Introduction
Atherosclerosis (AS) is a common vascular pathogenic
condition characterized by artery wall thickening and reduced
elasticity due to the formation of atherosclerotic plaques (fatty
deposits) in arterial intima. The plaques are primarily composed of
cholesterol, fat, calcium and other constitutes of human blood
(1,2). The hardening of plaques with time
usually results in arterial stenosis and reduces tissue blood
supply, which can lead to a number of severe conditions, such as
coronary heart disease, carotid artery disease, peripheral artery
disease, chronic kidney disease, stroke, angina and even death
(2,3). Currently, the clinical management of
AS greatly depends on altering the patient's lifestyle, the use of
statin medications and even surgical treatments, such as
percutaneous coronary intervention and coronary artery bypass
grafting, for patients with severe AS (2–4). A
previous study demonstrated that major risk factors for AS include
high low density lipoprotein (LDL) levels, hypertension, insulin
resistance, obesity, diabetes, inflammation and unhealthy lifestyle
habits, such as smoking and a high-saturated fat diet (5). However, the molecular mechanisms
underlying plaque formation and AS pathogenesis are not completely
understood.
The abnormal function of vascular smooth muscle
cells (VSMCs) is closely associated with AS development (6–8).
Accumulating evidence has suggested that VSMCs undergo significant
phenotypic alterations that drive plaque formation and AS
progression. Among those alterations are increased capacities for
cell proliferation and migration, a reduced rate of VSMC apoptosis,
and alterations in various extracellular matrix proteins and
secreted cytokines (6,9). For example, VSMCs in healthy vascular
vessel walls usually have extremely low turnover and proliferation
rates; however, VSMC proliferation is greatly increased during the
early stage of atherogenesis and in response to a vascular injury
(6). A previous report revealed
that high glucose-induced advanced glycosylation end products could
remarkably promote VSMC proliferation and apoptosis, resulting in
vein graft AS via activation of MAPK signaling pathways (10). Moreover, a previous investigation
suggested that suppression of VSMC proliferation, inflammatory
responses and adhesion by vasostatin-2 overexpression might serve
as a novel therapeutic strategy for AS (11). Collectively, the aforementioned
studies indicated that VSMC phenotypic switching is linked to
plaque formation and AS development, suggesting that inhibiting the
phenotype switch of VSMCs might serve as a promising therapeutic
strategy for AS.
MicroRNAs (miRNAs/miRs) are a large group of
single-stranded non-coding RNA molecules that consist of 19–25
nucleotides, and serve as regulators of gene expression by
modulating mRNA degradation and interfering with protein
translation (12–14). miRNAs are closely linked to various
pathogenic processes, such as cancer, neural degeneration, diabetic
nephropathy and vascular diseases (13–18).
More importantly, miRNAs also serve essential pathogenic roles
during AS development and progression (19,20). A
previous study demonstrated that miR-140-5p could inhibit the
angiogenesis process in a rat model of middle cerebral artery
occlusion by regulating vascular endothelial growth factor (VEGF)A
expression (21). Furthermore,
miR-140-5p inhibits glioma cell proliferation and invasion by
modulating the VEGF and matrix metallopeptidase 2 (MMP2) signaling
pathways (22). Liu et al
(23) reported that miR-140-5p
aggravates AS by mediating oxidative stress responses by targeting
nuclear factor, erythroid 2 like 2 and sirtuin 2 via the kelch like
ECH associated protein 1/heme oxygenase 1 signaling pathway
(23). Another study demonstrated
that miR-140-5p serves a key role in pulmonary arterial
hypertension by targeting SMAD-specific E3 ubiquitin protein ligase
1 (24). Despite the widespread
expression of microRNAs, the current understanding of the
association between microRNAs and AS pathogenesis is limited.
A bioinformatics analysis predicted that miR-140-5p
might target the roundabout guidance receptor 4 (ROBO4) gene, which
codes for a member of the neuronal Robo family that serve as
vascular-specific receptors, and are involved in angiogenesis and
vascular permeability regulation (25–27).
ROBO4 expression is mediated by methylation of its promoter, and
has been demonstrated to serve a key role in endothelial cell
differentiation (28). Moreover, a
previous study indicated that ROBO4 could modulate microvascular
endothelial cell migration during angiogenesis (29,30).
However, how miR-140-5p might affect ROBO4 gene expression during
the AS pathogenesis process, and the roles served by miR-140-5p in
VSMCs and AS development are not completely understood.
The present study aimed to investigated the role of
ROBO4 and miR-140-5p in an oxidized LDL (ox-LDL)-induced cell
model. The mechanism was identified by investigating the expression
levels of miR-140-5p and ROBO4 expression in AS tissues and human
VSMCs treated with ox-LDLs, as well as the effects of ox-LDL
treatment on VSMC viability, migration, invasion and apoptosis.
This study provided a novel perspective on the molecular
pathogenesis of AS, and may aid in developing novel strategies for
the prevention and treatment of the disease.
Materials and methods
Tissues, cell culture and
treatments
Artery tissues containing AS plaque were collected
from patients who had been diagnosed with AS (n=20; age, 48–71
years; 55% men and 45% women) and undergone surgical treatment in
the Department of Vascular Surgery of The First Affiliated Hospital
of Chongqing Medical University between July 2017 and December
2018. Samples of AS tissue were obtained from patients with lower
extremity atherosclerotic occlusive disease who were undergoing
surgery, and samples of healthy control tissue were obtained from
individuals who had suffered an injury, such as amputation (20%),
fracture (70%) or other (10%). The present study was reviewed and
approved by the Medical Ethics Committee of The First Affiliated
Hospital of Chongqing Medical University. Written informed consent
was obtained from each participant.
Human VSMCs were purchased from the BeNa Culture
Collection (cat. no. BNCC340087) and cultured in high-glucose DMEM
(cat. no. 12100046; Gibco; Thermo Fisher Scientific, Inc.)
containing 10% FBS (cat. no. 10099-141; Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin, and 100 U/ml streptomycin
at 37°C in a humidified cell culture chamber containing 5% CO2. All
cells were used within 7 passages and stored in liquid nitrogen
prior to use. Cells (5×105 cells/ml) were treated with
50 or 100 µg/ml ox-LDL (cat. no. YB-002; Guangzhou Yiyuan
Biological Technology Co., Ltd.) for 24 h at 37°C. Cells treated
with PBS were set as the control group.
Cell transfection
An miR-140-5p inhibitor
(3′-GUCACCAAAACGGGAUACCAUC-5′) and its negative control (NC,
5′-UUCUCCGAACGUGUCACGUTT-3′) were synthesized by Shanghai
GenePharma Co., Ltd. and used to induce miR-140-5p knockdown. VSMCs
(1×105 cells/ml) were seeded into 96-well plates and
cultured overnight at 37°C. Subsequently, VSMCs were incubated with
25 µl serum-free DMEM containing 5 pmol miR-140-5p inhibitor or NC
and 0.25 µl Lipofectamine® 2000 Transfection Reagent
(cat. no. 11668027; Thermo Fisher Scientific, Inc.) at 37°C
according to the manufacturer's protocol. To induce ROBO4
overexpression, ROBO4 genomic sequences were amplified by PCR using
specific primers (forward, 5′-GGGGTACCATGGTGGCTGTGGTGGGTGAGC-3′ and
reverse, 5′-CCAAGCTTTCAGGAGTAATCTACAGGAGAAGCACCAGC-3′) and inserted
into pcDNA3.0 plasmids (Biovector Science Lab, Inc.). VSMCs were
transfected with recombinant pcDNA3.0-ROBO4 plasmids (1.5 µg) using
Lipofectamine 2000 Transfection Reagent at 37°C. At 24 h
post-transfection, transfection efficiency was assessed via reverse
transcription-quantitative PCR (RT-qPCR). An empty vector was
transfected into VSMCs as a negative control.
