Introduction
Osteoarthritis is a common chronic degenerative
joint disease characterized by synovial inflammation and cartilage
protein degradation (1).
Osteoarthritis can cause pain and disability, and it will become
the largest disability problem in the United States by 2030
(2). Unfortunately, there are no
effective treatments that can change the progression of
osteoarthritis (3). Currently,
osteoarthritis is mainly treated with a regimen that controls
symptoms and relieves pain (4).
Therefore, it is essential to explore the pathogenesis of
osteoarthritis for the development of effective osteoarthritis
treatment programs.
Long non-coding RNAs (lncRNAs) are a type of
non-protein-encoding RNAs >200 nucleotides in length, which
exert crucial regulatory roles in gene regulatory networks
(5). lncRNAs are implicated in
vital physiological processes, such as cell lineage determination,
cell differentiation, organogenesis and tissue homeostasis
(6). Previous studies have
demonstrated that lncRNAs are involved in the progression of
osteoarthritis (7–9). For example, lncRNA FOXD2-AS1 was
revealed to modulate the proliferation of chondrocytes in
osteoarthritis (9). lncRNA
plasmacytoma variant translocation 1 (PVT1), also known as
LINC00079, MIR1204HG, or onco-lncRNA-100, is associated with the
progression of a range of tumors (10). PVT1 has been revealed to accelerate
lipopolysaccharide (LPS)-induced septic acute kidney injury
(11). PVT1 was demonstrated to
impede cardiac function and facilitate the secretion of
inflammatory factors in a sepsis model (12). Additionally, PVT1 has been revealed
to modulate the apoptosis of chondrocytes in osteoarthritis
(13). Nevertheless, the role of
PVT1 and its molecular mechanisms in the pathogenesis of
osteoarthritis remain to be elucidated.
MicroRNAs (miRNAs) are another crucial class of
non-coding RNAs that bind to complementary mRNAs, resulting in
translation inhibition or degradation of mRNA (14). miRNAs serve vital roles in cell
differentiation and metabolism, organ development, viral infection
and tumorigenesis (15).
miRNA-93-5p (miR-93-5p) serves different functions in different
tumors (16,17). In addition, miR-93-5p can suppress
osteogenic differentiation (18).
miR-93-5p has also been implicated in the inflammatory and
antiproliferative processes of human bronchial epithelial cell
lines induced by neodymium oxide (19). Furthermore, miR-93-5p has been
revealed to regulate IL-1β-induced cartilage degradation and
chondrocyte apoptosis in osteoarthritis (20). However, the role of miR-93-5p in
osteoarthritis remains to be elucidated.
Exosomes are small extracellular vesicles with a
lipid bilayer member structure that can be secreted by most cells
and have a diameter of ~30–100 nm (21). Exosomes contain cell-specific
proteins, lipids and nucleic acids that can be transmitted as
signal molecules to other cells to alter their function (22,23).
Small extracellular vesicles may exert a role in the pathogenesis
of a variety of inflammatory diseases (24). For instance, HIF-1α-induced
exosomal miR-23a can activate macrophages and promote
tubulointerstitial inflammation (25). In addition, the exosomal MALAT1
derived from adipose-derived stem cells regulates
inflammation-related networks and promotes regeneration after
traumatic brain injury (26). A
previous study reported that exosomes released from synovial
fibroblasts in patients with rheumatoid arthritis contain TNF-α
membrane forms, which could make activated T cells resistant to
apoptosis and contribute to the pathogenic process of rheumatoid
arthritis (27). However, the
function of exosomal PVT1 in the development of osteoarthritis is
unclear.
Hence, the present study explored the expression of
exosomal PVT1 in the serum of patients with osteoarthritis and in
an osteoarthritis cell model (LPS-stimulated C28/I2 cells) in
vitro. The effect of exosomes of LPS-stimulated C28/I2 cells
and the serum of patients with osteoarthritis on the expression of
exosomal PVT1 in C28/I2 cells was investigated. The present study
also explored the role of PVT1 in the viability, apoptosis and
inflammation responses of LPS-stimulated C28/I2 cells and its
molecular mechanisms to provide possible therapeutic tactics for
the treatment of osteoarthritis.
Materials and methods
Osteoarthritis subjects
The present study was authorized by the Ethics
Committee of Weihai Municipal Hospital. A total of 30
osteoarthritis patients (19 females and 1 males, age range from 50
to 70 years old) and 30 healthy volunteers (19 females and 11
males, age range from 50 to 70 years old) were recruited from
Weihai Municipal Hospital between October 2017 and June 2019. Whole
blood (20 ml) was drawn from each osteoarthritis patient and
healthy volunteer. The blood sample was kept at room temperature
for 2 h and centrifuged (4°C, 1,200 × g, 20 min) to acquire serum.
Serum was stored in liquid nitrogen for subsequent studies. All
patients with osteoarthritis and the healthy subjects who
participated in the present study signed written informed
consent.
Cell culture and treatment
Human normal chondrocytes C28/I2 were acquired from
BeNa Culture Collection. Dulbecco's modified Eagle's medium/F-12
medium (1:1, DMEM/F-12; HyClone; Cytiva) containing fetal bovine
serum (10%, FBS, HyClone; Cytiva) was employed to maintain the
C28/I2 cells. A humidified incubator with 5% CO2 at 37°C
was used to maintain the C28/I2 cells. C28/I2 cells were exposed to
different concentrations of LPS (1, 5 and 10 µg/ml; Sigma-Aldrich;
Merck KGaA) for 48 h to construct an osteoarthritis state cell
model.
Exosome isolation
Exosome-free medium (Sigma-Aldrich; Merck KGaA) was
used to culture C28/I2 cells or LPS-stimulated C28/I2 cells at 37°C
for 48 h. The culture medium was collected and centrifuged (1,500 ×
g, 15 min) at 4°C and the supernatant filtered through a 0.22-µm
polyvinylidene difluoride (PVDF) filter (EMD Millipore). ExoQuick™
Exosome Precipitation Solution (System Biosciences, LLC) was added
to the serum or filter medium and mixed. Following refrigeration
for 24 h at 4°C, the mixture was centrifuged (1,500 × g, 30 min) at
4°C to remove the supernatant. The exosome pellets were suspended
in the exosome-free medium and the ExoELISA Exosome Quantification
Assay kit (System Biosciences, LLC) was used to quantify the
concentration of exosomes. Western blot analysis was performed to
examine the exosomal markers CD9 and CD63. C28/I2 cells were
treated with 5 µg/ml exosomes.
Transmission electron microscopy
(TEM)
The exosome pellets were suspended in PBS and then
fixed in paraformaldehyde (4%) and glutaraldehyde (4%) in PBS (0.1
M, pH 7.4) at 4°C for 5 min. After adding a drop of the exosomal
sample, the carbon-coated copper grid was immersed in a
phosphotungstic acid solution (2%, pH 7.0) for 30 sec. A
transmission electron microscope (JEM-1200EX; JEOL, Ltd.) at
magnification, ×100,000 was used to observe and assess the
morphology and size of the exosomes.
Cell transfection
Small interfering RNA (siRNA) targeting PVT1
(si-PVT1) and negative control (si-NC) were procured from Shanghai
GenePharma Co., Ltd., as were miRNA mimics and inhibitors targeting
miR-93-5p (miR-93-5p and anti-miR-93-5p) and their negative control
(miR-NC and anti-miR-NC). The sequence of high mobility
groupprotein B1 (HMGB1; accession: NM_001363661) was cloned into
the pcDNA3.1 vector (pcDNA; Invitrogen; Thermo Fisher Scientific,
Inc.) to construct the overexpression vector of HMGB1 (HMGB1).
Oligonucleotides (si-PVT1 (40 nM), si-NC (40 nM), miR-93-5p (50
nM), miR-NC (50 nM), anti-miR-93-5p (50 nM), anti-miR-NC (50 nM))
or plasmids were transiently transfected into C28/I2 cells using
Lipofectamine® 3000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) and then cultured in DMEM/F-12 containing 5 µg/ml
LPS for 48 h. The sequences were: si-PVT1
(5′-GGGUACUGGAAGUAGAAUAUU−3′) and si-NC
(5′-UUCUCCGAACGUGUCACGUTT−3′).
Western blot analysis
Radio-immunoprecipitation assay (RIPA) lysis buffer
(Sigma-Aldrich; Merck KGaA-Aldrich; Merck KGaA) was used to extract
the protein of serum and cells. Total protein concentration was
quantified using a BCA protein quantification kit (Invitrogen;
Thermo Fisher Scientific, Inc.). The extracted total protein (20
µg) was separated through the sodium dodecyl
sulphate-polyacrylamide gel electrophoresis by 10% running gel and
5% stacking gel. Then, the separated protein was transferred onto
PVDF membranes. Following immersion in TBST buffer with 5% skimmed
milk at room temperature for 1 h, the PVDF membranes were incubated
with primary antibodies overnight at 4°C. The primary antibodies
were obtained from Abcam or Santa Cruz Biotechnology, including
anti-CD9 (cat. no. ab92726, 1:100), anti-CD63 (cat. no. ab59479,
1:200), anti-B cell lymphoma 2 (Bcl-2; cat. no. ab182858, 1:200),
anti-cleaved caspase-3 (cat. no. ab32042, 1:500),
anti-Bcl-2-associated X (Bax; cat. no. ab32503, 1:1,000),
anti-GAPDH (cat. no. ab8245, 1:1,000), anti-interleukin (IL)-6
(cat. no. ab6672, 1:500), anti-IL-1β (cat. no. ab9722, 1:250),
anti-TNF-α (sc-52B83, 1:500), anti-HMGB1 (cat. no. ab79823,
1:1,000), anti-Toll-like receptor 4 (TLR4; cat. no. ab13556,
1:500), anti-aggrecan (cat. no. ab3778, 1:100), anti-matrix
metallopeptidase 13 (MMP13; cat. no. ab84594, 1:200), anti-NF-κB
p65 (cat. no. ab16502, 1:200), anti-phosphorylated (p)-p65 (cat.
no. ab97726, 1:1,000), anti-nuclear factor κ-B (NF-κB) inhibitor α
(IκBα; cat. no. ab7217, 1:1,000) and anti-p-IκB-α (cat. no.
ab24783, 1:1,000). Then the membranes were incubated with the
secondary antibody goat anti-mouse IgG (cat. no. ab205719,
1:10,000) or anti-rabbit IgG (cat. no. ab205718, 1:1,000, both from
Abcam) at room temperature for 1 h. GAPDH was used as a loading
control. The Immobilon Western Chemiluminescent HRP Substrate (EMD
Millipore) was used to visualize the protein bands. Quantification
of protein bands was performed using ImageJ software 1.50 (National
Institutes of Health).
