Introduction
Colorectal cancer (CRC) is one of the most common
malignant cancers (1,2). Although significant improvements have
been made in the diagnosis and treatment of CRC, the 5-year
survival rate of patients with CRC remains low (3). Numerous patients are diagnosed at an
advanced stage, at which point current therapeutic strategies are
less efficient due to invasion and metastasis (2). Therefore, it is important to increase
understanding of the underlying mechanisms that may contribute to
the development of CRC.
MicroRNAs (miRNA/miRs) are a class of small,
single-stranded RNA with a length of ~22 nucleotides and lack
protein-coding ability (4,5). miRNAs regulate gene expression at a
post-translational level by binding to the 3′-untranslated region
(UTR) of target mRNAs, leading to mRNA degradation or translation
inhibition (6,7). Due to the widespread function of
miRNAs in protein-coding genes, miRNAs play critical roles in
multiple cellular processes, including cell proliferation,
differentiation and migration (8).
Notably, growing evidence has revealed that miRNAs are dysregulated
in human cancers and modulate tumor development such as cancer cell
metastasis and chemo- or radiotherapeutic sensitivity (9–13).
Aberrant expression of miRNA has previously been revealed in CRC
and possibly serves as a potential target to disrupt CRC
progression (14–16). For example, a decreased serum level
of miR-98 predicted an unfavorable clinical outcome of patients
with CRC (17). A recent study also
demonstrated that miR-361 promoted the sensitivity of CRC cells to
5-fluorouracil, indicating miR-361 as a possible drug target to
suppress tumorigenesis (18).
Notably, recent studies revealed the significantly dysregulated
expression of miR-744-5p in multiple cancers, such as
hepatocellular carcinoma, gastric cancer and pancreatic cancer
(19–21). Dysfunction of miR-744-5p has been
suggested to play suppressive roles in the development of cancers.
However, its role in CRC and the underlying mechanisms are
unknown.
Septin 2 (SEPT2) is a novel GTP-binding cytoskeletal
protein that is evolutionarily conserved and structurally
associated with the RAS oncogene (22). Numerous studies have indicated the
link between the altered expression of SEPT2 in tumorigenesis
(23–25). Downregulation of SEPT2 suppressed
the growth of hepatocellular carcinoma cells and induced cell cycle
arrest in glioma, and led to reduced cell viability (26). Overexpression of SEPT2 results in
cytokinesis failure, centrosome amplification, and the formation of
multipolar spindle bodies that frequently occur in cancer cells
(27). Although the function of
SEPT2 has been demonstrated in cancers, the involvement of SEPT2 in
CRC warrants further study.
The present study investigated the expression
pattern of miR-744-5p in CRC tissues and characterized the
functional mechanisms. It was revealed that miR-744-5p was
downregulated in CRC tissues and cell lines. Consequently,
overexpression of miR-744-5p inhibited the growth of CRC cells via
targeting SEPT2. The findings presented reveal a novel role of
miR-744-5p/SEPT2 signaling in the progression of CRC.
Materials and methods
Tissue samples
The paired CRC tissues and matched adjacent normal
tissues were obtained from 50 patients (age, 36–75 years; 22 women
and 28 men) who underwent surgery at the People's Hospital of
Yichun City between May 2011 and September 2013. Patients who were
treated with chemotherapy or radiotherapy were excluded from this
study. Tissues were frozen in the liquid immediately and stored at
−80°C until use. Written informed consent was obtained from all the
patients. The usage of the tissues was approved by the Ethics
Committee of the People's Hospital of Yichun City (approval no.
2014041566) on April 15, 2014.
Cell culture and transfection
Human CRC cell lines HCT116, SW480, SW620, RKO and
normal colon cell line CCD-18Co were obtained from the Cell Bank of
the Chinese Academy of Sciences. Cells were grown in DMEM (Thermo
Fisher Scientific, Inc.) containing 10% FBS (Thermo Fisher
Scientific, Inc.) at 37°C in a humidified atmosphere containing 5%
CO2.
The miR-744-5p mimics (5′-UGCGGGGCUAGGGCUAACAGCA-3′)
and miRNA negative control (miR-NC; 5′-GAGCUACGGUAGAGCCGGUAGC-3′)
were obtained from Guangzhou RiboBio Co., Ltd. A total of 50 nM
miRNA was transfected into cells using the
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
After transfection for 48 h, cells were harvested for subsequent
experiments.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from tissues and cells using
TRIzol® reagent. (Beyotime Institute of Biotechnology)
cDNA was synthesized with the Maxima First Strand cDNA Synthesis
kit (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The expression level of miR-744-5p was
determined using the AceQ qPCR SYBRGreen Master Mix (Vazyme Biotech
Co.) on the CFX Connect Real-Time System (Bio-Rad Laboratories
Inc). The expression of U6 was also detected for normalization. The
specific primer sequences were as follows: miR-744-5p forward,
5′-AATGCGGGGCTAGGGCTA-3′ and reverse, 5′-GTGCAGGGTCCGAGGT-3′; U6
forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′; SEPT2 forward,
5′-TAAACAGCCTATTCCTAACT-3′ and reverse, 5′-CATAAACGCCACATCTAA-3′;
HNRNPC forward, 5′-AGAACCCGGGAGTAGGAGAC-3′ and reverse,
5′-TCTCACAAAGCCGAAAACAA-3′; NFIX forward, 5′-ACTCCCCGTACTGCCTCAC-3′
and reverse, 5′-TGCAGGTTGAACCAGGTGTA-3′; c-Myc forward,
5′-CGTCCTCGGATTCTCTGCTC-3′ and reverse, 5′-GCTGGTGCATTTTCGGTTCT-3′;
PAX2 forward, 5′-CCTCGCTCCAATGGTGAGAA-3′ and reverse,
5′-TGCTGCTGGGTGAAGGTGTC-3′; GAPDH forward,
5′-ACACCCACTCCTCCACCTTT-3′ and reverse, 5′-TTACTCCTTGGAGGCCATGT-3′.
The PCR conditions were set as: Denaturation at 95°C for 5 min,
followed by 40 cycles of denaturation at 95°C for 15 sec, and
annealing and extension at 60°C for 45 sec. The relative gene
expression of miR-744-5p was analyzed using the 2−ΔΔCq
method (28).
Cell proliferation assay
The proliferation of CRC cells transfected with
miR-744-5p mimics or miRNA negative control was determined by the
Cell Counting Kit-8 (CCK-8, cat. no. C0038; Beyotime Institute of
Biotechnology) assay according to the manufacturer's instructions.
