Introduction
Cervical cancer is the fourth most common malignancy
in women worldwide, resulting in considerable economic and medical
burden within society (1). Cervical
cancer is a global public health issue (1) with the number of associated deaths per
year estimated to reach 460,000 by 2040 (2). Cervical cancer treatment typically
includes surgery, chemotherapy and radiotherapy; however, the
treatment strategies are not always sufficient (3). Various chemical drugs, such as
bevacizumab, topotecan hydrochloride and gemcitabine/cisplatin, are
the first-line treatments used in cervical cancer chemotherapy;
however, patients often display severe side effects and drug
resistance, resulting in tumor recurrence and further progression
(4,5). Therefore, improving the current
understanding of the biological properties of cervical cancer cells
and developing effective novel chemotherapeutic agents is
important.
Metformin, a biguanide derivative approved by the US
Food and Drug Administration, is a well-tolerated first-line
therapy for type 2 diabetes mellitus (6). Clinical and laboratory studies have
demonstrated that metformin inhibits cellular proliferation, and
induces apoptosis and cell cycle arrest in vitro (7–13).
Moreover, metformin reduced the growth of experimental tumors in
vivo, including in prostate, lung, breast, colon and pancreatic
cancer, as well as in oral squamous cell carcinoma and melanoma
(7–13). Moreover, several studies have
reported that in cancer therapeutics, metformin exerts its effects
via several molecular signaling pathways, including the
AMP-activated protein kinase (AMPK), mTOR, insulin-like growth
factor-1, JNK/p38 MAPK, human epidermal growth factor receptor-2
and NF-κB signaling pathways (14–20).
However, Kowall et al (21)
and Iliopoulos et al (22)
revealed that when used alone, metformin is not an effective
anticancer treatment for colorectal, lung, breast or prostate
cancer; however, when combined with multiple chemotherapeutic
agents, metformin can effectively reduce tumor progression in
various types of cancer. In vitro and in vivo studies
have indicated that when combined with caffeic acid or nelfinavir,
metformin synergistically inhibits cancer cell proliferation and
tumor growth (23,24). The combined use of metformin and
chemotherapeutics (including, gemcitabine, paclitaxel/carboplatin,
megestrol acetate and erlotinib) is being investigated in several
ongoing phase I/II clinical trials (clinicaltrials.gov) to assess whether these
combinations increase patient survival and inhibit pancreatic
(NCT02005419), ovarian (NCT02312261), endometrial (NCT01968317) and
breast cancer metastasis (NCT01650506).
Pollak (25)
revealed that the antineoplastic mechanisms underlying metformin
may display indirect and/or direct effects on cancer biology.
Indirectly, metformin binds components of respiratory complex I to
limit mitochondrial oxidative phosphorylation, resulting in induced
hepatic energy stress and reduced liver gluconeogenesis, which
ultimately leads to a decrease in circulating glucose and insulin
levels, and in turn may inhibit tumor growth in patients with
insulin-responsive cancer (26).
Regarding its direct effects on cancer, metformin may adequately
accumulate in neoplastic tissues resulting in reduced ATP
production, which triggers AMPK activation, but inhibits mTOR and
fatty acid synthases expression, ultimately reducing cellular
energy consumption and promoting energetic stress, resulting in
tumor cell death (27,28). Another study revealed that metformin
reduces ATP production, which when coupled with the loss of AMPK,
p53 or liver kinase B1 (LKB1) function, energy deficient tumor
cells may not exhibit a high compensatory rate of glycolysis,
resulting in a lethal energetic crisis and cytotoxic effects
(29–31). Thus, understanding the mechanisms
underlying the therapeutic effects of metformin is crucial for the
development of potential cervical cancer therapies.
AKT is a serine/threonine kinase with a key role in
the PI3K/AKT signaling pathway, which mediates various biological
functions, including cell survival, proliferation, apoptosis,
angiogenesis, glucose metabolism, mitochondrial membrane gradient
and protein synthesis (32,33). Increased AKT activity has been
reported in 30–50% of breast, ovarian, prostate and pancreatic
cancer tumors (34,35). The results of clinical studies
indicated that the levels of phosphorylated (p)-AKT were also high
in esophageal squamous cell carcinoma (90.4%), lung cancer (76.4%)
and breast cancer (20–26%), which often resulted in drug resistance
and lower patient survival rates (36–40).
In the present study, the anticancer mechanisms underlying
metformin were evaluated by performing an apoptosis assay, and the
activation of the AMPK/p53 and PI3K/AKT signaling pathways in human
cervical cancer cell lines (CaSki, C33A and HeLa) following
metformin treatment were evaluated.
Materials and methods
Cell line and culture
The CaSki, HeLa and C33A human cervical cancer cell
lines were purchased from the Bioresource Collection and Research
Center. CaSki cells were cultured in RPMI-1640 (BioConcept AG) and
HeLa and C33A cells were cultured in Eagle's minimum essential
medium (MEM; HyClone: Cytiva) supplemented with 10% fetal bovine
serum (FBS; HyClone: Cytiva) in a humidified incubator at 37°C with
5% CO2, respectively.
Cell viability assay
Cells were seeded into a 96-well plate
(2×104 cells/well) in RPMI-1640 or MEM. Subsequently,
cells were pre-treated with or without Compound C (an AMPK
inhibitor; Merck KGaA) for 2 h at 37°C, CaSki and C33A were cells
treated with 1 µM Compound C and HeLa cells were treated with 5 µM
Compound C, followed by treatment with metformin (0–20 mM; Cayman
Chemical Company) for 48 h at 37°C. Following treatment, 10 µl Cell
Counting Kit-8 (CCK-8) solution (Dojindo Molecular Technologies,
Inc.) was added to each well and incubated at 37°C for 1 h.
Absorbance was measured at a wavelength of 450 nm using a FLUOstar
Galaxy microplate reader (BMG Labtech GmbH).
Migration assay
Cell migration was assessed by performing a
Transwell migration assay using Transwell chambers (24-well
inserts; 8.0-micron PET; BD Biosciences). Cells were seeded
(2×104 cells/well) into the upper chamber in serum-free
RPMI-1640 or MEM medium containing 0, 5 or 10 mM metformin; these
concentrations of metformin having been selected based on the cell
viability assay test. The lower chamber was filled with 700 µl
RPMI-1640 or MEM medium supplemented with 10% FBS. Following
incubation for 48 h at 37°C, a wet cotton swab was used to remove
the non-migratory cells from the upper surface of the Transwell
membrane. Subsequently, migratory cells were fixed with 3.7%
formalin for 2 min and 100% methanol for 20 min at room temperature
(RT). Migratory cells were stained with 0.1% crystal violet for 20
min at RT and observed using a BX61 fluorescence microscope
(magnification, ×100; Olympus Corporation).
Apoptosis and cell cycle analyses
Apoptosis and cell cycle analyses were performed
using an FITC Annexin V Apoptosis Detection kit (BD Biosciences)
and PI/RNase staining buffer (BD Pharmingen; BD Biosciences),
respectively, according to manufacturer's instructions. Cells were
seeded (1×106 cells/well) into 6-well plates and treated
with 0, 5 or 10 mM metformin for 48 h at 37°C. Subsequently, cells
were collected, fixed with cold 70% ethanol at room temperature and
stored at −20°C until analysis. Prior to analysis, cells were
centrifuged at 400 × g for 10 min at 4°C and washed with cold PBS.
For cell cycle analysis, cells were stained with 0.5 ml PI/RNase
staining buffer for 15 min at RT in the dark. For apoptosis
analysis, cells were double-stained with 5 µl Annexin V-FITC and 5
µl PI for 15 min at RT in the dark. Stained cells were analyzed
using a Cytomics FC500 flow cytometer (Beckman Coulter, Inc.), CXP
software (version 2.3; Beckman Coulter, Inc.) and late apoptosis
was assessed.
Western blotting
Cells were seeded (2×106 cells/dish) into
10-cm dishes, pre-treated with or without Compound C (1 or 5 µM)
for 2 h at 37°C and incubated with 0, 5 or 10 mM metformin for 48 h
at 37°C. Total protein was isolated from cells using RIPA buffer
(EMD Millipore) and protein concentrations were determined using a
BCA protein assay kit (Thermo Fisher Scientific, Inc.). Proteins
(30 µg) were incubated at 95°C for 10 min, separated via 10–12%
SDS-PAGE and transferred to 0.2-µm PVDF membranes (Bio-Rad
Laboratories, Inc.). Following blocking with BlockPRO blocking
buffer (Energenesis Biomedical Co., Ltd.) for 1 h at room
temperature, the membranes were incubated overnight at 4°C with the
following primary antibodies (all 1:1,000): Monoclonal anti-p-AMPKα
(Thr172; cat. no. 2535; Cell Signaling Technology, Inc.),
monoclonal anti-AMPKα (cat. no. 5832; Cell Signaling Technology,
Inc.), polyclonal anti-p-p53 (cat. no. 9284; Cell Signaling
Technology, Inc.), polyclonal anti-p53 (cat. no. 9282; Cell
Signaling Technology, Inc.), monoclonal anti-Bcl-2 (cat. no. 15071;
Cell Signaling Technology, Inc.), monoclonal anti-Bax (cat. no.
