Introduction
Pancreatic ductal adenocarcinoma (PDAC) is a common
malignant tumor of the digestive tract, which is characterized by
rapid progression, high mortality and high invasiveness (1,2). It
has been reported that ~50% of patients have been confirmed to
possess metastasized cancer cells (3). Despite continuous improvements in
diagnosis and treatment, this type of cancer is almost invariably
fatal, and the 5-year survival rate of patients is ~8% (4,5). PDAC
has a high degree of malignant biological characteristics,
including invasion and metastasis, which leads to a poor prognosis.
Therefore, revealing the molecular mechanisms underlying PDAC
progression, and developing corresponding targeted therapies, is
essential to improve the outcome of patients with PDAC.
Laminin is the main component of the extracellular
matrix (ECM); it is located on the basement membrane and plays an
important role in cell differentiation, adhesion and migration, as
well as signal transduction (6,7).
Laminin-332 (LM-332) is a primary member of the laminin family. It
is composed of LNα3, LNβ3 and LNγ2 chains, which are encoded by the
LAMA3, LAMB3 and LAMC2 genes, respectively (8,9). The
three genes each express three strands, and the subsequent
formation of a heterotrimer is a key step in the formation of the
LM-332 complex. It has been demonstrated that LM-332 promotes
various cell functions, including scattering, migration,
polarization, adhesion, proliferation and apoptosis, through
adhesion plaques and hemidesmosomes formed by interacting with
integrin α3β1 and α6β4 (10,11).
In addition, these integrins interact with adhesion molecules to
affect signal transduction, which plays a key role in tumor
invasion and metastasis. These characteristics of LM-332 suggest
that it may play a key role in the cancerous transformation of
normal cells.
The epithelial-to-mesenchymal transition (EMT)
refers to the biological process in which epithelial cells are
transformed into cells with a mesenchymal phenotype (12). EMT plays a vital role in tumor cell
invasion and metastasis (13,14).
When tumor cells undergo EMT, the cell skeleton undergoes
reorganization; the epithelial cell phenotype is lost; and the
expression levels of proteins that enhance intercellular adhesion,
such as E-cadherin, desmoplakin, zonula occludens-1 and claudin,
are decreased. Simultaneously, molecular markers of interstitial
cells, such as vimentin, fibronectin, matrix metalloproteinases and
N-cadherin, exhibit increased expression levels, resulting in
decreased or lost adhesion between cells, loose connection between
cells, loss of polarity, increased migratory ability,
anti-apoptotic effects and degradation of the ECM, which are
important biological processes for tumor invasion and metastasis to
distant organs (15,16). By detecting the expression levels of
classic markers associated with EMT in malignant tumors, their
roles in the prognosis of malignant tumors can be clarified, and
key molecules can be selected to screen patient populations with a
high risk of metastasis and provide effective evidence for
predicting prognosis and metastasis, and provide personalized
treatment options.
The present study investigated the roles of LM-332
subunits (α3, β3 and γ2) in the malignant biological behavior of
PDAC. It was revealed that LAMA3, LAMB3 and LAMC2 were upregulated
in PDAC cells. It could decrease cell proliferation, invasion and
migration by inhibiting the expression of α3, β3 and γ2 subunits.
The present study identified a novel molecular mechanism of action
of LM-332 in PDAC, which may provide a novel therapeutic strategy
to block PDAC invasion and metastasis.
Materials and methods
Cell culture
The human pancreatic cancer cell lines PANC1 and
BxPC3, and the normal human pancreatic ductal epithelial cell line
HPDE6c7 (H6C7) were purchased from The Cell Bank of Type Culture
Collection of the Chinese Academy of Sciences. All cell lines were
cultured in Dulbecco's modified Eagles medium (Gibco; Thermo Fisher
Scientific, Inc.) containing 10% fetal bovine serum (FBS; Gibco;
Thermo Fisher Scientific, Inc.) in an incubator with 5%
CO2 at 37°C.
RNA interference
Small interfering (si)RNA duplexes against LAMA3
(5′-GCACAAACATTAAACAACA-3′), siRNA duplexes against LAMB3
(5′-GATGGAGGAAGATGTCAGA-3′), siRNA duplexes against LAMC2
(5′-GTCAAAGCCTGTCCTTTGA-3′) and control duplex (si-NC;
5′-UUCUCCGAACGUGUCACGU-3′) were designed by Guangzhou RiboBio Co.,
Ltd. Cells not transfected with siRNA served as the control group.
Cells were transfected with 50 nM siRNA duplex using
Lipofectamine® RNAiMAX (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions.
PANC1 cells (1×106 cells/well) were cultured in 6-well
plates and were transfected with siRNA for 48 h.
Reverse transcription-quantitative PCR
(RT-qPCR)
For the RT-qPCR analysis, total RNA was isolated
from H6C7, PANC1 and BxPC3 cells using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. cDNA was synthesized from total RNA (2 µg)
using iScript™ cDNA Synthesis Kit (Bio-Rad Laboratories, Inc.).
Amplification reactions included 2 µl Mix, 2 µl RNA and 6 µl
dH2O. The temperature protocol was as followed: 37°C for
15 min, 85°C for 5 sec and 4°C for 10 min. qPCR was performed with
an ABI PRISM 7900 Sequence Detection System (Applied Biosystems;
Thermo Fisher Scientific, Inc.) using the iTaq™ Universal
SYBR-Green Supermix (Bio-Rad Laboratories, Inc.). Amplification
reactions included 1 µl cDNA template, 0.3 µl each of the forward
and reverse primers (10 µM), 0.2 µl 50X ROX Reference Dye II
(Takara Biotechnology Co., Ltd.), and 5 µl 2X SYBR Premix
DimerEraser in a total volume of 10 µl. The following primer
sequences were used: β-actin forward, 5′-ACTCTTCCAGCCTTCCTTCC-3′
and reverse, 5′-CGTCATACTCCTGCTTGCTG-3′; LAMA3 forward,
5′-AAAGCGTATGTGGATAAATGTGG-3′ and reverse,
5′-CGGAAAGCAGGCGTAGAAA-3′; LAMB3 forward,
5′-GGCAGATGATTAGGGCAGCCGAGGAA-3′ and reverse,
5′-CGGACCTGCTGGATTAGGAGCCGTGT-3′; LAMC2 forward,
5′-TTCTACAACGATCCGCACGAC-3′ and reverse,
5′-ACACCACCTCCTCCGTCTCC-3′. Amplification of the transcripts
involved an initial denaturation at 95°C for 30 sec, followed by 40
cycles at 95°C for 5 sec, 55°C for 30 sec, and 72°C for 34 sec. The
cycle quantification (Cq) comparison method was used for relative
quantification. β-actin was used as the internal control for
normalization. All qPCR analyses were performed in triplicate.
