Introduction
Allergic diseases, including asthma, affect 30–40%
of the global population (1). The
symptoms of allergic diseases range from local mild reactions to
severe anaphylaxis. A polarized T helper type 2 (Th2) immune
response and degranulation of mast cells are characteristic in
allergic diseases (2,3). Despite the fact that the exact
mechanisms underlying these diseases have been extensively
investigated, they remain to be fully elucidated. Although allergic
diseases, such as asthma, are inheritable (4), accumulating evidence has indicated
that epigenetic factors, such as non-coding RNAs [e.g., microRNAs
(miRNAs/miRs), long non-coding RNAs (lncRNAs) or circular RNAs
(circRNAs)] may have a role in these diseases (5,6).
Alterations in the expression of several miRNAs have been reported
to be associated with the development of allergic diseases. In
allergic inflammation, miR-155 was revealed to be significantly
upregulated (7,8); it targeted the cytotoxic
T-lymphocyte-associated protein 4 mRNA in patients with atopic
dermatitis and protected group 2 innate lymphoid cells from
apoptosis, thereby promoting type 2 immunity. Furthermore,
inhibition of miR-155 resulted in dampened expression of the Th2
cytokines, interleukin (IL)-5 and IL-13 (9). Abnormal lncRNA expression has also
been observed in allergies. Zhang et al (10) profiled human CD4+ T-cell
subsets and identified a Th2-specific lncRNA, GATA3-AS1. Notably,
peripheral blood mononuclear cells and CD4+ T-cell
clones stimulated by an allergic challenge displayed increased
expression levels of GATA3-AS1.
circRNAs are single-stranded, highly conserved RNAs
that take the form of a covalently closed continuous loop (11). circRNAs are involved in the
pathogenesis of various diseases (12). Notably, circRNAs have been reported
to act as miRNA sponges through multiple miRNA response elements,
modulating the expression of parental genes (13). For example, circHIPK3 has been
shown to bind to miR-558, resulting in upregulation of its target
protein heparanase, thus suppressing metastasis and angiogenesis of
bladder cancer (14). Recently,
certain circRNAs have been demonstrated to encode peptides
(15). However, the functions of
circRNAs in allergic diseases remain unclear. In the present study,
the expression levels of aberrantly expressed circRNAs were
profiled in a murine asthma model and their possible contributions
were investigated, with the aim of providing novel insights into
the etiology and pathogenesis of asthma.
Materials and methods
Mice
A total of six female BALB/c mice (age, 6–8 weeks;
weight, 33±0.2 g) were purchased from Guangdong Medical Laboratory
Animal Center and maintained under specific pathogen-free
conditions in the animal facility of Shenzhen University (Shenzhen,
China). A total of five mice were housed per cage in a laminar air
flow cabinet, maintained at 23±2°C at a relative humidity of 55±5%
with a 12 h dark/light cycle. All mice received a standard diet and
water ad libitum. The experimental protocols of the present
study were reviewed and approved by the Animal Care and Use
Committee of Shenzhen University. The mice were divided into
control (n=3, namely N1, N2 and N3) and asthma model groups (n=3,
namely M1, M2 and M3).
Development of a murine asthma
model
A house dust mite (HDM) allergen-induced asthma
mouse model was established, as previously described (16), the method of mouse nose drops was
changed to subcutaneous injection for the consistency of the
sensitization process. Female BALB/c mice were sensitized
subcutaneously with 20 µg crude Dermatophagoides farinae
(HDM) extract (Greer Laboratories, Inc.) in 100 µl saline on days
0, 7 and 14, and were challenged intranasally with 20 µg HDM in 50
µl saline for 5 consecutive days from day 20 onwards. The control
group only received treatment with PBS. Airway hyperresponsiveness
(AHR) to inhaled methacholine was analyzed to evaluate changes in
airway resistance in response to allergen exposure and challenge.
On day 26, the mice were sacrificed using CO2 at flow
rate of 20% volume/min (17), and
murine lung tissues were dissected and analyzed for inflammation.
Mice lung tissues were dissected and RNA was isolated for circRNA
profiling. Bronchoalveolar lavage fluid (BALF) was harvested via
lung tissue lavage with ice-cold PBS. The methods for
differentiating BALF cells were previously described in detail
(18). Sections from mouse lungs
were fixed at room temperature for 12 min with 4% neutral buffered
formalin, and were then stained with hematoxylin and eosin and
periodic acid-Schiff. Blood was collected to screen for serum
antibodies against HDM allergens.
Histopathological examination of lung
tissue
The right middle lung of mice was harvested and
fixed in 4% paraformaldehyde at room temperature for 24 h. After
dehydration and paraffin embedding, glass slides were coated with
4-µm sections. Subsequently, the sections were stained with
hematoxylin and eosin (cat. no. G1005; Servicebio, Inc.) for 5 min
at room temperature and with periodic acid-Schiff reagent (cat. no.
G1008; Servicebio, Inc.) for 10 min at room temperature. The lung
tissue sections were examined at ×200 magnification using an
optical microscope (DS-Ri2; Nikon Corporation).
Measurement of serum levels of
HDM-specific IgE and IgG1
Blood samples (100 µl per mouse) from the retro
orbital venous plexus were collected using a heparinized capillary
after the mice were anesthetized by isoflurane (RWD Life Science),
and stored at room temperature for 2 h. The samples were
centrifuged (3,000 × g, 10 min, 4°C) to isolate the serum and
stored at −80°C until further analysis. The serum levels of
HDM-specific IgE and IgG1 were assayed using an ELISA kit (cat. no.
DY008; R&D Systems, Inc.) according to the manufacturer's
protocol. The 96-well ELISA plates were coated overnight at 4°C
with 100 µl of a 4 µg/ml solution of HDM in carbonate-bicarbonate
buffer (pH 9.6; Sigma-Aldrich; Merck KGaA). ELISA was performed
using the dilutions of each serum samples, and the dilutions used
were 1/4 for sIgE and 1/500 for sIgG1. ELISA was prepared using the
Reagent diluent (cat. no. DY995; R&D Systems, Inc.) and Goat
Anti-Mouse IgG1-horseradish peroxidase (HRP; 1:2,000; cat. no.
1071-05; Southern Biotech) or Goat Anti-Mouse IgE-HRP (1:2,000;
cat. no. 1110-05; Southern Biotech). The other steps were performed
following the manufacturer's instructions.
Flow cytometry
For detection of eosinophils, red blood cells were
lysed using ammonium chloride lysis buffer (Sigma-Aldrich; Merck
KGaA). Absolute counting beads (cat. no. C36950; Invitrogen; Thermo
Fisher Scientific) were mixed with the BALF cells and assessed via
flow cytometry. The numbers of BALF cells were calculated by
comparing the ratio of beads events to cell events according to the
manufacturer's protocol. The BALF cells were stained with
antibodies against CD3-FITC (cat. no. 100203; Biolegend, Inc.),
B220-FITC (cat. no. 103205, Biolegend, Inc.), CCR3-PE (cat. no.
144505; Biolegend, Inc.), CD11c-PerCP/Cyanine5.5 (cat. no. 117327;
Biolegend, Inc.) and MHCII-APC (cat. no. 116417; BD Biosciences)
for 30 min at 4°C. The cells were stained for neutrophils,
mononuclear cells and lymphocytes (data not shown), and eosinophils
using a CytoFLEX flow cytometer (Beckman Coulter, Inc.), and data
collected were analyzed with FlowJo V10 software (FlowJo, LLC).
