Introduction
Gestational diabetes mellitus (GDM) is one of the
common complications noted in the perioperative period, which
affects normal glucose metabolism before pregnancy (1). Different degrees of abnormal glucose
tolerance and elevated fasting blood glucose (FBG) have been
reported during pregnancy. GDM is a serious life-threatening
disease affecting the mother and fetus and its incidence is
increasing every year (2) in China.
High blood glucose (BG) may increase the incidence of premature
delivery and miscarriage. However, the pathogenesis of GDM is still
unclear (2). The majority of
previous studies have demonstrated that insulin resistance (IR) is
one of the key factors involved in the pathogenesis of GDM. In
addition, inflammatory factors and oxidative stress also play an
important role in this process (3,4).
Several studies have reported that the expression levels of
antioxidant genes thioredoxin reductase 1 cytoplasmic (TXNRD1) and
superoxide dismutase (SOD) are upregulated in type 2 diabetes
(5,6).
Moreover, a direct link between GDM and persistent
inflammation has been reported in pregnant women (7). Previous studies have also demonstrated
that GDM induces abnormal microRNA (miRNA/miR) expression (8,9).
miRNAs can be used as diagnostic tools for GDM and may affect the
regulation of several cell functions (10). The expression levels of miR-875-5p
have been demonstrated to be downregulated in patients with GDM
(11). It has also been revealed
that miR-875-5p can be used as a biomarker for GDM, suggesting its
vital role in the diagnosis of this disease (11). It has also been illustrated that
miR-876-5p is involved in the development of cancer and that it can
be used for the development of potential novel anticancer therapies
(12,13). TXNRD1 gene encodes thioredoxin
reductase, and is also known as the TrxR1 gene. TrxR1 is a member
of the thioredoxin system, which plays an important role in redox
regulation and antioxidant defense by reducing Trx levels in order
to maintain the reduction state of intracellular proteins (14). TXNRD1 is predominantly present in
the cytoplasm. Cells can fight oxidative stress by upregulating
TXNRD1 levels to eliminate hydrogen peroxide and inhibit apoptosis.
A previous study indicated that the increase in TXNRD1 levels could
play a significant role in treating patients with uncontrolled
diabetes (15,16). The TrxR/Trx system plays an
important role in the inhibition of oxidative stress, reduction of
apoptosis (17), DNA synthesis and
the regulation of the NF-κB signaling pathway (18,19). A
close association has been demonstrated between the inflammatory
response and TXNRD1 (20).
Inflammation can regulate TXNRD1 production (21). Therefore, the present study aimed to
assess the mechanism of action of miR-875-5p and TXNRD1 in GDM
rats.
Materials and methods
Animals
A total of 200 Wistar rats (weight, 180–270; age, 8
weeks; female, 100, male, 100) were purchased from Junke Biological
Co., Ltd. The female rats were fed with normal feed (control group,
n=10 rats) or a high fat and high sucrose diet (GDM group, n=90
rats) for 8 weeks at 23–25°C with a relative humidity of 65–70% in
a 12 h light/dark cycle. Subsequently, female rats that were in
proestrus were caged with males overnight. The rats were identified
as pregnant (day 0) according to the observed sperm or mucus plug
by microscopy the following day. A total of 100 female rats were
identified to be pregnant. The rats in the GDM group were injected
intraperitoneally with streptozotocin (35 mg/kg, Sigma-Aldrich;
Merck KGaA) to induce gestational diabetes. If the FBG level was
stable at 13.5 mmol/l following 24 and 72 h, the GDM rat model was
considered to have been successfully established (22). GDM rats were randomly divided into
the following groups (n=10 per group): i) GDM group, no treatment;
ii) GDM+vector group, 4 days following induction, rats were
injected with miR-875-5p inhibitor negative control (NC, 10 µM
vector) every 3 days via the tail vein; and iii) GDM+miR-875-5p
inhibitor group, 4 days following induction, rats were injected
with miR-875-5p inhibitor (10 µM vector) every 3 days via the tail
vein. To further investigate the molecular mechanism of miR-875-5p
and TXNRD1, the remaining GDM rats were randomly divided into five
groups (n=10 per group): i) GDM, no treatment; ii) GDM+sh-NC; iii)
GDM+sh-TXNRD1; iv) GDM+miR-875-5p inhibitor+sh-NC (100 µg
pGPU6/GFP/Neo-shRNA-NC); and v) GDM+miR-875-5p inhibitor+sh-TXNRD1
(100 µg pGPU6/GFP/Neo-shTXNRD1). The sequences used for the
inhibition of miR-875-5p or TXNRD1 were as follows: miR-875
inhibitor, 5′-CACCUGAUAAAACUGAGGUAUA-3′; inhibitor NC,
5′-AUCGACAGGGUUAACUCCACGA-3′; sh-TXNRD1 forward,
5′-CACCGCAATGATCTTGAGTTCTATGCGAACATAGAACTCAAGATCATTGC-3′ and
reverse, 5′-AAAAGCAATGATCTTGAGTTCTATGTTCGCATAGAACTCAAGATCATTGC-3′;
shRNA-NC forward,
5′-CACCGACATACTCTGTTAGTGTAGTCGAAACTACACTAACAGAGTATGTC-3′ and
reverse, 5′-AAAAGACATACTCTGTTAGTGTAGTTTCGCACTACACTAACAGAGTATGTC-3′.