Results were calculated using the 2−ΔΔCq method
(17).
Western blot analysis
Proteins extracted from H6C7, PANC1 and BxPC3 cells
using radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology) containing 1 mM phosphatase and
protease inhibitors. The protein concentration was quantified using
a bicinchoninic protein assay kit (Beyotime Institute of
Biotechnology), and then 20 µg protein were separated by sodium
dodecyl sulfate polyacrylamide gel electrophoresis (8 to 12% gels)
and transferred to polyvinylidene difluoride membranes (Merck
KGaA). The membrane was blocked with 5% skimmed milk powder at room
temperature for 2 h. Then, the membrane was incubated with primary
antibodies against LAMB3 (1:2,000; cat. no. LAMB3:26795-1-AP;
ProteinTech Group, Inc.) LAMC2 (1:2,000; cat. no. LAMC2:19698-1-AP;
ProteinTech Group, Inc.), E-cadherin (1:2,000; cat. no.
E-cadherin:20874-1-AP; ProteinTech Group, Inc.) vimentin (1:2,000;
cat. no. Vimentin:10366-1-AP; ProteinTech Group, Inc.) and LAMA3
(1:1,000; cat. no. ab151715; Abcam) overnight at 4°C. Primary
antibodies were detected with a HRP-conjugated goat anti-mouse IgG
(1:5,000; cat. no. A0216) or goat anti-rabbit IgG (1:5,000; cat.
no. A0218; both Beyotime Institute of Biotechnology) secondary
antibodies for 2 h at room temperature. Proteins were visualized
using an enhanced chemiluminescence kit (Merck KGaA) according to
the manufacturer's instructions. Protein bands were normalized to
α-tubulin and protein expression was semi-quantified by ImageJ
(version 1.8.0; National Institutes of Health).
Cell viability assay
A total of 3×103 PANC1 cells were seeded
in 96-well plates overnight. All cells were transfected with siRNA
for 2 days. Cell Counting Kit-8 (CCK-8) solution (Beyotime
Institute of Biotechnology) was added to each well and incubated
for 2 h according to the manufacturer's instructions. The
absorbance was measured using a BioTek® Synergy™ HT
reader (BioTek Instruments, Inc.) at 450 nm. All assays were
repeated in triplicate.
Colony formation assay
An initial density of 500 cells/well were seeded in
6-well plates, and cultured for 2 weeks. The cells were fixed with
4% paraformaldehyde for 30 min at room temperature and stained with
0.1% crystal violet for 1 h at room temperature (Sigma Aldrich;
Merck KGaA). Colonies containing >50 cells were counted using a
light microscope at ×10–20 magnification.
Cell invasion and migration
assays
The Transwell membrane (Corning, Inc.) was coated
with Matrigel for 2 h at room temperature during the invasion
measurement, and Matrigel was not used for the migration
measurement. PANC1 cells (5×104) were seeded into the
upper chambers in serum-free medium, and 600 µl medium containing
20% FBS was added to the lower chambers. After incubation for 24 h,
the cells adhering to the upper surface of the membrane were
removed. The invaded or migrated cells that had adhered to the
lower surface were stained with 0.1% crystal violet for 1 h at room
temperature and calculated by counting the number of cells in nine
random fields of view using a fluorescence microscope at ×10–20
magnification,.
Cell apoptosis analysis
After transfection of PANC1 cells for 48 h, the
cells were digested with trypsin and then washed twice with cold
phosphate buffered saline to collect the cells. Subsequently,
according to the manufacturer's instructions, using a FITC Annexin
V Apoptosis Detection Kit (Beyotime Institute of Biotechnology),
Annexin V FITC and propidium iodide were used to double stain the
cells. The percentage of early + late apoptotic cells was analyzed
by a BD FACSCanto™ II (BD Biosciences) flow cytometer using BD
FACSDiva software (version 6.1.3; BD Biosciences).
Statistical analysis
All experiments were performed in triplicate. All
values were presented as the mean ± SD. GraphPad Prism 6.02
(GraphPad Software, Inc.) was used to determine the statistical
significance of differences in the means of experimental groups. A
paired Student's t-test was used to analyze the statistical
significance between two groups of data. One-way ANOVA followed by
a Student-Newman-Keuls post hoc test were used for multiple
comparisons. P<0.05 was considered to indicate a statistical
significant difference.