RT-qPCR
The relative expression levels of miR-140-5p and
ROBO4 mRNA in cultured VSMCs and artery tissues were analyzed via
RT-qPCR. Total RNA was isolated from cultured cells or tissues
using TRIzol® reagent (cat. no. 9109; Takara Bio, Inc.)
according to the manufacturer's protocol. Subsequently, total RNA
(2.0 µg) was reverse transcribed into cDNA using the Bestar qPCR RT
kit (cat. no. 2220; DBI Bioscience) according to the manufacturer's
protocol. Subsequently, qPCR was performed using a Stratagene
Mx3000P real-time PCR system (Agilent Technologies, Inc.) and
Bestar™ qPCR MasterMix (cat. no. 2043; DBI Bioscience) according to
the manufacturer's instructions. The following thermocycling
conditions were used for qPCR: 40 cycles of 94°C for 2 min,
followed by 94°C for 20 sec, 58°C for 20 sec and 72°C for 20 sec.
The sequences of the primers used for qPCR are presented in
Table I. miRNA and mRNA expression
levels were quantified using the 2−∆∆Cq method (31) and normalized to the internal
reference genes U6 and GAPDH, respectively.
 | Table I.Primer sequences used for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences used for reverse
transcription-quantitative PCR.
| Gene | Sequences
(5′→3′) | Product length
(bp) |
|---|
| GAPDH | F:
TGTTCGTCATGGGTGTGAAC | 131 |
|
| R:
ATGGCATGGACTGTGGTCAT |
|
| Bcl-2 | F:
GAGGATTGTGGCCTTCTTTG | 170 |
|
| R:
ACAGTTCCACAAAGGCATCC |
|
| MMP2 | F:
ATGACAGCTGCACCACTGAG | 174 |
|
| R:
ATTTGTTGCCCAGGAAAGTG |
|
| ROBO4 | F:
GTGGTGGGTGAGCAGTTTAC | 270 |
|
| R:
CATGTAGGTCCCTTCGTCAC |
|
| miR-140-5p | F:
ACACTCCAGCTGGGTGGTGGTTTTACCCTATGGT | N/A |
|
| R:
CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCTACCAT |
|
| U6 | F:
CTCGCTTCGGCAGCACA | N/A |
|
| R:
AACGCTTCACGAATTTGCGT |
|
Western blotting
Total protein was extracted from cultured VSMC cells
using RIPA Lysis Buffer (Thermo Fisher Scientific, Inc.) according
to the manufacturer's protocol. Total protein was quantified using
the bicinchoninic acid method. Protein (~20 µg) was separated via
10% SDS-PAGE and transferred onto PVDF membranes (EMD Millipore),
which were then blocked with 5% lipid-free milk solution for 2 h at
room temperature. Subsequently, the membranes were incubated for
1–2 h at room temperature with primary antibodies targeted against:
ROBO4 (cat. no. ab180824; 1:1,000; Abcam) and GAPDH (cat. no.
ab8245; 1:5,000; Abcam). Following primary incubation, the
membranes were incubated with rabbit anti-mouse HPR-conjugated
secondary antibodies (cat. no. ab6728; 1:3,000; Abcam) for 2 h at
room temperature. Immunoreactive bands were visualized using an
Enhanced Chemiluminescence Substrate kit (cat. no. KLS0500; Merck
KGaA). GAPDH was used as the loading control. Image-Pro Plus
software (version 6.0; Media Cybernetics, Inc.) was used for
semi-quantification.
Cell Counting Kit-8 (CCK-8) assay
VSMC viability was evaluated using a CCK-8 assay
(cat. no. CK04; Dojindo Molecular Technologies, Inc.) according to
the manufacturer's protocol. Briefly, cells (1×105 cells/well) in a
96-well plate were incubated in a humidified incubator at 37°C for
24 h. Subsequently, 10 µl CCK-8 solution was added to each well and
incubated for 24, 48 or 72 h at 37°C. The absorbance of each well
was measured at a wavelength of 450 nm (optical
density450) using a microplate reader to calculate the
cell viability rate.
Cell migration and invasion
VSMC migration and invasion were detected using a
Transwell culture system. To assess cell migration, VSMCs
(2×104) were re-suspended in serum-free DMEM containing
0.2% BSA (cat. no. ST025; Beyotime Institute of Biotechnology) and
seeded into the upper chambers. The lower chambers were filled with
700 µl complete DMEM containing 10% FBS. Following incubation at
37°C for 48 h, cells in the lower chambers were fixed with 4%
paraformaldehyde (cat. no. P0099; Beyotime Institute of
Biotechnology), stained with crystal violet staining solution for
5–10 min at room temperature and counted using a CX41 light
microscope (Olympus Corporation). To detect cell invasion, the
inner sides of the Transwell chambers were pre-coated with
Matrigel® Basement Membrane Matrix (cat. no. 356234BD;
Biocoat, Inc.) at room temperature, after which, the aforementioned
procedure was performed to assess cell invasion.
Cell apoptosis
VSMC apoptosis was detected by using an Annexin
V-FITC/PI kit (Nanjing KeyGen Biotech Co., Ltd.) in combination
with flow cytometry. Briefly, cultured VSMCs were seeded
(4×105 cells/well) into a 6-well plate and treated with
ox-LDL. After treatment, cells were collected and incubated with
Annexin V-FITC solution (1 ml) for 12 min in the dark at room
temperature, and incubated with propidium iodide solution (5 µl)
for 5 min in the dark. Subsequently, the percentage of apoptotic
VSMCs was determined by flow cytometry (BD FACSCalibur™; BD
Biosciences) using FlowJo™ version 10 software (FlowJo LLC). Early
+ late apoptosis rate was regarded as total apoptosis
percentage.
Immunofluorescence
VSMCs were fixed with 4% paraformaldehyde for 30 min
at room temperature followed by permeabilization with 0.3% Triton
X-100 for 15 min. After permeabilization, cells were washed with
PBS and blocked with 5% goat serum (cat. no. C0265; Beyotime
Institute of Biotechnology) for 30 min at room temperature, and
subsequently incubated with an anti-α-smooth muscle actin (SMA)
primary antibody (cat no. BM3902; 1:100; Boster Biological
Technology) overnight at 4°C. Subsequently, cells were incubated
with a secondary Cy5-labeled goat anti-rabbit antibody (cat no.
ab97077; 1:300; Abcam). Cells were stained with DAPI at room
temperature for 10 min for nuclear staining and observed using a
fluorescence microscope (magnification, ×400).
Construction of vectors and
transfection
The potential binding site of miR-140-5p on the
3′UTR of ROBO4 was predicted using TargetScan (http://www.targetscan.org/vert_72/) (32). siRNA against ROBO4
(5′-CCAAGACUACGAGUUCAAA-3′) and the NC (5′-AUUCGACGCAUGACACAAA-3′)
were purchased from Shanghai GenePharma Co., Ltd. siRNA and NC were
used at concentration of 20 µM diluted with diethyl pyrocarbonate
H2O. DNA was extracted from the VSMCs using PrimeSTAR®
HS DNA Polymerase (Takara Bio, Inc.). The following primers were
used to amplify the 3′-untranslated region (UTR) sequences from
VSMCs by PCR: Wild-type (WT) 3′UTR forward,
5′-CCCTCGAGACCGTGTCCCTGAGACTTCCC-3′ and reverse,
5′-ATTTGCGGCCGCTAGGAGTCAGGTGGAGATGATGTTT-3′. The PCR process was
according to the information included in Table II. The resulting PCR product was
ligated into psi-CHECK2 vectors (cat. no. 97157; Addgene, Inc.).
Point mutations were introduced by performing PCR with the
following primers: forward,
5′-CTTCCCAGACGGGAATCAGCTTACTGTCTCCTGTCCACCCACAAG-3′ and reverse,
5′-ACTTGTGGGTGGACAGGAGACAGTAAGCTGATTCCCGTCTGGGAAG-3′. Similarly,
the resulting PCR products were inserted into psi-CHECK2 vectors.
