Introduction
Clinically, heterotopic ossification (HO) is a
pathologic process defined as the formation of ectopic bone in
muscle and soft tissues. HO has long been described as a congenital
or inherited malformation, or a complication of various conditions,
such as paraplegia, traumatic injury, post hip arthroplasty, as
well as ossification of the posterior longitudinal ligament (OPLL)
(1). The histopathology of HO is
well investigated; however, the molecular mechanism that regulates
the pathogenesis of extraskeletal ossification remains largely
unknown. Recent data have suggested that the bone morphogenetic
protein (BMP) family may play a central role in HO development. For
example, the expression of BMP2, BMP4, BMP7 and BMP9 was
substantially elevated in spinal cord injury-induced HO (2). HO formation also frequently occurred
in patients with osteonecrosis of femoral head (ONFH) receiving
intraoperative recombinant human BMP2 protein (3). There is currently a lack of effective
drug therapy against HO. It is imperative to explore the signaling
pathway underlying the development of ectopic bone formation to
develop novel molecular therapy for this disease.
As a tumor suppressor, the phosphatase and tensin
homolog deleted from chromosome 10 (PTEN) functions primarily via
dephosphorylation of phosphatidylinositol (3,4,5)-triphosphate
(PIP3) to phosphatidylinositol (4,5)-bisphosphate (PIP2), leading
to the negative regulation of the phosphatidylinositol 3-kinase
(PI3K)/AKT activity (4). The
PTEN/PI3K/AKT nexus has been implicated in diverse physiological
and pathological conditions and plays a crucial role in the
regulation of cell growth, apoptosis, metabolism and other
processes (5). This signaling axis
is also involved in embryonic bone development and fracture
healing. Loss of PTEN in mature osteoblasts leads to profoundly
accumulated postnatal bone mass at all skeletal sites and notably
decreased apoptosis (6). The
knockout of PTEN in osteoblasts also enhances intramembranous and
late endochondral fracture repair (7). Recently, micro (mi)RNA-26a-5p has been
shown to inhibit PTEN expression and promote the bone-healing
process in fracture patients with traumatic brain injury (8). Being the direct target of PTEN,
PI3K/AKT is also involved in osteoblast function and bone
development. The whole-body deletion of AKT1 causes decreased bone
mass, which associates with enhanced apoptosis and impaired
osteoblast differentiation (9).
PI3K is required for BMP2 to induce osteoblast differentiation
(10). Interestingly, an inverse
association between BMP2 and PTEN has been reported, in which BMP2
could downregulate PTEN expression at the protein level, an effect
mediated via the RAS/ERK signaling pathway (11).
Although the PTEN/PI3K/AKT pathway is critically
involved in skeletogenesis and fracture healing, it remains unknown
whether this pathway impacts HO. In the present study, the mouse
mesenchymal stem cell line C3H10T1/2 were cultured and treated with
recombinant human BMP2 (rhBMP2) to induce osteoblast
differentiation. An ectopic ossification model was also established
by implantation of rhBMP2 into the muscle of mice. It was
hypothesized that PTEN inhibits BMP2-induced osteoblast
differentiation, whereas BMP2 activates PI3K/AKT via its inhibition
of PTEN. It was also hypothesized that PI3K/AKT signaling activity
is indispensable for BMP2 to induce osteogenic differentiation
in vitro and ectopic bone formation in vivo.
Materials and methods
Cell culture and stable
transfection
Mouse fibroblast-like pluripotent stem cell line
C3H10T1/2 (clone 8; American Type Culture Collection) was used in
the present study; this cell line undergoes osteoblast lineage
commitment, early and late osteoblast differentiation, and
osteoblast mineralization in response to osteogenic agents (such as
BMP2), thus providing an ideal model for osteoblast-associated
studies in the literature (12–15).
C3H10T1/2 cells were cultured at 37°C in humidified
air with 5% CO2 in a growth medium containing Dulbecco's
μodified Eagle's μedium (DMEM; Invitrogen; Thermo Fisher
Scientific, Inc.) with 10% fetal calf serum (FCS; Invitrogen;
Thermo Fisher Scientific, Inc.). Cells were authenticated using
small tandem repeat analysis and routinely tested to assure no
mycoplasma contamination. For stable transfection, the human
PTEN gene was subcloned into the pcDNA3.1 plasmid vector
(Invitrogen; Thermo Fisher Scientific, Inc.). The pcDNA-PTEN was
transfected into C3H10T1/2 cells using Lipofectamine®
2000 transfection reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) following the manufacturer's protocol. After transfection,
cells were treated with G418 at a dosage of 800 µg/ml at 37°C for
10 days to select for positive clones. Individual colonies were
pooled together and transferred to a new culture dish. After
selection, stable cell lines overexpressing PTEN were cultured in
DMEM-10% FCS at 37°C. Cells transfected with pcDNA3.1 alone were
used as controls.
Proliferation and apoptosis assay
The proliferation of C3H10T1/2 cells was evaluated
using an MTT Proliferation kit (Roche Applied Science). Cells were
plated in 96-well plates at a density of 5×103
cells/well in 100 µl medium. After 24 h, cells were incubated with
10 µl MTT reagent for 2 h. Then, 100 µl of the solubilization
solution from the kit was added to dissolve the formazan product.
Absorbances at 570 nm were measured.
For the apoptosis assay, C3H10T1/2 cells were
cultured under serum-starved conditions (DMEM-0.5% FCS) for 24 h.
The proapoptotic activity was determined by assessing the caspase-3
activity using the Caspase-3 Colorimetric Assay kit (R&D
Systems, Inc.) according to the manufacturer's protocol.
Absorbances at 405 nm were read on a microplate reader.
Reverse transcription-quantitative PCR
(RT-qPCR)
For RT-qPCR analysis, total RNA was isolated from
cells using the TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). RNA (2 µg) was reverse transcribed to cDNA at
50°C for 50 min using SuperScript III Reverse Transcriptase (Thermo
Fisher Scientific, Inc.). PCR reactions were performed with SYBR
green master mix (Thermo Fisher Scientific, Inc.). The cycling
conditions were: 50°C for 2 min; 95°C for 10 min; then 95°C for 15
sec and 60°C for 1 min for 40 cycles. The PCR primers for the mouse
genes are listed in Table I. PCR
products were analyzed with the ABI PRISM 7900HT Sequence
Detections System (Applied Biosystems; Thermo Fisher Scientific,
Inc.). To ensure primer specificity, one dissociation stage was
added to produce the melting curve at the end of the cycling
condition. The relative mRNA concentrations of the target genes
were determined with ABI software (RQ Manager; Version 1.2), which
normalizes the target gene threshold cycle to that of endogenous
B2M transcripts (ΔΔCt), using the formula 2−ΔΔCq to
define fold change (16).
 | Table I.PCR primers. |
Table I.