CRC cells were seeded in a 96-well plate at the density of 1,000
cells/ well and cultured overnight. Then, 10 µl CCK-8 reagent was
added into the medium after 24 h and incubated for an additional 4
h at 37°C. The absorbance at 450 nm for each well was detected by
an Epoch microplate (Bio-Tek Instruments). The assay was performed
in triplicate.
Colony formation assay
The CRC cells transfected with the miR-744-5p mimics
or miRNA negative control were seeded into the 6-well plate with
600 cells/well. Cells were grown with DMEM containing 10% FBS and
cultured for 10 days. The colonies were stained with 0.5% crystal
violet (Beyotime Institute of Biotechnology) room temperature (RT)
for 10 min after fixation with 70% ethanol (Solarbio) at RT for 10
min. The colonies were counted using a light microscope
(magnification, ×50).
Dual-luciferase reporter assay
The wild-type (WT) or mutant (MT) 3′-UTR sequences
of SEPT2 containing miR-744-5p binding sites were amplified and
inserted into the psiCHECK2 vector (Promega Corporation). CRC cells
were transfected with miR-744-5p mimics or miR-NC with
psiCHECK2-WT-SEPT2-3′UTR or psiCHECK2-MT-SEPT2-3′UTR. After
transfection with Lipofectamine® for 48 h, cells were
harvested and the luciferase activity was determined using the
Dual-Luciferase Reporter Assay (Promega Corporation) according to
the manufacturer's protocol. The activity of Renilla
luciferase was also detected for normalization.
Western blotting
Total proteins were extracted from CRC cells using
an NP-40 lysis buffer (Beyotime Institute of Biotechnology) and the
protein concentration was determined using a Bicinchoninic Acid
Protein Assay kit (Beyotime Institute of Biotechnology). A total of
20 µg proteins were separated by 15% SDS-PAGE and transferred onto
the nitrocellulose membranes (EMD Millipore). After blocking with
5% non-fat milk for 1 h at room temperature (RT), the membranes
were probed with primary antibodies against SEPT2 (1:2,000; cat.
no. 11397-1-AP; Thermo Fisher Scientific, Inc.) or GAPDH (1:3,000;
cat. no. ab9485; Abcam) at 4°C overnight. Subsequently, membranes
were incubated with HRP-conjugated goat anti-Rabbit IgG H&L
secondary antibody (1:3,000; cat. no. ab205718, Abcam) at RT for 1
h. The blots were developed by an Enhanced Chemiluminescence (ECL)
system (Thermo Fisher Scientific, Inc.). The antibodies used in
this study, including anti-SEPT2 and anti-GAPDH (both from
ProteinTech Group, Inc.) were commercially obtained.
Cell apoptosis
The cell apoptosis was detected using the FITC
Annexin V Apoptosis Detection kit (Beijing Solarbio Science &
Technology Co., Ltd.) according to the manufacturer's instructions.
Briefly, 5×105 CRC cells were transfected and harvested
after 48 h. Cells were washed with pre-cold PBS and re-suspended
with the provided binding buffer. Cells were then incubated with
Annexin V-FITC for 15 min at RT in the dark and subsequently