The coding sequences of ROBO4 were cloned from VSMCs using the
following primers: forward, 5′-GGGGTACCATGGTGGCTGTGGTGGGTGAGC-3′
and reverse, 5′-CCAAGCTTTCAGGAGTAATCTACAGGAGAAGCACCAGC-3′. All PCR
products were identified by sequencing conducted by Sangon Biotech
Co., Ltd. Plasmid (~10 µg) was transfected into VSMCs
(1×105 per ml). Transfections were performed using
Lipofectamine 3000 (Thermo Fisher Scientific, Inc.) according to
manufacturer's instructions. All primers used were synthesized by
Sangon Biotech Co., Ltd. Subsequent assays were performed 24 h
after transfection.
| Table II.PCR amplification procedure. |
Table II.
PCR amplification procedure.
| Reagent | Volume (µl) |
|---|
|
PrimeSTAR® HS DNA
Polymerase | 0.5 |
| 5X PrimeSTAR
buffer | 5.0 |
| dNTP Mix (2.5 mM
each) | 2.0 |
| F (10 µM) | 0.5 |
| R (10 µM) | 0.5 |
| cDNA | 1.0 |
|
ddH2O | ≤25 |
| Total volume | 25 |
Dual-luciferase reporter assay
The binding of miR-140-5p to ROBO4 mRNA was verified
by performing a dual-luciferase reporter assay using a
Dual-Luciferase Reporter Assay System (cat. no. E1910; Promega
Corporation) according to the manufacturer's instructions. Briefly,
the WT and mutant (MUT) 3′UTR ROBO4 gene sequences were separately
inserted into psi-CHECK2 plasmids (cat. no. 97157; Addgene, Inc.).
miR-140-5p mimic (5′-CAGUGUUUUACCCUAUGGUAG-3′) and its NC
(5′-UUCUCCGAACGUGUCACGUTT-3′) were synthesized by Shanghai
GenePharma Co., Ltd. VSMCs were transfected with the WT or MUT
recombinant plasmids (8 µg) and miR-140-5p mimic or the NC (100
ppm) using Lipofectamine 2000 Transfection Reagent (Thermo Fisher
Scientific, Inc.). Luciferase activity was detected using a GloMax
20/20 Luminometer (Promega Corporation). Renilla luciferase
activity was set as the internal reference.
Statistical analysis
Statistical analyses were performed using SPSS
software (version 20.0; IBM Corp). Data are presented as the mean ±
SD. All experiments were repeated three times. Comparisons among
groups were analyzed using the Student's t test or non-parametric
Kruskal-Wallis followed by the Dunn's post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
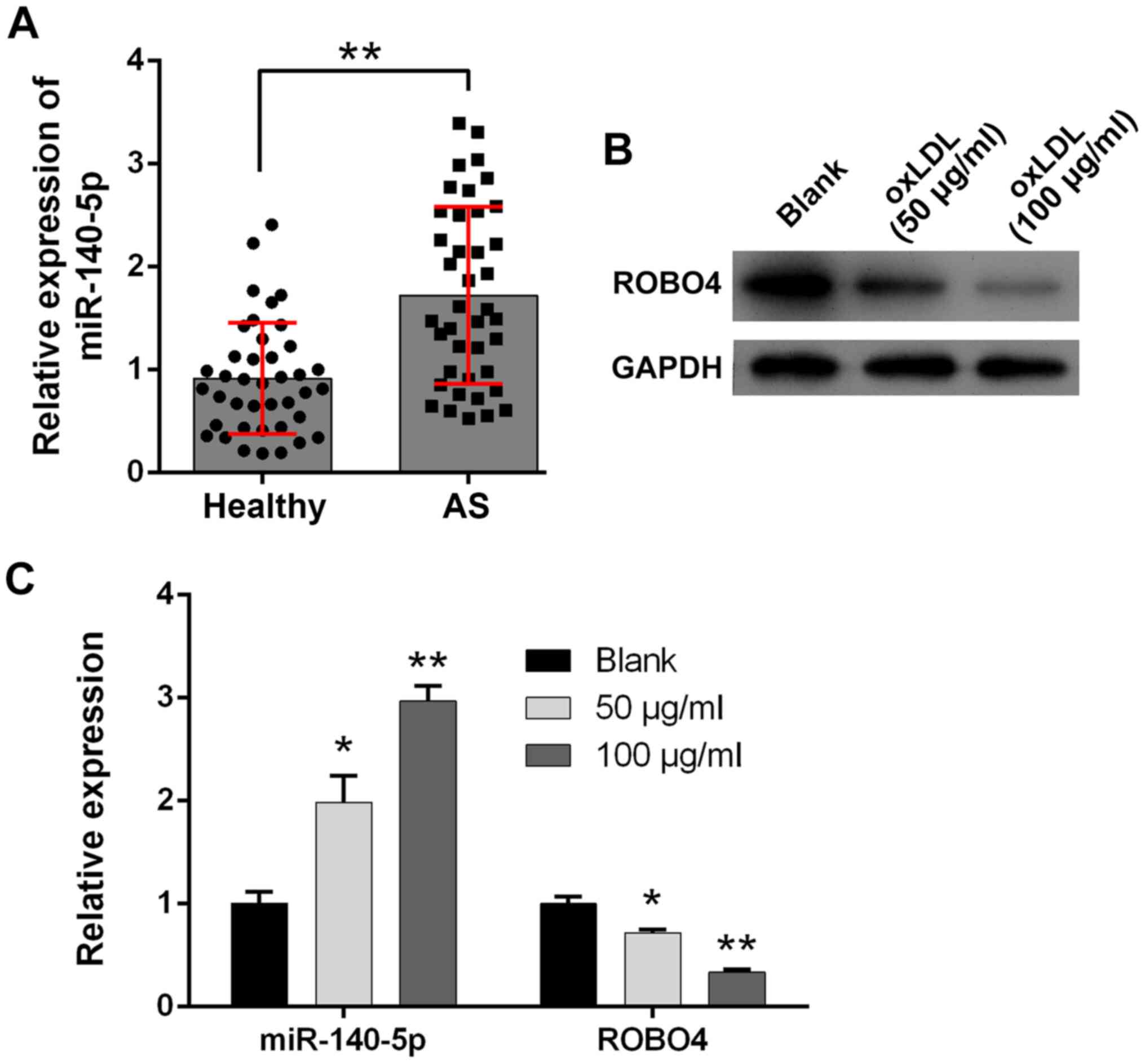
Increased miR-140-5p expression and
decreased ROBO4 expression during AS
To investigate the potential pathogenic roles of
miR-140-5p during AS development, artery tissues containing plaque
were collected from 20 patients who had been diagnosed with AS and
healthy artery tissues were collected from 20 healthy volunteers
who served as healthy control subjects. miR-140-5p was expressed at
significantly higher levels in the artery tissues of patients with
AS compared with healthy control subjects (Fig. 1A). Moreover, cultured human VSMCs
were treated with ox-LDL to establish a cellular model of AS
development. A bioinformatics analysis predicted ROBO4 as a target
of miR-140-5p. In the present study, ROBO4 protein expression
levels in human VSMCs were notably reduced by ox-LDL treatment in a
concentration-dependent manner compared with the blank control
group (Fig. 1B). Furthermore, the
RT-qPCR results indicated that miR-140-5p expression was
significantly increased and ROBO4 gene expression was significantly
decreased in ox-LDL-treated human VSMCs compared with blank control
VSMCs (Fig. 1C).
miR-140-5p inhibition suppresses VSMC
viability, migration and invasion, and promotes apoptosis following
ox-LDL treatment
To analyze the cellular functions of miR-140-5p
during AS development, cultured human VSMCs were transfected with
miR-140-5p inhibitor. VSMCs were identified by detecting α-SMA
expression via immunofluorescence. The results suggested that the
VSMCs expressed α-SMA (Fig. 2A).
miR-140-5p expression levels in ox-LDL-treated human VSMCs were
significantly downregulated by miR-140-5p inhibitor compared with
the NC group (Fig. 2B).
Furthermore, ROBO4 protein expression levels in ox-LDL-treated
VSMCs were significantly downregulated compared with control VSMCs
(Fig. 2C). ROBO4 protein expression
levels in ox-LDL-treated human VSMCs were upregulated by
transfection with miR-140-5p inhibitor (Fig. 2D). The RT-qPCR results indicated
that Bcl-2 and MMP2 expression levels were significantly higher in
ox-LDL-treated VSMCs compared with control VSMCs, which was
reversed by miR-140 inhibitor (Fig.