PCR primers.
| Gene | Forward, 5′-3′ | Reverse, 5′-3′ |
|---|
| Alpl |
GTTGCCAAGCTGGGAAGAACAC |
CCCACCCCGCTATTCCAAAC |
| Col1α1 |
CCTGCCTGCTTCGTGTAA |
GGTCACGTTCAGTTGGTCA |
|
Osteocalcin |
GGGAGACAACAGGGAGGAAAC |
CAGGCTTCCTGCCAGTACCT |
| Runx2 |
CGGTCTCCTTCCAGGATGGT |
GCTTCCGTCAGCGTCAACA |
| Sp7 |
AGCGACCACTTGAGCAAACAT |
GCGGCTGATTGGCTTCTTCT |
| Pten |
AATTCCCAGTCAGAGGCGCTATGT |
GATTGCAAGTTCCGCCACTGAACA |
| B2M |
AAATGCTGAAGAACGGGAAA |
GATGCTTGATCACATGTCTCG |
Western blotting
Cell lysates were prepared using RIPA Lysis buffer
(Sigma-Aldrich; Merck KGaA). The samples (25 µg) were
electrophoresed on a 10% SDS polyacrylamide gel (Invitrogen; Thermo
Fisher Scientific, Inc.) and were transferred onto a polyvinylidene
difluoride membrane (EMD Millipore). Membranes were washed with
TBS/0.1% Tween-20 (TBST) and blocked with 1% BSA in TBST for 1 h at
room temperature. Primary antibodies (Table II) were added to the membranes
overnight at 4°C. Blots were washed three times with TBST, followed
by incubation with horseradish peroxidase-conjugated secondary
antibodies (Bio-Rad Laboratories, Inc.) for 1 h at room
temperature. After washing with TBST, the membrane was detected
with ECL Detection reagents (Thermo Fisher Scientific, Inc.) and
exposed to X-ray film. Densitometry of the protein bands was
analyzed using ImageJ software (v1.51j8; National Institutes of
Health).
| Table II.Antibodies used. |
Table II.
Antibodies used.
| Antibody | Dilution | Manufacturer | Cat. no. |
|---|
| PTEN | 1:500 | Santa Cruz
Biotechnology, Inc. | SC-7974 |
| Phospho-AKT | 1:1,000 | Cell Signaling
Technology, Inc. | 9271 |
| AKT | 1:1,000 | Cell Signaling
Technology, Inc. | 9272 |
| β-actin | 1:1,000 | Cell Signaling
Technology, Inc. | 4967 |
| Anti-mouse IgG | 1:3,000 | Cell Signaling
Technology, Inc. | 7076 |
| Anti-rabbit
IgG | 1:3,000 | Cell Signaling
Technology, Inc. | 7074 |
Osteogenic differentiation
Confluent C3H10T1/2 cells were incubated in
osteogenic medium containing DMEM-10% FCS, 50 µg/ml ascorbic acid,
10 mM sodium β-glycerophosphate, and 200 ng/ml recombinant human
BMP2 (rhBMP2; Sigma-Aldrich; Merck KGaA). The osteogenic medium was
replaced every 48 h. The activity of alkaline phosphatase (ALP) was
examined on day 3 using a colorimetric assay kit (Abcam). This kit
used p-nitrophenyl phosphate as a phosphatase substrate that turned
yellow at OD 405 when dephosphorylated by ALP. For ALP staining,
cells were washed with PBS, fixed with 70% ethanol for 10 min at
room temperature, and incubated with NBT/BCIP solution for 20 min
at room temperature. The positively stained cells were monitored
using a light microscope (×200; Olympus). The secretion of
osteocalcin was examined using a mouse ELISA kit (cat. no.
MBS2020904; MyBioSource, Inc.) after cells were cultured for 7
days. Cells were stained using the von Kossa method for calcium
deposition at room temperature after 2 weeks. Briefly, cells were
fixed with 4% paraformaldehyde for 20 min and washed with water
twice. Cells were then treated with 1% silver nitrate solution
under ultraviolet light for 30 min. After washing with distilled
water, cells were incubated with 5% sodium thiosulfate for 5 min
and were counterstained with 1% nuclear fast red solution for 5
min. Cells were also stained with Alizarin Red for mineralization
at room temperature after being cultured in the osteogenic medium
after 4 weeks. In brief, cells were fixed with 4% paraformaldehyde
for 20 min and washed with water. Cells were then treated with 2%
Alizarin Red S solution for 5 min. The quantification of stains was
performed using the ARed-Q assay (cat. no. 8678, ScienCell).
Following von Kossa and Alizarin Red S staining, cells were
monitored using a light microscope (magnification, ×200; Olympus
Corporation). The expression of Runx2 and osterix (the two key
transcription factors for osteoblasts), ALP (early osteoblast
differentiation marker), collagen type I (COL1A1), and osteocalcin
(late osteoblast differentiation marker) was examined using qPCR
analysis.
Ectopic bone-formation model
A total of 18 male BALB/c mice (12–14 weeks old)
were utilized for the present study. All animal procedures were
reviewed and approved by the Institutional Animal Care and Use
Committee of the Shandong Provincial Hospital (approval no.
M2018143). Mice were anesthetized using an intraperitoneal
injection of a cocktail of ketamine (80 mg/kg)/xylazine (10 mg/kg).
A 5-mm skin incision was made on the dorsal surface of the right
lower limb; then a muscle pouch was created in the gluteus of mice.
Freeze-dried absorbable collagen sponges (5×5×5 mm, Helistat) were
mixed with 20 µg rhBMP2 and placed into the muscle pouch. The
incision was closed with sutures. Animals were subcutaneously
injected with an analgesic agent, buprenorphine, at a dosage of 0.1
mg/kg twice a day for 3 days. Following the operation, animals were
divided into two groups (9 mice/each). One group of mice were
injected intraperitoneally with PI3K inhibitor LY294002
(Calbiochem) at a dosage of 25 mg/kg immediately and twice weekly.
Other animals were injected with the same volume of DMSO (vehicle)
as controls. For radiography, animals were examined for
ossification using a Faxitron MX20 X-ray system (Faxitron Bioptics)
at 3 weeks following the operation. Animals were monitored daily
for their health and behaviors. All mice were in good health and
survived well after implantation of rhBMP2.
For histological analysis, mice were sacrificed
using standard CO2 euthanasia at 9 days and 3 weeks,
respectively, after the operation. A fill rate of 30% of the
chamber volume/min of CO2 to the existing air in the
chamber was used for the animals to achieve fast unconsciousness
with minimal distress. The death of mice was confirmed by
ascertaining cardiac and respiratory arrest.