All sequences were obtained from Guangzhou RiboBio Co., Ltd.
After 7 days, the rats were anesthetized by
intraperitoneal injection of 1% pentobarbital sodium (50 mg/kg),
followed by exsanguination 10 h after fasting. After anesthesia,
blood (5 ml) was collected from the abdominal aorta for subsequent
experiments. Blood, pancreatic islets and placental tissues of the
rats were collected. After the blood samples were collected, they
were stored overnight in the refrigerator at 4°C. Subsequently, the
serum was collected by centrifugation at 4°C at 1,409 × g for 15
min and divided into 100 µl samples, which were stored at −80°C.
Other tissues and organs were separated and stored at −80°C or
fixed in 4% formaldehyde solution at room temperature for 48 h. The
experimental process is presented in Fig. S1. All experimental procedures were
approved by the Animal Experimental Ethics Committee of The First
Hospital of Lanzhou University (approval no. LDYYLL2020-207).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted using the TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocol, and reverse transcribed into cDNA
using miScript II RT Kit (Shanghai Beinuo Biotechnology Co., Ltd.)
at 37°C for 20 min. miRNA was amplified using the miScript SYBR
Green PCR Kit (Shanghai Beinuo Biotechnology Co., Ltd.). The other
part of collected serum or pancreatic islet tissues was used to
reverse transcribe into cDNA with the PrimeScript RT Reagent Kit
(Takara Bio, Inc.) according to the manufacturer's protocol. mRNA
was amplified using the SYBR Premix ExTaq (Takara Bio, Inc.). The
thermocycling conditions were performed for qPCR as follows:
Initial denaturation at 95°C for 3 min, followed by 45 cycles of
95°C for 5 sec, 60°C for 20 sec and 72°C for 15 sec. The Cq value
of the target gene of each sample was normalized against U6 or
GADPH, and the relative expression levels were measured using the
2−ΔΔCq method (23). The
primer sequences used were as follows: miR-875-5p forward,
5′-TATACCTCAGTTTTATCAGGTG-3′ and reverse,
5′-GCGGCCGCGTGCATAGCTTCTGTAAAGG-3′; TXNRD1 forward,
5′-CTTCCACGTACTGGGTCCAAATG-3′ and reverse,
5′-TCACCGACAGCGTTGTAAATATCTC-3′; CRP forward,
5′-CATCTGTGCCACCTGGGAGTC-3′ and reverse, AAGCCACCGCCATACGAGTC-3′;
TNF-α forward, 5′-TGAGCACAGAAAGCATGATC-3′ and reverse,
5′-CATCTGCTGGTACCACCAGTT-3′; IL-6 forward,
5′-GGCCCTTGCTTTCTCTTCG-3′ and reverse, 5′-ATAATAAAGTTTTGATTATGT-3′;
U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′; and GADPH forward,
5′-TCACCATCTTCCAGGAGCGA-3′ and reverse, 5′- ACGCCAGTAGACTCCACG
ACA-3′.
Detection of FBG, serum fasting
insulin (FINS), homeostatic model assessment of insulin resistance
(HOMA-IR), total cholesterol (TC), triglyceride (TG) and
high-density lipoprotein (HDL) levels
Following fasting for 10 h, the blood samples of the
rats in each group were collected from the abdominal aorta (5 ml).
The blood samples were placed on blood glucose test strips to
record FBG levels (Nova Biomedical Corporation). The detection of
FINS was performed by enzyme linked immunosorbent assay (cat. no.
hzA448Ra; Shanghai Huzhen Biological Technology Co., Ltd.). The
HOMA-IR was calculated using the following formula: [FBG (mmol/l) ×
FINS (mU/l)] / 22.5. TG and HDL levels were determined by an BK-400
automatic biochemical analyzer (BioBase).