Results
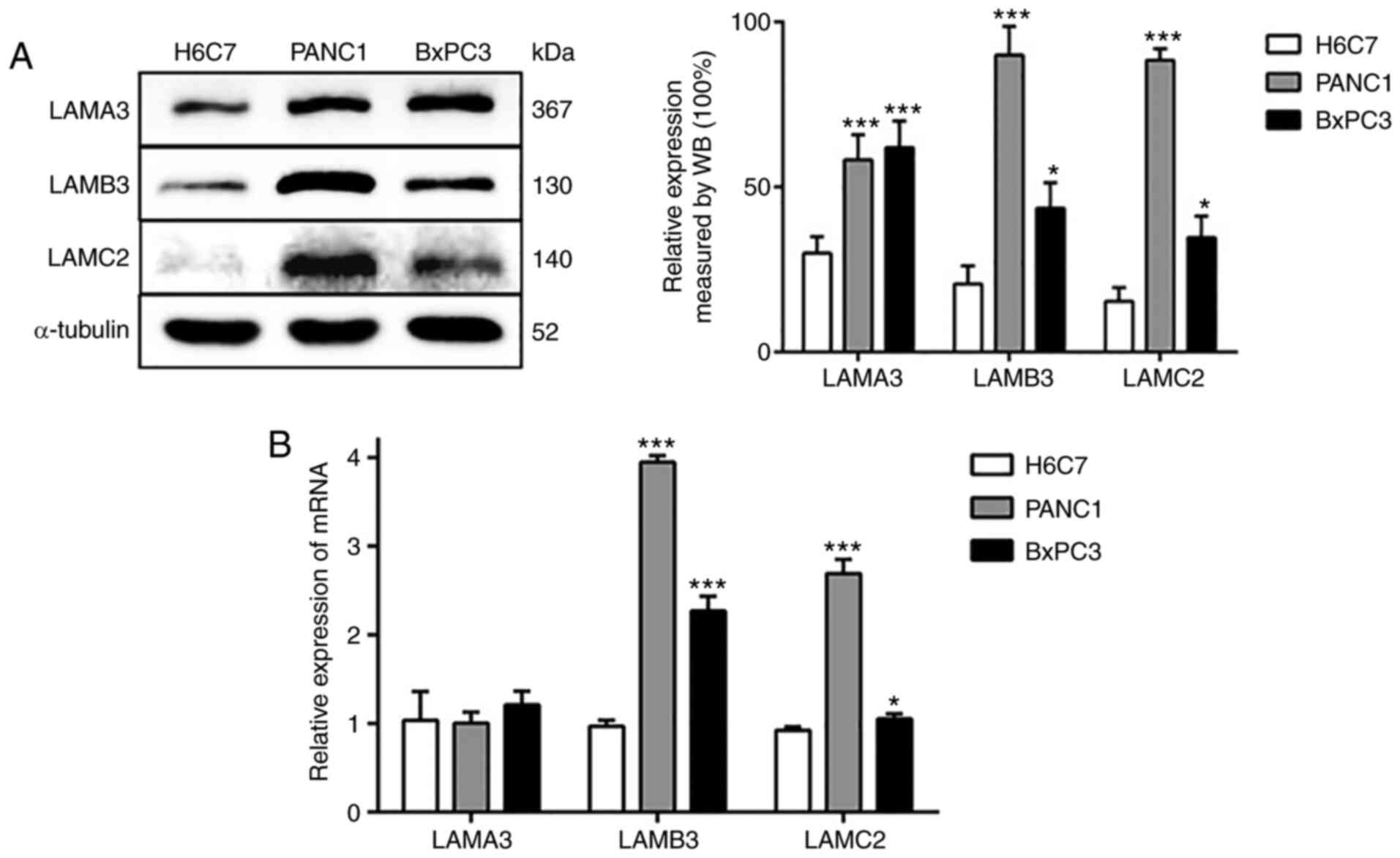
α3, β3 and γ2 chains of LM-332 are
upregulated in PDAC cells
In order to determine the expression levels of the
α3, β3 and γ2 chains of LM-332, the present study assessed LAMA3,
LAMB3 and LAMC2 expression levels in two PDAC cell lines (PANC1 and
BxPC3) and a human immortalized, non-tumorigenic pancreatic ductal
epithelial cell line (H6C7). LAMB3 and LAMC2 expression levels were
significantly increased in PANC1 and BxPC3 cells, both at the
protein and mRNA levels (Fig. 1).
In addition, the protein expression level of LAMA3 in the PDAC cell
line was increased compared with the normal pancreatic cell line.
However, there was no difference in the mRNA expression of LAMA3.
These results indicated that the expression of LAMA3, LAMB3 and
LAMC2 in the PDAC cell line is increased (particularly the level of
LAMB3) compared with the normal pancreatic cell line. Subsequently,
the present study selected PANC1 cells for the subsequent
experiments, as there were higher differences in the expression
levels of LAMB3 and LAMC2 when compared with the findings in BxPC3
cells.
Knockdown of LAMA3 inhibits
proliferation, migration and invasion, promotes apoptosis, and
regulates EMT in PANC1 cells
To further investigate the function of LAMA3, the
present study used siRNAs designed specifically against LAMA3 to
test the biological behavior of PDAC cells. The CCK-8 and colony
formation assays were used to detect cell proliferation. Transwell
migration and invasion assays were used to detect cell migratory
and invasive abilities, respectively. Flow cytometry was used to
investigate whether LAMA3 promoted PDAC proliferation by regulating
cell apoptosis. The RT-qPCR results confirmed that transfection
with si-LAMA3 led to the downregulation of LAMA3 in PANC1 cells
when compared with the findings in si-NC cells (Fig. 2A). The CCK-8 assay demonstrated that
the viability of PANC1 cells transfected with si-LAMA3 was
significantly impaired when compared with the findings in si-NC
cells (Fig. 2B). Similarly, the
results of the colony formation assay indicated that si-LAMA3
decreased the number of colonies (Fig.
2C). In addition, the migration and invasion assays
demonstrated that the migratory and invasive abilities of PANC1
cells transfected with si-LAMA3 were significantly reduced
(Fig. 2D). To be certain of the
requirement of LAMA3 for EMT, western blotting was used to evaluate
the EMT-associated protein vimentin and the epithelial marker
E-cadherin. It was revealed that si-LAMA3 significantly reduced
vimentin expression, but there was no difference in E-cadherin
expression when compared with the findings in si-NC cells (Fig. 2E). Simultaneously, in
LAMA3-knockdown cells, the number of apoptotic cells increased,
particularly those in the early stages of apoptosis (Fig. 2F). Overall, these results indicated
that LAMA3 knockdown could inhibit proliferation, migration and
invasion; promote apoptosis; and regulate EMT in PDAC cells.