Reverse transcription-quantitative
(RT-q) PCR
Total RNA of the exosomes, serum and cells
(1×106) was extracted through TRIzol® reagent
(Thermo Fisher Scientific, Inc.). The first-strand complementary
DNA (cDNA) for PVT1 and HMGB1 was generated through a Moloney
Murine Leukemia Virus (M-MLV) First Strand kit (Thermo Fisher
Scientific, Inc.). The first-strand cDNA for miR-93-5p was
synthesized using MiRNA Reverse Transcription kit (Thermo Fisher
Scientific, Inc.). RNA extraction and cDNA synthesis were performed
in accordance with the manufacturer's protocols. RT-qPCR was
conducted using SYBR Green PCR Master mixes (Thermo Fisher
Scientific, Inc.) in a reaction volume of 20 µl (10 µl SYBR Premix
Ex Taq II, 6 µl ddH2O, 2 µl cDNA, and 2 µl primer) with the
following reaction program: 95°C for 30 sec, followed by 40 cycles
of 95°C for 5 sec and 60°C for 20 sec. The primers used were: PVT1:
5′-TTGGCACATACAGCCATCAT-3′ forward (F) and
5′-GCAGTAAAAGGGGAACACCA-3′ reverse (R); HMGB1:
5′-CTCAGAGAGGTGGAAGACCATGT-3′ (F) and
5′-GGGATGTAGGTTTTCATTTCTCTTTC-3′ (R); GAPDH:
5′-GACTCCACTCACGGCAAATTCA-3′ (F) and 5′-TCGCTCCTGGAAGATGGTGAT−3′
(R) miR-93-5p: 5′-GCCGCCAAAGTGCTGTTC−3′ (F) and
5′-CAGAGCAGGGTCCGAGGTA-3′ (R); as well as U6 small nuclear (sn)
RNA: 5′-GCTCGCTTCGGCAGCACA−3′ (F) and 5′-GAGGTATTCGCACCAGAGGA-3′
(R). The expression of PVT1, HMGB1 and miR-93-5p was calculated by
the 2−ΔΔCq method (28) and GAPDH or U6 snRNA was used as an
internal control. All RT-qPCR reactions were performed in
triplicate.
Cell viability assay
Cell Counting Kit-8 (CCK-8) (Dojindo Molecular
Technologies, Inc.) was applied to assess the viability of cells
according to the manufacturer's protocol. In short, C28/I2 cells
(5×103 cells/well) were treated with LPS (1, 5 and 10
µg/ml) for 48 h or LPS-stimulated C28/I2 cells transfected with
oligonucleotides or plasmids were cultured for 48 h. Then, CCK-8
(10 µl) reagent was supplemented to each well and incubated at 37°C
for 2 h. A microplate absorbance reader (Thermo Fisher Scientific,
Inc.) was used for the evaluation of the color reaction at 450
nm.
Flow cytometric assay
An Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) apoptosis detection kit
(Sigma-Aldrich; Merck KGaA) was employed to assess the apoptosis
rate of cells according to the manufacturer's protocol. In brief,
the C28/I2 cells were treated in the same manner as for the cell
viability assay. Following washing, treated C28/I2 cells
(1×105) were resuspended in binding buffer. Annexin
V-FITC (5 µl) and PI (10 µl) were added to the binding buffer and
incubated at room temperature for 15 min in the dark. A FACScan
flow cytometer (BD Biosciences) with CellQuest software (version
5.1, BD Biosciences) was used to analyze the apoptosis rate of
treated C28/I2 cells.
Enzyme-linked immunosorbent assay
(ELISA)
The levels of inflammatory cytokines IL-6, IL-1β and
TNF-α in the supernatant of treated-C28/I2 cells were analyzed with
human IL-1β (cat. no. SLB50)/IL-6 (cat. no. S6050)/TNF-α (cat. no.
STA00D) Quantikine ELISA kits (R&D Systems Europe, Ltd.).
Dual-luciferase reporter assay
StarBase v2.0 (http://starbase.sysu.edu.cn/) was employed to predict
the binding sites between PVT1 or HMGB1 and miR-93-5p. Then, the
sequences of wild-type (WT) PVT1 (with predicted miR-93-5p binding
sites) and mutant (MUT) PVT1 were amplified and inserted into the
psiCHECK-2 vector (Promega Corporation) to establish the luciferase
reporter vectors WT-PVT1 and MUT-PVT1 for the verification of the
binding sites between PVT1 and miR-93-5p. The same method was used
to construct the reporter vectors HMGB1 3′ untranslated region
(UTR)-WT and HMGB1 3′UTR-MUT for the confirmation of the binding
sites between HMGB1 and miR-93-5p. C28/I2 cells were cotransfected
miR-NC (30 nM) or miR-93-5p (30 nM) and a luciferase reporter
vector (100 ng) Lipofectamine® 3000 reagent (Invitrogen;
Thermo Fisher Scientific, Inc.). After transfection for 48 h, the
luciferase activities of the luciferase reporter vectors were
evaluated with dual-luciferase reporter assay kit (Promega
Corporation). Relative Renilla luciferase activity was
normalized to that of firefly luciferase.
Statistical analysis
SPSS 20.0 software (IBM Corp.) and GraphPad Prism
5.0 (GraphPad Software, Inc.) were used for statistical analysis.
The data in the present study were derived from ≥3 independent
experiments. Data are presented as the mean ± standard deviation.
Unpaired Student's t-test or one-way variance analysis (ANOVA) with
Tukey's post hoc test was employed. Pearson's correlation analysis
method was used to assess the relationship between miR-93-5p and
PVT1 as well as HMGB1 and miR-93-5p expression in the serum of
osteoarthritis patients. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of PVT1, exosomal PVT1,
miR-93-5p in the serum of osteoarthritis patients and
LPS-stimulated C28/I2 cells
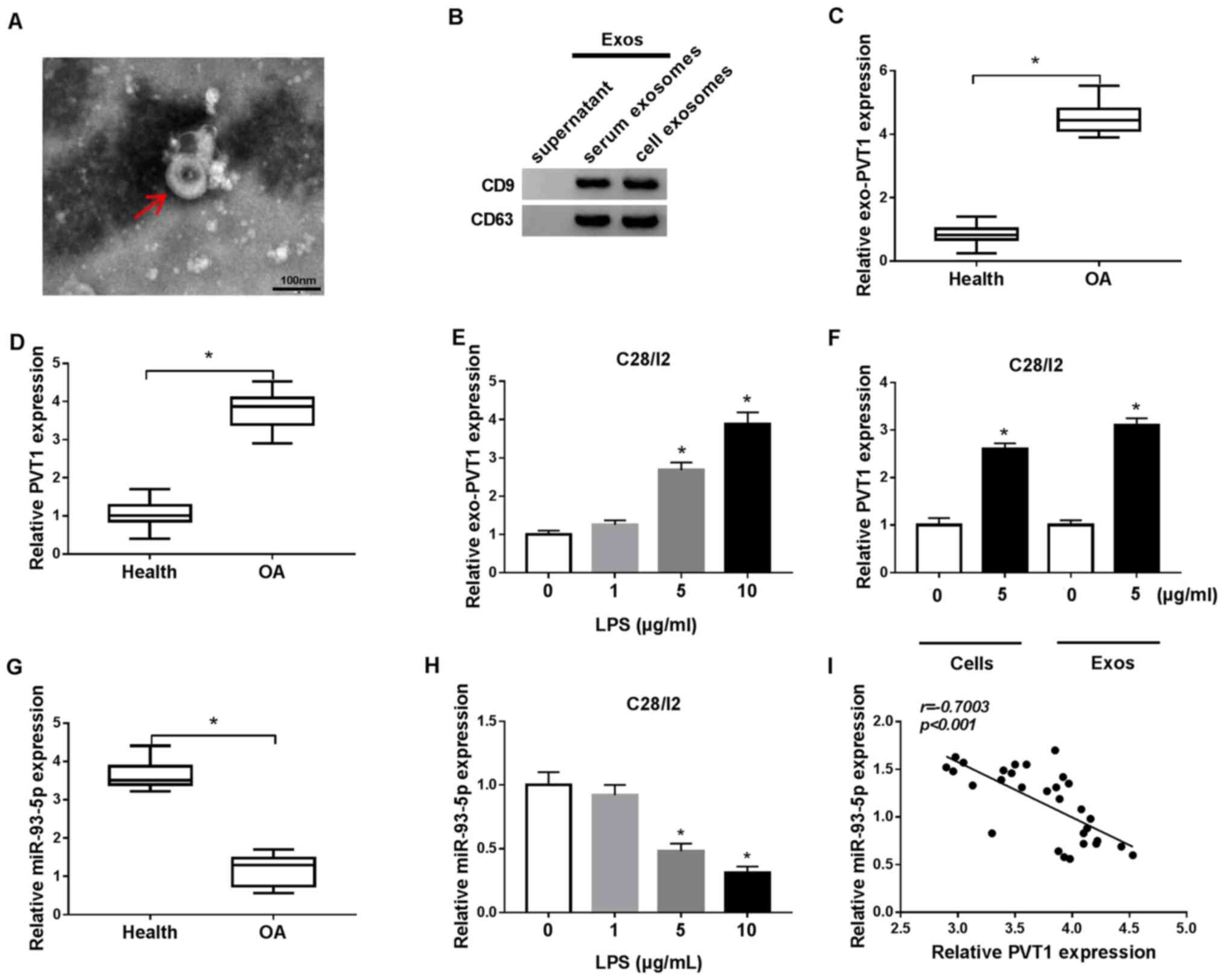
Exosomes were extracted from C28/I2 cells and from
the serum of patients with osteoarthritis to analyze
exosome-encapsulated PVT1 in osteoarthritis. The exosomes were
circular in shape and 40–100 nm in the C28/I2 cells and the serum
of osteoarthritis patients and healthy volunteers (Fig. 1A). Western blot analysis
demonstrated that the exosome marker proteins CD9 and CD63 were
present in the exosomes of the C28/I2 cells and the serum of
osteoarthritis patients (Fig. 1B).