stained with PI for at RT for 1 min. The cell apoptosis was
determined with the FACScan flow cytometer (BD Biosciences). The
data were analyzed using FlowJo software (v10.7, http://www.flowjo.com/solutions/flowjo/downloads).
Targets prediction
The possible targets of miR-744-5p were predicted
with the miRDB (http://mirdb.org/; version 6.0) online
dataset.
Bioinformatics
Gene expression in CRC tissues and normal tissues
was evaluated against The Cancer Genome Atlas Colon Adenocarcinoma
(TCGA-COAD) database (http://ualcan.path.uab.edu/index.html) (29).
Statistical analysis
Data were presented as the mean ± standard deviation
and analyzed with SPSS 22.0 (IBM Corp.). Differences between groups
were determined with unpaired Student's t-test or one-way ANOVA
followed by Tukey's post hoc tests. The difference shown in
Figs. 1A and 4A was analyzed using paired Student's
t-test. The correlation between miR-744-5p and SEPT2 was analyzed
by a Spearman's correlation test. P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-744-5p is downregulated in
CRC
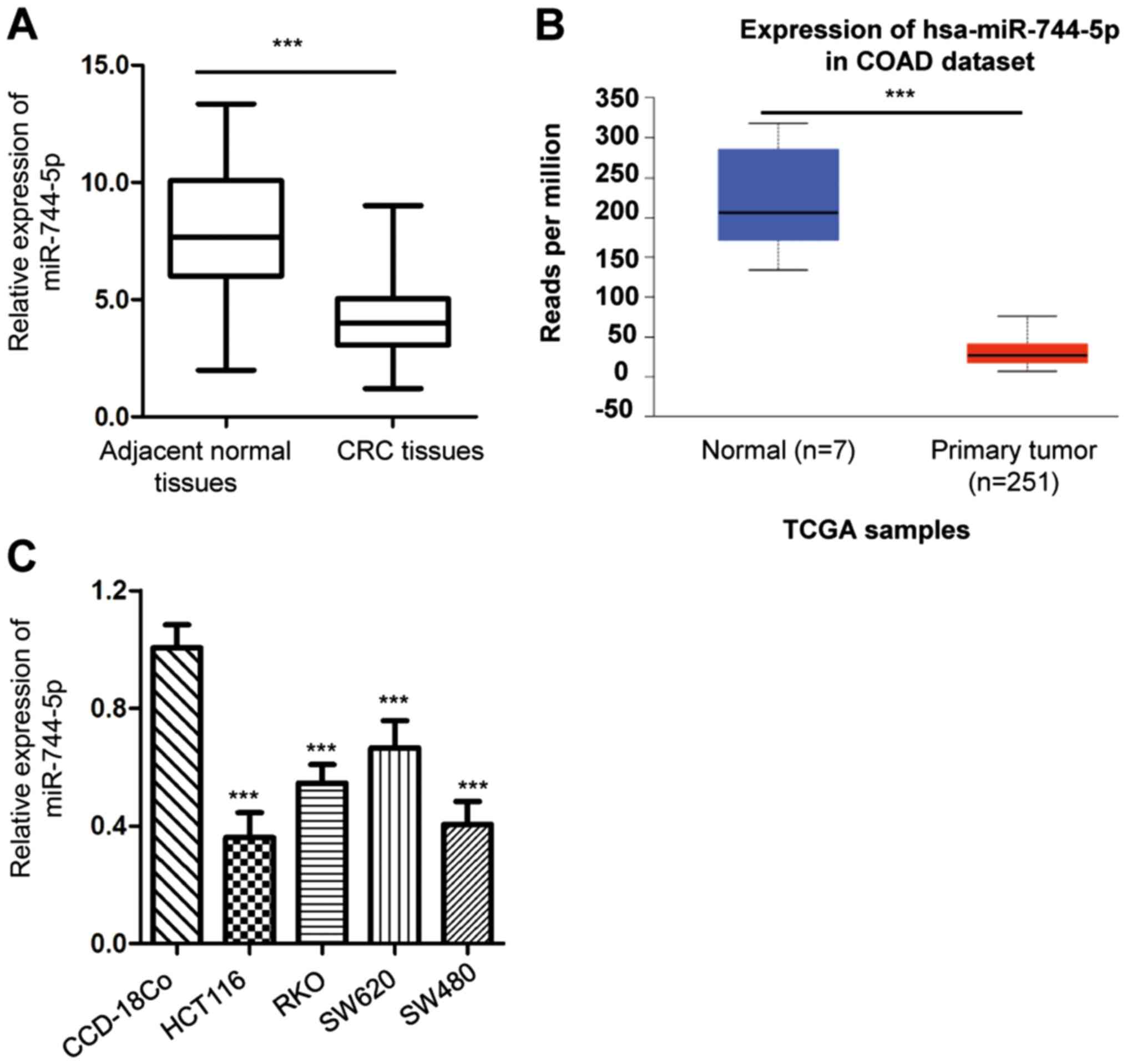
To explore the potential involvement of miR-744-5p
in CRC, the expression pattern of miR-744-5p in CRC tissues and
paired adjacent normal tissues was detected by RT-qPCR. The results
revealed that the expression level of miR-744-5p was significantly
reduced in CRC tissues compared with the surrounding non-cancerous
tissues (Fig. 1A). To support this
result, the expression of miR-744-5p in CRC tissues and normal
tissues was also evaluated against the TCGA-COAD database
(http://ualcan.path.uab.edu/index.html) (29). Consistent with the presented
findings, there was a significantly reduced level of miR-744-5p
observed in CRC tissues compared with the normal tissues (Fig. 1B). Additionally, miR-744-5p
expression in CRC cells and normal cells was analyzed. It was
determined that miR-744-5p was significantly decreased in CRC cell
lines compared with that in the normal cell line CCD-18Co (Fig. 1C). These results indicated the
downregulation of miR-744-5p in CRC.
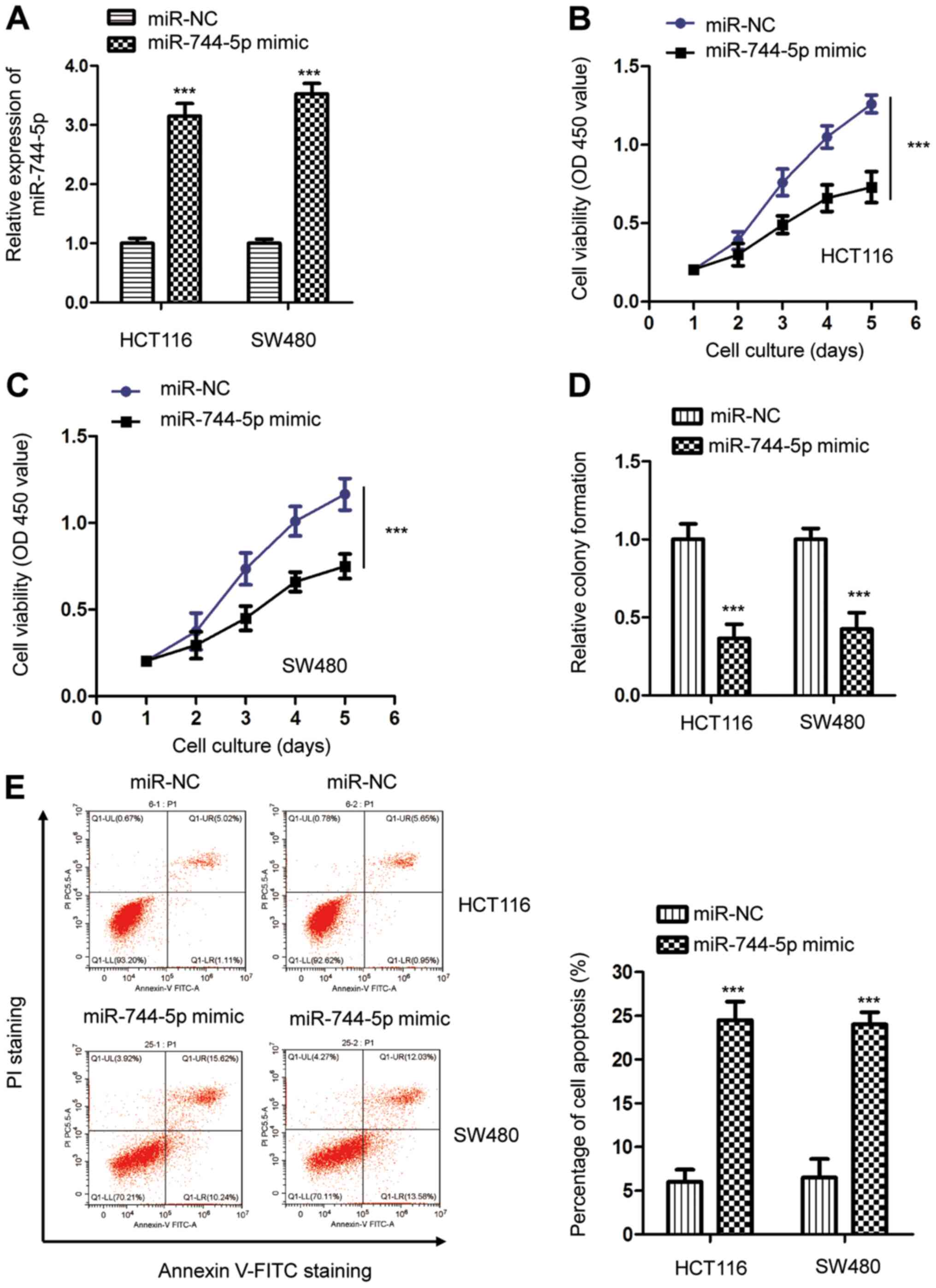
miR-744-5p inhibits the proliferation
and induces apoptosis in CRC cells
To investigate the function of miR-744-5p in CRC,
both HCT116 and SW480 cells were selected for biological analysis
due to the relatively low miR-744-5p in these cells. The miR-744-5p
mimic oligonucleotides were transfected into CRC cells and the
expression of miR-744-5p was detected by RT-qPCR (Fig. 2A). The CCK-8 assay revealed that
miR-744-5p overexpression significantly inhibited the proliferation
of both HCT116 and SW480 cells compared with cells transfected with
miR-NC (Fig. 2B and C). A colony
formation assay was performed to confirm the suppressive role of
miR-744-5p in CRC. The results indicated that overexpression of
miR-744-5p inhibited the colony formation ability of both HCT116
and SW480 cells (Fig. 2D).