Western blot analysis
Total protein was isolated from pancreatic islet
tissue using RIPA lysis buffer (Sigma-Aldrich; Merck KGaA). The
concentration of total protein of the pancreatic islet tissue was
determined using the Bradford method. Proteins (50 µg) were
separated via SDS-PAGE on a 10% gel, and then transferred to PVDF
membranes, which were blocked using 5% skimmed milk powder for 1 h
at 4°C. The membranes were incubated with the following primary
antibodies for 2 h at room temperature: Anti-C reactive protein
(CRP; cat. no. ab207756; 1:1,000), TNF-α (cat. no. ab205587;
1:1,000), IL-6 (cat. no. ab208113; 1:1,000), TXNRD1 (cat. no.
ab124954; 1:1,000) and GAPDH (cat. no. ab9485; 1:1,000) (all
purchased from Abcam). Subsequently, membranes were incubated with
the HRP-conjugated secondary antibody (cat. no. ab7090; 1:10,000;
Abcam) for 1 h at room temperature. Chemiluminescence enhancement
reagent (Thermo Fisher Scientific, Inc.) was added to react with
the protein band at room temperature for 1 min. The results were
analyzed using the ImageJ software (version 1.46r; National
Institutes of Health).
Oxidative stress
Following blood collection, the pancreatic tissues
were removed, cut into pieces and further homogenized. Following
centrifugation at 1,917 × g for 5 min at 4°C, the supernatant was
obtained. The activity of SOD and catalase (CAT), and the content
of malondialdehyde (MDA) in the pancreatic tissues were determined
by UV-visible spectrophotometry using assay kits (SOD, cat. no.
S0086; CAT, cat. no. S0082; MDA, cat. no. S0131S; all purchased
from Beyotime Institute of Biotechnology), according to the
manufacturer's instructions. The OD value of each group was
detected at 560, 405 or 532 nm for SOD activity, CAT activity and
MDA content, respectively.
Hematoxylin and eosin (H&E)
staining
Following anesthesia of the animals with 1% sodium
pentobarbital (50 mg/kg, abdominal injection), pancreatic and
placental tissues were obtained. The tissues were fixed in 4%
formaldehyde solution at room temperature for 48 h, embedded in
paraffin (thickness of 5 µm) and stained with H&E. Hematoxylin
staining was performed for 9 min, followed by eosin staining for 2
min. The pathophysiological changes of the pancreatic and placental
tissues were observed by light microscopy (magnification,
×200).
Cell lines and luciferase reporter
assay
INS-1 cells (American Type Culture Collection) were
cultured in RPMI-1640 medium (containing 11.1 mM glucose; Thermo
Fisher Scientific, Inc.), in the presence of 10% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.), 0.11 g/l sodium pyruvate,
50 µM β-mercaptoethanol, and 1% penicillin-streptomycin. The cells
were maintained at 37°C with 5% CO2. Cells were seeded
into 96-well plates at a density of 40–60% and cultured in
RPMI-1640 medium without penicillin-streptomycin for 24 h, and
subsequently transfected with specific plasmids. miR-875-5p mimic
(5′-UAUACCUCAGUUUUAUCAGGUG-3′; 50 nM; Guangzhou RiboBio Co., Ltd.)
or scrambled miR-875-5p (5′-AUGCAAGGUCGGGGUAACCUCC-3′; 50 nM;
Guangzhou RiboBio Co., Ltd.) were transfected into INS-1 cells with
Lipofectamine® 2000 for 48 h at 37°C (Invitrogen; Thermo
Fisher Scientific, Inc.). Then, cells were collected and the
expression of miR-875-5p was determined using RT-qPCR. The
relationship between miR-875-5p and the 3′ untranslated region
(UTR) of TXNRD1 was predicted using Starbase (starbase.sysu.edu.cn/). The interaction was further
confirmed by performing dual-luciferase reporter assays. Mutant
(MUT) 3′UTR of TRXR1 dual-luciferase reporter vectors (Zeye
Bio-Web) were constructed using a Site-directed Mutagenesis Kit
(cat. no. B639281; Sangon Biotech Co., Ltd.). Wild-type (WT) or MUT
vectors and miR-875-5p mimic or scrambled miR-875-5p were
co-transfected into the cells using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). At 48 h
post-transfection, luciferase activities were detected using a
Luciferase Reporter Assay Kit (BioVision, Inc.) and a
GloMax® Discover Microplate Reader (Promega
Corporation). Firefly luciferase activities were normalized to
Renilla luciferase activities.
Statistical analysis
Data were analyzed using GraphPad Prism software
(version 8; GraphPad Software, Inc.). The data are presented as the
mean ± SD. The comparisons among different groups were performed
using ANOVA followed by Tukey's post hoc test. The unpaired
Student's test was performed between two groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-875-5p regulates FBG, IR and blood
lipid levels in GDM rats
The miR-875-5p expression levels in GDM rats were
initially analyzed by RT-qPCR. The results indicated that the serum
miR-875-5p levels of GDM rats were significantly decreased compared
with those of the control group (Fig.