Tissue samples were harvested, fixed in 4%
paraformaldehyde at 4°C for 2 days, decalcified in 20% EDTA (pH
7.4) at 4°C for 10 days, and embedded in paraffin. Sections (7 µm)
were stained with Safranin O (SO) and hematoxylin and eosin
(H&E). For SO staining, deparaffinized and rehydrated sections
were successively incubated with 0.05% fast green solution for 5
min, 1% acetic acid solution for 30 sec, and 0.1% SO solution for 5
min at room temperature. For HE staining, sections were incubated
with hematoxylin solution for 3 min and eosin solution for 1 min at
room temperature. Bone histomorphometric analysis was performed as
previously reported (17). An
analysis image window measuring 2 mm2 was established
for evaluation of bone volume as a percentage of total tissue
volume (BV/TV %), trabecular thickness (µm), trabecular number (per
mm), and trabecular separation (µm). The measurements were
performed using BoneJ, a plugin for bone image analysis in ImageJ
software (v1.51j8, National Institute of Health).
Statistical analyses
GraphPad Prism software (GraphPad Prism Software,
Inc.; version 5.0) was used for statistical analyses. Data are
shown as mean ± SD. The significance of differences was evaluated
using the Student's t-test between two groups, or one-way ANOVA
among multiple groups, followed by Tukey's post hoc test. P<0.05
was considered to indicate a statistically significant
difference.
Results
PTEN inhibits proliferation but
promotes apoptosis of C3H10T1/2 cells
To investigate the role of PTEN in BMP2-induced
osteogenic differentiation, a gain-of-function approach was used to
overexpress PTEN in C3H10T1/2 cells. Using qPCR analysis, it was
demonstrated that transfection with pcDNA-PTEN resulted in a
3.82-fold increase in PTEN mRNA level compared with that of
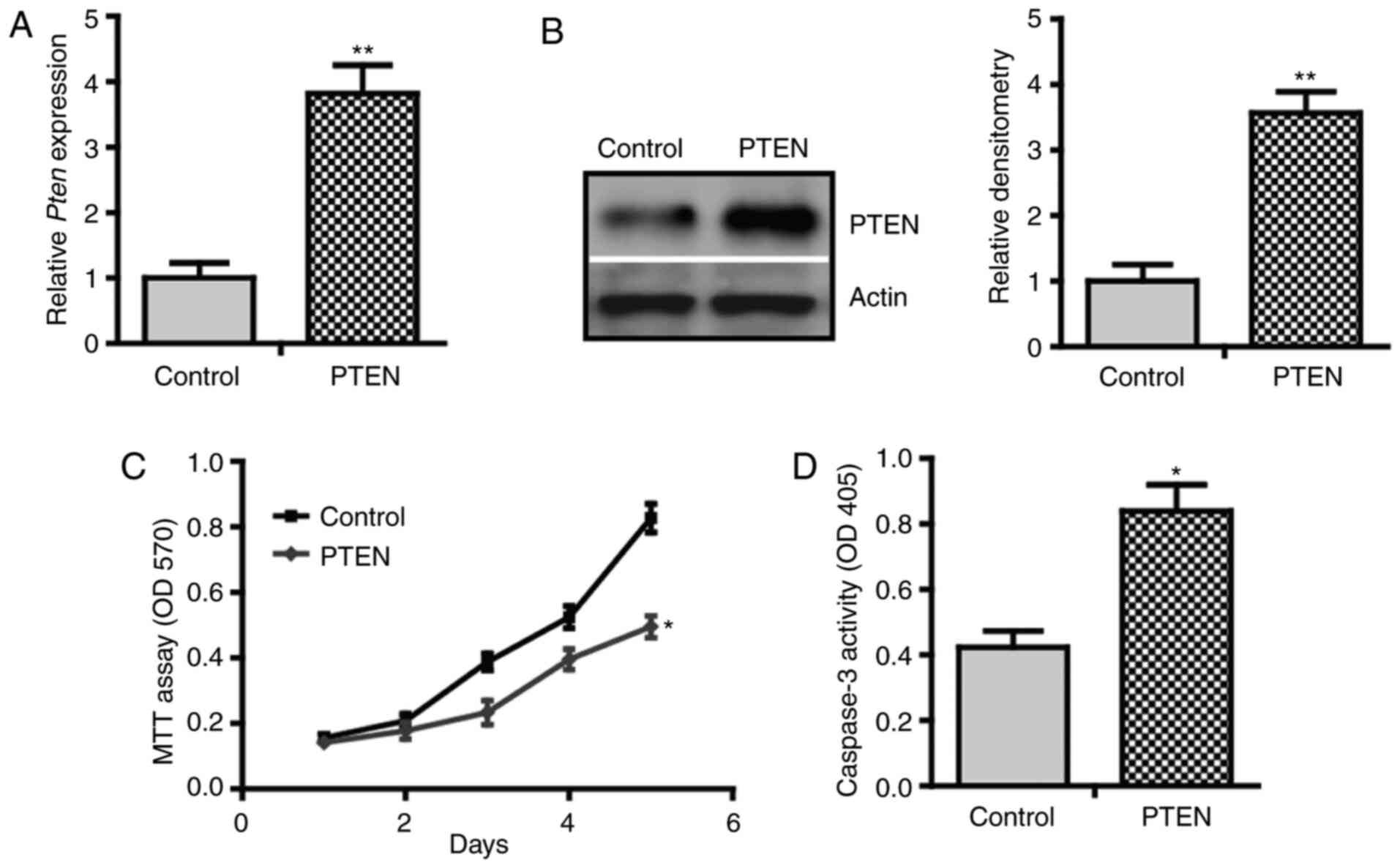
cells transfected with pcDNA3.1 empty vectors (Fig. 1A). Immunoblotting of cell lysate
confirmed the induction of PTEN at the protein level (Fig. 1B). Using MTT assay, it was found
that PTEN inhibited the proliferation of C3H10T1/2 cells (Fig. 1C). Notably, a higher caspase-3
activity in PTEN-overexpressing cells was observed compared with
the control cells (Fig. 1D). These
results suggest that PTEN inhibits proliferation but promotes
apoptosis of C3H10T1/2 cells, indicating a growth-inhibitory
effect.