Consistent with these results, highly expressed miR-744-5p also
significantly upregulated the apoptosis of CRC cells (Fig. 2E). These findings indicated the
potential tumor-suppressive function of miR-744-5p in the
development of CRC.
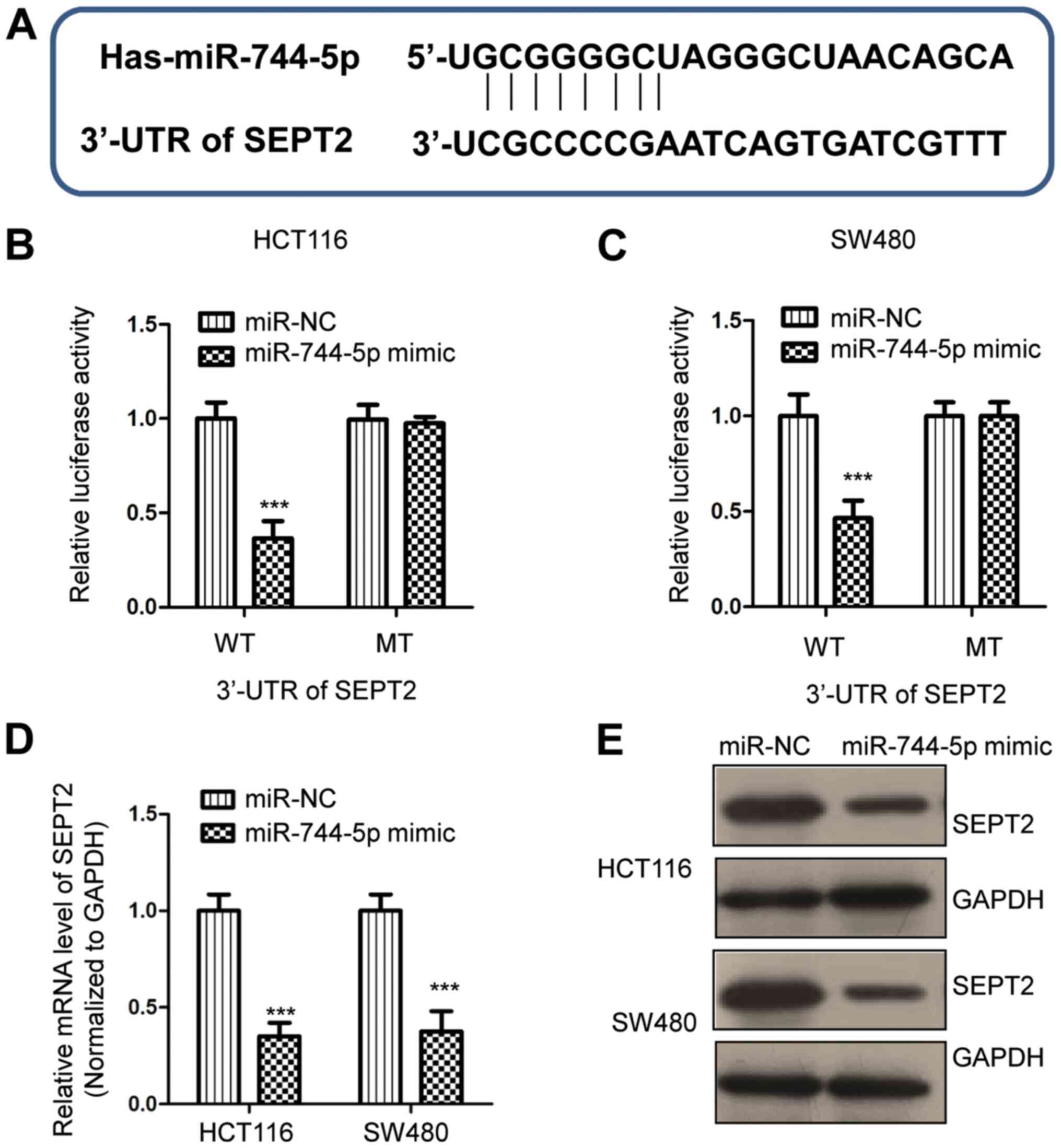
SEPT2 is a target of miR-744-5p in
CRC
To understand the molecular mechanism underlying the
tumor-suppressive role of miR-744-5p in CRC, the miRDB database was
used to predict the potential binding targets of miR-744-5p
(29). It was determined that the
3′-UTR of SEPT2 contains a putative binding site for miR-744-5p
(Fig. 3A). To confirm this
predicted binding, a dual-luciferase assay was performed by
transfecting miR-744-5p mimics and the luciferase reporter vector
carrying WT or MT 3′-UTR of SEPT2. The data revealed that
overexpression of miR-744-5p significantly decreased the luciferase
activity of WT but not MT 3′-UTR of SEPT2 (Fig. 3B and C). To detect whether
miR-744-5p regulates the expression of SEPT2, RT-qPCR and western
blot assays were performed to examine the mRNA and protein levels
of SEPT2, respectively. The results revealed that overexpression of
miR-744-5p significantly reduced the mRNA level of SEPT2 in CRC
cells (Fig. 3D). Similarly, the
protein expression of SEPT2 was also suppressed by the transfection
of miR-744-5p in both HCT116 and SW480 cells (Fig. 3E). These results demonstrated that
miR-744-5p targeted SEPT2 and negatively modulated the expression
of SEPT2 in CRC.