2E). Moreover, human VSMC viability was greatly increased by
ox-LDL treatment, but ox-LDL-mediated alterations to cell viability
were reversed by miR-140-5p inhibitor (Fig. 2F). Similarly, the number of
migratory and invading VSMCs was significantly increased following
ox-LDL treatment compared with control VSMCs, and ox-LDL-mediated
increases were attenuated by miR-140-5p inhibitor (Fig. 2G). By contrast, the number of
apoptotic VSMCs was significantly decreased by ox-LDL treatment
compared with control VSMCs, but restored by transfection with
miR-140-5p inhibitor (Fig. 2H).
Collectively, the results indicated that human VSMC viability,
migration and invasion were enhanced, whereas VSMC apoptosis was
suppressed in ox-LDL-treated VSMCs; however, miR-140-5p inhibitor
reversed ox-LDL-mediated effects.
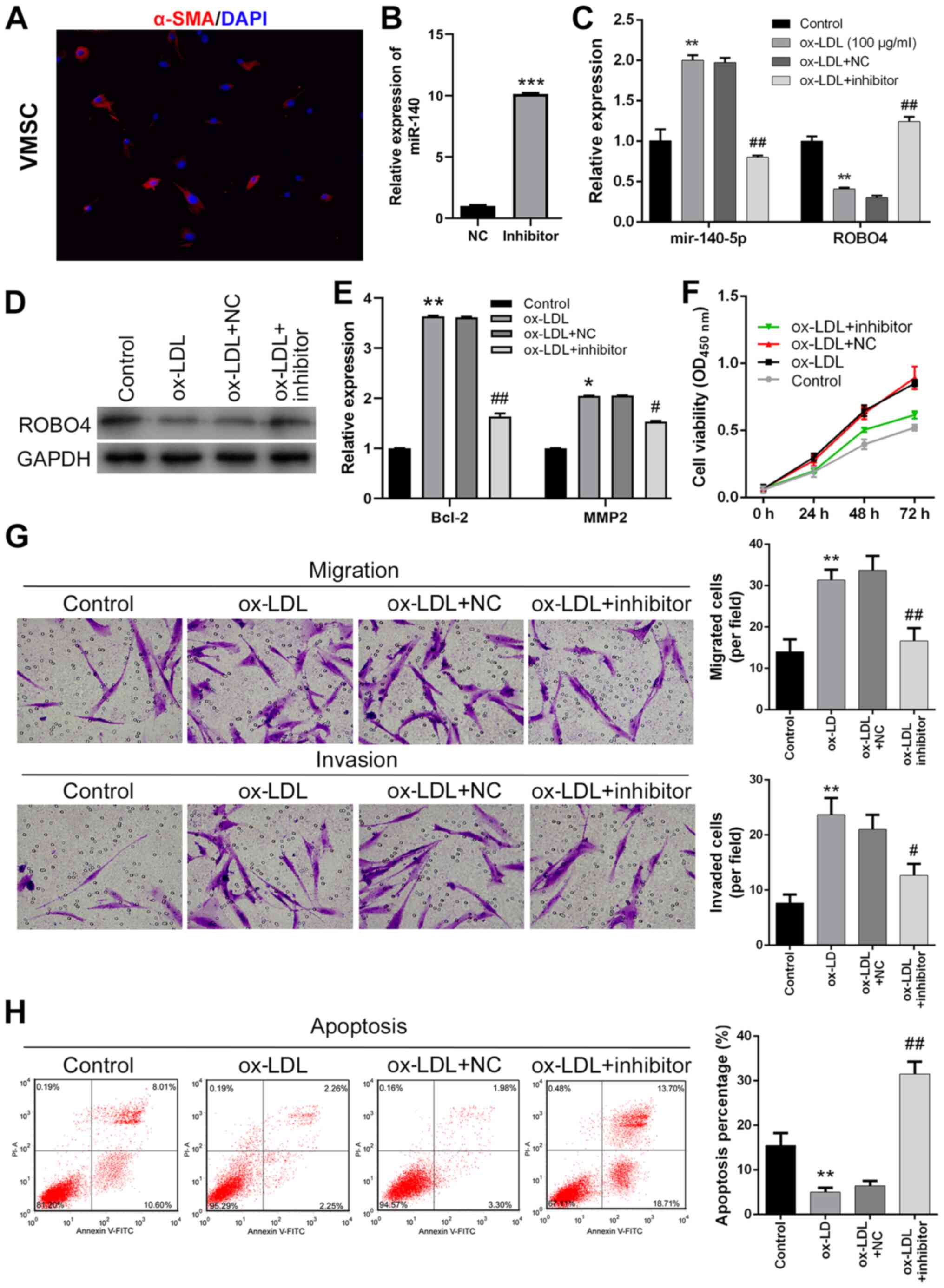 | Figure 2.Effect of miR-140-5p knockdown and
ox-LDL treatment on VSMC viability, migration, invasion and
apoptosis. (A) VSMCs were identified via immunofluorescence
(magnification, ×200). (B) Transfection efficiency of miR-140-5p
inhibitor. (C) miR-140-5p and ROBO4 expression levels in miR-140-5p
inhibitor-transfected and ox-LDL-treated VMSCs. (D) ROBO4 protein
expression levels in miR-140-5p inhibitor-transfected and
ox-LDL-treated human VSMCs (E) Bcl-2 and MMP2 expression levels
were measured by reverse transcription-quantitative PCR. Human VSMC
(F) viability, (G) migration, invasion (magnification, ×200) and
(H) apoptosis following miR-140-5p inhibitor transfection and
ox-LDL treatment. *P<0.05, **P<0.01, ***P<0.001 vs. NC or
control; #P<0.05, ##P<0.01 vs. ox-LDL +
NC. miR, microRNA; ox-LDL, oxidized-low density lipoprotein; VSMC,
vascular smooth muscle cell; ROBO4, roundabout guidance receptor 4;
MMP2, matrix metallopeptidase 2; α-SMA, α-smooth muscle actin; NC,
negative control; OD, optical density. |
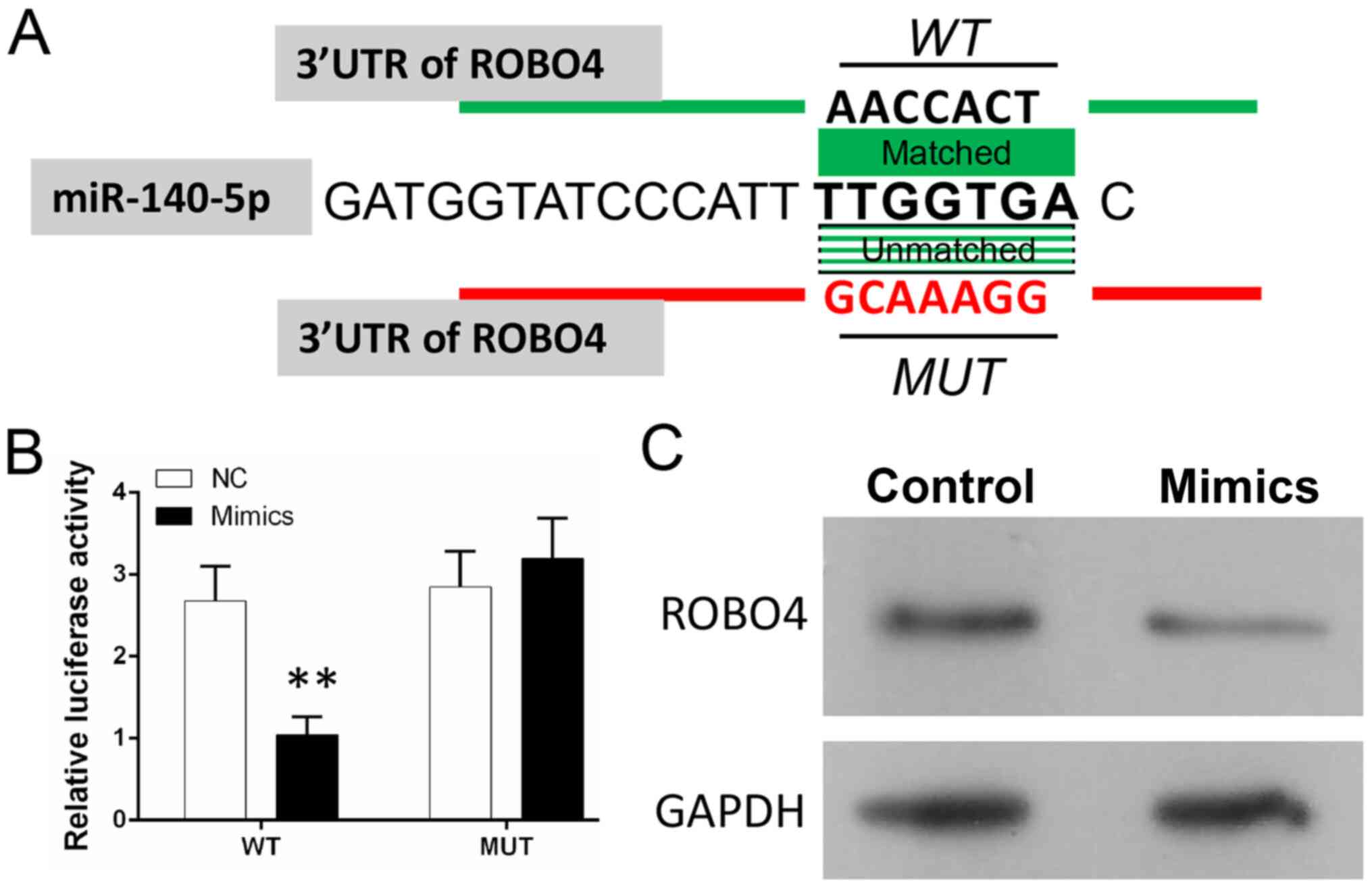
miR-140 directly targets the ROBO4
gene in VSMCs
A bioinformatics analysis predicted that ROBO4 might
be a target gene of miR-140-5p by binding to the 3′-UTR of ROBO4
mRNA (Fig. 3A). To investigate the
relationship between miR-140-5p and ROBO4, a dual-luciferase
reporter assay was conducted to assess the binding between
miR-140-5p and the 3′-UTR of ROBO4 mRNA (Fig. 3B). The results suggested that the