Granulocytes were recognized as non-autofluorescent highly granular
(SSChi) cells, and within this gate, eosinophils were
defined as cells expressing the eotaxin receptor CCR3 and with low
expression of MHCII, B220 and CD3. Neutrophils had a similar
scatter profile as eosinophils but lacked CCR3 expression.
Lymphocytes were identified as FSClo/SSClo
and expressing CD3 or B220. Mononuclear cells expressed high levels
of MHCII and CD11c.
Non-invasive measurement of airway
function
Airway responsiveness was measured 24 h after the
final exposure to HDM extract by recording respiratory pressure
curves using barometric unrestrained whole-body plethysmography
(Buxco; emka TECHNOLOGIES S.A.S) in response to inhaled
methacholine (acetyl-β-methylcholine chloride; Sigma-Aldrich; Merck
KGaA) in conscious unrestrained mice. Airway responsiveness was
expressed as enhanced pause (Penh), as described previously
(19). Briefly, mice were placed
in a whole-body chamber and basal readings were obtained and
averaged for a 3-min period. Subsequently, increasing doses of
methacholine (0–100 mg/ml) were aerosolized for 3 min, and readings
were taken and averaged for 3 min after each nebulization.
RNA extraction and library
construction
Total RNA was extracted from the tissues using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), according to the manufacturer's instructions. Total RNA was
qualified and quantified using a NanoDrop (NanoDrop; Thermo Fisher
Scientific, Inc.) and Agilent 2100 bioanalyzer (Agilent
Technologies, Inc.). Subsequently, total RNA was treated with DNase
I, which degraded double-stranded and single-stranded DNA.
Ribosomal RNA was removed using the Ribo-off rRNA Depletion kit
(Vazyme Biotech Co., Ltd.) and linear RNA was removed using RNase R
(Epicentre; Illumina, Inc.). Purification was performed using
Agencourt RNAClean XP magnetic beads (Invitrogen; Thermo Fisher
Scientific, Inc.). All steps were performed according to the
manufacturer's protocols. The distribution of the fragment sizes
was determined using the Agilent 2100 bioanalyzer and quantitated
using a BMG microplate reader (Omega Bio-Tek, Inc.). Finally, the
libraries were sequenced on the BGISEQ-500 (BGI Group). Raw reads
were filtered out for low quality, linker contamination and
excessively high levels of unknown base N (SOAPnuke software
v1.5.2; http://github.com/BGI-flexlab/SOAPnuke), and then
clean reads were aligned to the reference genome (Mus_musculus,
UCSC_mm9; ftp://hgdownload.soe.ucsc.edu/goldenPath/mm9/).
Differentially expressed circRNAs
circRNAs were detected and identified using CIRI
(20,21) and Find_circ (22,23),
and the results of the two types of software were integrated based
on the circRNA start and stop positions (combining circRNAs with
start and stop positions within the first and last 10 bases into
one class). The expression of circRNAs was calculated based on the
number of junction reads aligned to the two ends of the circRNA.
Since two softwares, CIRI (BWA-MEM genome alignment algorithm) and
Find_circ (Bowtie2 genome alignment algorithm), were used in the
profiling, the final numbers of junction reads were the mean of the
two results. Junction reads per billion mapped reads was used to
homogenize each sample, and the EBseq (24) algorithm was used to detect
differentially expressed circRNAs between control and model groups.
The circRNAs included in circBase (25) were provided with corresponding ID
numbers in the present study. circRNAs exhibiting fold changes ≥2.0
with P-values ≤0.05 were classified as significantly differentially
expressed circRNAs. The differentially expressed genes (DEGs)
obtained from all groups were analyzed by bidirectional clustering
using the Pheatmap package (v1.0.12; rdocumentation.org/packages/pheatmap/versions/1.0.12)
and presented as heatmaps.
Validation of differentially expressed
circRNAs and mRNAs
Total RNA from lung tissues was extracted using
TRIzol® reagent. Total RNA (1 µg) was treated with RNase
R for circRNA RT-qPCR, or without RNase R for mRNA RT-qPCR, and
first-strand cDNA was synthesized using the First Strand cDNA
Synthesis kit for reverse transcription-quantitative PCR (RT-qPCR)
analysis (Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer's instructions. First-strand cDNA was synthesized
using 8 µl First Strand Synthesis Act D Mix and SuperScript II
Reverse Transcriptase at 40°C for 1 h, 90°C for 5 min and 0°C for 5
min. qPCR was performed on an ABI ViiA7 real-time PCR detection
system (Applied Biosystems; Thermo Fisher Scientific, Inc.) using
QuantiFast SYBR Green PCR kit (Qiagen, Inc.). The PCR mixture was
incubated under the following conditions: One cycle at 95°C for 2
min, followed by 40 cycles at 94°C for 10 sec, 60°C for 10 sec and
72°C for 40 sec, and a final extension step of 72°C for 5 min. Data
were analyzed using the 2−ΔΔCq method (26) and expressed as fold change relative
to untreated controls. The circRNA levels were normalized to the
internal control gene GAPDH. Differentially expressed circRNAs were
identified by unpaired two-tailed Student's t-test between two
groups and fold change between two samples. All the reactions were
assessed with three biological replicates. The primers were
designed by ABI Primer Express Software v2.0 (Applied Biosystems;
Thermo Fisher Scientific, Inc.) and synthesized by BGI Group. The
primer sequences used in the present study are listed in Table I.
 | Table I.Primers used in reverse
transcription-quantitative PCR analysis. |
Table I.
Primers used in reverse
transcription-quantitative PCR analysis.
| Primer | Sequence (5′ to
3′) |
|---|
|
mmu_circ_0000909 | Forward,
GCTGAGGAAGTGGTGGATGA |
|
| Reverse,
TGTGATGAGTACGCGCATGT |
|
mmu_circ_0000629 | Forward,
TCTGTGGCAGGCTCTATGGC |
|
| Reverse,
GCTTTGCTCCCGAAACGACT |
|
mmu_circ_0000455 | Forward,
TAAGAAAATGCCTCGCACGA |
|
| Reverse,
GCAGGTTACAAAAGGCTGGC |
|
mmu_circ_0001454 | Forward,
GGCAACCACCGAAGCAGTAA |
|
| Reverse,
CAAGGCAGAGAGGTGGCAGA |
|
mmu_circ_0000723 | Forward,
CACCTCGAGTCACTGTCGCA |
|
| Reverse,
TCCCGGTTAATTCAGGAGCC |
|
mmu_circ_0001389 | Forward,
GGCATTGCAGAGGACCTCAA |
|
| Reverse,
AGGAGGACGCACTGCTTGTG |
| mmu_IL4 | Forward,
GGTCTCAACCCCCAGCTAGT |
|
| Reverse,
GCCGATGATCTCTCTCAAGTGAT |
| mmu_IL5 | Forward,
CTCTGTTGACAAGCAATGAGACG |
|
| Reverse,
TCTTCAGTATGTCTAGCCCCTG |
| mmu_IL13 | Forward,
CCTGGCTCTTGCTTGCCTT |
|
| Reverse,
GGTCTTGTGTGATGTTGCTCA |
| mmu_GAPDH | Forward,
AGGTCGGTGTGAACGGATTTG |
|
| Reverse,
TGTAGACCATGTAGTTGAGGTCA |
Gene Ontology (GO) and Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathway analyses
GO was divided into three major functional
categories: Molecular function, cellular component and biological
process. According to the results of the DEGs, the clusterProfiler
package (version 3.5.0; bioconductor.org/packages/release/bioc/html/clusterProfiler.html)
in R software (version 3.6.0; cran.r-project.org) was used for GO and KEGG pathway
enrichment analysis Subsequently, the P-value underwent false
discovery rate (FDR) correction for q-value, and the pathways with
q≤0.01 were considered as being significantly enriched. The
directed acyclic graph (DAG) was a graphical display of the GO
enrichment analysis results of genes from different circRNA
sources.