RT-qPCR demonstrated that PVT1 was significantly upregulated in
exosomes of the serum of osteoarthritis patients when compared with
the serum of healthy volunteers (Fig.
1C). In addition, PVT1 was significantly upregulated in the
serum of osteoarthritis patients compared with healthy volunteers
(Fig. 1D). LPS (1, 5 and 10 µg/ml)
was used to treat C28/I2 cells for the construction of the
osteoarthritis status in vitro. It was determined that the
expression of PVT1 was significantly enhanced in exosomes of 5 and
10 µg/ml LPS-stimulated C28/I2 cells (Fig. 1E). Furthermore, it was identified
that the exosomes of 5 µg/ml LPS-stimulated C28/I2 cells and the
serum of osteoarthritis patients could increase the expression of
exosomal PVT1 in C28/I2 cells (Fig.
1F).
miR-93-5p in the serum of patients with
osteoarthritis was decreased compared with the serum of healthy
volunteers (Fig. 1G). In addition,
miR-93-5p was significantly downregulated in LPS (5 and 10
µg/ml)-treated C28/I2 cells (Fig.
1H). Pearson correlation analysis demonstrated that the
expression of miR-93-5p and PVT1 was negatively correlated in the
serum of patients with osteoarthritis (Fig. 1I). Collectively, these results
indicated that the upregulation of PVT1 mediated by exosomes and
the reduction of miR-93-5p may be associated with the pathogenesis
of osteoarthritis.
LPS accelerates cell apoptosis and
inflammation responses and restrains cell viability in C28/I2
cells
The influence of LPS on the viability, apoptosis and
inflammation responses of C28/I2 cells was investigated. A CCK-8
assay demonstrated that the viability of C28/I2 cells was
significantly suppressed with increased concentration of LPS
(Fig. 2A). Flow cytometry revealed
that the apoptosis rate of C28/I2 cells was significantly increased
with increased concentration of LPS (Fig. 2B). The levels of apoptosis-related
proteins (Bcl-2, Bax and cleaved caspase-3) in LPS-stimulated
C28/I2 cells were analyzed with western blot analysis. With
increased concentration of LPS, Bcl-2 protein level in C28/I2 cells
was reduced, while Bax and cleaved caspase-3 protein levels were
increased (Fig. 2C). The levels of
inflammatory cytokines IL-6, IL-1β and TNF-α in the supernatant of
LPS-stimulated C28/I2 cells were measured using ELISA. The results
demonstrated that the levels of IL-6, IL-1β and TNF-α were
increased in the supernatant of C28/I2 cells with increased
concentration of LPS (Fig. 2D).
LPS increased the protein levels of IL-6, IL-1β and TNF-α in C28/I2
cells in a concentration-dependent manner (Fig. 2E). Aggrecan levels were reduced
while MMP13 levels were increased in C28/I2 cells with increased
concentration of LPS, indicating that LPS treatment induced
collagen degradation in C28/I2 cells (Fig. 2E). Collectively, these findings
demonstrated that LPS could induce cell apoptosis and inflammation
responses and inhibit cell viability in C28/I2 cells.
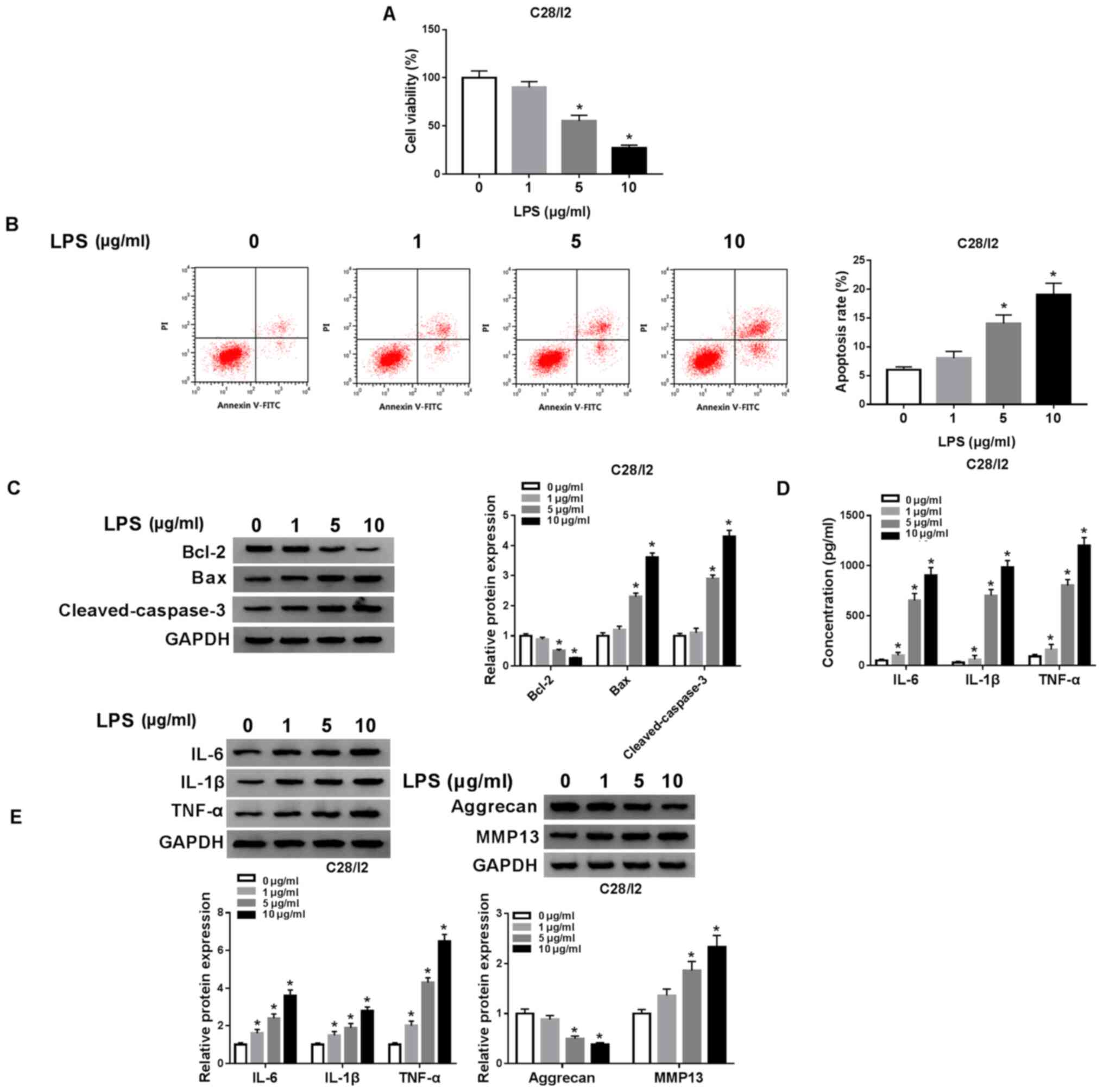 | Figure 2.Effects of LPS on viability and
induced cell apoptosis, inflammation responses and collagen
degradation of C28/I2 cells. (A) Influence of various
concentrations of LPS (0, 1, 5 and 10 µg/ml) on viability of C28/I2
cells as determined by CCK-8 assay. (B) The apoptosis of C28/I2
cells cultured in medium with various concentrations of LPS was
detected by flow cytometry. (C) Western blot analysis was performed
to assess the protein levels of Bcl-2, Bax and cleaved caspase-3 in
the C28/I2 cells cultured in medium with various concentrations of
LPS. (D) The levels of IL-6, IL-1β and TNF-α in C28/I2 cells
stimulated with various concentrations of LPS were assessed using
ELISA. (E) The protein levels of IL-6, IL-1β, TNF-α, aggrecan and
MMP13 in C28/ I2 cells treated with various concentrations of LPS
were assessed by western blot analysis. *P<0.05. LPS,
lipopolysaccharide; IL, interleukin; MMP, matrix
metallopeptidase. |
Depletion of PVT1 overturns
LPS-mediated viability, apoptosis and inflammation responses and
collagen degradation of C28/I2 cells
To investigate the function of PVT1 in
osteoarthritis, a function-loss-experiment was performed in C28/I2
cells treated with LPS (5 µg/ml) for 48 h. The results demonstrated
that PVT1 expression was significantly increased in C28/I2 cells
following exosome treatment. In addition, the upregulation of PVT1
in C28/I2 cells caused by exosome treatment was reversed after
si-PVT1 transfection (Fig. S1).
The transfection of si-PVT1 abolished the upregulation of PVT1 in
LPS-stimulated C28/I2 cells compared with the si-NC group (Fig. 3A). A CCK-8 assay demonstrated that
PVT1 downregulation reversed the suppressive influence of LPS on
the viability of C28/I2 cells (Fig.
3B). The increase of apoptosis in LPS-treated C28/I2 cells was
reversed by the downregulation of PVT1 (Fig. 3C). In addition, the enhancement of
Bax and cleaved caspase-3 proteins and the decrease of Bcl-2
protein in LPS-induced C28/I2 cells were overturned by PVT1
inhibition (Fig. 3D). ELISA
revealed that reduced PVT1 expression abrogated the increase in
IL-6, IL-1β and TNF-α in C28/I2 cells treated with LPS (Fig. 3E). Western blot analysis also
demonstrated that PVT1 silencing reversed LPS-mediated effects on
the protein levels of IL-6, IL-1β, TNF-α, aggrecan and MMP13 in
C28/I2 cells (Fig. 3F). In brief,
PVT1 downregulation abolished LPS-mediated viability, apoptosis,
inflammation responses and collagen degradation in C28/I2
cells.