luciferase activity of WT-VSMCs was significantly decreased in
miR-140-5p mimic-transfected cells compared with the NC group
(Fig. 3B). However, the luciferase
activity of MUT-VSMCs was not significantly altered in miR-140-5p
mimic-transfected cells compared with the NC group (Fig. 3B). The results indicated that
miR-140-5p could bind to the 3′UTR of ROBO4 mRNA in VSMCs and
verified ROBO4 as a target of miR-140-5p. Furthermore, miR-140-5p
mimic downregulated ROBO4 expression levels compared with the NC
group (Fig. 3C). Therefore, the
results indicated that the relationship between ROBO4 and
miR-140-5p may be associated with the ability of miR-140-5p to
regulate AS pathogenesis.
ROBO4 overexpression reverses
ox-LDL-mediated effects on VSMCs
To explore the pathogenic roles of
miR-140-5p-mediated inhibition of ROBO4 gene expression,
ROBO4-overexpression human VSMCs were established. The results
indicated that ROBO4 expression was significantly increased by
transfection with the ROBO4-overexpression vector compared with the
NC group (Fig. 4A). ROBO4
expression levels were markedly increased in ROBO4-overexpression
ox-LDL-treated human VSMCs compared with empty vector-transfected
ox-LDL-treated human VSMCs (Fig. 4B and
C). Bcl-2 and MMP2 expression levels were significantly
increased by ox-LDL treatment compared with control cells, but
ROBO4 overexpression reversed ox-LDL-mediated alterations (Fig. 4D). In addition, ox-LDL-induced VSMC
viability was notably decreased by ROBO4 overexpression (Fig. 4E). Similarly, ox-LDL-induced VSMC
migration and invasion were significantly reduced by ROBO4
overexpression (Fig. 4F).
Furthermore, the inhibitory effect of ox-LDL treatment on human
VSMC apoptosis was also reversed by ROBO4 overexpression, which
resulted in significantly increased VSMC apoptosis compared with
ox-LDL-treated cells (Fig. 4G).
Collectively, the results indicated that ox-LDL-mediated effects on
human VSMC viability, migration, invasion and apoptosis could be
attenuated by ROBO4 gene overexpression.
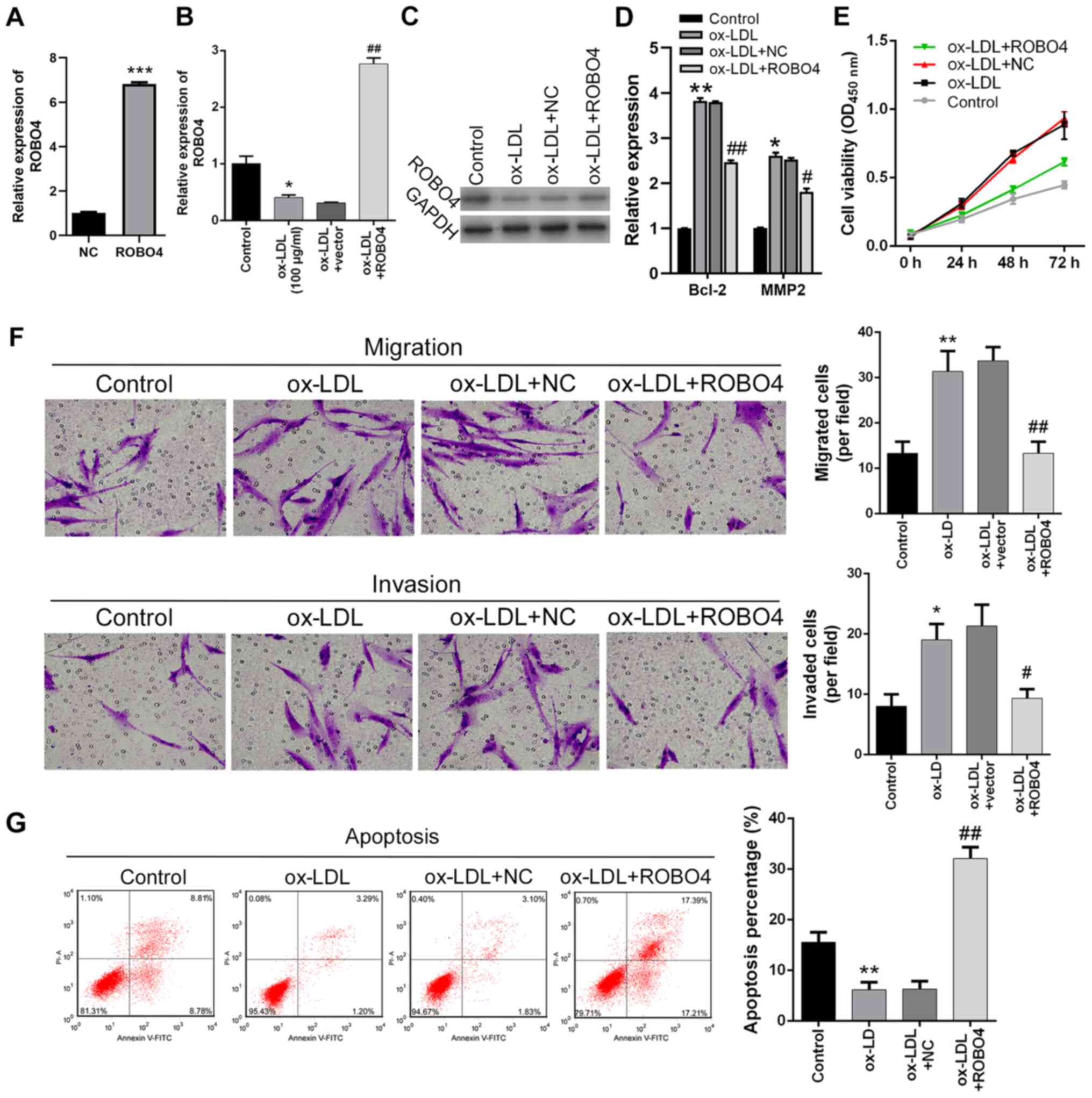 | Figure 4.Effects of ROBO4 overexpression and
ox-LDL treatment on VSMC viability, migration, invasion and
apoptosis. (A) Transfection efficiency of ROBO4 overexpression.