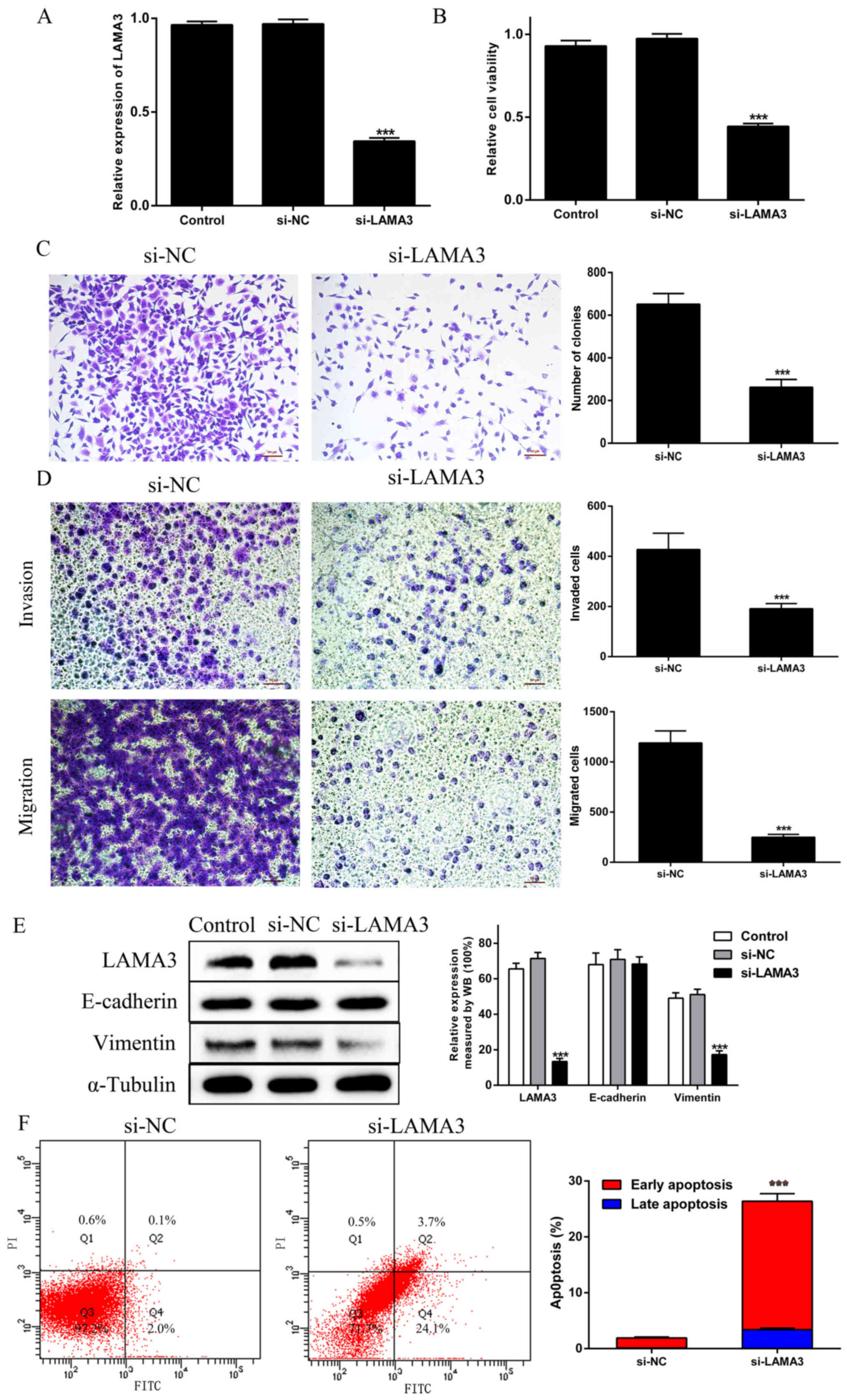 | Figure 2.Knockdown of LAMA3 inhibits PDAC cell
proliferation, migration, invasion and EMT, and promotes PDAC cell
apoptosis. (A) LAMA3 mRNA expression was evaluated via reverse
transcription-quantitative PCR in PANC-1 cells transfected with
si-LAMA3. (B) Cell proliferation was analyzed by Cell Counting
Kit-8 assays in the Control, si-NC and si-LAMA3 groups. (C) Colony
formation assays were performed in the si-NC and si-LAMA3 groups.
(D) Transwell invasion and migration assays. (E) EMT-associated
proteins E-cadherin and vimentin were examined via WB analysis in
the Control, si-NC and si-LAMA3 groups. (F) The percentage of
apoptotic cells was demonstrated by flow cytometry in the si-NC and
si-LAMA3 groups. Scale bar, 100 µm. ***P<0.01 vs. si-NC group.
PDAC, pancreatic ductal adenocarcinoma; EMT,
epithelial-to-mesenchymal transition; siRNA, small interfering RNA;
NC, negative control; LAMA3, laminin subunit α3; WB, western
blotting. |
Knockdown of LAMB3 inhibits
proliferation, migration and invasion, promotes apoptosis, and
regulates EMT in PANC1 cells
To confirm the requirement of LAMB3 in the
biological behavior of tumor cells, si-LAMB3 was transfected into
PANC1 cells. Its expression was significantly decreased when
compared with the findings in si-NC cells (Fig. 3A). Likewise, cell proliferation was
significantly reduced by si-LAMB3 (Fig.
3B and C). Similarly, si-LAMB3 decreased migration and invasion
in PANC1 cells (Fig. 3D). The
protein expression levels of E-cadherin and vimentin were assessed
via western blotting. It was noted that the expression of
E-cadherin was significantly higher, and the expression of vimentin
was significantly lower in cells transfected with si-LAMB3
(Fig. 3E). Simultaneously, in
LAMB3-knockdown cells, the number of apoptotic cells increased,
particularly those in the early stages of apoptosis (Fig. 3F). These results revealed that LAMB3
knockdown could inhibit proliferation, migration and invasion;
promote apoptosis; and regulate EMT in PDAC cells.
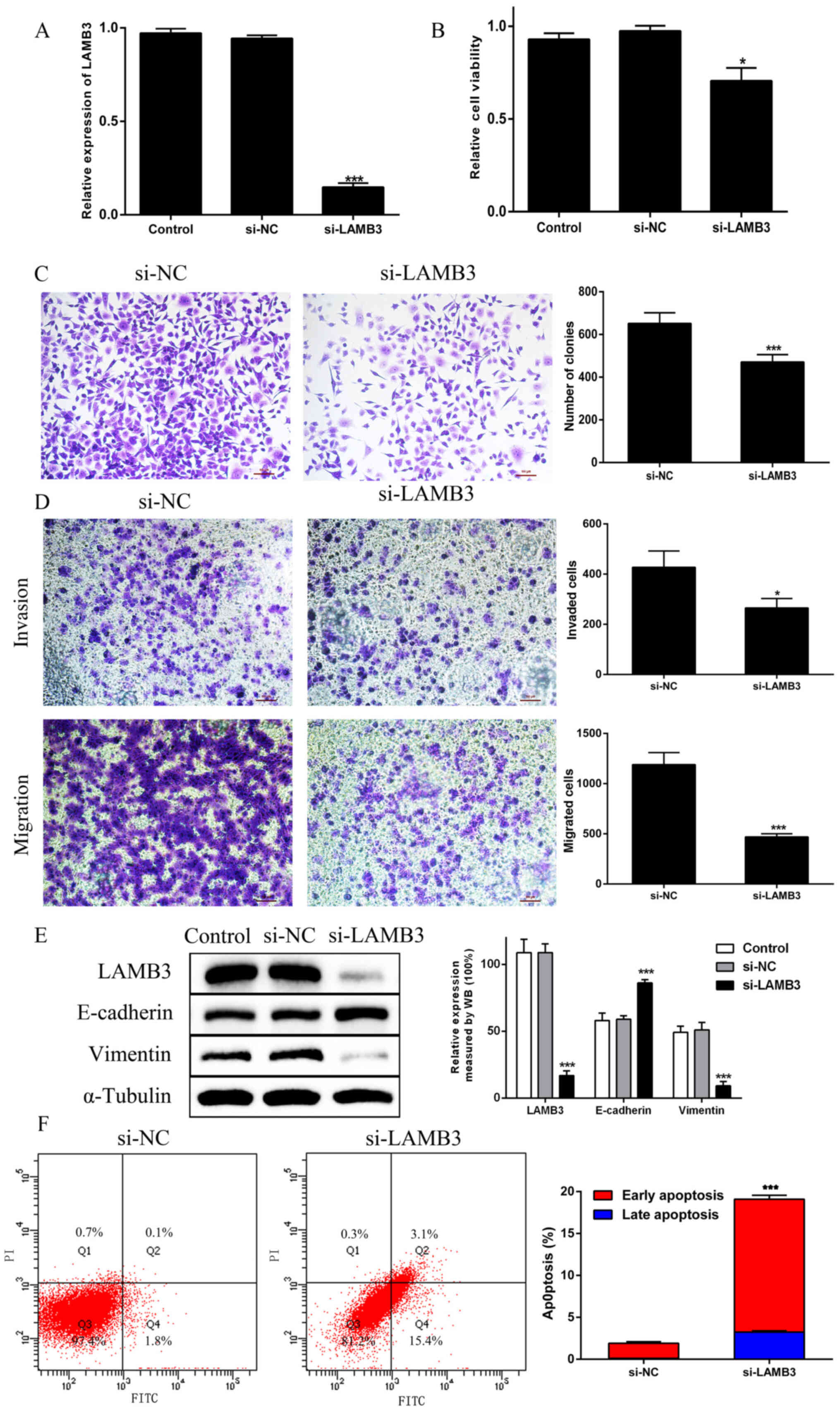 | Figure 3.Knockdown of LAMB3 inhibits PDAC cell
proliferation, migration, invasion and EMT, and promotes PDAC cell
apoptosis. (A) LAMB3 mRNA expression was evaluated via reverse
transcription-quantitative PCR in PANC-1 cells transfected with
si-LAMB3. (B) Cell proliferation was analyzed by Cell Counting
Kit-8 assays in the Control, si-NC and si-LAMB3 groups. (C) Colony
formation assays were performed in the si-NC and si-LAMB3 groups.
(D) Transwell invasion and migration assays. (E) EMT-associated
proteins E-cadherin and vimentin were examined via WB analysis in
the Control, si-NC and si-LAMB3 groups. (F) The percentage of
apoptotic cells was shown by flow cytometry in the si-NC and
si-LAMB3 groups. *P<0.05, ***P<0.01 vs. si-NC group. PDAC,
pancreatic ductal adenocarcinoma; EMT, epithelial-to-mesenchymal
transition; siRNA, small interfering RNA; NC, negative control;
LAMB3, laminin subunit β3; WB, western blotting. |
Knockdown of LAMC2 inhibits
proliferation, migration and invasion, promotes apoptosis, and
regulates EMT in PANC1 cells
The RT-qPCR results confirmed that transfection with