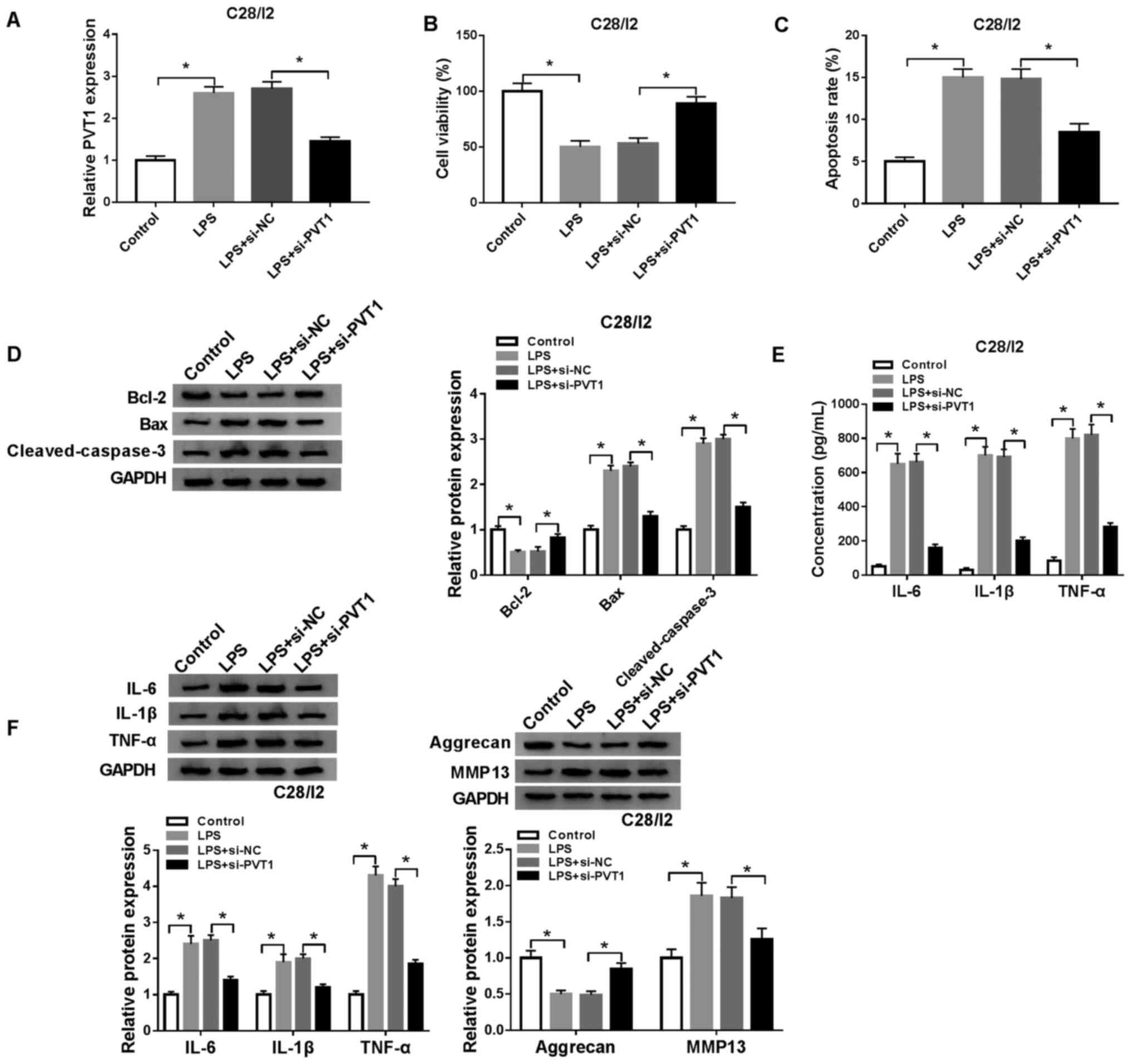 | Figure 3.Effects of PVT1 depletion on
viability, apoptosis, inflammation responses and collagen
degradation in LPS-stimulated C28/I2 cells. LPS-stimulated C28/I2
cells were transfected with si-NC or si-PVT1. (A) RT-qPCR was
employed to analyze the expression of PVT1 in LPS-stimulated C28/I2
cells. (B) Effect of PVT1 inhibition on cell viability in
LPS-stimulated C28/I2 cells was analyzed using a CCK-8 assay. (C)
Influence of PVT1 inhibition on the apoptosis rate of
LPS-stimulated C28/I2 cells as determined using flow cytometry. (D)
Effects of PVT1 inhibition on Bcl-2, Bax and cleaved caspase-3
protein expression levels were evaluated with western blot
analysis. (E) ELISA was used to analyze the effects of PVT1
silencing on IL-6, IL-1β and TNF-α expression levels in the
supernatant of LPS-stimulated C28/I2 cells. (F) Western blot
analysis was performed to assess the influence of PVT1
downregulation on the protein levels of IL-6, IL-1β, TNF-α,
aggrecan and MMP13 in LPS-stimulated C28/I2 cells. *P<0.05.
PVT1, plasmacytoma variant translocation 1; LPS,
lipopolysaccharide; si, short interfering RNA; NC, negative
control; RT-qPCR, reverse transcription-quantitative PCR; IL,
interleukin; MMP, matrix metallopeptidase. |
miR-93-5p acts as a target for
PVT1
To understand the molecular mechanism of PVT1 in
osteoarthritis, starBase v2.0 was used to predict the potential
binding sites for PVT1. miR-93-5p was revealed to be a possible
target for PVT1 (Fig. 4A).
Subsequently, the luciferase reporter vectors WT-PVT1 and MUT-PVT1
were constructed to verify the potential binding sites between PVT1
and miR-93-5p. Dual-luciferase reporter assay demonstrated that
miR-93-5p introduction suppressed the luciferase activity of
WT-PVT1 when compared with the miR-NC, while no significant
difference was observed in luciferase activity between the miR-NC
and MUT-PVT1 groups (Fig. 4B).
RT-qPCR demonstrated that PVT1 expression level was decreased in
LPS-stimulated C28/I2 cells transfected with si-PVT1, but the
expression of PVT1 was enhanced in LPS-stimulated C28/I2 cells
transfected with PVT1 (Fig. 4C).
Decreased PVT1 expression effectively increased miR-93-5p
expression in LPS-stimulated C28/I2 cells, whereas increased levels
of PVT1 suppressed miR-93-5p expression in LPS-stimulated C28/I2
cells (Fig. 4D). The introduction
of anti-miR-93-5p overturned PVT1 downregulation-mediated
enhancement of miR-93-5p in LPS-stimulated C28/I2 cells (Fig. 4E). Whether PVT1 exerted its
function via miR-93-5p in LPS-stimulated C28/I2 cells was then
investigated. A CCK-8 assay demonstrated that the promotion of
proliferation of LPS-stimulated C28/I2 cells caused by PVT1
silencing was reversed by the inhibition of miR-93-5p (Fig. 4F). miR-93-5p silencing overturned
the suppression of apoptosis in LPS-stimulated C28/I2 cells induced
by PVT1 knockdown (Fig. 4G).
Western blot analysis revealed that the downregulation of miR-93-5p
abrogated PVT1 reduction-mediated protein levels of Bcl-2, Bax and
cleaved caspase-3 of LPS-stimulated C28/I2 cells (Fig. 4H). ELISA revealed that miR-93-5p
reduction reversed the suppression of IL-6, IL-1β and TNF-α in
LPS-stimulated C28/I2 cells induced by PVT1 downregulation
(Fig. 4I). Western blot analysis
also revealed that downregulated miR-93-5p expression reversed the
PVT1 inhibition-mediated influence on the protein levels of IL-6,
IL-1β, TNF-α, aggrecan and MMP13 in C28/I2 cells following LPS
treatment (Fig. 4J). Therefore,
these data indicated that PVT1 mediated cell viability, apoptosis,
inflammation responses and collagen degradation in LPS-stimulated
C28/I2 cells via miR-93-5p.
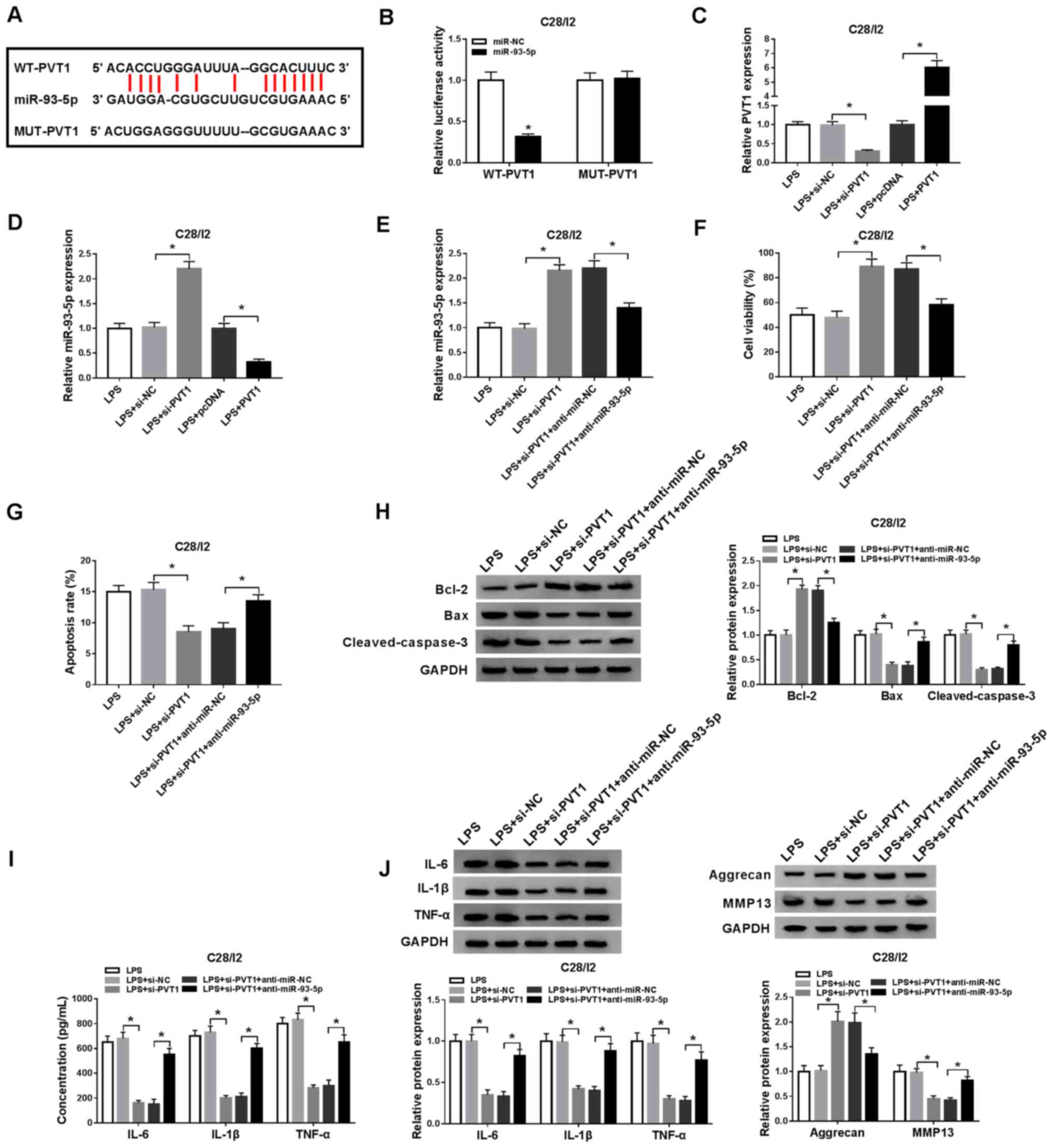 | Figure 4.PVT1 performs its role through
miR-93-5p in C28/I2 cells. (A) The binding sites of PVT1 in
miR-93-5p were predicted using starBase v2.0. (B) The luciferase
intensity in C28/I2 cells cotransfected WT-PVT1 or MUT-PVT1 and
miR-NC or miR-93-5p were determined with dual-luciferase reporter
assay. (C) RT-qPCR was performed to analyze PVT1 expression level
in LPS-stimulated C28/I2 cells transfected with si-PVT1 or PVT1.