5023; Cell Signaling Technology, Inc.), monoclonal anti-cleaved
caspase-3 (cat. no. 9664; Cell Signaling Technology, Inc.),
monoclonal anti-p-AKT (Ser473; cat. no. 4060; Cell Signaling
Technology, Inc.), monoclonal anti-Akt (cat. no. 4298; Cell
Signaling Technology, Inc.), monoclonal anti-p-p70S6 kinase
(p70S6K; Thr389; cat. no. 9234; Cell Signaling Technology, Inc.),
monoclonal anti-p70S6K (cat. no. 2708; Cell Signaling Technology,
Inc.), polyclonal anti-Bcl-2 antagonist/killer 1 (Bak; cat. no.
GTX100063; GeneTex, Inc.), polyclonal
anti-phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic
subunit α (PIK3CA; cat. no. NBP2-19804; Novus Biologicals, LLC) and
monoclonal anti-GAPDH (cat. no. AC002; ABclonal Biotech Co., Ltd.).
Following washing with TBST (Tris-buffered saline, 0.1% Tween-20),
the membranes were incubated with an IgG HRP-conjugated secondary
antibody (polyclonal anti-mouse; cat. no. 115-035-003; 1:50,000;
Jackson ImmunoResearch Laboratories, Inc.; polyclonal anti-rabbit;
cat. no. 31460; 1:100,000; Thermo Fisher Scientific Inc.) for 1 h
at room temperature. Proteins bands were visualized using enhanced
chemiluminescence reagent (EMD Millipore). Densitometry was
performed using Fusion-Capt Advanced FX7 software (version 16.08a;
Labtech International, Ltd.) with GAPDH as the loading control.
Statistical analysis
All experiments were performed at least three times.
Data are presented as the mean ± standard deviation. Comparisons
among groups were analyzed using one-way ANOVA followed by
Bonferroni's post hoc test using SPSS v22.0 (IBM Corp.). P<0.05
was considered to indicate a statistically significant
difference.
Results
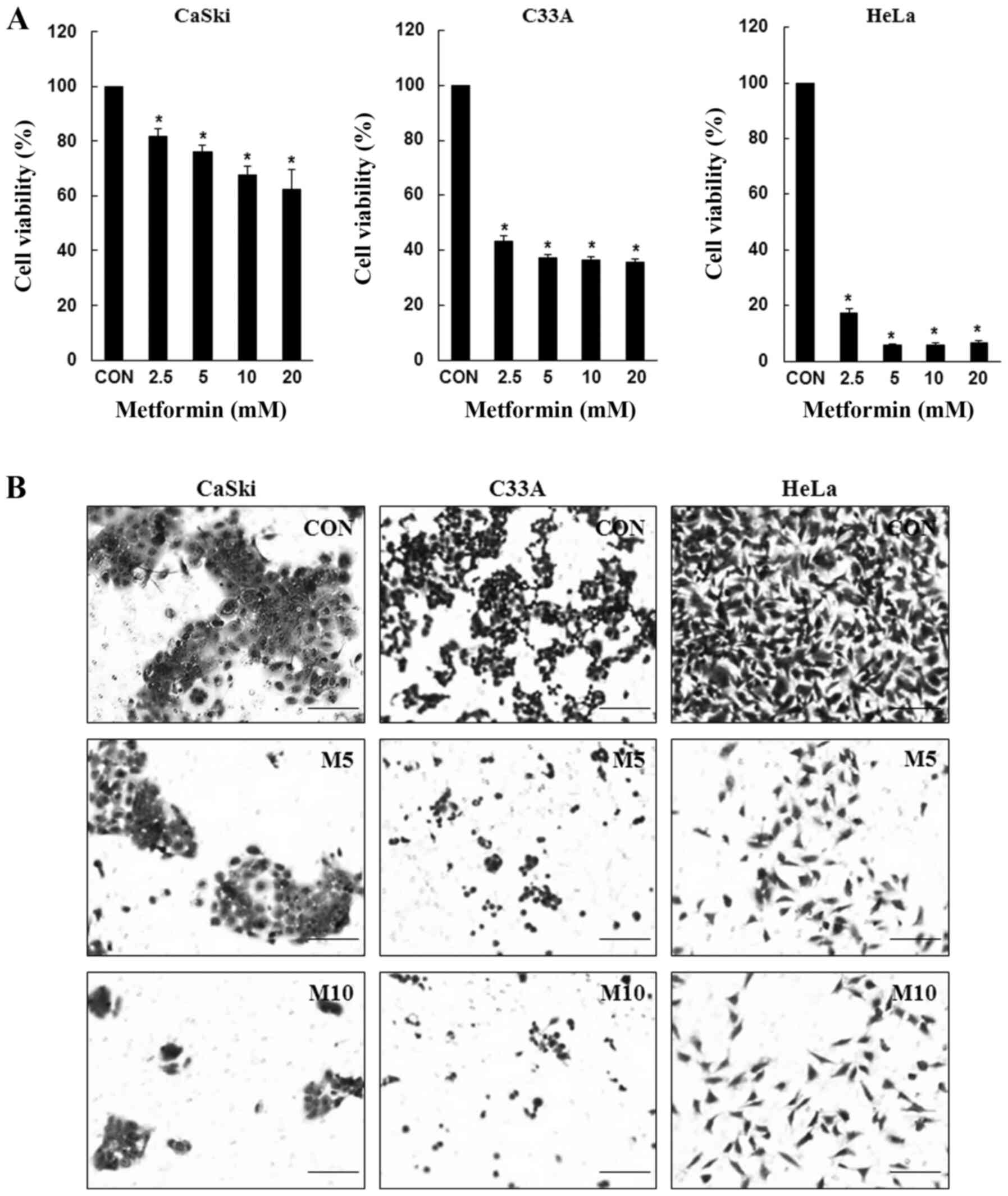
Metformin reduces human cervical
cancer cell viability and migration
To determine the inhibitory effects of metformin,
CaSki, C33A and HeLa cells were treated with various concentrations
of metformin for 48 h, and cell viability was determined using the
CCK-8 assay. Compared with the control group, metformin
significantly inhibited CaSki, C33A and HeLa cell viability in a
dose-dependent manner (Fig. 1A).
Similarly, compared with the control group, CaSki, C33A and HeLa
cell migration was also markedly decreased following treatment with
metformin at 48 h (Fig. 1B). The
results suggested that metformin inhibited cervical cancer cell
proliferation and migration.