Construction of a circRNA regulatory
network
The targeted associations between mouse miRNAs and
circRNAs were predicted using the miRanda (27) algorithm. The circRNA-miRNA
interaction network was visualized using Cytoscape 3.7.1 software
(28).
Statistical analysis
Data are expressed as mean ± SEM. Statistical
significance for normally distributed samples was assessed using an
independent two-tailed Student's t-test or one-way ANOVA followed
by the Bonferroni correction for multiple comparisons. All analyses
were performed with GraphPad Prism 7 software (GraphPad Software,
Inc.). P≤0.05 was considered to indicate a statistically
significant difference.
Results
Establishment of an asthma model
A mouse model of asthma was established as
aforementioned (Fig. 1A). Crude
HDM challenge induced AHR (Fig.
1B), and increased titers of HDM-specific IgE and IgG1
(Fig. 1C). In addition,
recruitment of inflammatory cells to the lungs was observed in
response to HDM challenge, with dense bronchial cell infiltrates
(Fig. 1D). Total and differential
cell counts of BALF samples indicated an increased number of total
inflammatory cells, with significant eosinophil recruitment to the
lungs compared with that in the normal group (Fig. 1E). Lung tissue exposed to HDM also
exhibited higher mRNA expression levels of IL-4, IL5 and IL-13
(Fig. 1F). These results indicated
that HDM induced evident Th2 type pulmonary inflammation.
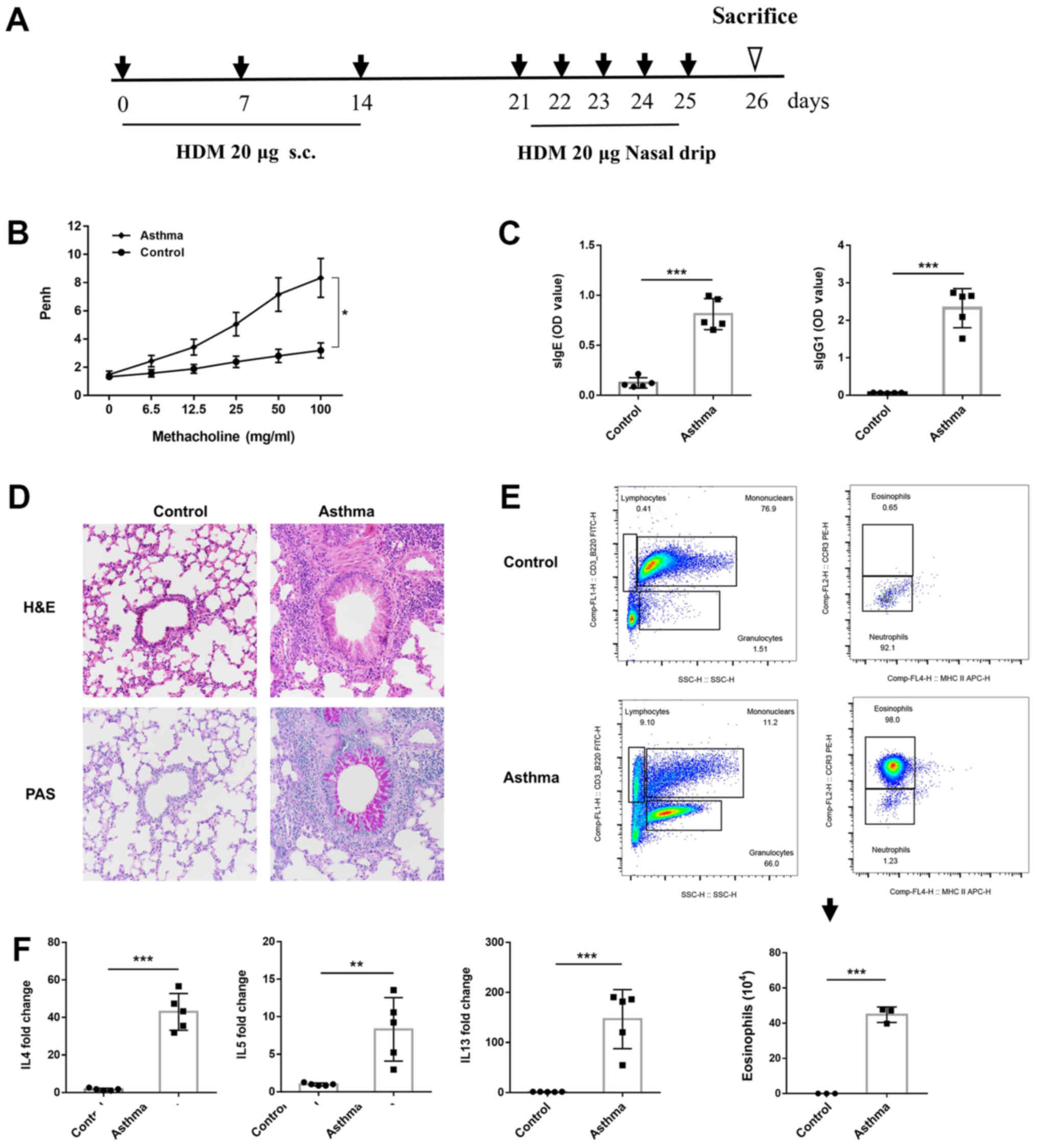 | Figure 1.Establishment of a mouse model of HDM
allergen-induced asthma. (A) Protocol for the generation of a
murine asthma model. (B) AHR to inhaled methacholine (6.25–100
mg/ml). (C) ELISA of HDM-specific IgE and IgG1 in mouse serum. (D)
Lung tissue histopathology (magnification, ×200). Paraffin-embedded
tissue sections of the lung tissue were stained with H&E (left
panel) and PAS (right panel). (E) Gating strategies for the flow
cytometric analysis of BALF cells of mice challenged with PBS
(control, upper panel) and HDM (asthma, lower panel). Quantitation
of BALF eosinophils counts as indicated by lower panel histogram.