(D) Effect of PVT1 on the expression of miR-93-5p was evaluated
using RT-qPCR. LPS-stimulated C28/I2 cells were transfected with
si-NC, si-PVT1, si-PVT1+anti-miR-NC, si-PVT1+anti-miR-93-5p. (E)
Effect of miR-93-5p suppression on PVT1 knockdown-mediated
miR-93-5p expression of LPS-stimulated C28/I2 cells was analyzed
using RT-qPCR. (F) Effect of miR-93-5p inhibition on PVT1
downregulation-mediated viability of LPS-stimulated C28/I2 cells
was determined via CCK-8 assay. (G) Flow cytometry was performed
for the evaluation of the influence of miR-93-5p silencing on PVT1
inhibition-mediated apoptosis of LPS-stimulated C28/I2 cells. (H)
Western blot analysis was performed to detect the protein levels of
Bcl-2, Bax and cleaved caspase-3 in LPS-stimulated C28/ I2 cells.
(I) The levels of IL-6, IL-1β and TNF-α in LPS-stimulated C28/I2
cells were measured using ELISA. (J) The protein levels of IL-6,
IL-1β, TNF-α, aggrecan and MMP13 in LPS-stimulated C28/I2 cells
were explored using western blot analysis. *P<0.05. PVT1,
plasmacytoma variant translocation 1; miR, microRNA; WT, wild-type;
MUT, mutant; RT-qPCR, reverse transcription-quantitative PCR; LPS,
lipopolysaccharide; si, short interfering RNA; NC, negative
control; IL, interleukin; MMP, matrix metallopeptidase. |
HMGB1 serves as a target for
miR-93-5p
To further investigate the molecular mechanism of
miR-93-5p in osteoarthritis, the latent target for miR-93-5p was
explored in starBase v2.0. As illustrated in Fig. 5A, miR-93-5p possessed latent
binding sites in HMGB1. The luciferase reporter vectors HMGB1
3′UTR-WT and HMGB1 3′UTR-MUT were then established and a
dual-luciferase reporter assay demonstrated a significant
suppression of luciferase activity of HMGB1 3′UTR-WT in C28/I2
cells transfected with miR-93-5p compared with the negative control
group, while HMGB1 3′UTR-MUT luciferase activity was not noticeably
altered (Fig. 5B). In addition,
the mRNA and protein levels of HMGB1 were significantly upregulated
in the serum of osteoarthritis patients (Fig. 5C). The mRNA and protein levels of
HMGB1 were significantly increased in LPS (5 µg/ml)-treated C28/I2
cells, implying that HMGB1 may be associated with LPS-induced
osteoarthritis (Fig. 5D and E). A
negative correlation was observed between miR-93-5p and HMGB1 in
the serum of osteoarthritis patients (Fig. 5F). The introduction of miR-93-5p
significantly upregulated miR-93-5p in LPS-stimulated C28/I2 cells,
whereas the introduction of anti-miR-93-5p downregulated miR-93-5p
in LPS-stimulated C28/I2 cells (Fig.
5G). The mRNA and protein levels of HMGB1 were suppressed by
miR-93-5p introduction in LPS-stimulated C28/I2 cells, while these
effects were reversed by miR-93-5p silencing (Fig. 5H and I). These findings revealed
that miR-93-5p negatively regulated the expression of HMGB1 in
LPS-stimulated C28/I2 cells.
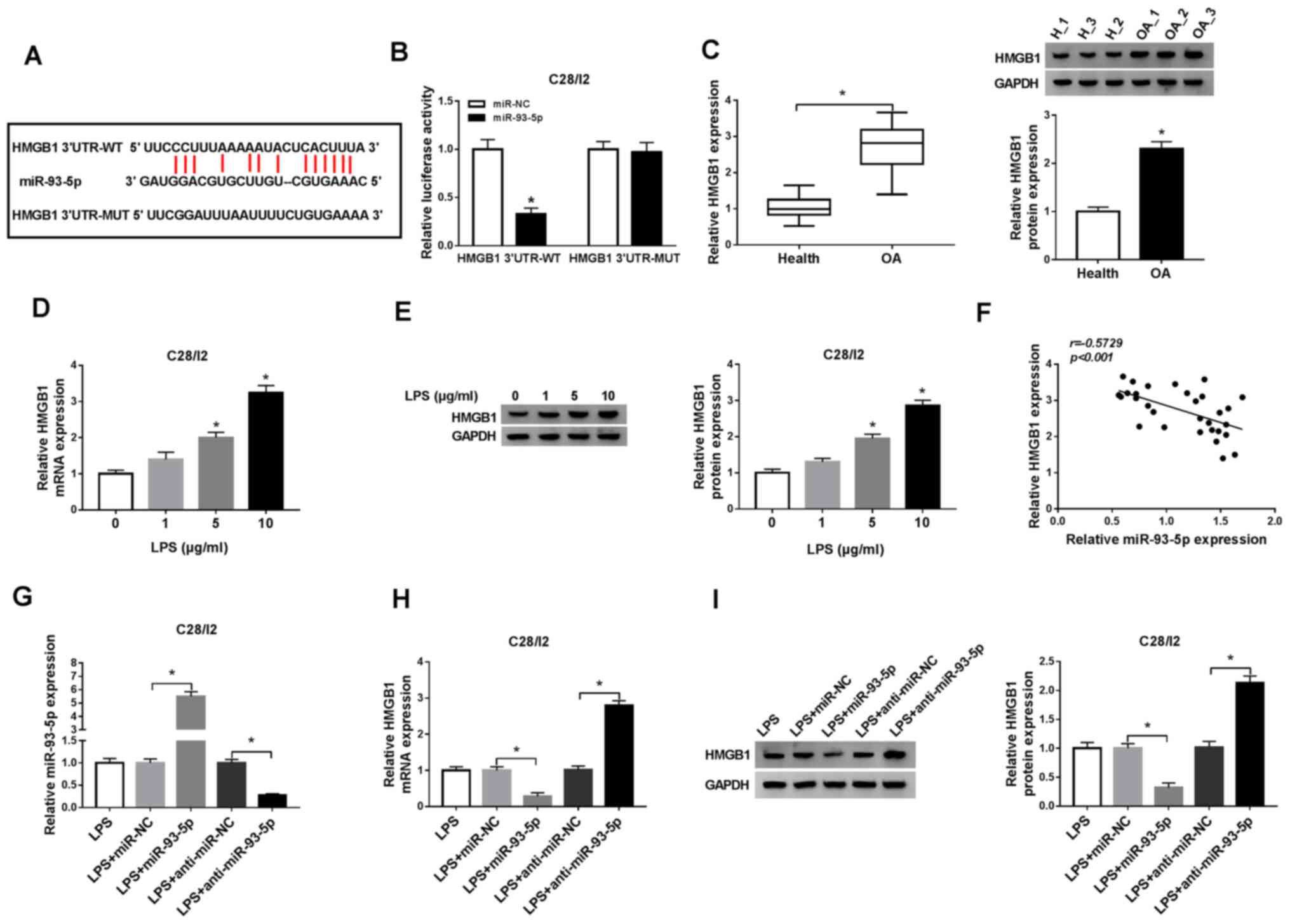 | Figure 5.HMGB1 serves as a target for
miR-93-5p. (A) miR-93-5p latent binding sites in HMGB1 as predicted
by starBase v2.0. (B) Dual-luciferase reporter assay was performed
to determine the luciferase intensity in C28/I2 cells cotransfected
with HMGB1 3′UTR-WT or HMGB1 3′UTR-MUT and miR-NC or miR-93-5p. (C)
RT-qPCR or western blot analysis was employed to evaluate the
levels of HMGB1 mRNA and protein in the serum of osteoarthritis
patients. The expression levels of HMGB1 mRNA and protein in C28/I2
cells treated with LPS (0, 1, 5 and 10 µg/ml) were measured using
(D) RT-qPCR or (E) western blot analysis. (F) The correlation
between HMGB1 and miR-93-5p in the serum of osteoarthritis patients
was analyzed via Pearson correlation analysis. (G) The effect of
miR-93-5p on the expression of miR-93-5p of LPS-stimulated C28/I2
cells was assessed by RT-qPCR. The effect of miR-93-5p on the mRNA
and protein levels of HMGB1 miR-93-5p of LPS-stimulated C28/I2
cells was determined by (H) RT-qPCR or (I) western blot analysis.
*P<0.05. HMGB1, high mobility groupprotein B1; UTR, untranslated
region; WT, wild-type; MUT, mutant; miR, microRNA; NC, negative
control; RT-qPCR, reverse transcription-quantitative PCR; LPS,
lipopolysaccharide. |
HMGB1 increase abolishes miR-93-5p
overexpression-mediated viability, apoptosis, inflammation
responses and collagen degradation of LPS-stimulated C28/I2
cells
To ascertain whether miR-93-5p-mediated viability,
apoptosis and inflammation responses of LPS-stimulated C28/I2 cells
were dependent on HMGB1, the expression of HMGB1 was explored in
LPS-stimulated C28/I2 cells transfected with miR-NC, miR-93-5p,
miR-93-5p+pcDNA or miR-93-5p+HMGB1. The results revealed that the
mRNA and protein levels of HMGB1 were inhibited by miR-93-5p
upregulation in LPS-stimulated C28/I2 cells, but this suppression
was overturned by the addition of HMGB1 (Fig. 6A and B). A CCK-8 assay revealed
that the enhanced miR-93-5p expression resulted in increased
viability of LPS-stimulated C28/I2 cells, whereas this influence
was reversed by the upregulation of HMGB1 (Fig. 6C). Flow cytometry revealed that
miR-93-5p overexpression served a repressive role in the apoptosis
of LPS-stimulated C28/I2 cells, while this influence was restored
by upregulation of HMGB1 expression (Fig. 6D). Overexpression of HMGB1
abolished the decrease of Bax and cleaved caspase-3 and the
upregulation of Bcl-2 caused by miR-93-5p increase in
LPS-stimulated C28/I2 cells (Fig.