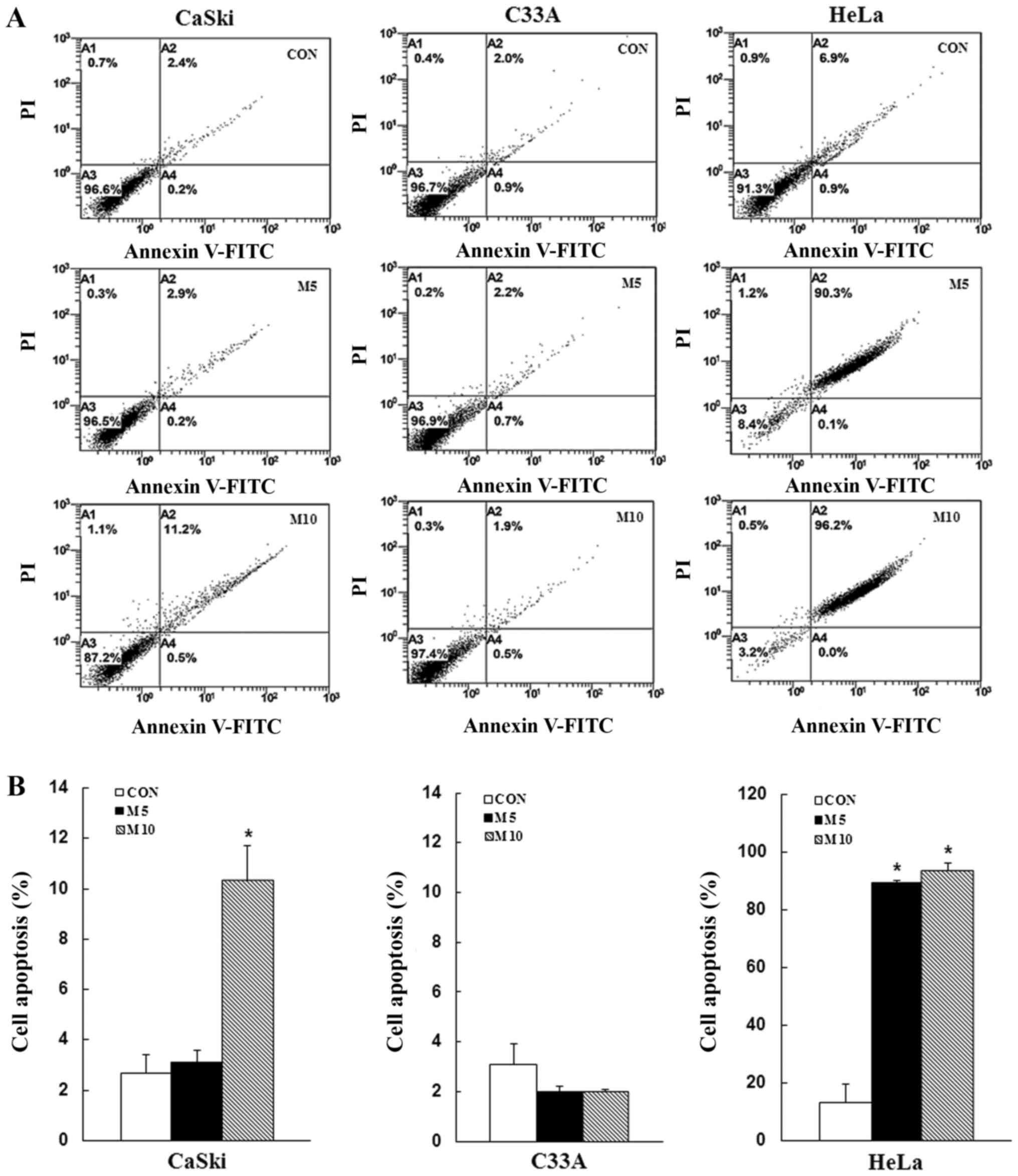
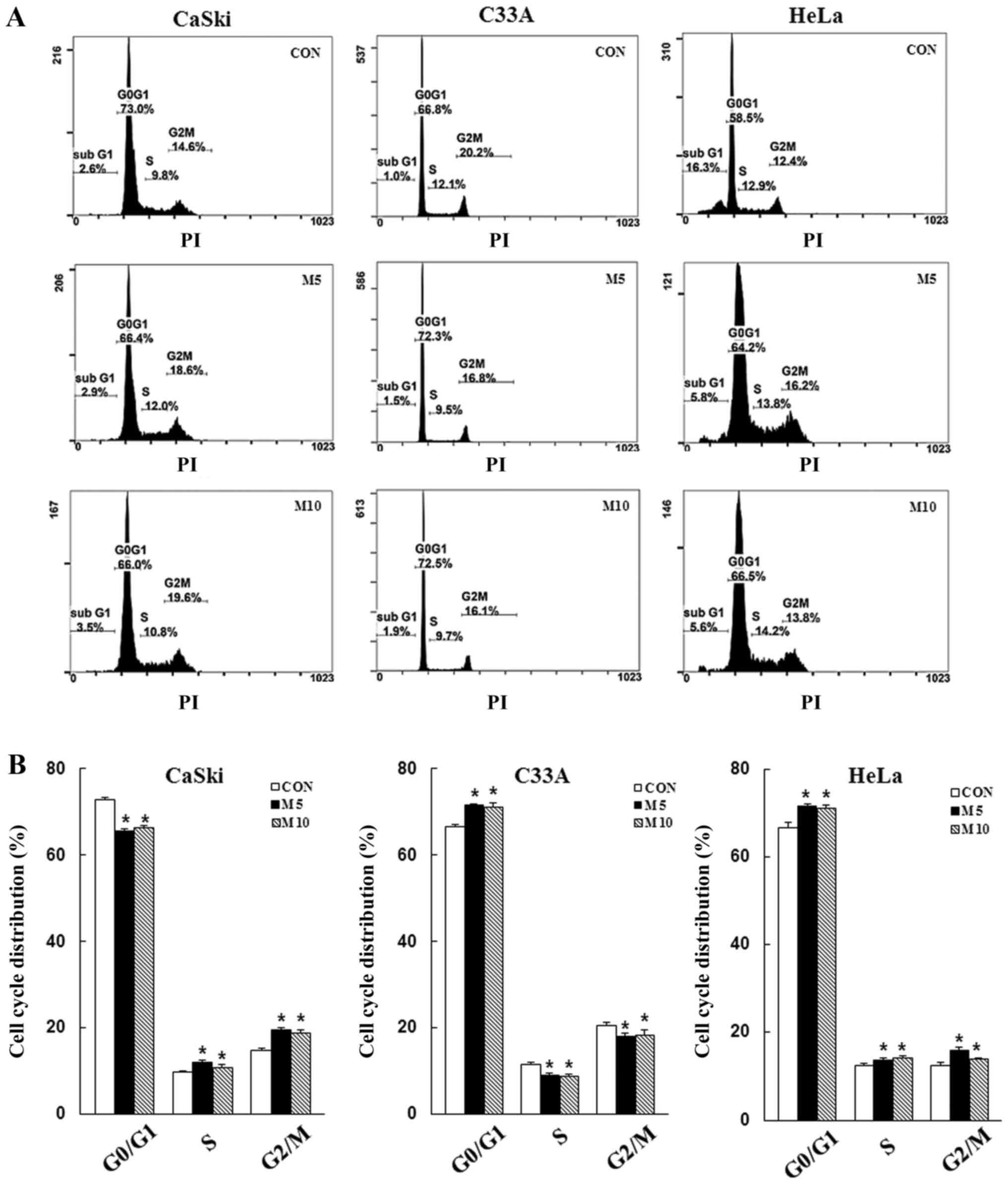
Metformin induces apoptosis and cell
cycle arrest in human cervical cancer cells
To determine whether the inhibitory effects of
metformin on cell proliferation were mediated via induction of
apoptosis and cell cycle arrest, CaSki, C33A and HeLa cells were
stained with Annexin V/PI or PI/RNase, respectively, and then
analyzed via flow cytometry. Compared with the control group,
metformin treatment increased the number of apoptotic cells in a
dose-dependent manner (CaSki cells, 10.3±1. vs. 2.7±0.7%; HeLa
cells, 93.8±2.5 vs. 13.3±6.6%), although no significant difference
was observed in C33A cells (2.0±0.1 vs. 3.1±0.8%) (Fig. 2). Furthermore, compared with the
control group, metformin treatment significantly increased the
number of CaSki and HeLa cells in the G2/M phase, as
well as the number of C33A and HeLa cells in the
G0/G1 phase (P<0.05; Fig. 3). The results indicated that
metformin mediated cervical cancer cell death and proliferation via
induction of apoptosis and cell cycle arrest.
Metformin activates the AMPK/p53
signaling pathway to mediate the mitochondrial apoptotic
pathway
Due to its involvement in apoptotic regulation
(41), AMPK/p53 signaling was
investigated to determine whether metformin-induced AMPK and p53
activation contributed to apoptosis. In the present study, CaSki,
C33A and HeLa cells were treated with 0, 5 or 10 mM metformin for
48 h, and the expression levels of AMPK, p53 and apoptosis-related
proteins were determined via western blotting. In CaSki and C33A
cells, compared with the control group, 5 mM metformin
significantly increased the expression levels of p-AMPK, p-p53, Bak
and Bax, and significantly decreased the expression levels of the
antiapoptotic protein Bcl-2 (Fig.
4). By contrast, p-p53, Bak, Bax and Bcl-2 expression levels
were significantly decreased in metformin-treated HeLa cells
compared with control cells. Compared with the control group, high
dose metformin (10 mM) treatment significantly increased the
expression levels of cleaved caspase-3 in CaSki, C33A and HeLa
cells. The results indicated that metformin mediated apoptosis via
targeting AMPK and mitochondria-mediated caspase-dependent
signaling pathways.