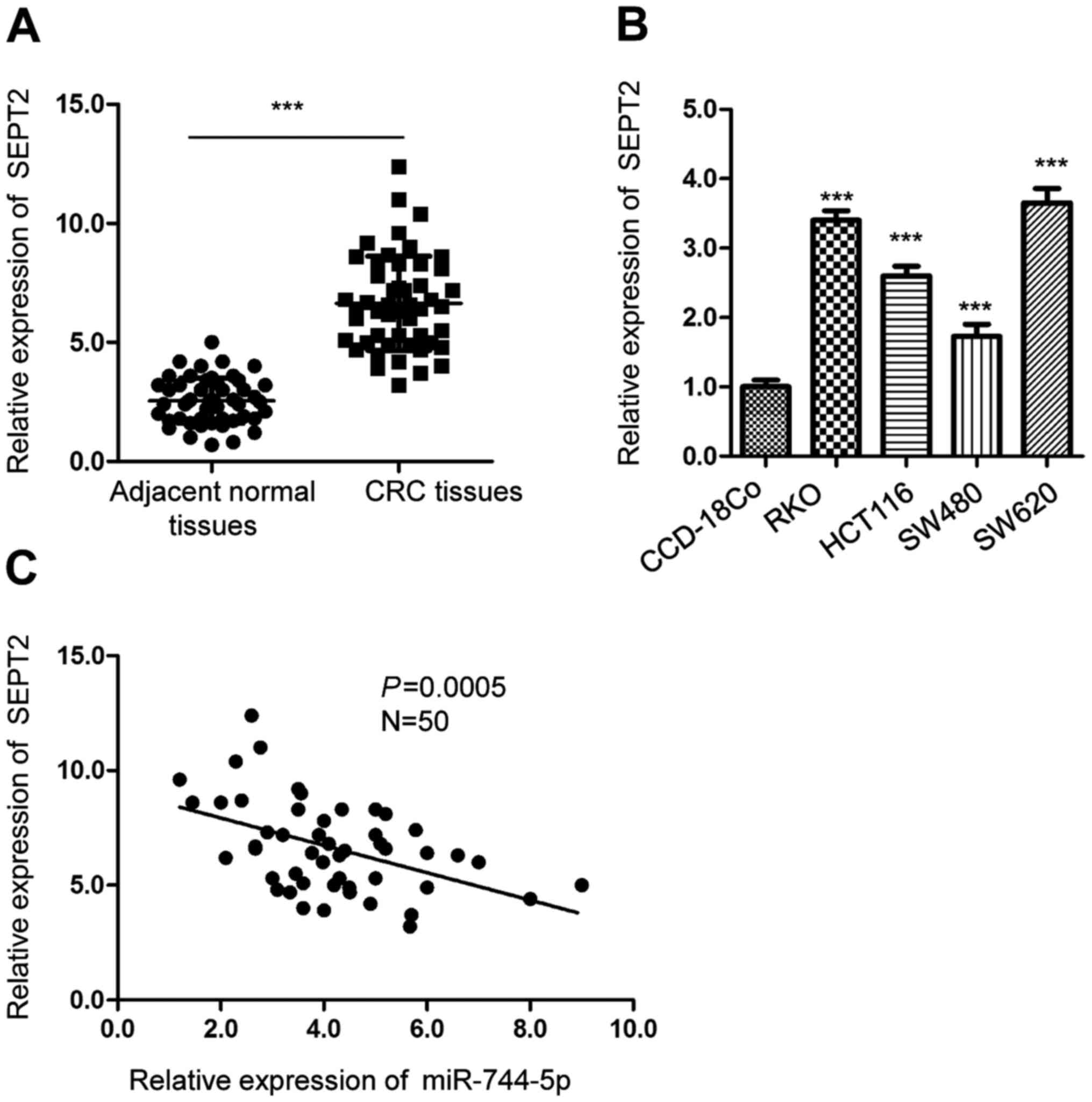
SEPT2 is upregulated in CRC and is
inversely correlated with the expression of miR-744-5p
To further explore the relationship between the
level of SEPT2 and miR-744-5p, the expression of SEPT2 in CRC
tissues and paired adjacent normal tissues was detected. The
RT-qPCR data revealed that SEPT2 expression was significantly
increased in CRC tissues compared with the surrounding
non-cancerous tissues (Fig. 4A).
The level of SEPT2 was also higher in CRC cells than in normal
CCD-18Co cells (Fig. 4B).
Additionally, the correlation between the expression of miR-744-5p
and SEPT2 was analyzed using Spearman's correlation test. The data
revealed that the expression of SEPT2 was inversely correlated with
the level of miR-744-5p in CRC tissues (Fig. 4C). The findings of the present study
supported the conclusion that SEPT2 was a target of miR-744-5p in
CRC.
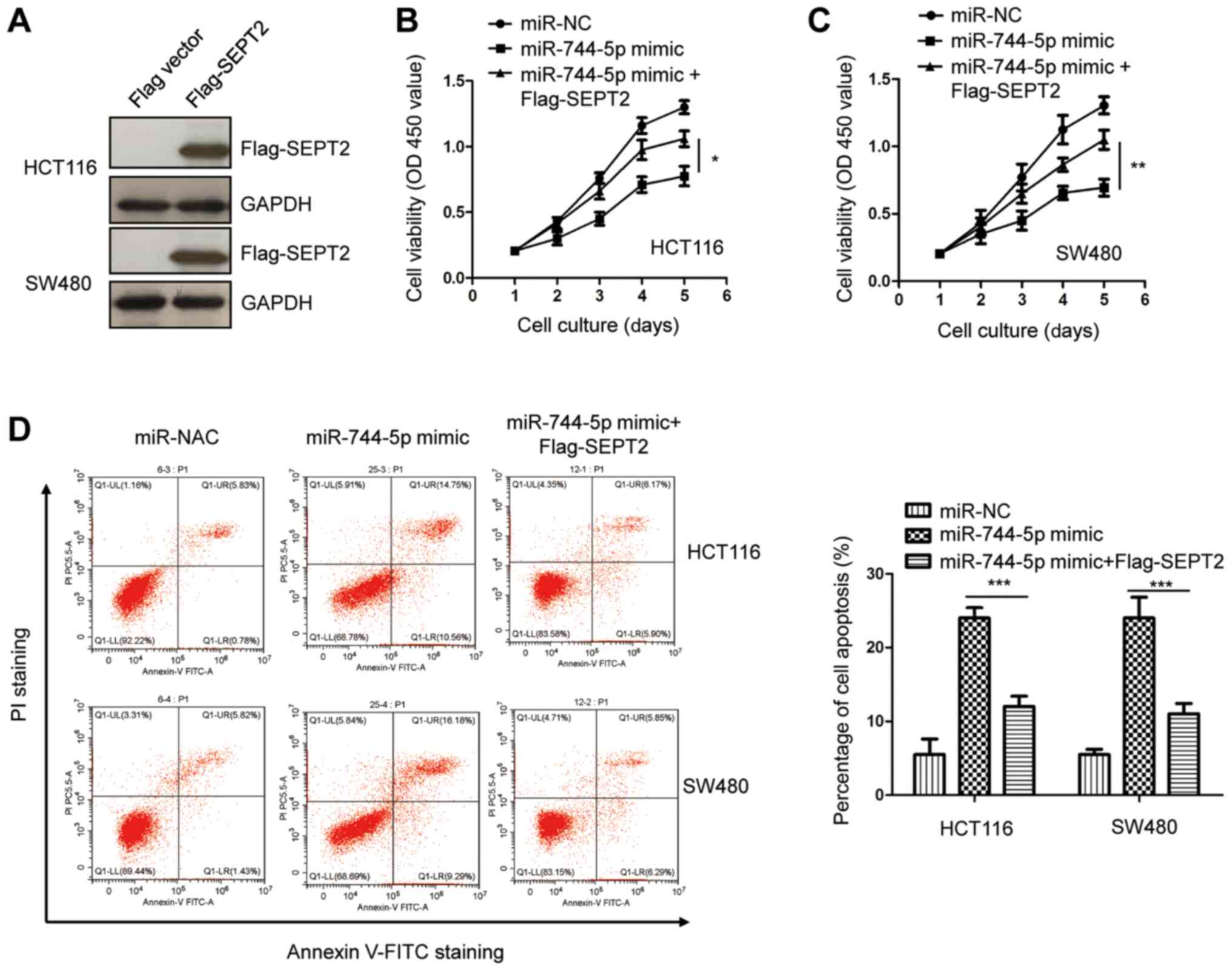
Overexpression of SEPT2 reverses the
tumor-suppressive effects of miR-744-5p in CRC
To investigate whether SEPT2 plays a suppressive
role for miR-744-5p in the malignancy of CRC, SEPT2 was
overexpressed by transfecting pcDNA-3Flag-SEPT2 into both HCT116
and SW480 cells. The ectopic expression of SEPT2 was validated by
western blotting (Fig. 5A). The
results of the CCK-8 assay revealed that SEPT2 overexpression
reversed the suppressive effects of miR-744-5p mimics on CRC cell
proliferation (Fig. 5B and C).
Additionally, the apoptosis of CRC cells was significantly
decreased with the co-transfection of SEPT2 compared with cells
only expressing miR-744-5p mimics (Fig.
5D). These results indicated that miR-744-5p partially
inhibited the progression of CRC by downregulating SEPT2. Notably,
recent studies reported that miR-744-5p targets c-Myc in papillary
thyroid carcinoma cells (30),
Heterogeneous Nuclear Ribonucleoprotein C (HNRNPC) and Nuclear
Factor I X (NFIX) in ovarian cancer cells (19), and Paired box gene 2 (PAX2) in
non-small cell lung cancer (NSCLC) (20). To determine the correlation between
miR-744-5p and the aforementioned gene expression, the expression
change of HNRNPC, NFIX, c-Myc and PAX2 in CRC tissues and normal
tissues was examined via TCGA-COAD database. No significant change
for the expression of NFIX and PAX2 were revealed in CRC tissues
compared with that of the normal tissues (Fig. 6A and B). It was also revealed that
the expression of NFIX and PAX2 was not significantly reduced by