6E). In addition, the downregulation of IL-6, IL-1β, TNF-α and
MMP13 and the upregulation of aggrecan in LPS-stimulated C28/I2
cells caused by miR-93-5p upregulation were overturned by HMGB1
expression (Fig. 6F and G). PVT1
depletion significantly downregulated the mRNA and protein levels
of HMGB1 in LPS-stimulated C28/I2 cells, but this reduction was
recovered by the inhibition of miR-93-5p (Fig. 6H and I). The aforementioned data
indicated that miR-93-5p mediated cell viability, apoptosis,
inflammation responses and collagen degradation in LPS-stimulated
C28/I2 cells through HMGB1.
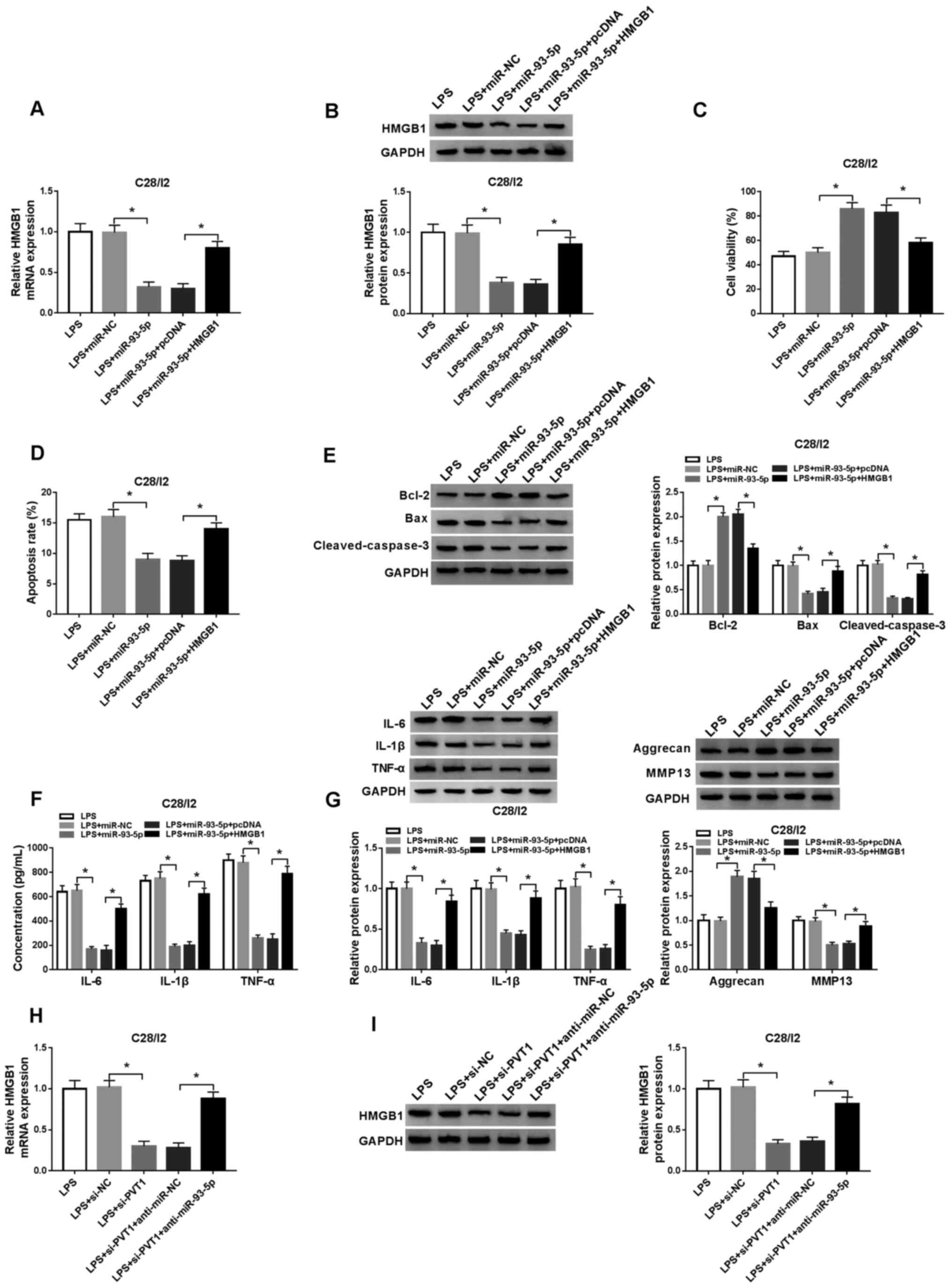 | Figure 6.miR-93-5p exerts its function in
LPS-stimulated C28/I2 cells through HMGB1. LPS-stimulated C28/I2
cells were transfected with miR-NC, miR-93-5p, miR-93-5p+pcDNA or
miR-93-5p+HMGB1. The mRNA and protein expression levels of HMGB1 in
LPS-stimulated C28/I2 cells were assessed through (A) RT-qPCR or
(B) western blot analysis. (C) A CCK-8 assay was performed to
determine the viability of LPS-stimulated C28/I2 cells. (D) The
apoptosis of LPS-stimulated C28/I2 cells was detected by flow
cytometry. (E) The protein levels of Bcl-2, Bax and cleaved
caspase-3 in LPS-stimulated C28/I2 cells were evaluated by western
blot analysis. (F) ELISA was applied to analyze the levels of IL-6,
IL-1β and TNF-α in LPS-stimulated C28/I2 cells. (G) Western blot
analysis was performed to assess the protein levels of IL-6, IL-1β,
TNF-α, aggrecan and MMP13 in the LPS-stimulated C28/I2 cells. (H)
RT-qPCR or (I) western blot analysis was employed to detect HMGB1
mRNA and protein expression levels in LPS-stimulated C28/I2 cells
transfected with si-NC, si-PVT1, si-PVT1+anti-miR-NC and
si-PVT1+anti-miR-93-5p. *P<0.05. miR, microRNA; LPS,
lipopolysaccharide; HMGB1, high mobility groupprotein B1; NC,
negative control; RT-qPCR, reverse transcription-quantitative PCR;
IL, interleukin; MMP, matrix metallopeptidase; si, short
interfering RNA. |
PVT1 silencing blocks the TLR4/NF-κB
pathway through the miR-93-5p/HMGB1 axis
The TLR4/NF-κB pathway is associated with the
secretion of pro-inflammatory cytokines (29). Hence, the present study
investigated whether the TLR4/NF-κB pathway was involved in
PVT1-mediated cell viability, apoptosis and inflammation responses
in LPS-stimulated C28/I2 cells. Western blot analysis demonstrated
that LPS treatment promoted the protein levels of p-p65, TLR4 and
p-IκB-α in C28/I2 cells (Fig. 7A).
Inhibition of PVT1 reversed the increase of TLR4, p-p65 and p-IκB-α
proteins in C28/I2 cells induced by LPS. However, inhibition of
miR-93-5p and HMGB1 upregulation abolished the suppressive impact
of PVT1 suppression on p-p65, TLR4 and p-IκB-α protein expression
levels in LPS-stimulated C28/I2 cells (Fig. 7B). Additionally, the data indicated
that LPS-induced PVT1 accelerated osteoarthritis by activating the
TLR4/NF-κB pathway through the miR-93-5p/HMGB1 axis in C28/I2 cells
(Fig. 7C). These findings
indicated that PVT1 regulated the TLR4/NF-κB pathway through the
miR-93-5p/HMGB1 axis.