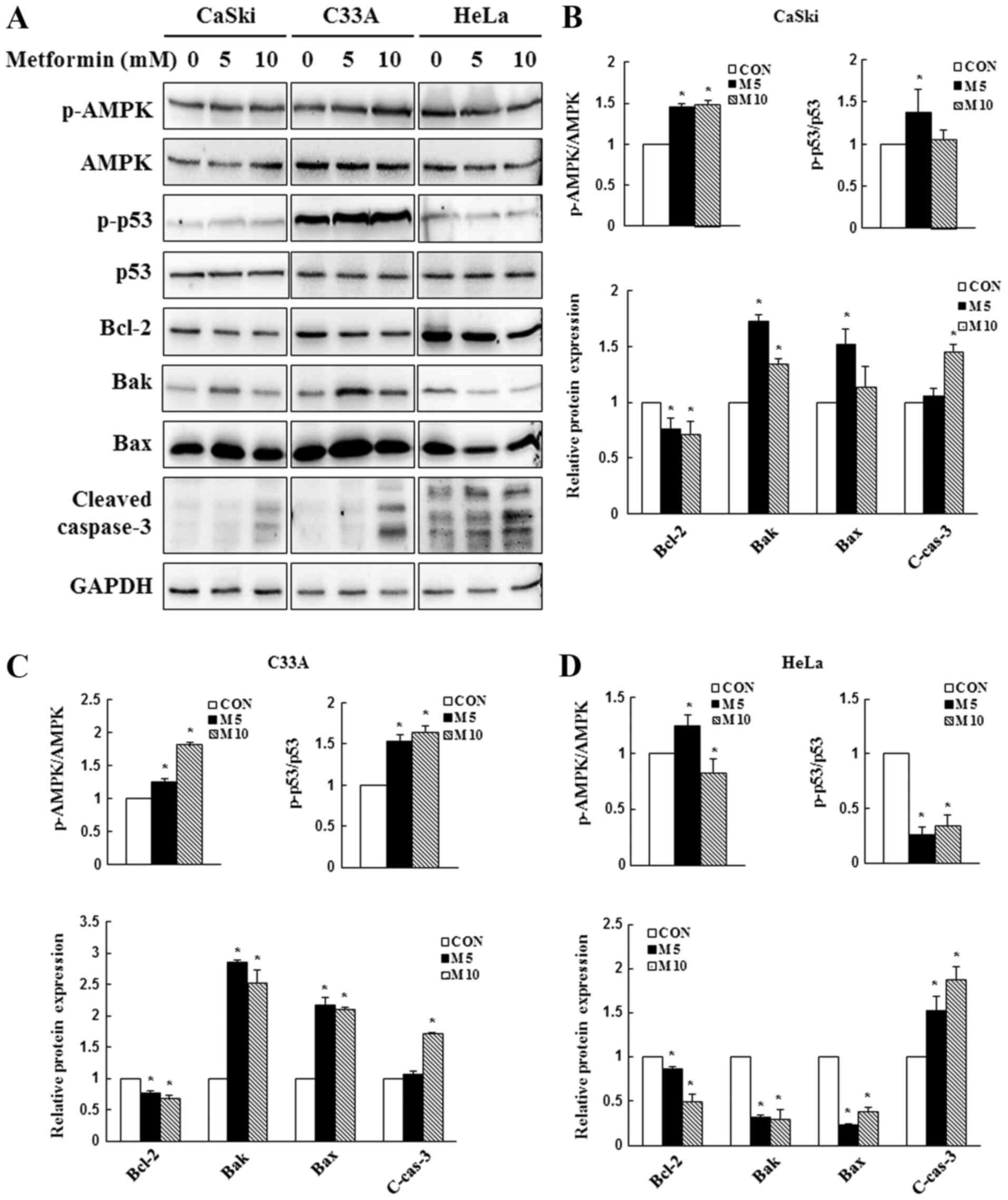 | Figure 4.Effect of metformin on AMPK and
caspase-dependent apoptosis signaling in cervical cancer cell
lines. CaSki, C33A and HeLa cells were treated with metformin (0, 5
or 10 mM) for 48 h. p-AMPK, AMPK, p-p53, p53, Bcl-2, Bak, Bax and
cleaved caspase-3 protein expression levels were (A) determined via
western blotting and semi-quantified in (B) CaSki, (C) C33A and (D)
HeLa cells. Data are presented as the mean ± SD from three
independent experiments. *P<0.05 vs. CON. AMPK, AMP-activated
protein kinase; p, phosphorylated; Bak, Bcl-2 antagonist/killer 1;
CON, 0 mM metformin; M5, 5 mM metformin; M10, 10 mM metformin;
C-cas-3, cleaved caspase-3. |
Metformin decreases PI3K/AKT
signaling
Previous studies have reported that the
PI3K/AKT/mTOR signaling pathway is overactivated in numerous cancer
types and its activation may promote cancer cell proliferation and
migration, as well as inhibit apoptosis (42,43).
In the present study, western blotting was performed to determine
the expression levels of PIK3CA, p-AKT and p-p70S6K. Compared with
the control group, 10 mM metformin significantly reduced PIK3CA,
p-AKT and p-p70S6K expression levels in CaSki, C33A and HeLa cells
(P<0.05; Fig. 5). The results
suggested that the PI3K/AKT signaling pathway might be involved in
regulating cellular physiology and apoptosis following metformin
treatment.
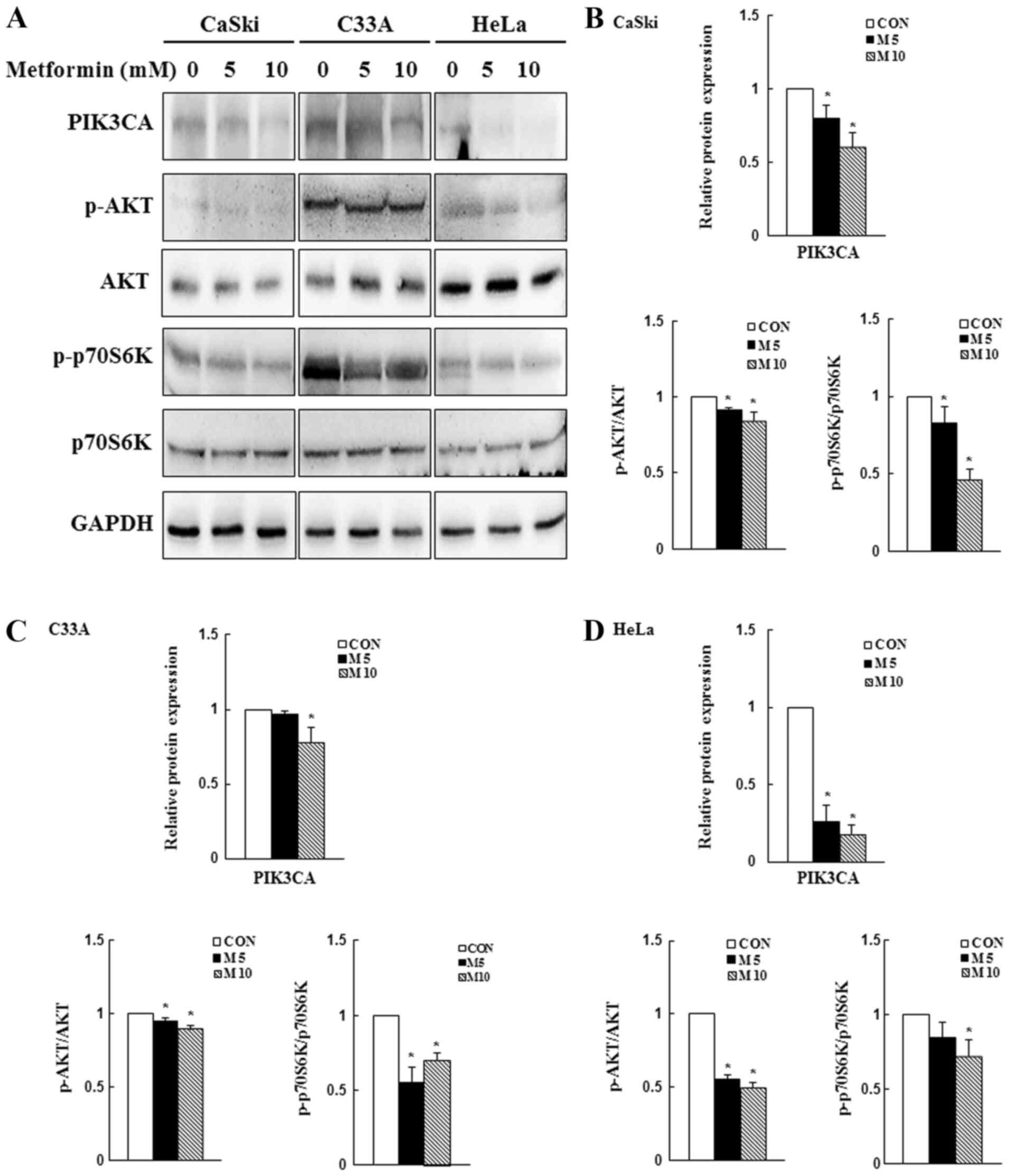 | Figure 5.Effects of metformin on PI3K/AKT/mTOR
signaling in cervical cancer cell lines. CaSki, C33A and HeLa cells
were treated with metformin (0, 5 or 10 mM) for 48 h. Protein
expression levels of PIK3CA, p-AKT, AKT, p-p70S6K, and p70S6K were
(A) determined via western blotting and semi-quantified in (B)
CaSki, (C) C33A and (D) HeLa cells. Data are presented as the mean
± SD from three independent experiments. *P<0.05 vs. CON. p,
phosphorylated; PIK3CA, phosphatidylinositol-4,5-bisphosphate
3-kinase catalytic subunit α; p70S6K, p70S6 kinase; CON, 0 mM
metformin; M5, 5 mM metformin; M10, 10 mM metformin. |
AMPK signaling pathway activity
contributes to metformin-induced cytotoxicity and apoptosis in
human cervical cancer cells
To assess whether the AMPK signaling pathway served
a key molecular role in metformin-treated cervical cancer cells,
CaSki, C33A and HeLa cells were pre-treated with or without 1 or 5
µM Compound C for 2 h, and then treated with or without 10 mM
metformin for 48 h. Cell viability and the expression levels of
p-AMPK, AMPK, p-p53, p53, Bcl-2 and cleaved caspase-3 were
determined by performing the CCK-8 assay and western blotting,
respectively. In CaSki and C33A cells, pre-treatment with Compound
C significantly reversed the effects of metformin on cell
viability, and suppressed p-AMPK, p-p53, cleaved caspase-3 and
increased Bcl-2 expression levels compared with metformin treatment
alone. By contrast, p-AMPK, p-p53 and Bcl-2 expression levels were
significantly increased in the metformin + Compound C group
compared with the metformin group in HeLa cells (P<0.05;
Fig. 6). The results indicated that
metformin enhanced apoptosis by targeting the AMPK/p53 signaling
pathway in CaSki, C33A and HeLa cells.