ROBO4 (B) mRNA and (C) protein expression levels in human VSMCs
following ROBO4 overexpression and ox-LDL treatment. (D) Bcl-2 and
MMP2 expression levels were measured by reverse
transcription-quantitative PCR. Effect of ROBO4 overexpression and
ox-LDL treatment on human VSMC (E) viability, (F) migration,
invasion (magnification, ×200) and (G) apoptosis. *P<0.05,
**P<0.01, ***P<0.001 vs. NC or control;
#P<0.05, ##P<0.01 vs. ox-LDL + NC.
ROBO4, roundabout guidance receptor 4; ox-LDL, oxidized-low density
lipoprotein; VSMC, vascular smooth muscle cell; MMP2, matrix
metallopeptidase 2; NC, negative control; OD, optical density. |
miR-140-5p mediates ox-LDL-induced
VSMC phonotypes by targeting ROBO4
miR-140-5p inhibitor and ROBO4 siRNA were used to
further investigate the role of miR-140-5p in VSMCs. The
transfection efficiency of ROBO4 siRNA was assessed via RT-qPCR,
which indicated that ROBO4 siRNA significantly downregulated ROBO4
expression levels compared with the NC group (Fig. 5A). ROBO4 expression was
significantly reduced by treatment with ox-LDL compared with the
control group, an effect that was partially reversed by miR-140-5p
inhibitor. Moreover, ROBO4 expression in ox-LDL- and
inhibitor-treated VSMCs was significantly decreased by treatment
with ROBO4 siRNA (Fig. 5B). Bcl-2
and MMP2 expression levels were significantly increased in
ox-LDL-treated cells compared with control cells, but treatment
with miR-140-5p inhibitor significantly decreased ox-LDL-induced
expression. In addition, Bcl-2 and MMP2 expression levels in
ox-LDL- and inhibitor-treated VSMCs were significantly increased by
ROBO4 siRNA (Fig. 5C). Compared
with the control group, VSMC apoptosis was significantly decreased
by ox-LDL treatment, but rescued by treatment with miR-140-5p
inhibitor. ROBO4 siRNA significantly decreased the rate of
apoptosis in ox-LDL and inhibitor-treated cells (Fig. 5D). The Transwell assays indicated
that the migration and invasion of ox-LDL-treated VSMCs were
significantly enhanced compared with the control group, but
miR-140-5p inhibitor suppressed ox-LDL-induced migration and
invasion. By contrast, ROBO4 siRNA significantly enhanced the
migration and invasion of ox-LDL- and inhibitor-treated cells
(Fig. 5E).
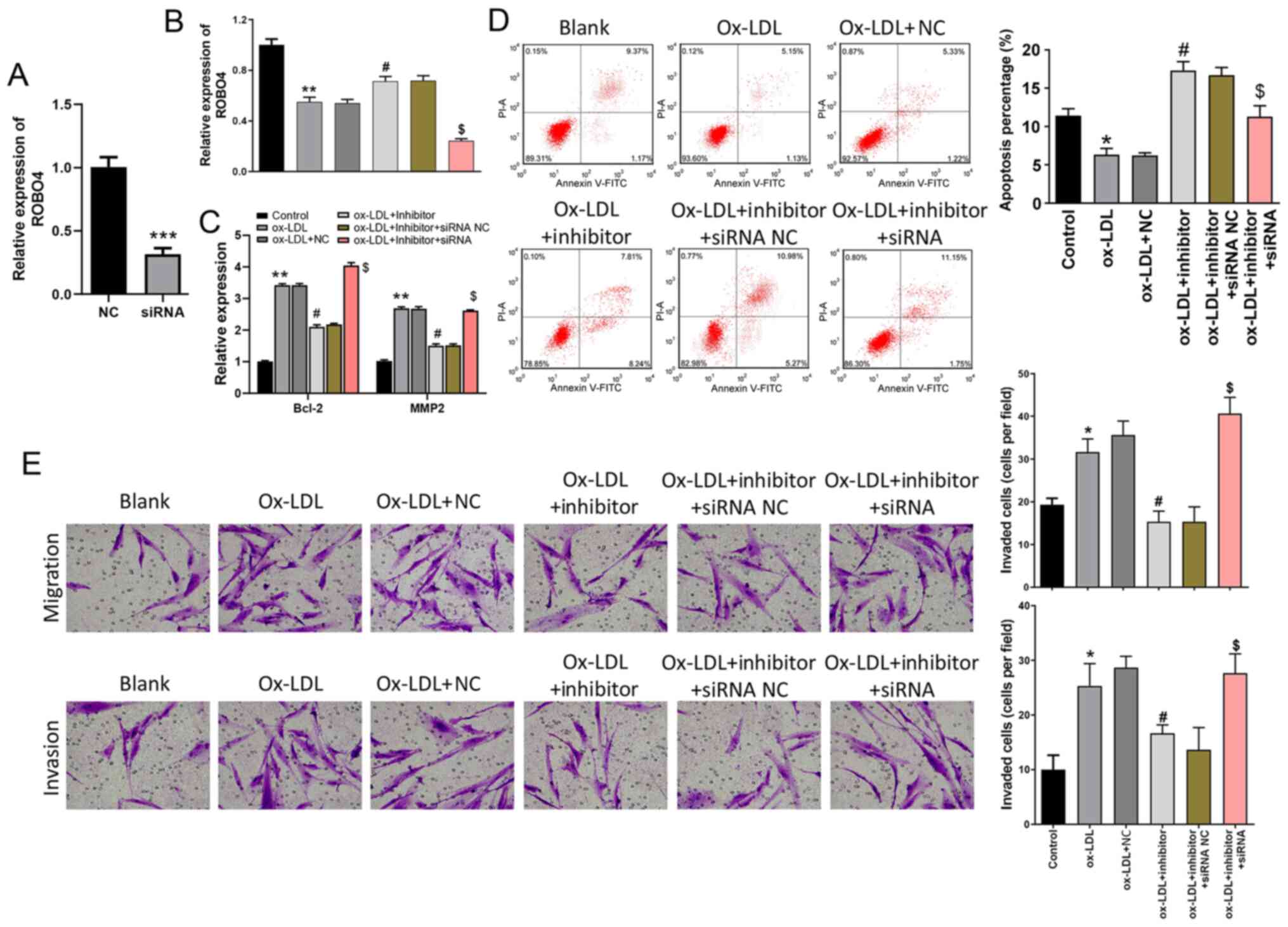 | Figure 5.miR-140-5p mediates VSMC apoptosis,
migration and invasion by targeting ROBO4. (A) Transfection
efficiency of ROBO4 siRNA. (B) Effects of ox-LDL, miR-140-5p
inhibitor and ROBO4 siRNA on ROBO4 expression levels in VSMCs. (C)
Bcl-2 and MMP2 expression levels were measured by reverse
transcription-quantitative PCR. Effect of ox-LDL, miR-140-5p
inhibitor and ROBO4 siRNA on VSMC (D) apoptosis, (E) migration and
invasion (magnification, ×200). *P<0.05, **P<0.01,
***P<0.001 vs. NC or control; #P<0.05 vs. ox-LDL +
NC; $P<0.05 vs. ox-LDL + inhibitor + siRNA NC. miR,
microRNA; VSMC, vascular smooth muscle cell; ROBO4, roundabout
guidance receptor 4; siRNA, small interfering RNA; ox-LDL,
oxidized-low density lipoprotein; MMP2, matrix metallopeptidase 2;
NC, negative control. |
Discussion
Increasing evidence has revealed that non-coding RNA
molecules, including miRNAs, are closely associated with various
vascular disorders, such as AS, and their related complications
(19,20,33).