(F) Reverse transcription-quantitative PCR for mRNA expression
levels of Th2 cytokines. Data are presented as mean ± SEM
(n=3-5/group). *P<0.05, **P<0.01, ***P<0.001. HDM, house
dust mite; AHR, airway hyperresponsiveness; H&E, hematoxylin
and eosin; PAS, periodic acid-Schiff; BALF, bronchoalveolar lavage
fluid. |
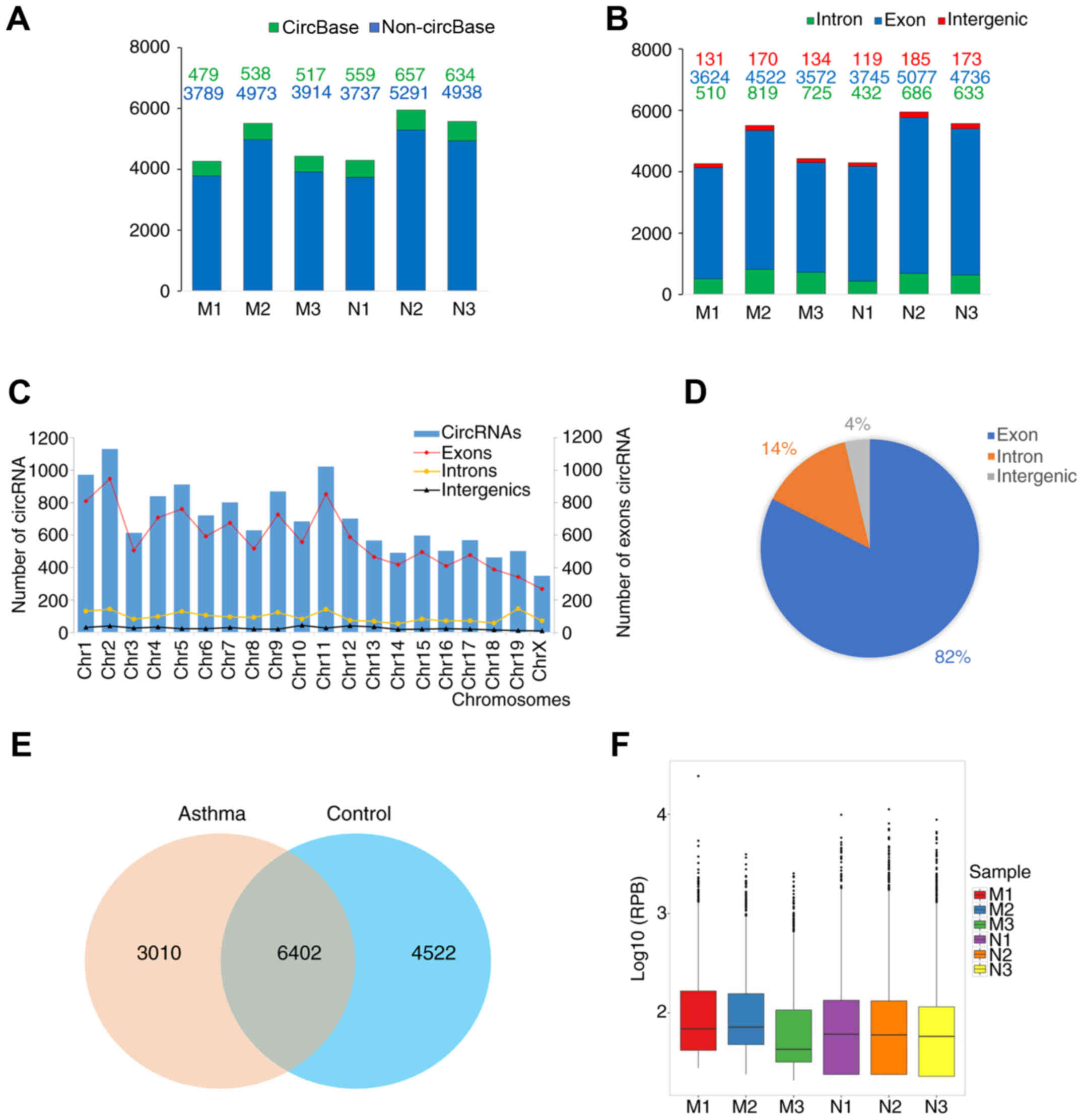
circRNA expression profile
As aforementioned, circRNA sequencing libraries were
constructed and sequenced. The total number of circRNAs annotated
or unannotated in circBase identified in each sample is shown in
Fig. 2A. The circRNA types
(exonic, intronic and intergenic) of each sample are shown in
Fig. 2B, and the types of circRNAs
at each chromosome position in the two groups are shown in Fig. 2C. In total, 10,924 circRNAs were
identified in the control group, whereas 9,412 circRNAs were
detected in the asthma model group, among which 6,402 circRNAs were
expressed in both groups (Fig.
2E). The box plot in Fig. 2F
shows the distribution and dispersion of circRNA expression levels
in each sample. Furthermore, the number of circRNAs distributed on
each chromosome was examined and it was observed that the majority
of circRNAs (82%) were derived from exons (Fig. 2D).
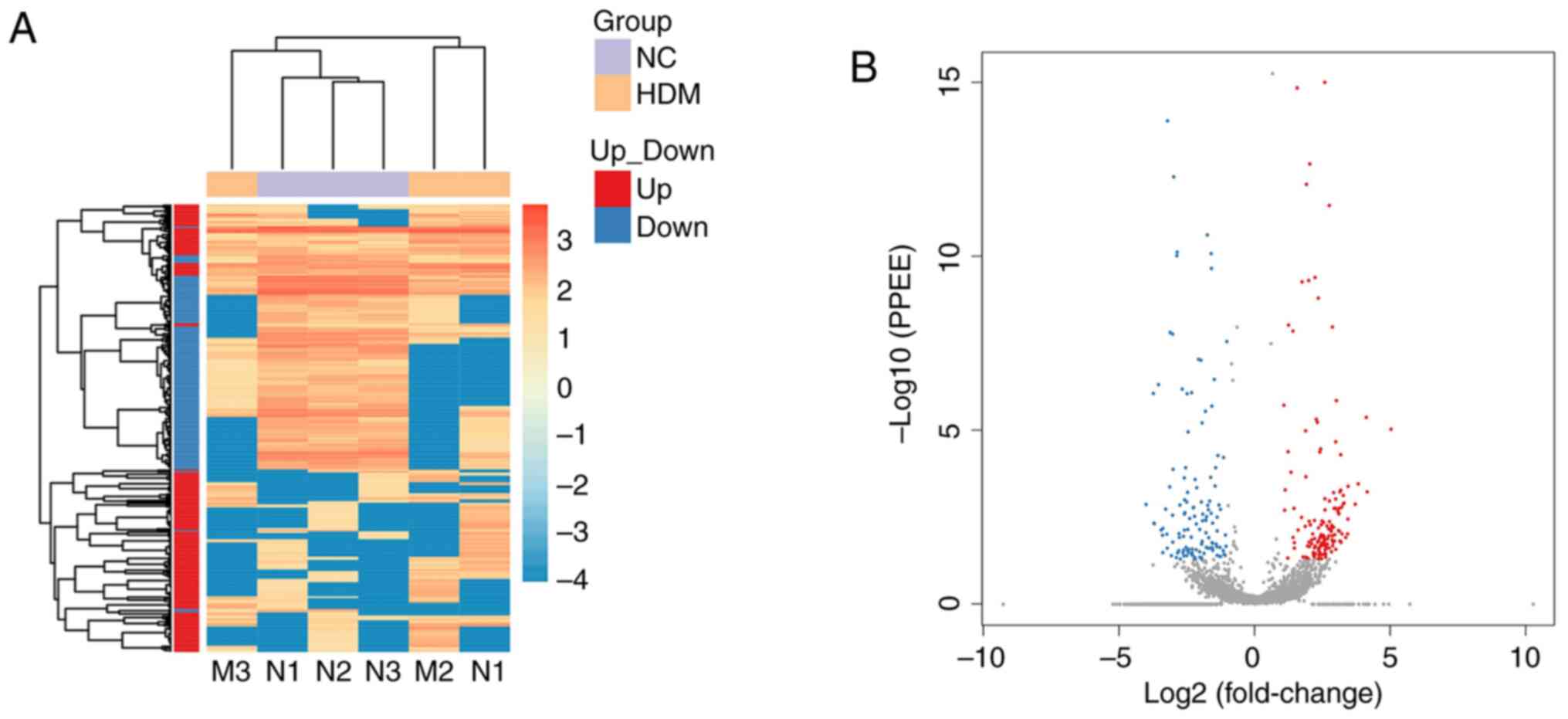
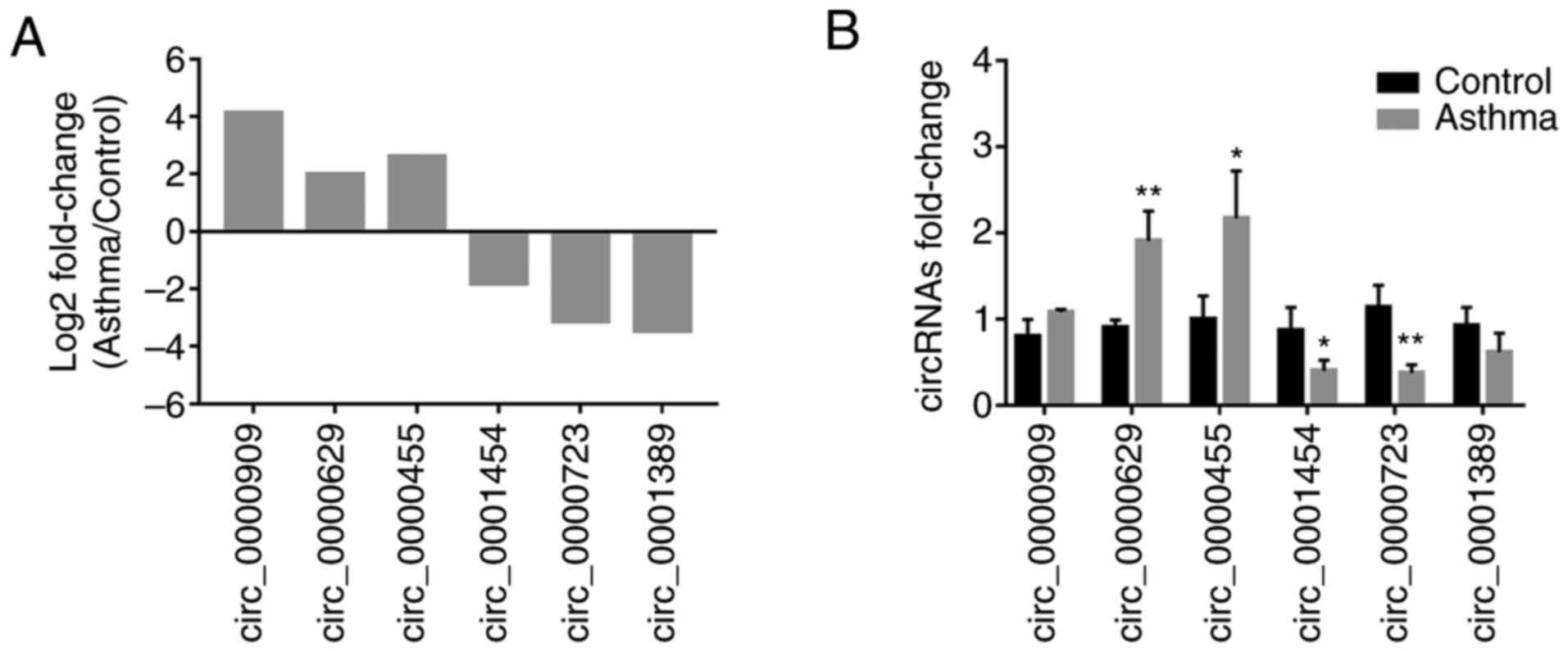
Analysis and validation of
differentially expressed circRNAs
A total of 282 circRNAs that were significantly
differentially expressed in the lung tissue between the control and