PTEN inhibits BMP2-induced osteogenic
differentiation
Next, the effect of PTEN on BMP2-induced osteogenic
differentiation was investigated. Overexpression of PTEN notably
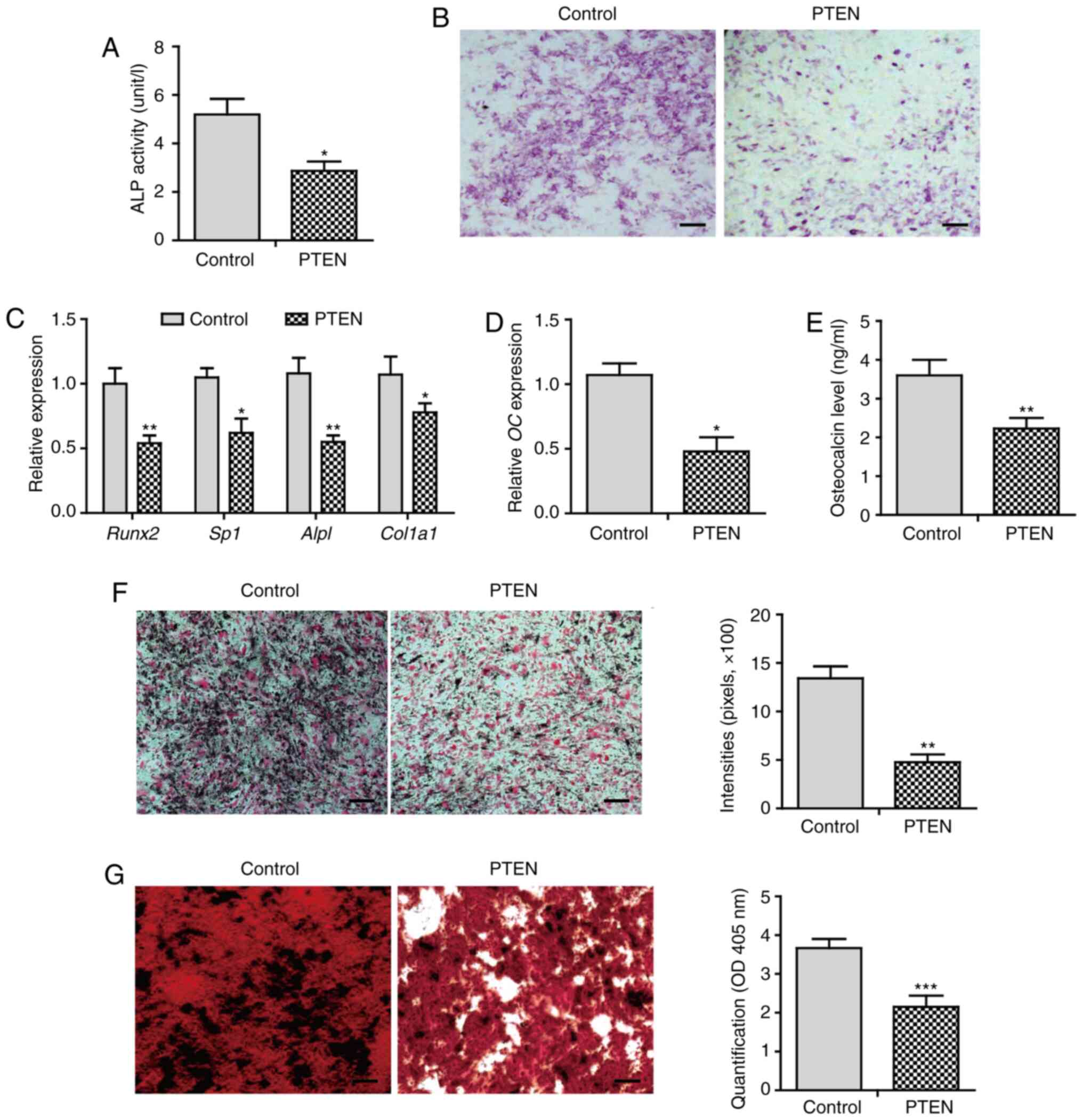
inhibited ALP activity by 45% compared with that of control cells
(Fig. 2A). ALP staining exhibited a
markedly decreased staining intensity in PTEN-overexpressing cells
(Fig. 2B). The expression of
osteoblast lineage commitment and early differentiation markers,
such as Runx2, osterix (Sp7), ALP (Alpl), and
collagen I (Col1α1), was downregulated in cells that
overexpressed PTEN (Fig. 2C). These
data suggest that PTEN inhibits early osteoblast
differentiation.
Furthermore, overexpression of PTEN significantly
(P<0.05) decreased the gene expression and secretion of
osteocalcin (Fig. 2D and E). The
calcium deposition level was decreased in cells that overexpressed
PTEN (Fig. 2F). Alizarin red
staining showed extensive mineralized nodule formation in control
cells. In contrast, the mineralization level was attenuated in
cells overexpressing PTEN (Fig.
2G). These results suggest that PTEN inhibits BMP2-induced late
osteogenic differentiation, leading to impaired osteoblast
maturation characterized by decreased calcium deposition and
mineralization.
BMP2 inhibits the expression of
PTEN
It was shown that PTEN suppresses BMP2-induced
osteogenic differentiation of C3H10T1/2 cells. These observations
imply a possible inverse association between BMP2 and PTEN. To test
this hypothesis, cells were treated with rhBMP2 at different doses
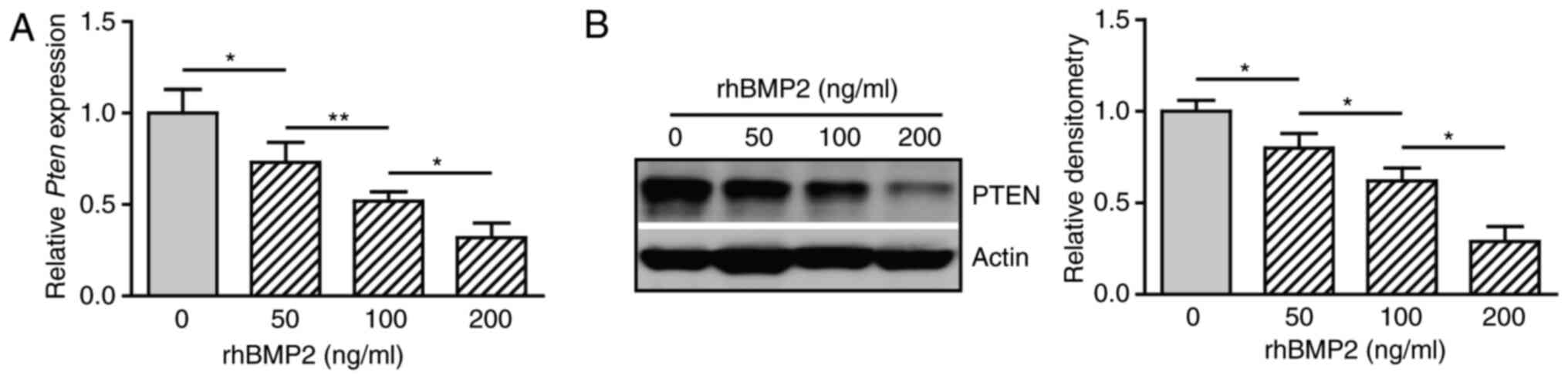
and the gene expression of PTEN was determined. BMP2 decreased the
expression of PTEN mRNA in a dose-dependent manner (Fig. 3A). Western blot analysis also
revealed an inhibitory effect of BMP2 on the expression of PTEN at
the protein level (Fig. 3B and C).
These results suggest that BMP2 suppresses the expression of PTEN
during osteogenic differentiation.