the transfection of miR-744-5p in HCT116 and SW480 cells (Fig. 6C and D). In contrast, highly
expressed HNRNPC and c-Myc was revealed in CRC tissues compared
with the normal tissues by the TCGA-COAD (Fig. 6E and F). Decreased levels of HNRNPC
and c-Myc were observed with the transfection of miR-744-5p in
HCT116 and SW480 cells by RT-qPCR (Fig.
6G and H). Additionally, the expression of HNRNPC and c-Myc in
CRC tissues was analyzed for their correlation with miR-744-5p. An
inverse correlation was observed between the levels of HNRNPC,
c-Myc and miR-744-5p, respectively (Fig. 6I and J). These results indicated the
possibility of HNRNPC and c-Myc as potential targets of miR-744-5p
in CRC in addition to SEPT2.
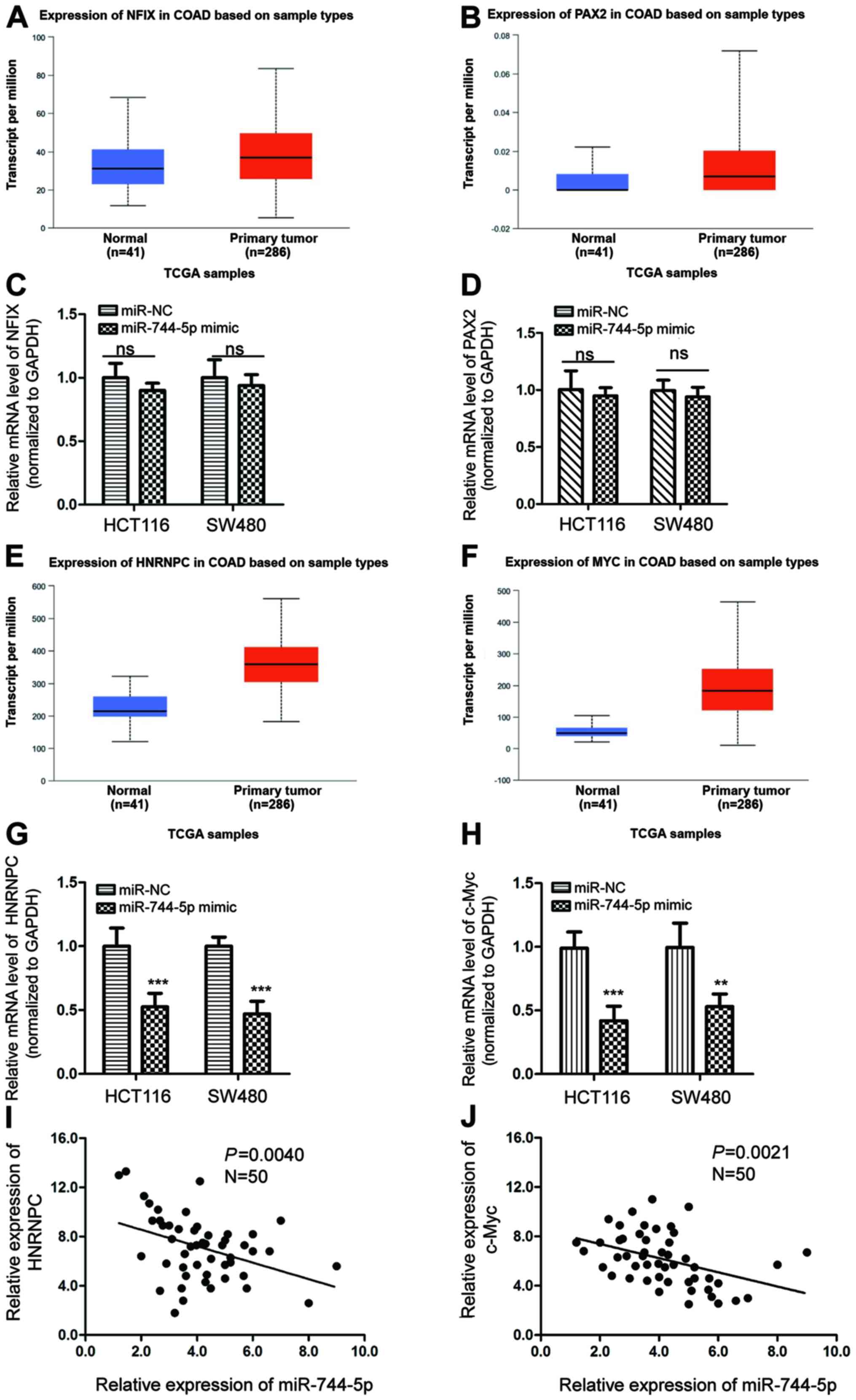 | Figure 6.Correlation between the levels of
miR-744-5p with other possible targets. (A and B) The expression of
NFIX and PAX2 in CRC tissues and normal tissues was evaluated with
the TCGA-COAD database. (C and D) HCT116 and SW480 cells were
transfected with miR-NC or miR-744-5p mimics, and the mRNA
abundance of NFIX and PAX2 was detected by RT-qPCR. (E and F) The
expression of HNRNPC and c-Myc in CRC tissues and normal tissues
was evaluated with the TCGA-COAD database. (G and H) The mRNA
levels of HNRNPC and c-Myc were examined by RT-qPCR with CRC cells
expressing miR-NC or miR-744-5p mimics. (I and J) The correlation
between the expression of miR-744-5p and HNRNPC, c-Myc in CRC
tissues was determined by the Spearman's correlation test,
respectively. **P<0.01; ***P<0.001. CRC, colorectal cancer;
miR, microRNA; RT-qPCR, reverse transcription-quantitative PCR;
NFIX, Nuclear Factor I X; PAX2, Paired box gene 2; HNRNPC,
Heterogeneous Nuclear Ribonucleoprotein C; miR-NC, micRNA control;
Ns, no significance. |
Discussion
CRC is one of the most common malignant tumors
worldwide and a major cause of cancer-related mortality (31). Growing evidence suggests that miRNA
acts as a tumor suppressor or promoter in the progression and
metastasis of CRC by regulating cell proliferation, apoptosis, and
cell cycles (14). The present
study provided novel insights into the tumor inhibitory effects of
miR-744-5p in the tumorigenesis of CRC. The present study detected
the expression level of miR-744-5p in CRC tissues and cell lines.
The function and underlying mechanisms of miR-744-5p that are
responsible for the effects of miR-744-5p in the development of CRC
were also investigated.