asthma model groups were identified, among which 152 were
upregulated (red spots) and 130 were downregulated (blue spots), as
shown in the heatmap (Fig. 3A) and
the volcano plot (Fig. 3B). To
verify sequencing expression data, six differentially expressed
circRNAs (according to the results of miRanda and the fold change
of circRNAs expression), including three upregulated (circ_0000909,
circ_0000629 and circ_0000455) and three downregulated
(circ_0001454, circ_0000723 and circ_0001389) circRNAs were
selected for RT-qPCR validation (Fig.
4A). The results revealed that the relative expression levels
of circ_0000629 and circ_0000455 in the asthma group were
significantly increased compared with those in the control group,
whereas the expression levels of circ_0000454 and circ_0000723 were
significantly decreased (Fig. 4B).
These results confirmed that the four circRNAs were differentially
expressed.
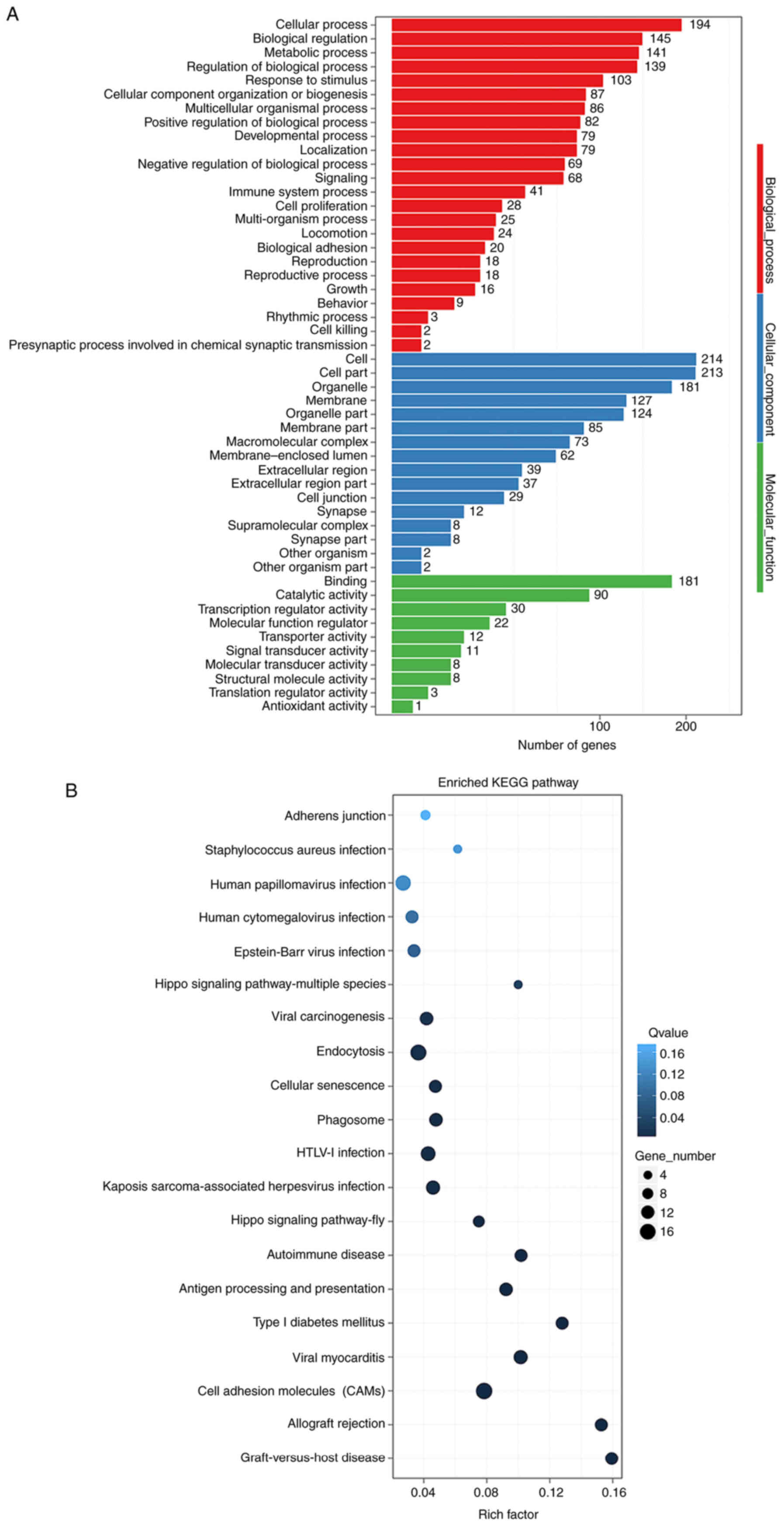
GO and KEGG pathway analyses of
differentially expressed circRNAs
GO functional classification and enrichment analysis
was conducted on genes derived from circRNAs. Deeper classification
and enrichment analysis of the three major functions, namely
biological process, cellular component and molecular function, was
performed. The results of GO functional classification were shown
in Fig. 5A. GO was a
classification system of function of gene, whereas DAG was a
graphical display of GO enrichment data. The DAG of biological
processed was shown in Fig. S1A.