Despite previous reports describing the pathogenic roles served by
miR-140-5p in angiogenesis and glioma cell proliferation/invasion
(21,22), the roles of miR-R140-5p in AS
development and progression are not completely understood. The
present study indicated that miR-140-5p expression levels were
significantly increased in artery plaque tissues collected from
patients with AS and in ox-LDL-treated human VSMCs compared with
healthy control tissues and control VSMCs, respectively. Moreover,
ROBO4 gene expression was markedly decreased in ox-LDL-treated
VSMCs compared with control VSMCs. The transfection experiments
indicated that miR-140-5p promoted cell viability, migration, and
invasion, but decreased apoptosis in ox-LDL-treated human VSMCs,
which also inhibited ROBO4 gene expression. Additionally, the
direct binding of miR-140-5p to the 3′-UTR of ROBO mRNA was
validated using a dual luciferase reporter assay. Finally, the
results indicated that the regulatory effects of ox-LDL treatment
on human VSMCs were markedly abrogated by ROBO4 overexpression.
Collectively, the results indicated that miR-140-5p served as an
AS-promoting factor in human VSMCs by directly targeting and
repressing ROBO4 gene expression.
The functions and molecular mechanisms underlying
miR-140-5p have been extensively investigated due to its widespread
and essential roles in various biological processes and pathogenic
conditions. For instance, a recent study demonstrated that
miR-140-5p could serve as a potent tumor suppressor and inhibit
pediatric renal cancer cell proliferation by targeting and
suppressing the expression of TGF-β receptor type 1 and
insulin-like growth factor 1 receptor (34). Besides its widespread involvement in
the development and progression of various human malignancies,
miR-140-5p has been reported to serve as a key non-coding RNA
molecule during the development of other human disorders, such as
skeletal dysplasia, pulmonary arterial hypertension,
inflammation-induced acute lung injuries and angiogenesis (35–37).
On the other hand, several miRNAs, including miR-155, miR-33,
miR-296 and miR-302a, have also been identified as essential
regulators of AS development (38–41).
Considering the functions of the microRNAs during AS pathogenesis,
it was hypothesized that miR-140-5p may be involved in AS. The high
levels of miR-140-5p expression detected in artery tissues from
patients with AS and in ox-LDL-treated VSMCs, as well as the
cellular functions of miR-140-5p in ox-LDL-treated VSMCs, were
further indicated in the present study by transfecting human VSMCs
with miR-140-5p inhibitor. The results provided a novel perspective
for the molecular processes underlying AS development.
Previous studies indicated that human VSMCs undergo
significant alterations in their cellular processes during AS
pathogenesis, as demonstrated by increased rates of proliferation,
migration and invasion, and a lower rate of apoptosis (6–8). For
instance, VSMC proliferation was demonstrated to be severely
limited by CD98, a deficiency of which greatly altered the number
of smooth muscle cells in the plaque tissues of an AS mouse model
(42). Wang et al (43) suggested that ox-LDL exerts biphasic
effects on VSMC proliferation and apoptosis via an
ox-LDL/β2-glycoprotein (GP)I/anti-β2GP complex. In the present
study, ox-LDL treatment significantly promoted human VSMC
viability, migration and invasion, but suppressed apoptosis, which
was consistent with previously reported effects (6). The discrepancy between studies may be
due to the sensitivity of VSMCs and the length of exposure to
ox-LDLs. For instance, the NF-κB and MAPK signaling pathways send a
variety of signals that regulate cell apoptosis and proliferation,
and the net balance between cell death and survival is determined
by the signaling pathway involved (43). Moreover, the present study indicated
that miR-140-5p inhibitor transfection could almost completely
reverse ox-LDL-induced alterations in VSMC viability, migration,
invasion and apoptosis. However, inhibition of AS development via
miR-140-5p inhibition requires further investigation in animal AS
models. The cellular assays conducted in the present study
characterized miR-140-5p as a novel AS-promoting miRNA that could
possibly serve as biomarker for diagnosing AS, and be useful for
the development of novel drugs for the treatment of AS.
ROBO4 is a major homolog of the neuronal Robo
family, and was previously identified as a critical
vascular-specific receptor that was specifically expressed in the
vascular endothelium and inhibited endothelial migration (26). Subsequent investigations indicated
that ROBO4 could interact with and activate the vascular Netrin
receptor Unc-5 homolog B to inhibit angiogenesis and maintain
vessel integrity by inhibiting the VEGF signaling pathway (25). A previous study also demonstrated
the essential roles served by the ROBO4 gene in regulating vascular
permeability and neovascularization (27). However, the pathogenic function of
the ROBO4 gene during artery stiffening and AS
development/progression is not completely understood. In the
present study, ROBO4 gene expression was downregulated in the
artery plaque tissues from patients with AS and in ox-LDL-treated
human VSMCs compared with healthy artery tissues and control VSMCs,
respectively. Consistent with a bioinformatics prediction, the
dual-luciferase reporter assay indicated that miR-140-5p bound to
the 3′-UTR regions of ROBO4 mRNA. Moreover, the results indicated
that the effects of ox-LDL treatment on human VSMCs were mitigated
by ROBO4 overexpression. The results characterized the ROBO4 gene
as the target of miR-140-5p in VSMCs during AS development.
In summary, the present study indicated that
miR-140-5p was highly expressed in human VSMCs during AS. The
results suggested that miR-140-5p might promote the AS process by
directly binding to ROBO4 mRNA and suppressing ROBO4 gene
expression. ROBO4 overexpression effectively alleviated the
cellular phenotypic alterations in VSMCs associated with AS
pathogenesis. The present study provided novel insights into the
pathogenic mechanisms underlying artery stiffening and AS
development, and also established a rationale for further
investigation into the use of miR-140-5p and ROBO4 as target
molecules for use in AS diagnosis and treatment.
Acknowledgements
Not applicable.
Funding
The present study was supported by the North Sichuan
Medical College City School Cooperation Program (grant nos.
18sxhz0364 and 18sxhz0408).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YL and YZ designed the experiments. YL, YML and HP
performed the experiments and collected the data. YL and YZ confirm
the authenticity of all the raw data. HP analyzed the experimental
data. YML described the results. YL drafted the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was reviewed and approved by the
Medical Ethics Committee of The First Affiliated Hospital of
Chongqing Medical University. Written informed consent was obtained
from each participant.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Schwertani A, Choi HY and Genest J: HDLs
and the pathogenesis of atherosclerosis. Curr Opin Cardiol.
33:311–316. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Weber C and Noels H: Atherosclerosis:
Current pathogenesis and therapeutic options. Nat Med.
17:1410–1422. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Spence JD: Recent advances in
pathogenesis, assessment, and treatment of atherosclerosis. F1000
Res. 5:18802016. View Article : Google Scholar
|
|
4
|
Nicorescu I, Dallinga GM, de Winther MPJ,
Stroes ESG and Bahjat M: Potential epigenetic therapeutics for
atherosclerosis treatment. Atherosclerosis. 281:189–197. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Musunuru K and Kathiresan S: Surprises
from genetic analyses of lipid risk factors for atherosclerosis.
Circ Res. 118:579–585. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bennett MR, Sinha S and Owens GK: Vascular
smooth muscle cells in atherosclerosis. Circ Res. 118:692–702.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hu D, Yin C, Luo S, Habenicht AJR and
Mohanta SK: Vascular smooth muscle cells contribute to
atherosclerosis immunity. Front Immunol. 10:11012019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lacolley P, Regnault V, Segers P and
Laurent S: Vascular smooth muscle cells and arterial stiffening:
Relevance in development, aging, and disease. Physiol Rev.
97:1555–1617. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Alexander MR and Owens GK: Epigenetic
control of smooth muscle cell differentiation and phenotypic
switching in vascular development and disease. Annu Rev Physiol.