Activation of PI3K/AKT is required for
BMP2 to induce osteogenic differentiation
Next, whether BMP2-mediated inhibition of PTEN could
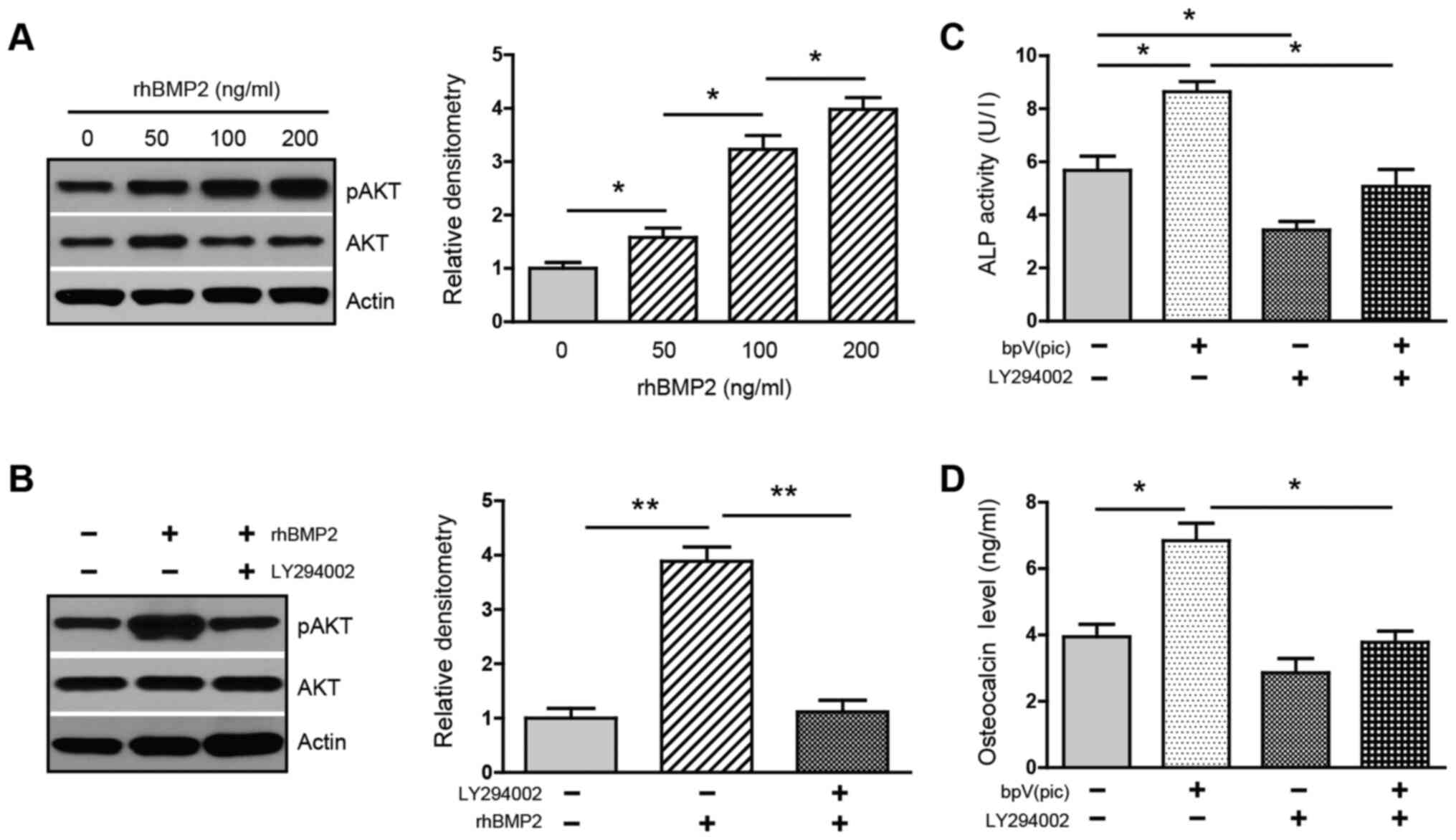
activate PI3K/AKT was investigated. Treatment with rhBMP2 at
various doses increased the phosphorylation of AKT in a
dose-dependent manner (Fig. 4A),
whereas this upregulation was greatly attenuated in the presence of
PI3K inhibitor LY294002 (Fig. 4B),
suggesting that BMP2 activates the PI3K/AKT pathway. The data
support the notion that BMP2 activates PI3K/AKT via its inhibition
of PTEN.
It was investigated whether the PI3K/AKT activity is
required for BMP2-induced osteogenic differentiation. Cells were
cultured in osteogenic medium and treated with PTEN inhibitor
bpV(pic) and PI3K inhibitor LY294002. Treatment with bpV(pic)
increased ALP activity. In contrast, the addition of LY294002
decreased both baseline and bpV(pic)-induced activity of ALP
(Fig. 4C). Similarly, the secretion
of osteocalcin was increased in the presence of bpV(pic). However,
this induction was strongly reversed by LY294002 (Fig. 4D). These results confirm that PTEN
acts as a suppressor for osteoblastogenesis and, more importantly,
support the idea that activation of the PI3K/AKT is indispensable
for BMP2 to induce osteogenic differentiation.
Inactivation of the PI3K/AKT pathway
suppresses BMP2-induced ectopic bone formation
Further, the role of PI3K/AKT during in vivo
ectopic bone formation was elucidated. Nine days after the
operation, implantation of rhBMP2 induced the formation of a large
area of a cartilaginous matrix in control mice, as indicated by the
positive red stain with Safranin O. Strikingly, animals treated
with LY294002 exhibited markedly decreased chondrogenic
differentiation at this time point, as characterized by a
substantially decreased area of the cartilaginous matrix (Fig. 5A and Di). At 3 weeks post-operation,
X-ray radiography in control mice detected the formation of ectopic
new bone. However, only a mildly ossified area was present in mice
treated with LY294002 (Fig. 5B).
Histological analysis revealed the development of mature bone
tissue, which was characterized by a well-defined perimeter of the
cortical bone rim, in which a large number of trabeculae and a
marrow cavity containing bone marrow and adipocyte-like cells were
identified. Osteoblasts along the trabeculae or osteocytes embedded
within the bone were also observed. Notably, mice treated with
LY294002 displayed highly decreased bone formation, in that only a
small number of trabeculae had developed, and the bone marrow
cavity was mainly filled with a large area of adipocyte-like cells
with only a few bone marrow cells present (Fig. 5C). The bone histomorphometric
analysis further indicated that LY294002-treated animals exhibited
a decreased ratio of bone volume/tissue volume (BV/TV), decreased
trabecular number, and decreased trabecular thickness, but
increased trabecular spacing, compared with those of control mice
(Fig. 5Dii-v). These results
suggest that treatment with LY294002 dramatically suppresses
intramuscular endochondral ossification, supporting the notion that
the activity of the PI3K/AKT pathway is critically implicated in
BMP2-induced ectopic bone formation.