The DAGs of cellular components and molecular functions were shown
in Fig. S1B and C. The results of
KEGG pathway enrichment analysis were shown in Fig. S1D demonstrating all the functions,
and the results of some specific pathway enrichments were shown in
Fig. 5B. The enriched GO terms
were mainly associated with ‘cellular process’, ‘biological
regulation’, ‘metabolic process’, ‘regulation of biological
process’, and ‘cellular component organization or biogenesis’
(Fig. 5A). Genes associated with
differentially expressed circRNAs were mainly enriched in
‘autoimmune disease’, ‘cell adhesion molecules (CAMs)’,
‘endocytosis’ and ‘lipid metabolism’ pathways, among others
(Figs. 5B and S1D).
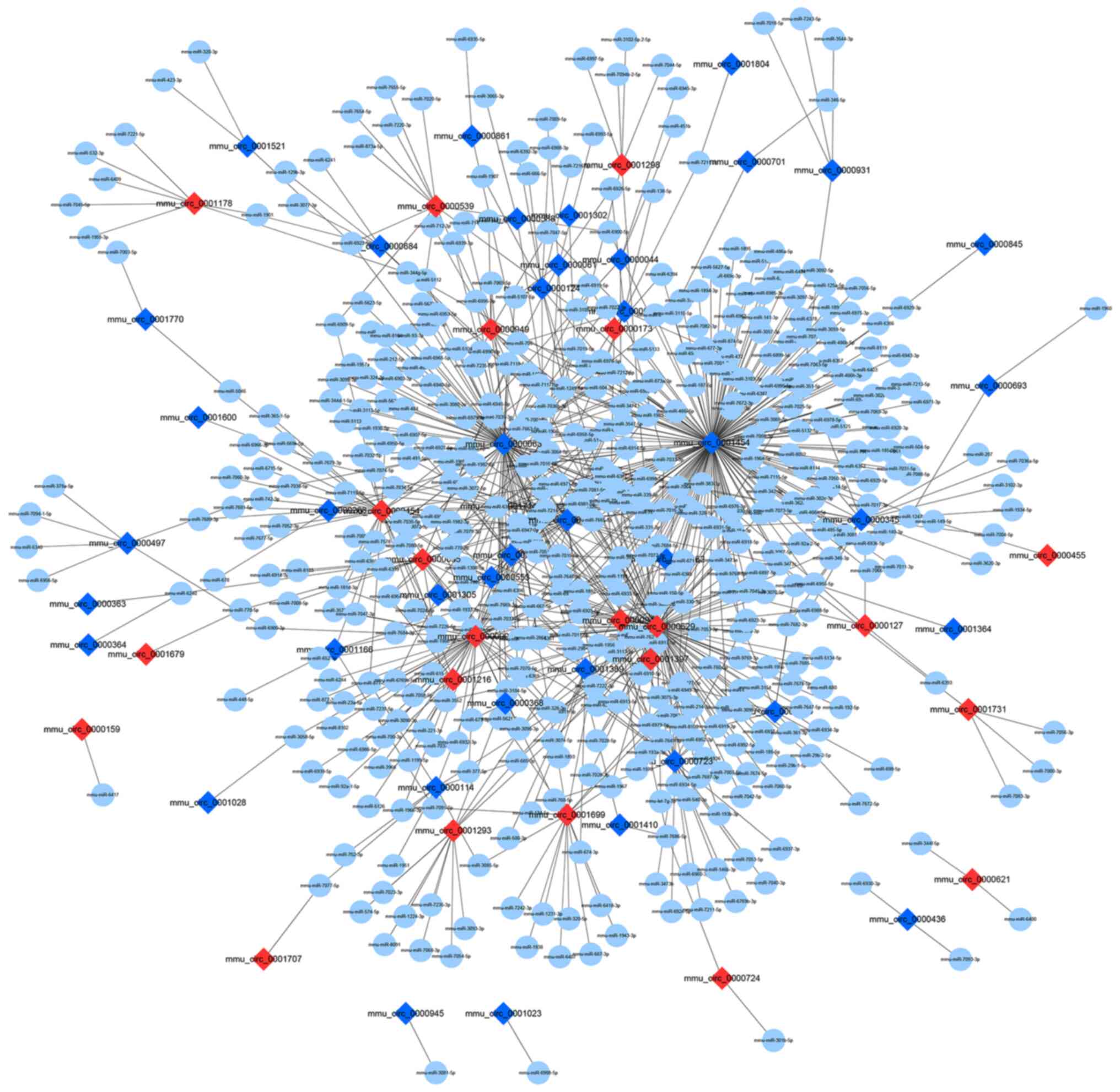
circRNA-miRNA regulatory network
construction
circRNAs comprise a large class of
post-transcriptional regulators that have been proven to act like
sponges for miRNAs, thereby indirectly regulating mRNA expression
(13,14). Based on the calculations of
miRanda, the results of differentially expressed circRNAs in
circBase combined with miRNAs in miRBase were provided. In the
constructed potential circRNA-miRNA network, 491 miRNAs interacted
with 63 differentially expressed circRNAs (23 upregulated and 40
downregulated) (Fig. 6). The
circRNA-miRNA regulatory network indicated that two of the
downregulated circRNAs (circ_0001454 and circ_0000723) targeted
miR-146b and miR-214, and two of these upregulated circRNAs
(circ_0000455 and circ_0000629) could target miR-29b and
miR-15a.
Discussion
In order to construct a reliable animal model for
circRNA profiling in allergic diseases, HDM, the most common and
causative allergen source, was selected (29). Notably, the model group developed
apparent hypersensitivities upon challenge with methacholine. A
typical Th2 type immune response was observed, with elevated
expression of type 2 cytokines, including IL-4, IL-5 and IL-13, as
well as IgE and IgG1 antibody titers. Pathology analysis
demonstrated that the lung tissues were extensively infiltrated by
inflammatory cells, such as eosinophils, which are considered
indicators of allergic inflammation (30). These findings confirmed that a
murine model of HDM-induced asthma was successfully established.