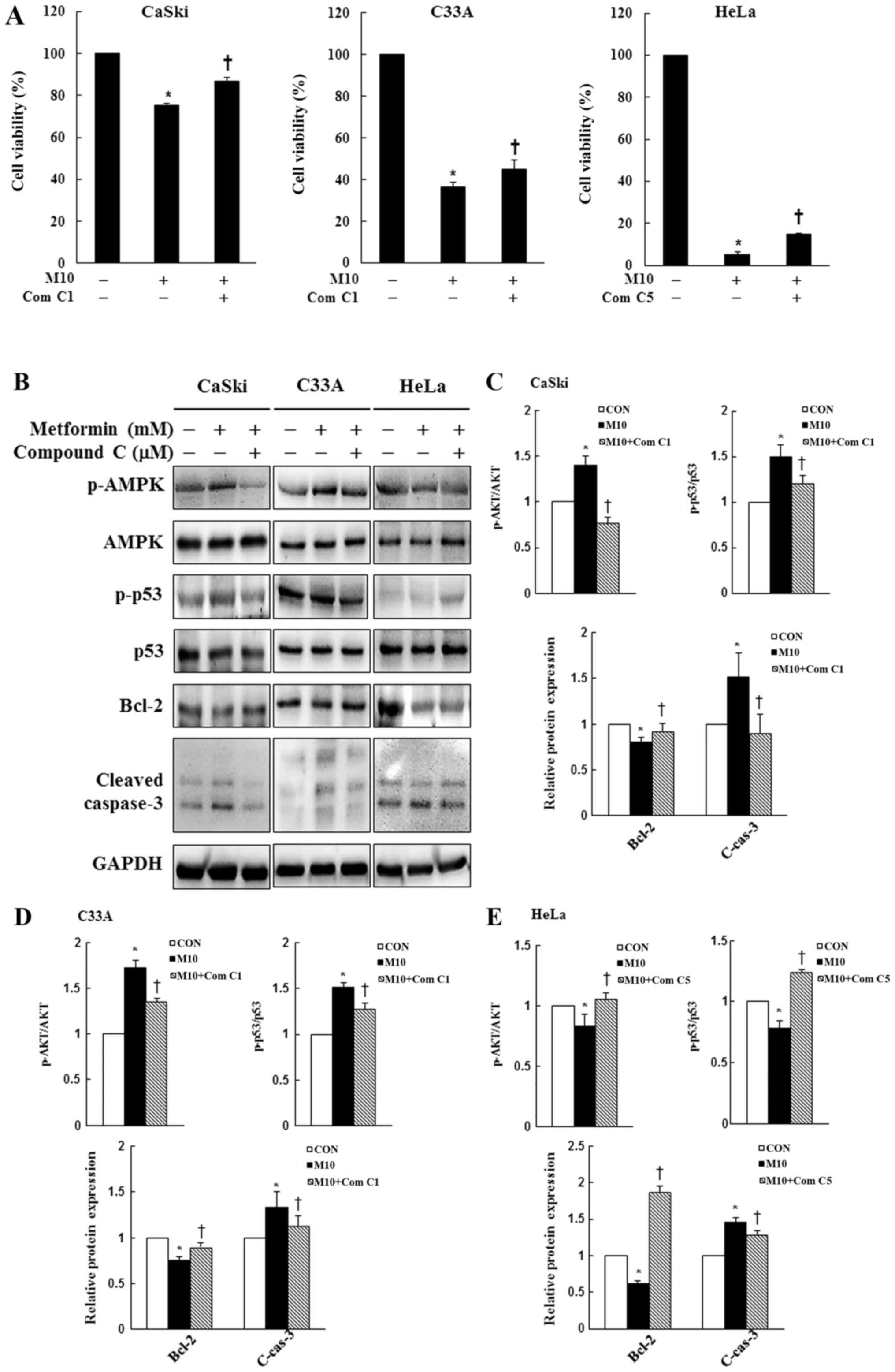 | Figure 6.Effects of Compound C on cell
viability, AMPK signaling and apoptotic signaling in cervical
cancer cell lines. CaSki, C33A and HeLa cells were pre-treated with
or without Compound C (1 or 5 µM; an AMPK inhibitor) for 2 h and
then treated with or without 10 mM metformin for 48 h, the
concentration of metformin selected for experiment was based on the
cell viability assay test. (A) Cell viability was determined by
performing the Cell Counting Kit-8 assay. Protein expression levels
of p-AMPK, AMPK, p-p53, p53, Bcl-2 and cleaved caspase-3 were (B)
determined by western blotting and semi-quantified in (C) CaSki,
(D) C33A and (E) HeLa cells. Data are presented as the mean ± SD
from three independent experiments. *P<0.05 vs. CON;
†P<0.05 vs. M10. AMPK, AMP-activated protein kinase;
p, phosphorylated; CON, 0 mM metformin; M10, 10 mM metformin; Com
C1, 1 µM Compound C; Com C5, 5 µM Compound C; C-cas-3, cleaved
caspase-3. |
Discussion
Metformin is a widely used antidiabetic drug that
has also been shown to reduce the risk of cancer and improve the
efficacy of cancer treatment in patients with diabetes (44,45).
However, multiple observational studies have demonstrated that
metformin displays a similar protective effect and can improve the
survival time of non-diabetic patients with cancer (46,47),
but the results are controversial. In the present study, CCK-8,
Transwell and flow cytometry assays were performed to evaluate the
effects of metformin on cell viability, migration, apoptosis and
the cell cycle in human cervical cancer cells, respectively.
Western blotting was conducted to quantify protein expression. The
results of the present study indicated that metformin significantly
reduced human cervical cancer cell viability (CaSki, C33A and HeLa)
in a dose-dependent manner (Fig.
1A), which was consistent with previous studies that
demonstrated that metformin significantly reduced the viability of
thyroid cancer, osteosarcoma, leukemia and bile duct cancer cells
by downregulating the expression of the antiapoptotic protein
Bcl-2, and upregulating the expression of the proapoptotic proteins
Bak and Bax (16,48–50).
The present study also indicated that HeLa cells were more
sensitive to metformin compared with CaSki and C33A cells, which
may be caused by cell-type specificity and mutations in
cancer-related genes resulting in resistance to the
anti-proliferative effects of metformin. CaSki cells, which are
derived from small bowel metastasis, have been reported to possess
an integrated human papilloma virus 16 genome (51). C33A cells are human papillomavirus
(HPV)-negative human cervical cancer cells that display upregulated
expression of oncogenes p53 and retinoblastoma protein (52). HeLa cells are a cervical
adenocarcinoma-derived cell line containing integrated HPV-18 DNA
and lower levels of p53 expression (53).
Consistent with the results of the present study,
Hsieh Li et al (53)
reported that metformin significantly induces apoptotic HeLa cell
death and reduces p53 expression levels. By contrast, Xiao et
al (54) reported that
metformin is less effective against HeLa cells and enhances AMPK
activation, but does not alter the expression levels of LKB1 or
p53. Irie et al (55) also
demonstrated that metformin is able to enhance LKB1
phosphorylation, promote AMPK and p53 activation, and inhibit cell
cycle progression, resulting in cervical cancer cell death.
Therefore, the mechanism underlying metformin-mediated effects on
cervical cancer requires further investigation.