miR-744-5p has previously been revealed to be
significantly downregulated in cancers (19,20,32).
Overexpression of miR-744-5p induced apoptosis of ovarian cancer
cells and high miR-744-5p expression was associated with prolonged
disease-free survival of patients with ovarian cancer (19). Additionally, the serum level of
miR-744-5p was dysregulated in various types of cancers, such as
pancreatic and biliary tract cancers, which indicates the potential
significance of miR-744-5p in the diagnosis of cancers (21). A recent study also reported
decreased expression of miR-744-5p in NSCLC (20). Overexpression of miR-744-5p
inhibited the proliferation, colony formation, and invasion of
NSCLC cells. In the present study, miR-744-5p was significantly
downregulated in CRC tissues and cell lines. Transfection of
miR-744-5p suppressed proliferation and colony formation, and
induced apoptosis of CRC cells. These observations suggested that
miR-744-5p may be a potential therapeutic target for the treatment
of patients with CRC.
To investigate the molecular mechanism underlying
the tumor-suppressive effects of miR-744-5p in CRC, the potential
targets of miR-744-5p were predicted with bioinformatics software.
The 3′-UTR of SEPT2 was found to carry the binding sequence for
miR-744-5p. SEPT2 is a conserved filamentous GTPase and is
overexpressed in a variety of human cancers (33). SEPT2 acts as a cancer-promoting gene
and is associated with the poor prognosis of patients with cancer
(34). However, the regulatory
mechanism of SEPT2 in cancer remains unclear. In the present study,
miR-744-5p bound the 3′-UTR of SEPT2 and inhibited the expression
of SEPT2 in CRC cells. The level of SEPT2 was increased in CRC
tissues compared with the matched normal tissues. The upregulation
of SEPT2 was significantly inversely correlated with the level of
miR-744-5p in CRC tissues. The reintroduction of exogenously
expressed SEPT2 abrogated the suppressive function of miR-744-5p in
the proliferation of CRC cells. The present results demonstrated
that miR-744-5p targeted SEPT2 and inhibited the malignant
phenotype of CRC cells.
In conclusion, the present study indicated that
miR-744-5p was downregulated in CRC. Overexpression of miR-744-5p
significantly restricted the proliferation and promoted apoptosis
of CRC cells by targeting SEPT2. Hence, the miR-744-5p/SEPT2 axis
may represent a potential therapeutic target for patients with CRC.