AHR was measured by Penh, the validity of which as an index of
airflow limitation was once considered controversial. Kirschvink
et al (31) argued that
although Penh was able to detect changes induced by pulmonary
cholinergic stimulation, it poorly detected tracheal calibre
modifications in rats. However, Nakaya et al (32) demonstrated that Penh could be a
useful noninvasive indicator for studying nasal hypersensitivity in
murine allergic rhinitis. In addition, a recent study indicated
that the non-invasive Penh system was more accurate compared with
the invasive ventilated lung resistance method, as ventilation
induced an additional cell influx into the airways (33). It was therefore suggested that Penh
could be used to evaluate hypersensitivity in asthma models.
In the present study, the majority of circRNAs (82%)
were derived from exons of protein-coding genes, whereas the others
originated from intronic or intergenic genomic regions. A total of
282 circRNAs (152 upregulated and 130 downregulated) were
differentially expressed in the murine allergy model group compared
with in the control group. KEGG pathway analysis revealed that the
differentially expressed circRNAs were strongly associated with
‘cellular process’ and ‘biological regulation’. The analyses
indicated that circRNAs performed critical functions in asthma, and
may act through endocytosis, or signaling pathways that involve
cell adhesion or lipid metabolism. Cell adhesion molecules have
been closely implicated in various immunological disorders,
including asthma, and may serve a key role in their pathogenesis by
upregulating the function of eosinophils (30,34).
Several clinical studies have established that macrophage
phagocytosis is disrupted in asthma (35,36),
and this disruption is more pronounced in more severe forms of
asthma (37). In addition, altered
sphingolipid metabolism has been reported to be associated with
asthma phenotype in patients allergic to HDMs (38).
Previous studies demonstrated that circRNAs are
highly evolutionarily conserved between humans and mice (39,40),
and some may function as miRNA sponges. In the present study, four
differentially expressed circRNAs were validated by qPCR, of which
two were significantly upregulated and two were significantly
downregulated. The regulatory network of circRNA and miRNA
indicated that two of these upregulated circRNAs (circ_0000455 and
circ_0000629) could target miR-29b and miR-15a, respectively, which
were previously reported to be negatively correlated with the
occurrence of allergic reactions (41,42).
The expression levels of inducible co-stimulator, a target gene of
miR-29b, were also previously shown to be elevated in the lungs of
asthmatic mice, and promoted Th2 cytokine production and
eosinophilic inflammation (43).
Furthermore, vascular endothelial growth factor, which is a target
gene of mir-15a, was shown to be overexpressed in cases of
Th2-mediated lung inflammation, such as asthma, and induced an
asthma-like phenotype (44). By
contrast, two of the downregulated circRNAs (circ_0001454 and
circ_0000723) targeted miR-146b and miR-214, respectively, which
were previously shown to be positively associated with asthma
(45,46). In a previous study, miR-146b could
further propagate or help maintain the Th2 response by suppressing
Th1 activation (47). Runx
transcription factor, a target gene of miR-214, has been identified
as a molecular link in TGF-β-induced Foxp3 expression in T
regulatory cell differentiation and function, and suppressed IL-4
to affect the balance of Th1/Th2 (48). Therefore, these four circRNAs
appear to be ideal circRNA candidates and future studies could be
performed to investigate their association with asthma.
In conclusion, the present study identified circRNAs
that were differentially expressed between the asthma model and
control groups. The findings suggested that there may be a positive
association between the expression of circRNAs and asthma, and
provided novel insights into the etiology and pathogenesis of
asthma. Future studies may employ gain- or loss-of-function assays
to verify the functions of circRNAs as miRNA sponges and their
involvement in the development and pathogenesis of asthma, in mice
as well as in humans.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Professor Liu
(Research Center of Allergy and Immunology, Shenzhen University
School of Medicine) for technical assistance, as well as his
critical editing of the manuscript.
Funding
The present study was supported by the Shenzhen
Fundamental Research Plan (grant no. JCYJ 20170818094217688), the
Science and Technology Foundation of Guangdong Province (grant no.
2014A020212422), the National Natural Science Foundations of China
(grant no. 81273275 and 31671766) and the Discipline Construction
Project of Biochemistry and Molecular Biology from National
Development and Reform Commission (grant no. 1452).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Author's contributions
HB and LX designed experiments and performed the
statistical analysis. HB interpreted the data. HB, QZ, QL and MN
were involved in performing the experiments. ZL, PY and SC aided in
the experimental design. HB and LX prepared the manuscript. LX
given final approval of the version to be published. LX planned and
coordinated the overall research efforts. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The experimental protocols of the present study were
reviewed and approved by the Animal Care and Use Committee of
Shenzhen University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chan TF, Ji KM, Yim AK, Liu XY, Zhou JW,
Li RQ, Yang KY, Li J, Li M, Law PT, et al: The draft genome,
transcriptome, and microbiome of Dermatophagoides farinae
reveal a broad spectrum of dust mite allergens. J Allergy Clin
Immunol. 135:539–548. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Roberts G, Almqvist C, Boyle R, Crane J,
Hogan SP, Marsland B, Saglani S and Woodfolk JA: Developments in
the field of allergy in 2017 through the eyes of clinical and
experimental allergy. Clin Exp Allergy. 48:1606–1621. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Corren J and Ziegler SF: TSLP: From
allergy to cancer. Nat Immunol. 20:1603–1609. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ortiz RA and Barnes KC: Genetics of
allergic diseases. Immunol Allergy Clin North Am. 35:19–44. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gomez JL: Epigenetics in Asthma. Curr
Allergy Asthma Rep. 19:562019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dai X, Zhang S and Zaleta-Rivera K: RNA:
Interactions drive functionalities. Mol Biol Rep. 47:1413–1434.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sonkoly E, Janson P, Majuri ML, Savinko T,
Fyhrquist N, Eidsmo L, Xu N, Meisgen F, Wei T, Bradley M, et al:
miR-155 is overexpressed in patients with atopic dermatitis and
modulates T-cell proliferative responses by targeting cytotoxic T
lymphocyte-associated antigen 4. J Allergy Clin Immunol.
126:581–589.e1-e20. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Knolle MD, Chin SB, Rana BMJ, Englezakis
A, Nakagawa R, Fallon PG, Git A and McKenzie ANJ: MicroRNA-155
protects group 2 innate lymphoid cells from apoptosis to promote
type-2 immunity. Front Immunol. 9:22322018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Daniel E, Roff A, Hsu MH, Panganiban R,
Lambert K and Ishmael F: Effects of allergic stimulation and
glucocorticoids on miR-155 in CD4+ T-cells. Am J Clin
Exp Immunol. 7:57–66. 2018.PubMed/NCBI
|
|
10
|
Zhang H, Nestor CE, Zhao S, Lentini A,
Bohle B, Benson M and Wang H: Profiling of human CD4+
T-cell subsets identifies the TH2-specific noncoding RNA GATA3-AS1.
J Allergy Clin Immunol. 132:1005–1008. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang PP, Sun J and Li W: Genome-wide
profiling reveals atrial fibrillation-related circular RNAs in
atrial appendages. Gene. 728:1442862020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Vidal AF, Sandoval GT, Magalhães L, Santos
SE and Ribeiro-dos-Santos Â: Circular RNAs as a new field in gene
regulation and their implications in translational research.
Epigenomics. 8:551–562. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kristensen LS, Hansen TB, Venø MT and
Kjems J: Circular RNAs in cancer: Opportunities and challenges in
the field. Oncogene. 37:555–565. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li Y, Zheng F, Xiao X, Xie F, Tao D, Huang
C, Liu D, Wang M, Wang L, Zeng F and Jiang G: CircHIPK3 sponges
miR-558 to suppress heparanase expression in bladder cancer cells.