In addition, the present study indicated that
metformin-induced cytotoxicity occurred via activation of the
caspase-dependent apoptotic signaling pathway. Moreover, the
results suggested that metformin induced cytotoxicity by promoting
cell cycle arrest in the G2/M phase and increasing
apoptosis in CaSki and HeLa cells, and promoting
G0/G1 phase cell cycle arrest in C33A and
HeLa cells. Previous studies have illustrated the effects of
metformin on the cell cycle in human osteosarcoma, demonstrating
increased K7M2 and MG63 cell numbers in the
G0/G1 phase, and increased U20S and 143B cell
numbers in the G2/M phase (49,56).
Additionally, metformin has been reported to enhance anticancer
effects by arresting human colon carcinoma cells in the
G0/G1 phase (SW480 cells) or the
G2/M phase (HCT116 p53−/− cells) (57,58).
The differences among cell lines may be associated with cancer cell
line specificity and individual metformin bioavailability.
AMPK is not only a sensor of cellular energetics,
but is also a crosstalk protein involved in apoptotic signaling
pathways, including the LKB1-AMPK and AMPK/p53 signaling pathways
(41,59). The results of the present study
revealed that AMPK activation was consistent with increased p-p53
expression following metformin treatment, and that the AMPK
inhibitor Compound C significantly alleviated p53 activation in
metformin-treated cells. The aforementioned results indicated that
the anticancer effect of metformin was mediated via the AMPK/p53
signaling pathway, although metformin did not induce p53 activation
in HeLa cells. Furthermore, the results of the present study were
consistent with previous reports that demonstrated that metformin
induces the phosphorylation and activation of p53, which inhibits
prostate, melanoma, lymphoma and acute myeloid leukemia cancer cell
proliferation (49,60–62).
Moreover, metformin can inhibit p53−/− colorectal cancer
cell proliferation both in vitro and in vivo
(30). Previous studies have
reported that metformin directly decreases the expression levels of
endogenous p53 in sensitive cells, which downregulates the
expression of target genes, including p21, Bax, Bak and BH3-only
proteins (Bid and Bim), ultimately resulting in apoptosis (53,63,64).
The results of the aforementioned previous studies were consistent
with the results of the present study, which indicated that
high-dose metformin (10 mM) induced CaSki cell apoptosis more
effectively compared with low-dose metformin (5 mM), despite
decreases in p-p53, Bak and Bax expression levels. Therefore, the
present study indicated that metformin activated AMPK, inhibiting
cervical cancer cell viability in both a p53-dependent and
-independent manner.
Metformin displays inhibitory effects on cell
proliferation, apoptosis, metastasis, angiogenesis and
chemoresistance in various malignancies in vitro and in
vivo, including ovarian (65),
endometrial (66) and
hepatocellular cancer (67). The
inhibitory effects of metformin were mediated via activation of the
PI3K/AKT/mTOR signaling pathway (65–67).
In an earlier study, Storozhuk et al (68) also demonstrated that metformin
inhibited tumor growth in non-small cell lung cancer (NSCLC) and
xenograft animal models by activating the ATM serine/threonine
kinase/AMPK/p53 signaling pathway and inhibiting the
AKT/mTOR/eukaryotic translation initiation factor 4E-binding
protein 1 signaling pathway, leading to an enhanced radiation
response in NSCLC. Likewise, the clinical studies conducted by
Dhillon et al (69) and
Sayed et al (70)
demonstrated that metformin treatment is associated with the
improved overall survival of patients with NSCLC. In the present
study, compared with the control group, metformin significantly
decreased PIK3CA, p-AKT and p-p70S6K expression levels in CaSki,
C33A and HeLa cells. The results indicated that metformin-mediated
alterations to the PI3K/AKT/mTOR signaling pathway were associated
with apoptosis induction in human cervical cancer cells.
In conclusion, the results of the present study
suggested that metformin induced apoptosis and cell cycle arrest by
modulating the AMPK/p53 and PI3K/AKT/mTOR signaling pathways.
Collectively, the results indicated that metformin might serve as a
novel therapeutic for human cervical cancer.
The present study had some limitations. First, it
focused on the effects of metformin on cell death-related pathways
in human cervical cancer cells but further studies should be
conducted to elucidate the underlying molecular mechanism of the
effects in human cervical cancer cells in more detail. Second, only
human cervical cancer cell lines were used; future studies using
animal models and patient tissues samples should be conducted to
elucidate the mechanism underlying metformin-mediated effects on
cervical cancer in vivo. Additional clinical studies are
required to assess the safety and clinical efficiency of metformin
for cervical cancer treatment.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Changhua
Christian Hospital Research Foundation (grant nos. 108-CCH-IRP-132
and 108-CCH-IRP-059).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YHC and YHH designed the study, prepared the figures
and drafted the manuscript. SFY and CKY designed the study. HDT,
THC and MCC contributed to the conception of the work, drafted the
manuscript and revised it critically for important intellectual
content. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Small W Jr, Bacon MA, Bajaj A, Chuang LT,
Fisher BJ, Harkenrider MM, Jhingran A, Kitchener HC, Mileshkin LR,
Viswanathan AN and Gaffney DK: Cervical cancer: A global health
crisis. Cancer. 123:2404–2412. 2017. View Article : Google Scholar
|
|
2
|
Simms KT, Steinberg J, Caruana M, Smith
MA, Lew JB, Soerjomataram I, Castle PE, Bray F and Canfell K:
Impact of scaled up human papillomavirus vaccination and cervical
screening and the potential for global elimination of cervical
cancer in 181 countries, 2020-99: A modelling study. Lancet Oncol.
20:394–407. 2019. View Article : Google Scholar
|
|
3
|
Ch PN, Gurram L, Chopra S and Mahantshetty
U: The management of locally advanced cervical cancer. Curr Opin
Oncol. 30:323–329. 2018. View Article : Google Scholar
|
|
4
|
Rosen VM, Guerra I, McCormack M,
Nogueira-Rodrigues A, Sasse A, Munk VC and Shang A: Systematic
review and network meta-analysis of bevacizumab plus first-line
topotecan-paclitaxel or cisplatin-paclitaxel versus
non-bevacizumab-containing therapies in persistent, recurrent, or
metastatic cervical cancer. Int J Gynecol Cancer. 27:1237–1246.
2017. View Article : Google Scholar
|
|
5
|
Bethesda. SEER Cancer Stat Facts. Cervical
Cancer. National Cancer Institute; simplehttps://seer.cancer.gov/statfacts/html/cervix.htmlMay.
2020
|
|
6
|
Nathan DM, Buse JB, Davidson MB, Heine RJ,
Holman RR, Sherwin R and Zinman B; Professional Practice Committee,
American Diabetes Association, European Association for the Study
of Diabetes: Management of hyperglycaemia in type 2 diabetes: A
consensus algorithm for the initiation and adjustment of therapy, :
A consensus statement from the American Diabetes Association and
the European Association for the study of Diabetes. Diabetologia.
49:1711–1721. 2006. View Article : Google Scholar
|
|
7
|
Crawley D, Chandra A, Loda M, Gillett C,
Cathcart P, Challacombe B, Cook G, Cahill D, Santa Olalla A, Cahill
F, et al: Metformin and longevity (METAL): A window of opportunity
study investigating the biological effects of metformin in
localised prostate cancer. BMC Cancer. 17:4942017. View Article : Google Scholar
|
|
8
|
Quinn BJ, Dallos M, Kitagawa H,
Kunnumakkara AB, Memmott RM, Hollander MC, Gills JJ and Dennis PA:
Inhibition of lung tumorigenesis by metformin is associated with
decreased plasma IGF-I and diminished receptor tyrosine kinase
signaling. Cancer Prev Res (Phila). 6:801–810. 2013. View Article : Google Scholar
|
|
9
|
El-Haggar SM, El-Shitany NA, Mostafa MF
and El-Bassiouny NA: Metformin may protect nondiabetic breast
cancer women from metastasis. Clin Exp Metastasis. 33:339–357.
2016. View Article : Google Scholar
|
|
10
|
Luo Q, Hu D, Hu S, Yan M, Sun Z and Chen
F: In vitro and in vivo anti-tumor effect of metformin as a novel
therapeutic agent in human oral squamous cell carcinoma. BMC
Cancer. 12:5172012. View Article : Google Scholar
|
|
11
|
Hosono K, Endo H, Takahashi H, Sugiyama M,
Sakai E, Uchiyama T, Suzuki K, Iida H, Sakamoto Y, Yoneda K, et al:
Metformin suppresses colorectal aberrant crypt foci in a short-term
clinical trial. Cancer Prev Res (Phila). 3:1077–1083. 2010.
View Article : Google Scholar
|
|
12
|
Tseng HW, Li SC and Tsai KW: Metformin
treatment suppresses melanoma cell growth and motility through
modulation of microRNA expression. Cancers (Basel). 11:2092019.