LAMC2 siRNA led to the downregulation of LAMC2 in PANC1 cells when
compared with the findings in si-NC cells (Fig. 4A). The CCK-8 assay indicated that
the viability of LAMC2-knockdown PANC1 cells was significantly
impaired when compared with the findings in si-NC cells (Fig. 4B). Likewise, the colony formation
results showed that si-LAMC2 decreased the colony numbers (Fig. 4C). Furthermore, the migration and
invasion assays showed that the migration and invasion of PANC1
cells with si-LAMC2 was significantly reduced (Fig. 4D). In addition, si-LAMC2
significantly decreased E-cadherin expression and increased
vimentin expression when compared with that in si-NC cells
(Fig. 4E). Furthermore, among cells
transfected with si-LAMC2, the number of apoptotic cells increased,
particularly those in the early stages of apoptosis (Fig. 4F). These results indicated that
LAMC2 knockdown could inhibit proliferation, migration and
invasion, promote apoptosis, and regulate EMT in PDAC cells.
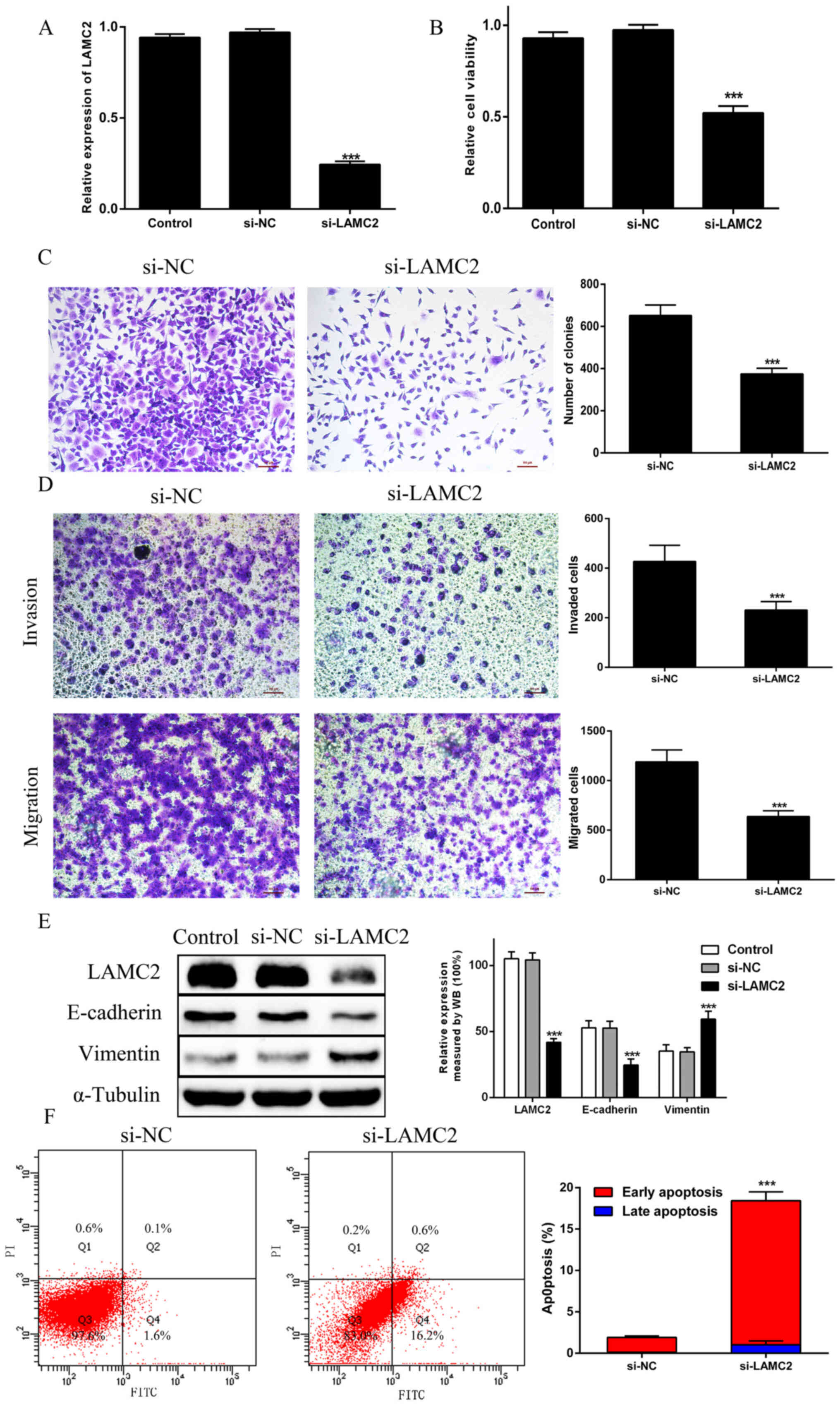 | Figure 4.Knockdown of LAMC2 inhibits PDAC cell
proliferation, migration and invasion, and promotes PDAC cell
apoptosis and EMT. (A) LAMC2 mRNA expression was evaluated via
reverse transcription-quantitative PCR in PANC-1 cells transfected
with si-LAMC2. (B) Cell proliferation was analyzed by Cell Counting
Kit-8 assays in the Control, si-NC and si-LAMC2 groups. (C) Colony
formation assays were performed in the si-NC and si-LAMC2 groups.
(D) Transwell invasion and migration assays. (E) EMT-associated
proteins E-cadherin and vimentin were examined via WB analysis in
the Control, si-NC and si-LAMC2 groups. (F) The percentage of
apoptotic cells was illustrated by flow cytometry in the si-NC and
si-LAMC2 groups. ***P<0.01 vs. si-NC group. PDAC, pancreatic
ductal adenocarcinoma; EMT, epithelial-to-mesenchymal transition;
siRNA, small interfering RNA; NC, negative control; LAMC2, laminin
subunit γ2; WB, western blotting. |
Discussion
Among patients diagnosed with advanced metastatic
disease, pancreatic cancer is one of the most malignant tumors and
deadly types of cancer, due to the difficulty in early detection
and lack of diagnostic and prognostic biomarkers (18,19).
Due to its highly malignant biological characteristics, pancreatic
cancer has a poor prognosis (20).
Although various comprehensive treatment methods have been enhanced
in recent years, the 5-year survival rate of patients remains very
low (21). It is necessary to
elucidate the invasion and metastasis mechanisms underlying
pancreatic cancer in order to improve the current treatment
situation.
A number of previous studies have demonstrated that
adhesion and extracellular matrix proteins promote the development
of various solid tumors, including pancreatic cancer (22–24).
Metastasis is a deadly feature of cancer, requiring the invasion of
cells through the basement membrane, which usually acts as a
barrier between tissue compartments. As with numerous types of
cancer of epithelial origin, LM-332 is a key element of the
basement membrane barrier (25). As
an ECM protein unique to epithelial cells, LM-332 plays a key role