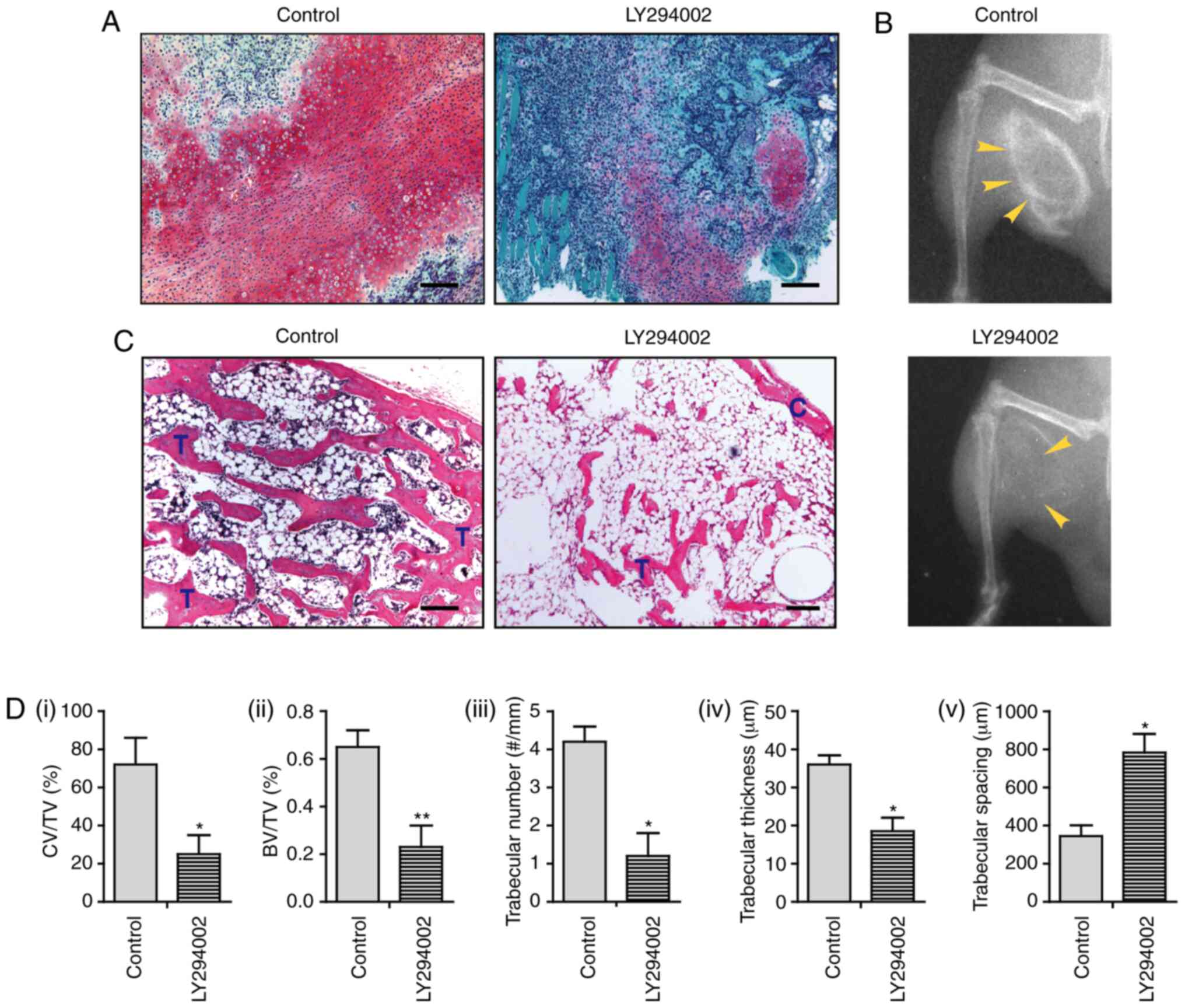 | Figure 5.Inactivation of PI3K/AKT suppresses
BMP2-induced endochondral ossification. (A) Safranin O staining at
9 days after the operation. (B) X-ray radiography for bone
formation at 3 weeks after the operation. (C) Hematoxylin and eosin
staining at 3 weeks after the operation. (D) Bone histomorphometric
analysis, including i) the ratio of CV/TV%, ii) the BV/TV%, iii)
trabecular number, iv) trabecular thickness and v) trabecular
spacing (n=3). Scale bar, 100 µm. *P<0.05; **P<0.01. BMP,
bone morphogenetic protein; C, cortical rim; T, trabecular
structure; CV/TV, cartilage volume/tissue volume; BV/TV, ratio of
bone volume/tissue volume. |
Discussion
The present study reports that PTEN inhibits
BMP2-induced osteoblast differentiation, whereas BMP2 represses
PTEN expression, leading to the activation of PI3K/AKT. In
vivo, the PI3K inhibitor suppresses BMP2-induced intramuscular
bone formation. The present results suggest an inverse association
between PTEN and BMP2, and also support the notion that PI3K/AKT
signaling activity is indispensable for BMP2 to induce ectopic
ossification. Hence, the present findings highlight a critical
implication of the PTEN/PI3K/AKT pathway during the pathogenesis of
HO.
Since BMP2 can drive the expression of Runx2 and
osterix and functions as a key regulator during osteoblast
differentiation (18,19), BMP2 was used as an osteogenic agent
to induce osteoblast differentiation in vitro and ectopic
bone formation in vivo. BMP2-induced osteoblastogenesis has
been widely reported in a variety of cell types, such as
mesenchymal stem cells, osteoblast precursors and osteoblasts
(12,20,21).
BMP2 also associates with the pathogenesis of HO (2,3).
Implantation of recombinant BMP2 has been a standard procedure to
induce endochondral bone formation in soft tissues, including
muscle, ligament and subcutaneous tissue (22–24).
The involvement of the PTEN/PI3K/AKT pathway in
skeletal development and bone repair has been previously
investigated. However, the role of this pathway nexus in HO remains
unknown. In the present study, PTEN was overexpressed in MSC-like
C3H10T1/2 cells and this tumor suppressor inhibited proliferation
but enhanced apoptosis, indicating a growth-inhibitory effect. This
result is not surprising, in that PTEN has been documented to
promote proapoptotic activity in a variety of cancer and
noncancerous cells, including MSCs and osteoblasts (6,25).
Interestingly, it was further reported that PTEN suppressed
BMP2-induced osteoblast differentiation, including the commitment
of osteoblast lineage, early and late osteoblast differentiation,
and osteoblast mineralization. The present results appear in
agreement with a previous study, in which osteoblasts deficient in
the PTEN gene displayed increased expression of several
osteoblast markers, including Runx2, ALP, collagen I and
osteocalcin (6). Based on these
findings, it seems highly plausible that PTEN functions as a
suppressor for osteoblast differentiation by inhibiting cell growth
and the entire differentiation process.