View Article : Google Scholar
|
|
13
|
Hwang AL, Haynes K, Hwang WT and Yang YX:
Metformin and survival in pancreatic cancer: A retrospective cohort
study. Pancreas. 42:1054–1059. 2013. View Article : Google Scholar
|
|
14
|
Podhorecka M, Ibanez B and Dmoszyńska A:
Metformin-its potential anti-cancer and anti-aging effects. Postepy
Hig Med Dosw (Online). 71:170–175. 2017. View Article : Google Scholar
|
|
15
|
Shi WY, Xiao D, Wang L, Dong LH, Yan ZX,
Shen ZX, Chen SJ, Chen Y and Zhao WL: Therapeutic metformin/AMPK
activation blocked lymphoma cell growth via inhibition of mTOR
pathway and induction of autophagy. Cell Death Dis. 3:e2752012.
View Article : Google Scholar
|
|
16
|
Wang CF, Zhang G, Zhao LJ, Qi WJ, Li XP,
Wang JL and Wei LH: Overexpression of the insulin receptor isoform
a promotes endometrial carcinoma cell growth. PLoS One.
8:e690012013. View Article : Google Scholar
|
|
17
|
He Y, Cao L, Wang L, Liu L, Huang Y and
Gong X: Metformin inhibits proliferation of human thyroid cancer
TPC-1 cells by decreasing LRP2 to suppress the JNK pathway. Onco
Targets Ther. 13:45–50. 2020. View Article : Google Scholar
|
|
18
|
Ling S, Xie H, Yang F, Shan Q, Dai H, Zhuo
J, Wei X, Song P, Zhou L, Xu X and Zheng S: Metformin potentiates
the effect of arsenic trioxide suppressing intrahepatic
cholangiocarcinoma: Roles of p38 MAPK, ERK3, and mTORC1. J Hematol
Oncol. 10:592017. View Article : Google Scholar
|
|
19
|
Vazquez-Martin A, Oliveras-Ferraros C and
Menendez JA: The antidiabetic drug metformin suppresses her2
(erbb-2) oncoprotein overexpression via inhibition of the mtor
effector p70s6k1 in human breast carcinoma cells. Cell Cycle.
8:88–96. 2009. View Article : Google Scholar
|
|
20
|
Saber S, Ghanim AMH, El-Ahwany E and
El-Kader EMA: Novel complementary antitumour effects of celastrol
and metformin by targeting IκBκB, apoptosis and NLRP3 inflammasome
activation in diethylnitrosamine-induced murine
hepatocarcinogenesis. Cancer Chemother Pharmacol. 85:331–343. 2020.
View Article : Google Scholar
|
|
21
|
Kowall B, Stang A, Rathmann W and Kostev
K: No reduced risk of overall, colorectal, lung, breast, and
prostate cancer with metformin therapy in diabetic patients:
Database analyses from Germany and the UK. Pharmacoepidemiol Drug
Saf. 24:865–874. 2015. View
Article : Google Scholar
|
|
22
|
Iliopoulos D, Hirsch HA and Struhl K:
Metformin decreases the dose of chemotherapy for prolonging tumor
remission in mouse xenografts involving multiple cancer cell types.
Cancer Res. 71:3196–3201. 2011. View Article : Google Scholar
|
|
23
|
Tyszka-Czochara M, Lasota M and Majka M:
Caffeic acid and metformin inhibit invasive phenotype induced by
TGF-β1 in C-4I and HTB-35/SiHa human cervical squamous carcinoma
cells by acting on different molecular targets. Int J Mol Sci.
19:2662018. View Article : Google Scholar
|
|
24
|
Xia C, Chen R, Chen J, Qi Q, Pan Y, Du L,
Xiao G and Jiang S: Combining metformin and nelfinavir exhibits
synergistic effects against the growth of human cervical cancer
cells and xenograft in nude mice. Sci Rep. 7:433732017. View Article : Google Scholar
|
|
25
|
Pollak MN: Investigating metformin for
cancer prevention and treatment: The end of the beginning. Cancer
Discov. 2:778–790. 2012. View Article : Google Scholar
|
|
26
|
Pollak M: The insulin and insulin-like
growth factor receptor family in neoplasia: An update. Nat Rev
Cancer. 12:159–169. 2012. View
Article : Google Scholar
|
|
27
|
Shackelford DB and Shaw RJ: The LKB1-AMPK
pathway: Metabolism and growth control in tumour suppression. Nat
Rev Cancer. 9:563–575. 2009. View
Article : Google Scholar
|
|
28
|
Foretz M, Carling D, Guichard C, Ferre P
and Foufelle F: AMP-activated protein kinase inhibits the
glucose-activated expression of fatty acid synthase gene in rat
hepatocytes. J Biol Chem. 273:14767–14771. 1998. View Article : Google Scholar
|
|
29
|
Choi YK and Park KG: Metabolic roles of
AMPK and metformin in cancer cells. Mol Cells. 36:279–287. 2013.
View Article : Google Scholar
|
|
30
|
Buzzai M, Jones RG, Amaravadi RK, Lum JJ,
DeBerardinis RJ, Zhao F, Viollet B and Thompson CB: Systemic
treatment with the antidiabetic drug metformin selectively impairs
p53-deficient tumor cell growth. Cancer Res. 67:6745–6752. 2007.
View Article : Google Scholar
|
|
31
|
Armando RG, Mengual Gómez DL and Gomez DE:
New drugs are not enough-drug repositioning in oncology: An update.
Int J Oncol. 56:651–684. 2020.
|
|
32
|
Datta SR, Brunet A and Greenberg ME:
Cellular survival: A play in three Akts. Genes Dev. 13:2905–2927.
1999. View Article : Google Scholar
|
|
33
|
Nitulescu GM, De Venter MV, Nitulescu G,
Ungurianu A, Juzenas P, Peng Q, Olaru OT, Grădinaru D, Tsatsakis A,
Tsoukalas D, et al: The Akt pathway in oncology therapy and beyond
(Review). Int J Oncol. 53:2319–2331. 2018.
|
|
34
|
Bellacosa A, Testa JR, Moore R and Larue
L: A portrait of AKT kinases: Human cancer and animal models depict
a family with strong individualities. Cancer Biol Ther. 3:268–275.
2004. View Article : Google Scholar
|
|
35
|
Cheng JQ, Lindsley CW, Cheng GZ, Yang H
and Nicosia SV: The Akt/PKB pathway: Molecular target for cancer
drug discovery. Oncogene. 24:7482–7492. 2005. View Article : Google Scholar
|
|
36
|
Shan ZZ, Chen PN, Wang F, Wang J and Fan
QX: Expression of P-EGFR and P-Akt protein in esophageal squamous
cell carcinoma and its prognosis. Oncol Lett. 14:2859–2863. 2017.
View Article : Google Scholar
|
|
37
|
Yu X, Yuan Y, Zhi X, Teng B, Chen X, Huang
Q, Chen Y, Guan Z and Zhang Y: Correlation between the protein
expression of A-kinase anchor protein 95, cyclin D3 and AKT and
pathological indicators in lung cancer tissues. Exp Ther Med.
10:1175–1181. 2015. View Article : Google Scholar
|
|
38
|
Yang SX, Costantino JP, Kim C, Mamounas
EP, Nguyen D, Jeong JH, Wolmark N, Kidwell K, Paik S and Swain SM:
Akt phosphorylation at Ser473 predicts benefit of paclitaxel
chemotherapy in node-positive breast cancer. J Clin Oncol.
28:2974–2981. 2010. View Article : Google Scholar
|
|
39
|
Lu C and Shervington A: Chemoresistance in
gliomas. Mol Cell Biochem. 312:71–80. 2008. View Article : Google Scholar
|
|
40
|
Jacobsen K, Bertran-Alamillo J, Molina MA,
Teixidó C, Karachaliou N, Pedersen MH, Castellví J, Garzón M,
Codony-Servat C, Codony-Servat J, et al: Convergent Akt activation
drives acquired EGFR inhibitor resistance in lung cancer. Nat
Commun. 8:4102017. View Article : Google Scholar
|
|
41
|
Nieminen AI, Eskelinen VM, Haikala HM,
Tervonen TA, Yan Y, Partanen JI and Klefström J: Myc-induced
AMPK-phospho p53 pathway activates Bak to sensitize mitochondrial
apoptosis. Proc Natl Acad Sci USA. 110:E1839–E1848. 2013.
View Article : Google Scholar
|
|
42
|
LoPiccolo J, Blumenthal GM, Bernstein WB
and Dennis PA: Targeting the PI3K/Akt/mTOR pathway: Effective
combinations and clinical considerations. Drug Resist Updat.
11:32–50. 2008. View Article : Google Scholar
|
|
43
|
Li Z, Dong H, Li M, Wu Y, Liu Y, Zhao Y,
Chen X and Ma M: Honokiol induces autophagy and apoptosis of
osteosarcoma through PI3K/Akt/mTOR signaling pathway. Mol Med Rep.