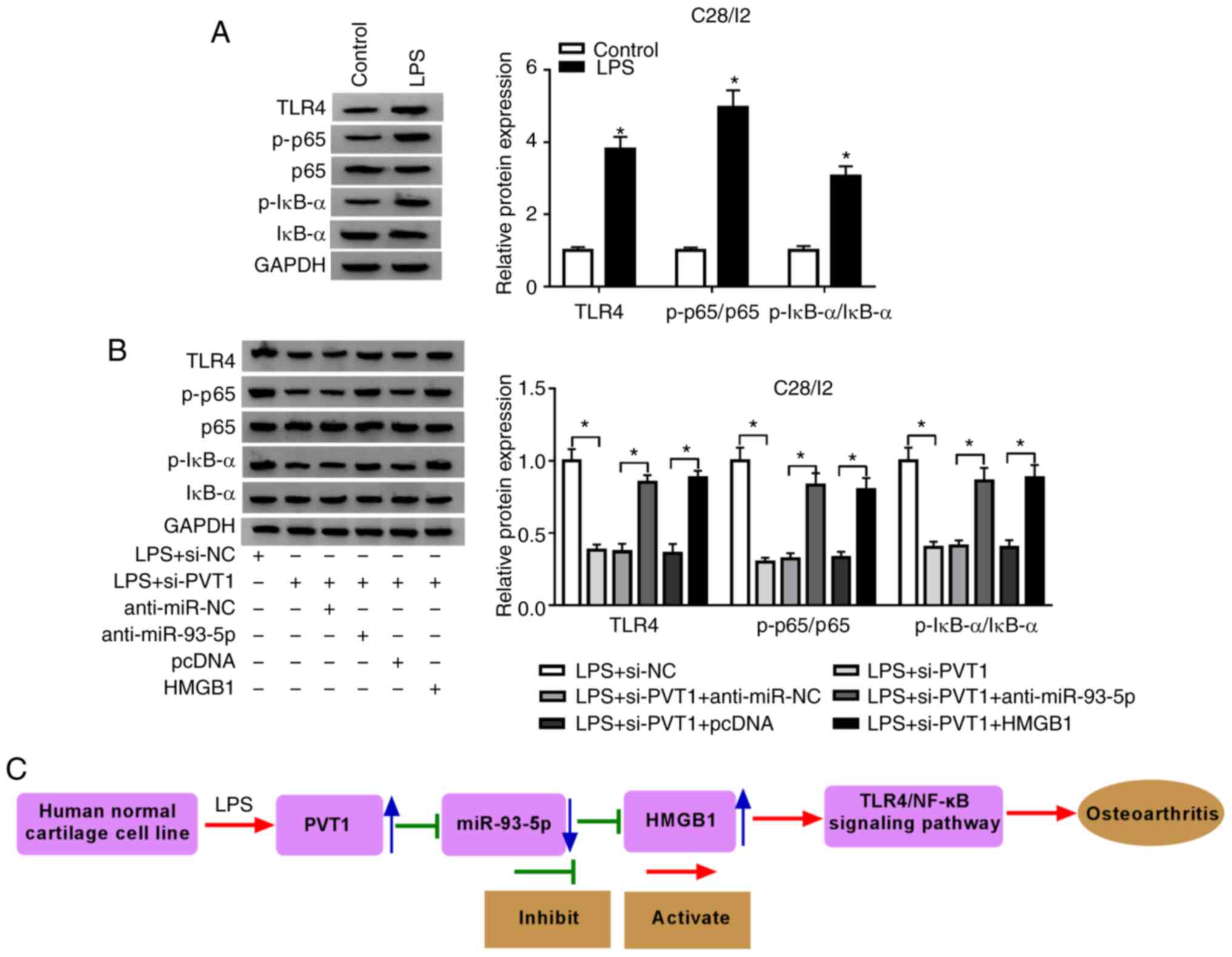 | Figure 7.PVT1 modulates the TLR4/NF-κB pathway
via the miR-93-5p/HMGB1 axis. (A) Effects of LPS on the expression
of TLR4, p-p65 and p-IκB-α proteins of C28/I2 cells were determined
with western blot analysis. (B) The expression of TLR4, p-p65 and
p-IκB-α proteins in LPS-stimulated C28/I2 cells transfected with
si-NC, si-PVT1, si-PVT1+anti-miR-NC, si-PVT1+anti-miR-93-5p,
si-PVT1+pcDNA, or si-PVT1+anti-HMGB1. (C) The schematic
presentation of the LPS/PVT1/miR-93-5p/HMGB1 axis in
osteoarthritis. *P<0.05. PVT1, plasmacytoma variant
translocation 1; TLR4,Toll-like receptor 4; miR, microRNA; HMGB1,
high mobility groupprotein B1; LPS, lipopolysaccharide; p-,
phosphorylated; IκBα, NF-κB inhibitor α; si, short interfering RNA;
NC, negative control. |
Discussion
Osteoarthritis mainly affects the health of
middle-aged and elderly people and its clinical manifestations
include joint pain, deformity and dysfunction (30). LPS is a key pro-inflammatory factor
associated with the onset of osteoarthritis (31). Studies have revealed that LPS can
be used to construct a cell injury model in vitro (32–34).
The present study used 5 µg/ml LPS to construct a cell injury model
in vitro. Increasing evidence emphasizes the association of
lncRNAs with the progression of osteoarthritis and the probability
that lncRNAs function as therapeutic targets and biomarkers
(8,9,35,36).
Exosomes can transport their loaded nucleic acids and proteins to
recipient cells, thereby playing a central role in cell-to-cell
communication (37). The present
study revealed that exosomal PVT1 was upregulated in the serum of
osteoarthritis patients and LPS-stimulated C28/I2 cells. In
addition, PVT1 was upregulated in the serum of osteoarthritis
patients compared with the healthy volunteers. Notably, the
exosomes of the serum of osteoarthritis patients and LPS-stimulated
C28/I2 cells could enhance the expression of PVT1 in C28/I2 cells.
Li et al (13) revealed
that PVT1 enhancement facilitates cell apoptosis in chondrocytes in
osteoarthritis. A previous study stated that increased PVT1
expression expedites inflammation and aberrant metabolic
dysfunction in IL-β-induced chondrocytes (38). PVT1 is associated with the
development of hyperglycemia-induced collagen degradation in
chondrocytes (36). In the present
study, inhibition of PVT1 reversed the decrease of viability and
the increase of apoptosis, inflammation responses and collagen
degradation in C28/I2 cells induced by LPS, implying that PVT1
acted as an unfavorable factor in osteoarthritis.
Previous studies suggest that PVT1 can be used as a
sponge for miRNAs to participate in the development of
osteoarthritis (13,38,39).
miR-93-5p-containing exosomes attenuated the myocardial injury
caused by acute myocardial damage (40). In addition, miR-93-5p was revealed
to reverse the inflammatory and antiproliferative processes of
neodymium oxide-induced human bronchial epithelial cell lines
caused by circular RNA 0039411 (19). Xue et al (20) noted that miR-93-5p was decreased in
IL-β-induced chondrocytes and human and rat osteoarthritis-affected
cartilage and that introduction of miR-93-5p suppressed cell
apoptosis, increased cell viability and maintained cell metabolism
balance in IL-β-induced chondrocytes. In the present study,
miR-93-5p was downregulated in the serum of osteoarthritis patients
and LPS-stimulated C28/I2 cells. Furthermore, miR-93-5p was a
target of PVT1. Downregulation of miR-93-5p abolished PVT1
silencing-mediated viability, apoptosis, inflammation responses and
collagen degradation of LPS-stimulated C28/I2 cells. These data
implied that PVT1 mediated cell viability, apoptosis and
inflammation responses in osteoarthritis through miR-93-5p.
HMGB1 is a member of the damage-associated molecular
patterns (41). It is released
from damaged or dead cells during tissue damage, necrosis,
inflammation and hypoxia, resulting in a sustained inflammatory
environment (42). The HMGB1-LPS
complex accelerates the transformation of osteoarthritis synovial
fibroblasts into the synovial fibroblast-like phenotype of
rheumatoid arthritis (43). HMGB1
has been revealed to promote cell apoptosis, NF-κB and the
production of IL-6, IL-1β and TNF-α in LPS-induced chondrocytes
(44). In addition, necrostatin-1
has been revealed to ameliorate IL-1β-induced apoptosis and
trauma-induced mouse osteoarthritis in primary mouse chondrocytes
by inhibiting HMGB1/TLR4/SDF-1 (45). Inhibition of the transcriptional
activity of HMGB1 and the NF-κB pathway by BRD4 silencing can
attenuate chondrocyte inflammation and catabolism (46). In the present study, HMGB1 was
enhanced in the serum of osteoarthritis patients and LPS-stimulated
C28/I2 cells. Overexpression of HMGB1 abolished the decrease of
viability and the increase of apoptosis, inflammation responses and
collagen degradation in LPS-stimulated C28/I2 cells caused by
miR-93-5p upregulation. In addition, PVT1 inhibition could block
the HMGB1/TLR4/NF-κB pathway through miR-93-5p. Therefore, the
present study indicated that PVT1 inhibition alleviated LPS-induced
damage in osteoarthritis by blocking the HMGB1/TLR4/NF-κB pathway
through miR-93-5p.
In summary, PVT1 was upregulated in serum and serum
exosomes of osteoarthritis patients, as well as LPS-stimulated
C28/I2 cells. PVT1 inhibition enhanced viability and suppressed
apoptosis, inflammation responses and collagen degradation of
LPS-stimulated C28/I2 cells. Therefore, exosome-mediated PVT1
regulated LPS-induced osteoarthritis progression by modulating the
HMGB1/TLR4/NF-κB pathway via miR-93-5p. The present study aids to
improve understanding of the role of PVT1 in osteoarthritis,
providing a novel possible target for osteoarthritis treatment.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author on reasonable request.
Authors' contributions
YM and SQ conceived and designed the experiments and
performed the experiments. LS wrote the paper and performed the
experiments. JZ conceived and designed the experiments and reviewed
drafts of the paper. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was authorized by the Ethics
Committee of Weihai Municipal Hospital. All patients with
osteoarthritis and the healthy subjects who participated in the
present study signed written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hussain SM, Neilly DW, Baliga S, Patil S
and Meek R: Knee osteoarthritis: A review of management options.
Scott Med J. 61:7–16. 2016.PubMed/NCBI
|
|
2
|
Kurtz SM, Ong KL, Lau E, Widmer M, Maravic
M, Gómez-Barrena E, de Pina Mde F, Manno V, Torre M, Walter WL, et
al: International survey of primary and revision total knee
replacement. Int Orthop. 35:1783–1789. 2011.PubMed/NCBI
|
|
3
|
Zhen G, Wen C, Jia X, Li Y, Crane JL,
Mears SC, Askin FB, Frassica FJ, Chang W, Yao J, et al: Inhibition
of TGF-β signaling in mesenchymal stem cells of subchondral bone
attenuates osteoarthritis. Nat Med. 19:704–712. 2013.PubMed/NCBI
|
|
4
|
Taylor N: Nonsurgical management of
osteoarthritis knee pain in the older adult. Clin Geriatr Med.
33:41–51. 2017.PubMed/NCBI
|
|
5
|
Batista PJ and Chang HY: Long noncoding
RNAs: Cellular address codes in development and disease. Cell.
152:1298–1307. 2013.PubMed/NCBI
|
|
6
|
Schmitz SU, Grote P and Herrmann BG:
Mechanisms of long noncoding RNA function in development and
disease. Cell Mol Life Sci. 73:2491–2509. 2016.PubMed/NCBI
|
|
7
|
Liu Q, Hu X, Zhang X, Dai L, Duan X, Zhou
C and Ao Y: The TMSB4 pseudogene lncRNA functions as a competing
endogenous RNA to promote cartilage degradation in human
osteoarthritis. Mol Ther. 24:1726–1733. 2016.PubMed/NCBI
|
|
8
|
Zhang Y, Wang F, Chen G, He R and Yang L:
LncRNA MALAT1 promotes osteoarthritis by modulating miR-150-5p/AKT3
axis. Cell Biosci. 9:542019.PubMed/NCBI
|
|
9
|
Cao L, Wang Y, Wang Q and Huang J: LncRNA
FOXD2-AS1 regulates chondrocyte proliferation in osteoarthritis by
acting as a sponge of miR-206 to modulate CCND1 expression. Biomed
Pharmacother. 106:1220–1226. 2018.PubMed/NCBI
|
|
10
|
Cui M, You L, Ren X, Zhao W, Liao Q and
Zhao Y: Long non-coding RNA PVT1 and cancer. Biochem Biophys Res
Commun. 471:10–14. 2016.PubMed/NCBI
|
|
11
|
Huang W, Lan X, Li X, Wang D, Sun Y, Wang
Q, Gao H and Yu K: Long non-coding RNA PVT1 promote LPS-induced
septic acute kidney injury by regulating TNFα and JNK/NF-κB
pathways in HK-2 cells. Int Immunopharmacol. 47:134–140.