Given that PTEN is a suppressor during
BMP2-triggered osteogenic differentiation, it was deduced that an
inverse association might exist between BMP2 and PTEN. Thus far,
the association between these two molecules seems controversial.
Waite and Eng (26) reported that
exposure to BMP2 increases PTEN protein expression in MCF7 human
breast cancer cells, yet this induction is rapid and attributed to
the inhibition of PTEN degradation rather than an increase in
protein synthesis. In contrast, Beck and Carethers (11) showed that BMP2 downregulates PTEN
expression in SW480 human colon cancer cells, an effect mediated
via the RAS/ERK signaling pathway. This discrepancy may be due to
different time lengths of BMP2 treatment used in these studies or
different cell types. The BMP2-mediated rapid increase in PTEN
protein occurred within 6 h in response to BMP2, peaking at 1 h
(26), whereas BMP2 resulted in a
reduction of PTEN after 36 h of BMP2 treatment, and this inhibitory
effect could last at least 7 days (11). The present study reported that BMP2
decreased PTEN expression at mRNA and protein levels in C3H10T1/2
cells cultured in osteogenic medium for 48 h. As such, the present
results suggest a negative association between BMP2 and PTEN and
also support the notion that BMP2 exerts its osteogenic
differentiation capacity via its negative regulation of PTEN.
The present study showed that BMP2-mediated
downregulation of PTEN was closely associated with activation of
PI3K/AKT, in that BMP2 increased the level of AKT phosphorylation.
However, this upregulation could be markedly attenuated by the PI3K
inhibitor LY294002. Functionally it was further demonstrated that
the PTEN inhibitor bpV(pic) increased the activity of ALP and the
secretion of osteocalcin, the two important markers for early and
late osteoblast differentiation, respectively. Notably,
bpV(pic)-induced upregulation of ALP and osteocalcin was
substantially reversed by LY294002. Hence, the present results
indicate that the PI3K/AKT signaling activity is indispensable for
BMP2 to induce osteogenic differentiation. In support of the
present study, Ghosh-Choudhury et al (10) reported that the expression of a
dominant-negative PI3K in 2T3 osteoblast precursor cells abolishes
the induction of ALP in response to BMP2, indicating the
requirement of PI3K for BMP2-induced osteoblastogenesis. Mukherjee
and Rotwein (12) also reported
that a dominant-negative AKT is potent in preventing osteoblast
differentiation by BMP2. Therefore, results from the present study
and others suggest that activation of PI3K/AKT plays a crucial role
during BMP2-induced osteogenic differentiation.
Importantly, it was reported that the inactivation
of PI3K/AKT dramatically inhibited ectopic bone formation. In the
present study, implantation of the recombinant BMP2 into muscle
induced chondrogenic differentiation 9 days after the operation, as
evidenced by the formation of a large area of cartilaginous matrix.
However, treatment with PI3K inhibitor induced only mild cartilage
deposition. This effect suggests that the antagonization of
PI3K/AKT decreases chondrogenesis. These findings are consistent
with Iwasa et al (27), who
reported that the knockdown of PTEN in adult human chondrocytes
activates PI3K/AKT and increases the expression and synthesis of
chondrocyte markers, including collagen II, aggrecan and
proteoglycan.
Furthermore, the implantation of BMP2 induced a
well-defined mature bone structure 3 weeks after the operation. In
contrast, treatment with LY294002 greatly suppressed new bone
development, suggesting that BMP2-induced bone-forming capacity is
impaired by the inactivation of PI3K/AKT. In support of the present
results, Ford-Hutchinson et al (28) reported that a collagen II
(COL2A1) promoter-driven PTEN gene deletion could
activate the PI3K signaling and induce a dramatic increase in the
amount of trabecular and cortical bone during postnatal bone
development. Therefore, the current findings support the idea that
PI3K/AKT activity is critically required during BMP2-induced
ectopic endochondral bone formation.
The results of the present study may have
significant clinical relevance. HO is defined as the formation of
extraskeletal bone in soft tissues (such as muscle and ligament),
and it may cause severe clinical problems. For example, the
ossification of the OPLL of the cervical spine can lead to serious
myelopathy due to compression of the spinal cord and nerve roots
(29), whereas HO following
arthroplasty can become symptomatic and result in functional
disability (30). Unfortunately,
there is currently a lack of effective drug therapy against HO. The
PI3K/AKT pathway has been an attractive target for cancer therapy
(31). It is proposed that
targeting PI3K/AKT using small-molecule inhibitor(s) may also be
seriously considered for the treatment of HO.
In general, the present study reports that PTEN
inhibits proliferation but promotes apoptosis of mesenchymal
pluripotent C3H10T1/2 cells, and inhibits BMP2-induced osteogenic
differentiation. On the other hand, BMP2 represses PTEN, leading to
the activation of PI3K/AKT, which is indispensable for BMP2 to
induce osteoblastogenesis. Inactivation of the PI3K/AKT pathway
markedly suppresses BMP2-induced endochondral ossification in
muscles. The present results demonstrate for the first time that
the PTEN/PI3K/AKT signaling nexus is critically involved in the
development of ectopic bone formation. Targeting the PI3K/AKT
pathway using inhibitor(s) may represent a potential molecular
therapy against HO.
Acknowledgements
Not applicable.
Funding
This study was supported by the Shandong Provincial
Medical Technology Development Project to J.D. (grant no.
2019WS459).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JD and BT conceived the study and wrote the
manuscript. JD, XX, QZ and ZY performed experiments and contributed
data. JD, XX, QZ and BT performed data analysis. All authors
approved the final version of the manuscript.
Ethics approval and consent to
participate
All animal procedures were reviewed and approved by
the Institutional Animal Care and Use Committee of the Shandong
Provincial Hospital Affiliated to Shandong First Medical University
(approval no. M2018143).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ALP
|
alkaline phosphatase
|
|
BMP
|
bone morphogenetic protein
|
|
H&E
|
hematoxylin and eosin
|
|
MSC
|
mesenchymal stem cells
|
|
PI3K
|
phosphatidylinositol 3-kinase
|
|
PIP2
|
phosphatidylinositol (4,5)-bisphosphate
|
|
PIP3
|
phosphatidylinositol (3,4,5)-triphosphate
|
|
PTEN
|
phosphatase and tensin homolog deleted
from chromosome 10
|
|
RT-qPCR
|
reverse transcription-quantitative
PCR
|
|
SO
|
Safranin O
|
References
|
1
|
Scott MA, Levi B, Askarinam A, Nguyen A,
Rackohn T, Ting K, Soo C and James AW: Brief review of models of
ectopic bone formation. Stem Cells Dev. 21:655–667. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Łęgosz P, Drela K, Pulik Ł, Sarzyńska S
and Małdyk P: Challenges of heterotopic ossification-molecular
background and current treatment strategies. Clin Exp Pharmacol
Physiol. 45:1229–1235. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shi L, Sun W, Gao F, Cheng L and Li Z:
Heterotopic ossification related to the use of recombinant human
BMP-2 in osteonecrosis of femoral head. Medicine (Baltimore).