The limitation of this study was that most of the conclusions were
obtained from in vitro assays. Therefore, further investigation is
required to examine the critical function of miR-744-5p/SEPT2
signaling in the progression of CRC using an in vivo study. Another
limitation is that only 50 patients were enrolled in the present
study to evaluate the clinical significance of miR-744-5p in CRC. A
larger sample size is required to investigate the association
between miR-744-5p expression and the prognosis of patients with
CRC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
All authors (WZ, KL and DL) contributed to the
project design, data acquisition and analysis, manuscript writing
and gave final approval of the version to be published. All authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
The usage of patient tissues was approved by the
Ethics Committee of the People's Hospital of Yichun City (approval
no. 2014041566) on April 15, 2014. Written informed consent was
obtained from all the patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Weinberg BA, Marshall JL and Salem ME: The
Growing Challenge of Young Adults With Colorectal Cancer. Oncology
(Williston Park). 31:381–389. 2017.PubMed/NCBI
|
|
2
|
Yiu AJ and Yiu CY: Biomarkers in
Colorectal Cancer. Anticancer Res. 36:1093–1102. 2016.PubMed/NCBI
|
|
3
|
Kuipers EJ, Grady WM, Lieberman D,
Seufferlein T, Sung JJ, Boelens PG, van de Velde CJ and Watanabe T:
Colorectal cancer. Nat Rev Dis Primers. 1:150652015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cai Y, Yu X, Hu S and Yu J: A brief review
on the mechanisms of miRNA regulation. Genomics Proteomics
Bioinformatics. 7:147–154. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mohr AM and Mott JL: Overview of microRNA
biology. Semin Liver Dis. 35:3–11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fabian MR, Sonenberg N and Filipowicz W:
Regulation of mRNA translation and stability by microRNAs. Annu Rev
Biochem. 79:351–379. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kwak PB, Iwasaki S and Tomari Y: The
microRNA pathway and cancer. Cancer Sci. 101:2309–2315. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Farazi TA, Spitzer JI, Morozov P and
Tuschl T: miRNAs in human cancer. J Pathol. 223:102–115. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qu H, Xu W, Huang Y and Yang S:
Circulating miRNAs: Promising biomarkers of human cancer. Asian Pac
J Cancer Prev. 12:1117–1125. 2011.PubMed/NCBI
|
|
12
|
Gentilin E, Degli Uberti E and Zatelli MC:
Strategies to use microRNAs as therapeutic targets. Best Pract Res
Clin Endocrinol Metab. 30:629–639. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Iorio MV and Croce CM: MicroRNA
dysregulation in cancer: Diagnostics, monitoring and therapeutics.
A comprehensive review. EMBO Mol Med. 9:8522017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sur DG, Colceriu M, Sur G, Aldea C,
Silaghi C, Samasca G, Lupan I, Căinap C, Burz C and Irimie A:
MiRNAs roles in the diagnosis, prognosis and treatment of
colorectal cancer. Expert Rev Proteomics. 16:851–856. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wei L, Wang X, Lv L, Zheng Y, Zhang N and
Yang M: The emerging role of noncoding RNAs in colorectal cancer
chemoresistance. Cell Oncol (Dordr). 42:757–768. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wan TMH, Iyer DN and Ng L: Roles of
microRNAs as non-invasive biomarker and therapeutic target in
colorectal cancer. Histol Histopathol. 35:225–237. 2020.PubMed/NCBI
|
|
17
|
Wang YG, He Q, Guo SQ and Shi ZZ: Reduced
serum miR-98 predicts unfavorable clinical outcome of colorectal
cancer. Eur Rev Med Pharmacol Sci. 23:8345–8353. 2019.PubMed/NCBI
|
|
18
|
Zhang L, Li B, Zhang B, Zhang H and Suo J:
miR-361 enhances sensitivity to 5-fluorouracil by targeting the
FOXM1-ABCC5/10 signaling pathway in colorectal cancer. Oncol Lett.
18:4064–4073. 2019.PubMed/NCBI
|
|
19
|
Kleemann M, Schneider H, Unger K, Sander
P, Schneider EM, Fischer-Posovszky P, Handrick R and Otte K:
miR-744-5p inducing cell death by directly targeting HNRNPC and
NFIX in ovarian cancer cells. Sci Rep. 8:90202018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen S, Shi F, Zhang W, Zhou Y and Huang
J: miR-744-5p inhibits non-small cell lung cancer proliferation and
invasion by directly targeting PAX2. Technol Cancer Res Treat.
18:15330338198769132019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim K, Yoo D, Lee HS, Lee KJ, Park SB, Kim
C, Jo JH, Jung DE and Song SY: Identification of potential
biomarkers for diagnosis of pancreatic and biliary tract cancers by
sequencing of serum microRNAs. BMC Med Genomics. 12:622019.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dolat L, Hu Q and Spiliotis ET: Septin
functions in organ system physiology and pathology. Biol Chem.
395:123–141. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cerveira N, Correia C, Bizarro S, Pinto C,
Lisboa S, Mariz JM, Marques M and Teixeira MR: SEPT2 is a new
fusion partner of MLL in acute myeloid leukemia with
t(2;11)(q37;q23). Oncogene. 25:6147–6152. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Angelis D and Spiliotis ET: Septin
Mutations in Human Cancers. Front Cell Dev Biol. 4:1222016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yu W, Ding X, Chen F, Liu M, Shen S, Gu X
and Yu L: The phosphorylation of SEPT2 on Ser218 by casein kinase 2
is important to hepatoma carcinoma cell proliferation. Mol Cell
Biochem. 325:61–67. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu D, Liu A, Wang X, Chen Y, Shen Y, Tan Z
and Qiu M: Repression of Septin9 and Septin2 suppresses tumor
growth of human glioblastoma cells. Cell Death Dis. 9:5142018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Neubauer K and Zieger B: The Mammalian
Septin Interactome. Front Cell Dev Biol. 5:32017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chandrashekar DS, Bashel B, Balasubramanya
SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BVSK and
Varambally S: UALCAN: A portal for facilitating tumor subgroup gene
expression and survival analyses. Neoplasia. 19:649–658. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yuan Q, Fan Y, Liu Z, Wang X, Jia M, Geng
Z, Zheng J and Lu X: miR-744-5p mediates lncRNA HOTTIP to regulate
the proliferation and apoptosis of papillary thyroid carcinoma
cells. Exp Cell Res. 392:1120242020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Azeem S, Gillani SW, Siddiqui A,
Jandrajupalli SB, Poh V and Syed Sulaiman SA: Diet and colorectal
cancer risk in Asia - a systematic review. Asian Pac J Cancer Prev.
16:5389–5396. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sui Y, Lin G, Zheng Y and Huang W: LncRNA
MAFG-AS1 boosts the proliferation of lung adenocarcinoma cells via
regulating miR-744-5p/MAFG axis. Eur J Pharmacol. 859:1724652019.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fung KY, Dai L and Trimble WS: Cell and
molecular biology of septins. Int Rev Cell Mol Biol. 310:289–339.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu M, Shen S, Chen F, Yu W and Yu L:
Linking the septin expression with carcinogenesis. Mol BiolRep.
37:3601–3608. 2010.
|