19:1997–2007. 2020.PubMed/NCBI
|
|
49
|
Yang S and Liu G: Targeting the
Ras/Raf/MEK/ERK pathway in hepatocellular carcinoma. Oncol Lett.
13:1041–1047. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Patel AL and Shvartsman SY: Outstanding
questions in developmental ERK signaling. Development.
145:dev1438182018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhao G, Han X, Cheng W, Ni J, Zhang Y, Lin
J and Song Z: Apigenin inhibits proliferation and invasion, and
induces apoptosis and cell cycle arrest in human melanoma cells.
Oncol Rep. 37:2277–2285. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhao M, Ma J, Zhu HY, Zhang XH, Du ZY, Xu
YJ and Yu XD: Apigenin inhibits proliferation and induces apoptosis
in human multiple myeloma cells through targeting the trinity of
CK2, Cdc37 and Hsp90. Mol Cancer. 10:1042011. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Jin XY and Ren CS: Effect and mechanism of
apigenin on VEGF expression in human breast cancer cells. Zhonghua
Zhong Liu Za Zhi. 29:495–499. 2007.(In Chinese). PubMed/NCBI
|
|
54
|
Lee SH, Ryu JK, Lee KY, Woo SM, Park JK,
Yoo JW, Kim YT and Yoon YB: Enhanced anti-tumor effect of
combination therapy with gemcitabine and apigenin in pancreatic
cancer. Cancer Lett. 259:39–49. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Pecorino L: Molecular biology of cancer:
Mechanisms, targets, and therapeutics. Oxford University Press; pp.
157–182. 2012
|
|
56
|
Yang Y, Zhu X, Chen Y, Wang X and Chen R:
p38 and JNK MAPK, but not ERK1/2 MAPK, play important role in
colchicine-induced cortical neurons apoptosis. Eur J Pharmacol.
576:26–33. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ballif BA and Blenis J: Molecular
mechanisms mediating mammalian mitogen-activated protein kinase
(MAPK) kinase (MEK)-MAPK cell survival signals. Cell Growth Differ.
12:397–408. 2001.PubMed/NCBI
|
|
58
|
Moos PJ and Fitzpatrick FA: Taxanes
propagate apoptosis via two cell populations with distinctive
cytological and molecular traits. Cell Growth Differ. 9:687–697.
1998.PubMed/NCBI
|