96:e74132017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pulido R: PTEN inhibition in human disease
therapy. Molecules. 23:2852018. View Article : Google Scholar
|
|
5
|
Papa A and Pandolfi PP: The PTEN-PI3K axis
in cancer. Biomolecules. 9:1532019. View Article : Google Scholar
|
|
6
|
Liu X, Bruxvoort KJ, Zylstra CR, Liu J,
Cichowski R, Faugere MC, Bouxsein ML, Wan C, Williams BO and
Clemens TL: Lifelong accumulation of bone in mice lacking Pten in
osteoblasts. Proc Natl Acad Sci USA. 104:2259–2264. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Burgers TA, Hoffmann MF, Collins CJ,
Zahatnansky J, Alvarado MA, Morris MR, Sietsema DL, Mason JJ, Jones
CB, Ploeg HL, et al: Mice lacking pten in osteoblasts have improved
intramembranous and late endochondral fracture healing. PLoS One.
8:e638572013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xiong Y, Cao F, Hu L, Yan C, Chen L,
Panayi AC, Sun Y, Zhou W, Zhang P, Wu Q, et al: miRNA-26a-5p
accelerates healing via downregulation of PTEN in fracture patients
with traumatic brain injury. Mol Ther Nucleic Acids. 17:223–234.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kawamura N, Kugimiya F, Oshima Y, Ohba S,
Ikeda T, Saito T, Shinoda Y, Kawasaki Y, Ogata N, Hoshi K, et al:
Akt1 in osteoblasts and osteoclasts controls bone remodeling. PLoS
One. 2:e10582007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ghosh-Choudhury N, Abboud SL, Nishimura R,
Celeste A, Mahimainathan L and Choudhury GG: Requirement of
BMP-2-induced phosphatidylinositol 3-kinase and Akt
serine/threonine kinase in osteoblast differentiation and
Smad-dependent BMP-2 gene transcription. J Biol Chem.
277:33361–33368. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Beck SE and Carethers JM: BMP suppresses
PTEN expression via RAS/ERK signaling. Cancer Biol Ther.
6:1313–1317. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mukherjee A and Rotwein P: Akt promotes
BMP2-mediated osteoblast differentiation and bone development. J
Cell Sci. 122:716–726. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mukherjee A, Wilson EM and Rotwein P:
Selective signaling by Akt2 promotes bone morphogenetic protein
2-mediated osteoblast differentiation. Mol Cell Biol. 30:1018–1027.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao L, Li G, Chan KM, Wang Y and Tang PF:
Comparison of multipotent differentiation potentials of murine
primary bone marrow stromal cells and mesenchymal stem cell line
C3H10T1/2. Calcif Tissue Int. 84:56–64. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee CH, Jin MU, Jung HM, Lee JT and Kwon
TG: Effect of dual treatment with SDF-1 and BMP-2 on ectopic and
orthotopic bone formation. PLoS One. 10:e01200512015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Egan KP, Brennan TA and Pignolo RJ: Bone
histomorphometry using free and commonly available software.
Histopathology. 61:1168–1173. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Matsubara T, Kida K, Yamaguchi A, Hata K,
Ichida F, Meguro H, Aburatani H, Nishimura R and Yoneda T: BMP2
regulates Osterix through Msx2 and Runx2 during osteoblast
differentiation. J Biol Chem. 283:29119–29125. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee MH, Kim YJ, Kim HJ, Park HD, Kang AR,
Kyung HM, Sung JH, Wozney JM, Kim HJ and Ryoo HM: BMP-2-induced
Runx2 expression is mediated by Dlx5, and TGF-beta 1 opposes the
BMP-2-induced osteoblast differentiation by suppression of Dlx5
expression. J Biol Chem. 278:34387–34394. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bain G, Müller T, Wang X and Papkoff J:
Activated beta-catenin induces osteoblast differentiation of
C3H10T1/2 cells and participates in BMP2 mediated signal
transduction. Biochem Biophys Res Commun. 301:84–91. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ogasawara T, Kawaguchi H, Jinno S, Hoshi
K, Itaka K, Takato T, Nakamura K and Okayama H: Bone morphogenetic
protein 2-induced osteoblast differentiation requires Smad-mediated
down-regulation of Cdk6. Mol Cell Biol. 24:6560–6568. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hashimoto K, Kaito T, Furuya M, Seno S,
Okuzaki D, Kikuta J, Tsukazaki H, Matsuda H, Yoshikawa H and Ishii
M: In vivo dynamic analysis of BMP-2-induced ectopic bone
formation. Sci Rep. 10:47512020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen Y, Whetstone HC, Youn A, Nadesan P,
Chow EC, Lin AC and Alman BA: Beta-catenin signaling pathway is
crucial for bone morphogenetic protein 2 to induce new bone
formation. J Biol Chem. 282:526–533. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hoshi K, Amizuka N, Sakou T, Kurokawa T
and Ozawa H: Fibroblasts of spinal ligaments pathologically
differentiate into chondrocytes induced by recombinant human bone
morphogenetic protein-2: Morphological examinations for
ossification of spinal ligaments. Bone. 21:155–162. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yamada KM and Araki M: Tumor suppressor
PTEN: Modulator of cell signaling, growth, migration and apoptosis.
J Cell Sci. 114:2375–2382. 2001.PubMed/NCBI
|
|
26
|
Waite KA and Eng C: BMP2 exposure results
in decreased PTEN protein degradation and increased PTEN levels.
Hum Mol Genet. 12:679–684. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Iwasa K, Hayashi S, Fujishiro T, Kanzaki
N, Hashimoto S, Sakata S, Chinzei N, Nishiyama T, Kuroda R and
Kurosaka M: PTEN regulates matrix synthesis in adult human
chondrocytes under oxidative stress. J Orthop Res. 32:231–237.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ford-Hutchinson AF, Ali Z, Lines SE,
Hallgrimsson B, Boyd SK and Jirik FR: Inactivation of Pten in
osteo-chondroprogenitor cells leads to epiphyseal growth plate
abnormalities and skeletal overgrowth. J Bone Miner Res.
22:1245–1259. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yan L, Gao R, Liu Y, He B, Lv S and Hao D:
The pathogenesis of ossification of the posterior longitudinal
ligament. Aging Dis. 8:570–582. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Biz C, Pavan D, Frizziero A, Baban A and
Iacobellis C: Heterotopic ossification following hip arthroplasty:
A comparative radiographic study about its development with the use
of three different kinds of implants. J Orthop Surg Res.
10:1762015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang J, Nie J, Ma X, Wei Y, Peng Y and Wei
X: Targeting PI3K in cancer: Mechanisms and advances in clinical
trials. Mol Cancer. 18:262019. View Article : Google Scholar : PubMed/NCBI
|