in cell adhesion and migration (8,26).
LM-332 is encoded by LAMA3, LAMB3 and LAMC2, which
are associated with tumor invasiveness in various types of
malignant tumor (27). The α3 chain
of LM-332 is essential for the interaction with α3β1 integrin,
which promotes cell adhesion, migration and invasion (28). It was recently demonstrated that
upregulation of LAMA3 is associated with poor prognosis in PDAC
(17). In our previous study, the
increased expression of LAMA3 was significantly associated with
worse differentiation, deeper depth of invasion, more advanced
stage, and shorter survival time in PDAC (17). In addition, LAMA3 expression was
assessed in human PDAC cells and it was revealed that it was
significantly higher in PANC1 cells than in H6C7 cells. LAMA3
knockdown had a significant effect on the proliferation of PANC1
cells, as demonstrated by the CCK-8 and colony formation assays.
Furthermore, the invasion and migration of PANC1 cells in the
Transwell assay were significantly decreased following LAMA3
knockdown.
LAMB3 encodes two transcript variants (29); nevertheless, it seems that only the
truncated form of LAMB3 is translated into proteins (30). In prostate carcinoma cell lines,
when this variant is expressed (where it is naturally lacking),
cell migration and tumorigenicity are promoted (31,32).
It was previously indicated that overexpression of LAMB3 is
associated with clinicopathological features and decreased survival
time of patients with PDAC (20).
The results of the present study suggested that LAMB3 expression
may play a key role in the progression and prognosis of PDAC. As a
potential biomarker of cancer invasion and metastasis, LAMB3
participates in the focal adhesion pathway (33). However, to the best of our
knowledge, the function of LAMB3 in pancreatic cancer has not yet
been investigated. Similarly, the present study found that LAMB3
knockdown has a significant effect on the proliferation, invasion,
migration and EMT of PANC1 cells. Further research is required in
order to validate whether loss of LAMB3 expression is associated
with methylation at the promoter region and clinicopathological
features of poor prognosis in PADC.
Research generally suggests that overexpression of
LM-332 or one of its subunits (particularly γ2) is associated with
poor prognosis in the majority of cancer types (34). It has been reported that the
cytoplasmic expression of LAMC2 demonstrates high invasive
potential of tumors and is correlated with distant metastasis,
particularly hepatic metastasis, and poor prognosis (27). Similarly, it has recently been
demonstrated that overexpression of LAMC2 is associated with poor
prognosis in PDAC (17). However,
the presence of γ2 appears to predict an improved prognosis in
breast and colon cancer (35,36).
The present study revealed that LAMC2 knockdown had a significant
effect on the proliferation, invasion and migration of PANC1 cells.
However, si-LAMC2 significantly decreased E-cadherin expression and
increased vimentin expression, indicating that si-LAMC2 promoted
the progression of EMT. Further research is required in order to
validate the transduction pathway and underlying mechanism of
LAMC2.