17:2719–2723. 2018.
|
|
44
|
Lee MS, Hsu CC, Wahlqvist ML, Tsai HN,
Chang YH and Huang YC: Type 2 diabetes increases and metformin
reduces total, colorectal, liver and pancreatic cancer incidences
in Taiwanese: A representative population prospective cohort study
of 800,000 individuals. BMC Cancer. 11:202011. View Article : Google Scholar
|
|
45
|
Evans JM, Donnelly LA, Emslie-Smith AM,
Alessi DR and Morris AD: Metformin and reduced risk of cancer in
diabetic patients. BMJ. 330:1304–1305. 2005. View Article : Google Scholar
|
|
46
|
Chen K, Li Y, Guo Z, Zeng Y, Zhang W and
Wang H: Metformin: Current clinical applications in nondiabetic
patients with cancer. Aging (Albany NY). 12:3993–4009. 2020.
View Article : Google Scholar
|
|
47
|
Quinn BJ, Kitagawa H, Memmott RM, Gills JJ
and Dennis PA: Repositioning metformin for cancer prevention and
treatment. Trends Endocrinol Metab. 24:469–480. 2013. View Article : Google Scholar
|
|
48
|
Li B, Zhou P, Xu K, Chen T, Jiao J, Wei H,
Yang X, Xu W, Wan W and Xiao J: Metformin induces cell cycle
arrest, apoptosis and autophagy through ROS/JNK signaling pathway
in human osteosarcoma. Int J Biol Sci. 16:74–84. 2020. View Article : Google Scholar
|
|
49
|
Zhou X, Kuang Y, Liang S and Wang L:
Metformin inhibits cell proliferation in SKM-1 cells via
AMPK-mediated cell cycle arrest. J Pharmacol Sci. 141:146–152.
2019. View Article : Google Scholar
|
|
50
|
Lee J, Hong EM, Kim JH, Jung JH, Park SW,
Koh DH, Choi MH, Jang HJ and Kae SH: Metformin induces apoptosis
and inhibits proliferation through the AMP-activated protein kinase
and insulin-like growth factor 1 receptor pathways in the bile duct
cancer cells. J Cancer. 10:1734–1744. 2019. View Article : Google Scholar
|
|
51
|
Pater MM and Pater A: Human papillomavirus
types 16 and 18 sequences in carcinoma cell lines of the cervix.
Virology. 145:313–318. 1985. View Article : Google Scholar
|
|
52
|
Donat U, Rother J, Schäfer S, Hess M,
Härtl B, Kober C, Langbein-Laugwitz J, Stritzker J, Chen NG,
Aguilar RJ, et al: Characterization of metastasis formation and
virotherapy in the human C33A cervical cancer model. PLoS One.
9:e985332014. View Article : Google Scholar
|
|
53
|
Hsieh Li SM, Liu ST, Chang YL, Ho CL and
Huang SM: Metformin causes cancer cell death through downregulation
of p53-dependent differentiated embryo chondrocyte 1. J Biomed Sci.
25:812018. View Article : Google Scholar
|
|
54
|
Xiao X, He Q, Lu C, Werle KD, Zhao RX,
Chen J, Davis BC, Cui R, Liang J and Xu ZX: Metformin impairs the
growth of liver kinase B1-intact cervical cancer cells. Gynecol
Oncol. 127:249–255. 2012. View Article : Google Scholar
|
|
55
|
Irie H, Banno K, Yanokura M, Iida M,
Adachi M, Nakamura K, Umene K, Nogami Y, Masuda K, Kobayashi Y, et
al: Metformin: A candidate for the treatment of gynecological
tumors based on drug repositioning. Oncol Lett. 11:1287–1293. 2016.
View Article : Google Scholar
|
|
56
|
Zhao B, Luo J, Wang Y, Zhou L, Che J, Wang
F, Peng S, Zhang G and Shang P: Metformin suppresses self-renewal
ability and tumorigenicity of osteosarcoma stem cells via reactive
oxygen species-mediated apoptosis and autophagy. Oxid Med Cell
Longev. 2019:92907282019. View Article : Google Scholar
|
|
57
|
Zhou XZ, Xue YM, Zhu B and Sha JP: Effects
of metformin on proliferation of human colon carcinoma cell line
SW-480. Nan Fang Yi Ke Da Xue Xue Bao. 30:1935–1938, 1942. 2010.(In
Chinese).
|
|
58
|
Kamarudin MNA, Sarker MMR, Zhou JR and
Parhar I: Metformin in colorectal cancer: Molecular mechanism,
preclinical and clinical aspects. J Exp Clin Cancer Res.
38:4912019. View Article : Google Scholar
|
|
59
|
Shaw RJ, Kosmatka M, Bardeesy N, Hurley
RL, Witters LA, DePinho RA and Cantley LC: The tumor suppressor
LKB1 kinase directly activates AMP-activated kinase and regulates
apoptosis in response to energy stress. Proc Natl Acad Sci USA.
101:3329–3335. 2004. View Article : Google Scholar
|
|
60
|
Ben Sahra I, Laurent K, Giuliano S,
Larbret F, Ponzio G, Gounon P, Le Marchand-Brustel Y,
Giorgetti-Peraldi S, Cormont M, Bertolotto C, et al: Targeting
cancer cell metabolism: The combination of metformin and
2-deoxyglucose induces p53-dependent apoptosis in prostate cancer
cells. Cancer Res. 70:2465–2475. 2010. View Article : Google Scholar
|
|
61
|
Cerezo M, Tichet M, Abbe P, Ohanna M,
Lehraiki A, Rouaud F, Allegra M, Giacchero D, Bahadoran P,
Bertolotto C, et al: Metformin blocks melanoma invasion and
metastasis development in AMPK/p53-dependent manner. Mol Cancer
Ther. 12:1605–1615. 2013. View Article : Google Scholar
|
|
62
|
Singh AR, Gu JJ, Zhang Q, Torka P,
Sundaram S, Mavis C and Hernandez-Ilizaliturri FJ: Metformin
sensitizes therapeutic agents and improves outcome in pre-clinical
and clinical diffuse large B-cell lymphoma. Cancer Metab. 8:102020.
View Article : Google Scholar
|
|
63
|
Chipuk JE, Kuwana T, Bouchier-Hayes L,
Droin NM, Newmeyer DD, Schuler M and Green DR: Direct activation of
Bax by p53 mediates mitochondrial membrane permeabilization and
apoptosis. Science. 303:1010–1014. 2004. View Article : Google Scholar
|
|
64
|
Chang YL, Lee HJ, Liu ST, Lin YS, Chen TC,
Hsieh TY, Huang HS and Huang SM: Different roles of p53 in the
regulation of DNA damage caused by1,2-heteroannelated
anthraquinones and doxorubicin. Int J Biochem Cell Biol.
43:1720–1728. 2011. View Article : Google Scholar
|
|
65
|
Fu YL, Zhang QH, Wang XW and He H:
Antidiabetic drug metformin mitigates ovarian cancer SKOV3 cell
growth by triggering G2/M cell cycle arrest and inhibition of
m-TOR/PI3K/Akt signaling pathway. Eur Rev Med Pharmacol Sci.
21:1169–1175. 2017.
|
|
66
|
Zhao Y, Sun H, Feng M, Zhao J, Zhao X, Wan
Q and Cai D: Metformin is associated with reduced cell
proliferation in human endometrial cancer by inbibiting
PI3K/AKT/mTOR signaling. Gynecol Endocrinol. 34:428–432. 2018.
View Article : Google Scholar
|
|
67
|
Zhang HH, Zhang Y, Cheng YN, Gong FL, Cao
ZQ, Yu LG and Guo XL: Metformin incombination with curcumin
inhibits the growth, metastasis, and angiogenesis of hepatocellular
carcinoma in vitro and in vivo. Mol Carcinog. 57:44–56. 2018.
View Article : Google Scholar
|
|
68
|
Storozhuk Y, Hopmans SN, Sanli T, Barron
C, Tsiani E, Cutz JC, Pond G, Wright J, Singh G and Tsakiridis T:
Metformin inhibits growth and enhances radiation response of
non-small cell lung cancer (NSCLC) through ATM and AMPK. Br J
Cancer. 108:2021–2032. 2013. View Article : Google Scholar
|
|
69
|
Dhillon SS, Groman A, Meagher A, Demmy T,
Warren GW and Yendamuri S: Metformin and not diabetes influences
the survival of resected early stage NSCLC patients. J Cancer Sci
Ther. 6:217–222. 2014.
|
|
70
|
Sayed R, Saad AS, ElWakeel L, Elkholy E
and Badary O: Metformin addition to chemotherapy in stage IV
non-small cell lung cancer: An open label randomized controlled
study. Asian Pac J Cancer Prev. 16:6621–6626. 2015. View Article : Google Scholar
|