EMBO Rep. 18:1646–1659. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dhamija S and Menon MB: Non-coding
transcript variants of protein-coding genes-what are they good for?
RNA Biol. 15:1025–1031. 2018.PubMed/NCBI
|
|
16
|
Cates EC, Fattouh R, Wattie J, Inman MD,
Goncharova S, Coyle AJ, Gutierrez-Ramos JC and Jordana M:
Intranasal exposure of mice to house dust mite elicits allergic
airway inflammation via a GM-CSF-mediated mechanism. J Immunol.
173:6384–6392. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Valentine H, Williams WO and Maurer KJ:
Sedation or inhalant anesthesia before euthanasia with
CO2 does not reduce behavioral or physiologic signs of
pain and stress in mice. J Am Assoc Lab Anim Sci. 51:50–57.
2012.PubMed/NCBI
|
|
18
|
Van Rijt LS, Kuipers H, Vos N, Hijdra D,
Hoogsteden HC and Lambrecht BN: A rapid flow cytometric method for
determining the cellular composition of bronchoalveolar lavage
fluid cells in mouse models of asthma. J Immunol Methods.
288:111–121. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang E, Liu X, Tu W, Do DC, Yu H, Yang L,
Zhou Y, Xu D, Huang SK, Yang P, et al: Benzo(a)pyrene facilitates
dermatophagoides group 1 (Der f 1)-induced epithelial cytokine
release through aryl hydrocarbon receptor in asthma. Allergy.
74:1675–1690. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao Y, Wang J and Zhao F: CIRI: An
efficient and unbiased algorithm for de novo circular RNA
identification. Genome Biol. 16:42015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li H and Durbin R: Fast and accurate short
read alignment with Burrows-Wheeler transform. Bioinformatics.
25:1754–1760. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Langmead B and Salzberg SL: Fast
gapped-read alignment with Bowtie 2. Nat Methods. 9:357–359. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Soneson C and Delorenzi M: A comparison of
methods for differential expression analysis of RNA-seq data. BMC
Bioinformatics. 14:912013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Glažar P, Papavasileiou P and Rajewsky N:
CircBase: A database for circular RNAs. RNA. 20:1666–1670. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Enright AJ, John B, Gaul U, Tuschl T,
Sander C and Marks DS: MicroRNA targets in Drosophila.
Genome Biol. 5:R12003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Miller JD: The role of dust mites in
allergy. Clin Rev Allergy Immunol. 57:312–329. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nakagome K and Nagata M: Involvement and
possible role of eosinophils in asthma exacerbation. Front Immunol.
9:22202018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kirschvink N, Vincke G, Onclinx C, Peck MJ
and Gustin P: Comparison between pulmonary resistance and penh in
anaesthetised rats with tracheal diameter reduction and after
carbachol inhalation. J Pharmacol Toxicol Methods. 51:123–128.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nakaya M, Dohi M, Okunishi K, Nakagome K,
Tanaka R, Imamura M, Baba S, Takeuchi N, Yamamoto K and Kaga K:
Noninvasive system for evaluating allergen-induced nasal
hypersensitivity in murine allergic rhinitis. Lab Invest.
86:917–926. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Verheijden KA, Henricks PA, Redegeld FA,
Garssen J and Folkerts G: Measurement of airway function using
invasive and non-invasive methods in mild and severe models for
allergic airway inflammation in mice. Front Pharmacol. 5:1902014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kong DH, Kim YK, Kim MR, Jang JH and Lee
S: Emerging roles of vascular cell adhesion molecule-1 (VCAM-1) in
immunological disorders and cancer. Int J Mol Sci. 19:10572018.
View Article : Google Scholar
|
|
35
|
Alexis NE, Soukup J, Nierkens S and Becker
S: Association between airway hyperreactivity and bronchial
macrophage dysfunction in individuals with mild asthma. Am J
Physiol Lung Cell Mol Physiol. 280:L369–L375. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Fitzpatrick AM, Holguin F, Teague WG and
Brown LA: Alveolar macrophage phagocytosis is impaired in children
with poorly controlled asthma. J Allergy Clin Immunol.
121:1372–1378. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liang Z, Zhang Q, Thomas CM, Chana KK,
Gibeon D, Barnes PJ, Chung KF, Bhavsar PK and Donnelly LE: Impaired
macrophage phagocytosis of bacteria in severe asthma. Respir Res.
15:722014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kowal K, Żebrowska E and Chabowski A:
Altered sphingolipid metabolism is associated with asthma phenotype
in house dust mite-allergic patients. Allergy Asthma Immunol Res.
11:330–342. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rybak-Wolf A, Stottmeister C, Glazar P,
Jens M, Pino N, Giusti S, Hanan M, Behm M, Bartok O, Ashwal-Fluss
R, et al: Circular RNAs in the mammalian brain are highly abundant,
conserved, and dynamically expressed. Mol Cell. 58:870–885. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jeck WR, Sorrentino JA, Wang K, Slevin MK,
Burd CE, Liu J, Marzluff WF and Sharpless NE: Circular RNAs are
abundant, conserved, and associated with ALU repeats. RNA.
19:141–157. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yan J, Zhang X, Sun S, Yang T, Yang J, Wu
G, Qiu Y, Yin Y and Xu W: miR-29b reverses T helper 1 cells/T
helper 2 cells imbalance and alleviates airway eosinophils
recruitment in OVA-induced murine asthma by targeting inducible
co-stimulator. Int Arch Allergy Immunol. 180:182–194. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Nakano T, Inoue Y, Shimojo N, Yamaide F,
Morita Y, Arima T, Tomiita M and Kohno Y: Lower levels of
hsa-mir-15a, which decreases VEGFA, in the CD4+ T cells
of pediatric patients with asthma. J Allergy Clin Immunol.
132:1224–1227.e12. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gonzalo JA, Tian J, Delaney T, Corcoran J,
Rottman JB, Lora J, Al-garawi A, Kroczek R, Gutierrez-Ramos JC and
Coyle AJ: ICOS is critical for T helper cell-mediated lung mucosal
inflammatory responses. Nat Immunol. 2:597–604. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lee CG, Link H, Baluk P, Homer RJ,
Chapoval S, Bhandari V, Kang MJ, Cohn L, Kim YK, McDonald DM and
Elias JA: Vascular endothelial growth factor (VEGF) induces
remodeling and enhances TH2-mediated sensitization and inflammation
in the lung. Nat Med. 10:1095–1103. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Feng MJ, Shi F, Qiu C and Peng WK:
MicroRNA-181a, −146a and −146b in spleen CD4+ T
lymphocytes play proinflammatory roles in a murine model of asthma.
Int Immunopharmacol. 13:347–353. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Qiu YY, Zhang YW, Qian XF and Bian T:
miR-371, miR-138, miR-544, miR-145, and miR-214 could modulate
Th1/Th2 balance in asthma through the combinatorial regulation of
Runx3. Am J Transl Res. 9:3184–3199. 2017.PubMed/NCBI
|
|
47
|
Lu TX and Rothenberg ME: Diagnostic,
functional, and therapeutic roles of microRNA in allergic diseases.
J Allergy Clin Immunol. 132:3–13; quiz 14.2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lee SH, Jeong HM, Choi JM, Cho YC, Kim TS,
Lee KY and Kang BY: Runx3 inhibits IL-4 production in T cells via
physical interaction with NFAT. Biochem Biophys Res Commun.
381:214–217. 2009. View Article : Google Scholar : PubMed/NCBI
|