EMT is a process that converts epithelial cells into
mesenchymal stem cells, with loss of polarity and adhesion, and
gain of migration and invasive properties (37). EMT can enhance the invasive,
migratory and metastatic ability of tumor cells, and it can help
cells escape apoptosis induced by certain factors in the process of
tumor development. However, tumor metastasis involves numerous
factors, such as cytokines and the internal microenvironment, and
EMT may be one of the most important factors in the overall process
(38). Tumor cells collected from
bone metastases in patients with prostate, breast and colon cancer
expressed EMT markers, and those with ovarian cancer showed EMT in
the process of metastasis (39–41).
There may be some limitations of the present study.
First, there were no investigations into the effects of LAMA3,
LAMB3 and LAMC2 overexpression. Secondly, this study did not
explore the underlying mechanism that mediates cell invasion and
migration, or EMT regulation by LM-332 in PDAC. Furthermore, there
was no investigation into the reason that the three LM-332 subunits
α3, β3 and γ2, exerted different effects on PDAC cells, nor was
there any investigation into the underlying mechanism of this
finding, which will form the main focus of future research. In our
previous research, enhanced expression of LAMA3, LAMB3 and LAMC2
were confirmed with tissue analysis (17,20).
The increased expression levels of the three subunits of LM-332
were significantly associated with worse differentiation, deeper
depth of invasion, a more advanced stage and shorter survival time
in PDAC. Zhang et al (33)
illustrated that LAMB3-mediated invasion and migration may occur
through the PI3K/Akt signaling pathway. However, the exact
underlying mechanism of LAMA3 and LAMC2 in PACD cells remains
unclear, and further studies are required to confirm this
hypothesis. We will try to explore functional analysis in a future
study. Such as, it will be necessary to confirm co-localization of
LAMA3, LAMB3 and LAMC2 in pancreatic cancer cells and to
investigate the interaction with specific integrins.
The present study revealed that LAMA3 knockdown was
closely associated with cell proliferation in PDAC. In addition,
LAMB3 knockdown regulated the EMT-associated proteins E-cadherin
and vimentin, indicating that LAMB3 is essential for EMT.
Furthermore, LAMC2 was associated with invasion and migration in
PDAC. These results may provide potential targets and novel
therapeutic strategies for the control of invasion and metastasis
in PDAC.
Acknowledgements
Not applicable.
Funding
The present work was supported by Science and
Technology Bureau of Jinhua City (grant no. 2015-3-005).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
JC and CH contributed to the conceptualization and
design of the study. JC drafted and critically revised the work. CH
performed the experiments. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Fan CS, Chen LL, Hsu TA, Chen CC, Chua KV,
Li CP and Huang TS: Endothelial-mesenchymal transition harnesses
HSP90α-secreting M2-macrophages to exacerbate pancreatic ductal
adenocarcinoma. J Hematol Oncol. 12:1382019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Erickson LA: Pancreatic ductal
adenocarcinoma. Mayo Clin Proc. 92:1461–1462. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li JT, Wang YP, Yin M and Lei QY:
Metabolism remodeling in pancreatic ductal adenocarcinoma. Cell
Stress. 3:361–368. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ayres Pereira M and Chio IC: Metastasis in
pancreatic ductal adenocarcinoma: Current standing and
methodologies. Genes (Basel). 11:112019. View Article : Google Scholar
|
|
5
|
Li B, Liu B, Zhang X, Liu H and He L:
KIF18B promotes the proliferation of pancreatic ductal
adenocarcinoma via activating the expression of CDCA8. J Cell
Physiol. 235:4227–4238. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Oh KH, Choi J, Woo JS, Baek SK, Jung KY,
Koh MJ, Kim YS and Kwon SY: Role of laminin 332 in lymph node
metastasis of papillary thyroid carcinoma. Auris Nasus Larynx.
44:729–734. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kang SG, Ha YR, Ko YH, Kang SH, Joo KJ,
Cho HY, Park HS, Kim CH, Kwon SY, Kim JJ, et al: Effect of laminin
332 on motility and invasion in bladder cancer. Kaohsiung J Med
Sci. 29:422–429. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Carpenter PM, Sivadas P, Hua SS, Xiao C,
Gutierrez AB, Ngo T and Gershon PD: Migration of breast cancer cell
lines in response to pulmonary laminin 332. Cancer Med. 6:220–234.
2017. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chiorean R, Danescu S, Virtic O, Mustafa
MB, Baican A, Lischka A, Hashimoto T, Kariya Y, Koch M and Sitaru
C: Molecular diagnosis of anti-laminin 332 (epiligrin) mucous
membrane pemphigoid. Orphanet J Rare Dis. 13:1112018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rousselle P and Beck K: Laminin 332
processing impacts cellular behavior. Cell Adhes Migr. 7:122–134.
2013. View Article : Google Scholar
|
|
11
|
Carpenter PM, Ziogas A, Markham EM,
Cantillep AS, Yan R and Anton-Culver H: Laminin 332 expression and
prognosis in breast cancer. Hum Pathol. 82:289–296. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
De Craene B and Berx G: Regulatory
networks defining EMT during cancer initiation and progression. Nat
Rev Cancer. 13:97–110. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wrighton KH: Cell migration: EMT promotes
contact inhibition of locomotion. Nat Rev Mol Cell Biol.
16:5182015. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee J, Choi JH and Joo CK: TGF-β1
regulates cell fate during epithelial-mesenchymal transition by
upregulating survivin. Cell Death Dis. 4:e7142013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kenda Suster N, Smrkolj S and Virant-Klun
I: Putative stem cells and epithelial-mesenchymal transition
revealed in sections of ovarian tumor in patients with serous
ovarian carcinoma using immunohistochemistry for vimentin and
pluripotency-related markers. J Ovarian Res. 10:112017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen J, Zhang H, Luo J, Wu X, Li X, Zhao
X, Zhou D and Yu S: Overexpression of α3, β3 and γ2 chains of
laminin-332 is associated with poor prognosis in pancreatic ductal
adenocarcinoma. Oncol Lett. 16:199–210. 2018.PubMed/NCBI
|
|
18
|
Murakami S, Shahbazian D, Surana R, Zhang
W, Chen H, Graham GT, White SM, Weiner LM and Yi C: Yes-associated
protein mediates immune reprogramming in pancreatic ductal
adenocarcinoma. Oncogene. 36:1232–1244. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pandey R, Zhou M, Islam S, Chen B, Barker
NK, Langlais P, Srivastava A, Luo M, Cooke LS, Weterings E, et al:
Carcinoembryonic antigen cell adhesion molecule 6 (CEACAM6) in
Pancreatic Ductal Adenocarcinoma (PDA): An integrative analysis of
a novel therapeutic target. Sci Rep. 9:183472019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen J, Wang W, Wei J, Zhou D, Zhao X,
Song W, Sun Q, Huang P and Zheng S: Overexpression of β3 chains of
laminin-332 is associated with clinicopathologic features and
decreased survival in patients with pancreatic adenocarcinoma. Appl
Immunohistochem Mol Morphol. 23:516–521. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gupta R, Amanam I and Chung V: Current and
future therapies for advanced pancreatic cancer. J Surg Oncol.