2017.PubMed/NCBI
|
|
12
|
Feng F, Qi Y, Dong C and Yang C: PVT1
regulates inflammation and cardiac function via the MAPK/NF-κB
pathway in a sepsis model. Exp Ther Med. 16:4471–4478.
2018.PubMed/NCBI
|
|
13
|
Li Y, Li S, Luo Y, Liu Y and Yu N: LncRNA
PVT1 regulates chondrocyte apoptosis in osteoarthritis by acting as
a sponge for miR-488-3p. DNA Cell Biol. 36:571–580. 2017.PubMed/NCBI
|
|
14
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297.
2004.PubMed/NCBI
|
|
15
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010.PubMed/NCBI
|
|
16
|
Wang X, Liao Z, Bai Z, He Y, Duan J and
Wei L: MiR-93-5p promotes cell proliferation through
down-regulating PPARGC1A in hepatocellular carcinoma cells by
bioinformatics analysis and experimental verification. Genes
(Basel). 9:512018.
|
|
17
|
Xiang Y, Liao XH, Yu CX, Yao A, Qin H, Li
JP, Hu P, Li H, Guo W, Gu CJ and Zhang TC: MiR-93-5p inhibits the
EMT of breast cancer cells via targeting MKL-1 and STAT3. Exp Cell
Res. 357:135–144. 2017.PubMed/NCBI
|
|
18
|
Zhang Y, Wei QS, Ding WB, Zhang LL, Wang
HC, Zhu YJ, He W, Chai YN and Liu YW: Increased microRNA-93-5p
inhibits osteogenic differentiation by targeting bone morphogenetic
protein-2. PLoS One. 12:e01826782017.PubMed/NCBI
|
|
19
|
Hua Q, Chen Y, Liu Y, Li M, Diao Q, Xue H,
Zeng H, Huang L and Jiang Y: Circular RNA 0039411 is involved in
neodymium oxide-induced inflammation and antiproliferation in a
human bronchial epithelial cell line via sponging miR-93-5p.
Toxicol Sci. 170:69–81. 2019.PubMed/NCBI
|
|
20
|
Xue H, Tu Y, Ma T, Wen T, Yang T, Xue L,
Cai M, Wang F and Guan M: miR-93-5p attenuates IL-1β-induced
chondrocyte apoptosis and cartilage degradation in osteoarthritis
partially by targeting TCF4. Bone. 123:129–136. 2019.PubMed/NCBI
|
|
21
|
Harding CV, Heuser JE and Stahl PD:
Exosomes: Looking back three decades and into the future. J Cell
Biol. 200:367–371. 2013.PubMed/NCBI
|
|
22
|
Valadi H, Ekström K, Bossios A, Sjöstrand
M, Lee JJ and Lötvall JO: Exosome-mediated transfer of mRNAs and
microRNAs is a novel mechanism of genetic exchange between cells.
Nat Cell Biol. 9:654–659. 2007.PubMed/NCBI
|
|
23
|
Lo Cicero A, Stahl PD and Raposo G:
Extracellular vesicles shuffling intercellular messages: For good
or for bad. Curr Opin Cell Biol. 35:69–77. 2015.PubMed/NCBI
|
|
24
|
Buzas EI, György B, Nagy G, Falus A and
Gay S: Emerging role of extracellular vesicles in inflammatory
diseases. Nat Rev Rheumatol. 10:356–364. 2014.PubMed/NCBI
|
|
25
|
Li ZL, Lv LL, Tang TT, Wang B, Feng Y,
Zhou LT, Cao JY, Tang RN, Wu M, Liu H, et al: HIF-1α inducing
exosomal microRNA-23a expression mediates the cross-talk between
tubular epithelial cells and macrophages in tubulointerstitial
inflammation. Kidney Int. 95:388–404. 2019.PubMed/NCBI
|
|
26
|
Patel NA, Moss LD, Lee JY, Tajiri N,
Acosta S, Hudson C, Parag S, Cooper DR, Borlongan CV and Bickford
PC: Long noncoding RNA MALAT1 in exosomes drives regenerative
function and modulates inflammation-linked networks following
traumatic brain injury. J Neuroinflammation. 15:2042018.PubMed/NCBI
|
|
27
|
Zhang HG, Liu C, Su K, Yu S, Zhang L,
Zhang S, Wang J, Cao X, Grizzle W and Kimberly RP: A membrane form
of TNF-alpha presented by exosomes delays T cell activation-induced
cell death. J Immunol. 176:7385–7393. 2006.PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI
|
|
29
|
Cao C, Yin C, Shou S, Wang J, Yu L, Li X
and Chai Y: Ulinastatin protects against LPS-induced acute lung
injury by attenuating TLR4/NF-κB pathway activation and reducing
inflammatory mediators. Shock. 50:595–605. 2018.PubMed/NCBI
|
|
30
|
Hare KB, Stefan Lohmander L, Kise NJ,
Risberg MA and Roos EM: Middle-aged patients with an MRI-verified
medial meniscal tear report symptoms commonly associated with knee
osteoarthritis. Acta Orthop. 88:664–669. 2017.PubMed/NCBI
|
|
31
|
Huang ZY, Stabler T, Pei FX and Kraus VB:
Both systemic and local lipopolysaccharide (LPS) burden are
associated with knee OA severity and inflammation. Osteoarthritis
Cartilage. 24:1769–1775. 2016.PubMed/NCBI
|
|
32
|
Zhao C, Wang Y, Jin H and Yu T: Knockdown
of microRNA-203 alleviates LPS-induced injury by targeting MCL-1 in
C28/I2 chondrocytes. Exp Cell Res. 359:171–178. 2017.PubMed/NCBI
|
|
33
|
Sun T, Yu J, Han L, Tian S, Xu B, Gong X,
Zhao Q and Wang Y: Knockdown of long non-coding RNA RP11-445H22.4
alleviates LPS-induced injuries by regulation of miR-301a in
osteoarthritis. Cell Physiol Biochem. 45:832–843. 2018.PubMed/NCBI
|
|
34
|
Li F, Sun J, Huang S, Su G and Pi G:
LncRNA GAS5 overexpression reverses LPS-induced inflammatory injury
and apoptosis through up-regulating KLF2 expression in ATDC5
chondrocytes. Cell Physiol Biochem. 45:1241–1251. 2018.PubMed/NCBI
|
|
35
|
Chen WK, Yu XH, Yang W, Wang C, He WS, Yan
YG, Zhang J and Wang WJ: lncRNAs: Novel players in intervertebral
disc degeneration and osteoarthritis. Cell Prolif.
50:e123132017.
|
|
36
|
Xu J and Xu Y: The lncRNA MEG3
downregulation leads to osteoarthritis progression via miR-16/SMAD7
axis. Cell Biosci. 7:692017.PubMed/NCBI
|
|
37
|
Barile L and Vassalli G: Exosomes: Therapy
delivery tools and biomarkers of diseases. Pharmacol Ther.
174:63–78. 2017.PubMed/NCBI
|
|
38
|
Zhao Y, Zhao J, Guo X, She J and Liu Y:
Long non-coding RNA PVT1, a molecular sponge for miR-149,
contributes aberrant metabolic dysfunction and inflammation in
IL-1β-simulated osteoarthritic chondrocytes. Biosci Rep.
38:BSR201805762018.PubMed/NCBI
|
|
39
|
Ding LB, Li Y, Liu GY, Li TH, Li F, Guan J
and Wang HJ: Long non-coding RNA PVT1, a molecular sponge of
miR-26b, is involved in the progression of hyperglycemia-induced
collagen degradation in human chondrocytes by targeting CTGF/TGF-β
signal ways. Innate Immun. 26:204–214. 2020.PubMed/NCBI
|
|
40
|
Liu J, Jiang M, Deng S, Lu J, Huang H,
Zhang Y, Gong P, Shen X, Ruan H, Jin M and Wang H:
miR-93-5p-containing exosomes treatment attenuates acute myocardial
infarction-induced myocardial damage. Mol Ther Nucleic Acids.
11:103–115. 2018.PubMed/NCBI
|
|
41
|
Laursen TL, Støy S, Deleuran B, Vilstrup
H, Grønbaek H and Sandahl TD: The damage-associated molecular
pattern HMGB1 is elevated in human alcoholic hepatitis, but does
not seem to be a primary driver of inflammation. APMIS.
124:741–747. 2016.PubMed/NCBI
|
|
42
|
Srikrishna G and Freeze HH: Endogenous
damage-associated molecular pattern molecules at the crossroads of
inflammation and cancer. Neoplasia. 11:615–628. 2009.PubMed/NCBI
|
|
43
|
Qin Y, Chen Y, Wang W, Wang Z, Tang G,
Zhang P, He Z, Liu Y, Dai SM and Shen Q: HMGB1-LPS complex promotes
transformation of osteoarthritis synovial fibroblasts to a
rheumatoid arthritis synovial fibroblast-like phenotype. Cell Death
Dis. 5:e10772014.PubMed/NCBI
|
|
44
|
Wang X, Guo Y, Wang C and Yu H, Yu X and
Yu H: MicroRNA-142-3p inhibits chondrocyte apoptosis and
inflammation in osteoarthritis by targeting HMGB1. Inflammation.
39:1718–1728. 2016.PubMed/NCBI
|
|
45
|
Liang S, Lv ZT, Zhang JM, Wang YT, Dong
YH, Wang ZG, Chen K, Cheng P, Yang Q, Guo FJ, et al: Necrostatin-1
attenuates trauma-induced mouse osteoarthritis and IL-1β induced
apoptosis via HMGB1/TLR4/SDF-1 in primary mouse chondrocytes. Front
Pharmacol. 9:13782018.PubMed/NCBI
|
|
46
|
Jiang Y, Zhu L, Zhang T, Lu H, Wang C, Xue
B, Xu X, Liu Y, Cai Z, Sang W, et al: BRD4 has dual effects on the
HMGB1 and NF-κB signalling pathways and is a potential therapeutic
target for osteoarthritis. Biochim Biophys Acta Mol Basis Dis.
1863:3001–3015. 2017.PubMed/NCBI
|