74:13–40. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ping S, Li Y, Liu S, Zhang Z, Wang J, Zhou
Y, Liu K, Huang J, Chen D, Wang J, et al: Simultaneous increases in
proliferation and apoptosis of vascular smooth muscle cells
accelerate diabetic mouse venous atherosclerosis. PLoS One.
10:e01413752015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hou J, Xue X and Li J: Vasostatin-2
inhibits cell proliferation and adhesion in vascular smooth muscle
cells, which are associated with the progression of
atherosclerosis. Biochem Biophys Res Commun. 469:948–953. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Eulalio A, Huntzinger E and Izaurralde E:
Getting to the root of miRNA-mediated gene silencing. Cell.
132:9–14. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Farazi TA, Spitzer JI, Morozov P and
Tuschl T: miRNAs in human cancer. J Pathol. 223:102–115. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu J, Zhu W and Wu W: MicroRNAs change
the landscape of cancer resistance. Methods Mol Biol. 1699:83–89.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kato M, Wang M, Chen Z, Bhatt K, Oh HJ,
Lanting L, Deshpande S, Jia Y, Lai JY, O'Connor CL, et al: An
endoplasmic reticulum stress-regulated lncRNA hosting a microRNA
megacluster induces early features of diabetic nephropathy. Nat
Commun. 7:128642016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ha M and Kim VN: Regulation of microRNA
biogenesis. Nat Rev Mol Cell Biol. 15:509–524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Salta E and De Strooper B: microRNA-132: A
key noncoding RNA operating in the cellular phase of Alzheimer's
disease. FASEB J. 31:424–433. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang Y, Liu Z, Zhou M and Liu C:
MicroRNA-129-5p inhibits vascular smooth muscle cell proliferation
by targeting Wnt5a. Exp Ther Med. 12:2651–2656. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Feinberg MW and Moore KJ: MicroRNA
Regulation of Atherosclerosis. Circ Res. 118:703–720. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu X and Li Z: MicroRNAs regulate vascular
smooth muscle cell functions in atherosclerosis (Review). Int J Mol
Med. 34:923–933. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun J, Tao S, Liu L, Guo D, Xia Z and
Huang M: miR-140-5p regulates angiogenesis following ischemic
stroke by targeting VEGFA. Mol Med Rep. 13:4499–4505. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hu Y, Li Y, Wu C, Zhou L, Han X, Wang Q,
Xie X, Zhou Y and Du Z: MicroRNA-140-5p inhibits cell proliferation
and invasion by regulating VEGFA/MMP2 signaling in glioma. Tumour
Biol. 39:10104283176975582017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu Q-Q, Ren K, Liu S-H, Li W-M, Huang C-J
and Yang X-H: MicroRNA-140-5p aggravates hypertension and oxidative
stress of atherosclerosis via targeting Nrf2 and Sirt2. Int J Mol
Med. 43:839–849. 2019.PubMed/NCBI
|
|
24
|
Rothman AMK, Arnold ND, Pickworth JA,
Iremonger J, Ciuclan L, Allen RMH, Guth-Gundel S, Southwood M,
Morrell NW, Thomas M, et al: MicroRNA-140-5p and SMURF1 regulate
pulmonary arterial hypertension. J Clin Invest. 126:2495–2508.
2016. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Koch AW, Mathivet T, Larrivée B, Tong RK,
Kowalski J, Pibouin-Fragner L, Bouvrée K, Stawicki S, Nicholes K,
Rathore N, et al: Robo4 maintains vessel integrity and inhibits
angiogenesis by interacting with UNC5B. Dev Cell. 20:33–46. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Park KW, Morrison CM, Sorensen LK, Jones
CA, Rao Y, Chien CB, Wu JY, Urness LD and Li DY: Robo4 is a
vascular-specific receptor that inhibits endothelial migration. Dev
Biol. 261:251–267. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang F, Prahst C, Mathivet T,
Pibouin-Fragner L, Zhang J, Genet G, Tong R, Dubrac A and Eichmann
A: The Robo4 cytoplasmic domain is dispensable for vascular
permeability and neovascularization. Nat Commun. 7:135172016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Okada Y, Funahashi N, Tanaka T, Nishiyama
Y, Yuan L, Shirakura K, Turjman AS, Kano Y, Naruse H, Suzuki A, et
al: Endothelial cell-specific expression of roundabout 4 is
regulated by differential DNA methylation of the proximal promoter.
Arterioscler Thromb Vasc Biol. 34:1531–1538. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tian R, Liu Z, Zhang H, Fang X, Wang C, Qi
S, Cheng Y and Su G: Investigation of the regulation of roundabout4
by hypoxia-inducible factor-1α in microvascular endothelial cells.
Invest Ophthalmol Vis Sci. 56:2586–2594. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dai C, Gong Q, Cheng Y and Su G:
Regulatory mechanisms of Robo4 and their effects on angiogenesis.
Biosci Rep. 39:BSR201905132019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
eLife. 4:e050052015. View Article : Google Scholar
|
|
33
|
Gao S, Wassler M, Zhang L, Li Y, Wang J,
Zhang Y, Shelat H, Williams J and Geng YJ: MicroRNA-133a regulates
insulin-like growth factor-1 receptor expression and vascular
smooth muscle cell proliferation in murine atherosclerosis.
Atherosclerosis. 232:171–179. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu Z, He F, OuYang S, Li Y, Ma F, Chang
H, Cao D and Wu J: miR-140-5p could suppress tumor proliferation
and progression by targeting TGFBRI/SMAD2/3 and IGF-1R/AKT
signaling pathways in Wilms' tumor. BMC Cancer. 19:4052019.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Grigelioniene G, Suzuki HI, Taylan F,
Mirzamohammadi F, Borochowitz ZU, Ayturk UM, Tzur S, Horemuzova E,
Lindstrand A, Weis MA, et al: Gain-of-function mutation of
microRNA-140 in human skeletal dysplasia. Nat Med. 25:583–590.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang Y, Liu D, Xi Y and Li J, Liu B and Li
J: Upregulation of miRNA-140-5p inhibits inflammatory cytokines in
acute lung injury through the MyD88/NF-κB signaling pathway by
targeting TLR4. Exp Ther Med. 16:3913–3920. 2018.PubMed/NCBI
|
|
37
|
Zhu TT, Zhang WF, Yin YL, Liu YH, Song P,
Xu J, Zhang MX and Li P: MicroRNA-140-5p targeting tumor necrosis
factor-α prevents pulmonary arterial hypertension. J Cell Physiol.
234:9535–9550. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li H, Ouyang XP, Jiang T, Zheng XL, He PP
and Zhao GJ: MicroRNA-296: A promising target in the pathogenesis
of atherosclerosis? Mol Med. 24:122018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Meiler S, Baumer Y, Toulmin E, Seng K and
Boisvert WA: MicroRNA 302a is a novel modulator of cholesterol
homeostasis and atherosclerosis. Arterioscler Thromb Vasc Biol.
35:323–331. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nazari-Jahantigh M, Wei Y, Noels H, Akhtar
S, Zhou Z, Koenen RR, Heyll K, Gremse F, Kiessling F, Grommes J, et
al: MicroRNA-155 promotes atherosclerosis by repressing Bcl6 in
macrophages. J Clin Invest. 122:4190–4202. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Rotllan N, Ramírez CM, Aryal B, Esau CC
and Fernández-Hernando C: Therapeutic silencing of microRNA-33
inhibits the progression of atherosclerosis in Ldlr−/−
mice - brief report. Arterioscler Thromb Vasc Biol. 33:1973–1977.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Baumer Y, McCurdy S, Alcala M, Mehta N,
Lee BH, Ginsberg MH and Boisvert WA: CD98 regulates vascular smooth
muscle cell proliferation in atherosclerosis. Atherosclerosis.
256:105–114. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang T, Zhou H, Chen Y, Zhang P and Wang
T: The biphasic effects of the oxLDL/β2GPI/anti-β2GPI complex on
VSMC proliferation and apoptosis. Cell Signal. 57:29–44. 2019.
View Article : Google Scholar : PubMed/NCBI
|