116:25–34. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kligys K, Wu Y, Hamill KJ, Lewandowski KT,
Hopkinson SB, Budinger GR and Jones JC: Laminin-332 and α3β1
integrin-supported migration of bronchial epithelial cells is
modulated by fibronectin. Am J Respir Cell Mol Biol. 49:731–740.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Erdogan B and Webb DJ: Cancer-associated
fibroblasts modulate growth factor signaling and extracellular
matrix remodeling to regulate tumor metastasis. Biochem Soc Trans.
45:229–236. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lugano R, Vemuri K, Yu D, Bergqvist M,
Smits A, Essand M, Johansson S, Dejana E and Dimberg A: CD93
promotes β1 integrin activation and fibronectin fibrillogenesis
during tumor angiogenesis. J Clin Invest. 128:3280–3297. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kamoshida G, Ogawa T, Oyanagi J, Sato H,
Komiya E, Higashi S, Miyazaki K and Tsuji T: Modulation of matrix
metalloproteinase-9 secretion from tumor-associated macrophage-like
cells by proteolytically processed laminin-332 (laminin-5). Clin
Exp Metastasis. 31:285–291. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kariya Y, Sato H, Katou N, Kariya Y and
Miyazaki K: Polymerized laminin-332 matrix supports rapid and tight
adhesion of keratinocytes, suppressing cell migration. PLoS One.
7:e355462012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guess CM and Quaranta V: Defining the role
of laminin-332 in carcinoma. Matrix Biol. 28:445–455. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang J, Wang H, Wang Y, Dong W, Jiang Z
and Yang G: Substrate-mediated gene transduction of LAMA3 for
promoting biological sealing between titanium surface and gingival
epithelium. Colloids Surf B Biointerfaces. 161:314–323. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fuentes I, Campos M, Repetto G, Morandé P,
Yubero MJ, Gonzalez S, Klausegger A, Schnitzhofer P, Pohla-Gubo G,
Bauer J, et al: Molecular epidemiology of junctional epidermolysis
bullosa: Discovery of novel and frequent LAMB3 mutations in Chilean
patients with diagnostic significance. Br J Dermatol.
176:1090–1092. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mayer B, Silló P, Mazán M, Pintér D,
Medvecz M, Has C, Castiglia D, Petit F, Charlesworth A, Hatvani Z,
et al: A unique LAMB3 splice-site mutation with founder effect from
the Balkans causes lethal epidermolysis bullosa in several European
countries. Br J Dermatol. 175:721–727. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Calaluce R, Bearss DJ, Barrera J, Zhao Y,
Han H, Beck SK, McDaniel K and Nagle RB: Laminin-5 beta3A
expression in LNCaP human prostate carcinoma cells increases cell
migration and tumorigenicity. Neoplasia. 6:468–479. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jung SN, Lim HS, Liu L, Chang JW, Lim YC,
Rha KS and Koo BS: LAMB3 mediates metastatic tumor behavior in
papillary thyroid cancer by regulating c-MET/Akt signals. Sci Rep.
8:27182018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang H, Pan YZ, Cheung M, Cao M, Yu C,
Chen L, Zhan L, He ZW and Sun CY: LAMB3 mediates apoptotic,
proliferative, invasive, and metastatic behaviors in pancreatic
cancer by regulating the PI3K/Akt signaling pathway. Cell Death
Dis. 10:2302019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kobayashi T, Masaki T, Nozaki E, Sugiyama
M, Nagashima F, Furuse J, Onishi H, Watanabe T and Ohkura Y:
Microarray analysis of gene expression at the tumor front of colon
cancer. Anticancer Res. 35:6577–6581. 2015.PubMed/NCBI
|
|
35
|
Moon YW, Rao G, Kim JJ, Shim HS, Park KS,
An SS, Kim B, Steeg PS, Sarfaraz S, Changwoo Lee L, et al: LAMC2
enhances the metastatic potential of lung adenocarcinoma. Cell
Death Differ. 22:1341–1352. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sato H, Higashi S and Miyazaki K:
Amino-terminal fragments of laminin γ2 chain stimulate migration of
metastatic breast cancer cells by interacting with CD44. Clin Exp
Metastasis. 32:405–415. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Crha K, Ventruba P, Žáková J, Ješeta M,
Pilka R, Vodička J and Serpa P: The role of mesenchymal-epithelial
transition in endometrial function and receptivity. Ceska Gynekol.
84:371–375. 2019.PubMed/NCBI
|
|
38
|
Kar R, Jha NK, Jha SK, Sharma A, Dholpuria
S, Asthana N, Chaurasiya K, Singh VK, Burgee S and Nand P: A
‘NOTCH’ deeper into the epithelial-to-mesenchymal transition (EMT)
program in breast cancer. Genes (Basel). 10:102019. View Article : Google Scholar
|
|
39
|
Peng YS, Syu JP, Wang SD, Pan PC and Kung
HN: BSA-bounded p-cresyl sulfate potentiates the malignancy of
bladder carcinoma by triggering cell migration and EMT through the
ROS/Src/FAK signaling pathway. Cell Biol Toxicol. 36:287–300. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hanrahan K, O'Neill A, Prencipe M, Bugler
J, Murphy L, Fabre A, Puhr M, Culig Z, Murphy K and Watson RW: The
role of epithelial-mesenchymal transition drivers ZEB1 and ZEB2 in
mediating docetaxel-resistant prostate cancer. Mol Oncol.
11:251–265. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li L, Liu J, Xue H, Li C, Liu Q, Zhou Y,
Wang T, Wang H, Qian H and Wen T: A TGF-beta-MTA1-SOX4-EZH2
signaling axis drives epithelial-mesenchymal transition in tumor
metastasis. Oncogene. 39:2125–2139. 2020. View Article : Google Scholar : PubMed/NCBI
|