1A). Subsequently, the functions of miR-875-5p were assessed
through a series of experiments. miR-875-5p knockdown vectors were
injected into GDM rats through the tail vein. Serum miR-875-5p
levels were significantly reduced following transfection with the
miR-875-5p inhibitor (Fig. 1B).
Furthermore, the miR-875-5p inhibitor significantly decreased FBG
and FINS levels in GDM rats compared with those in the GDM group
(Fig. 1C). In addition, GDM-induced
TC and TG levels were significantly decreased, whereas the level of
HDL was increased in the miR-875-5p inhibitor group compared with
the GDM group (Fig. 1D). The
results implied that decreased miR-875-5p expression levels played
protective roles in reducing FBG, IR and blood lipid levels in GDM
rats.
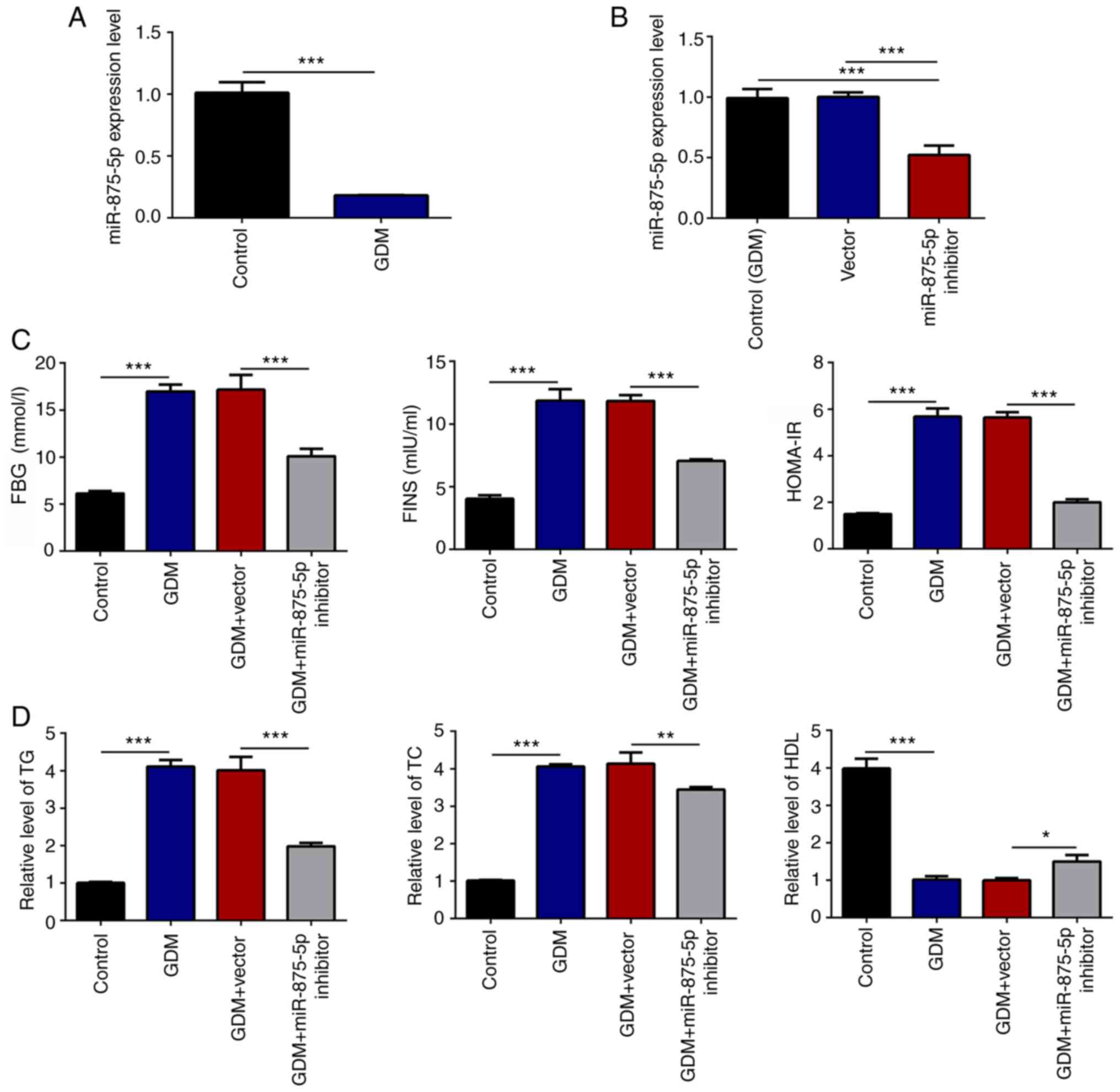 | Figure 1.miR-875-5p knockdown reduces FBG, IR
and blood lipid levels in GDM rats. (A) miR-875-5p levels in serum
were significantly reduced in GDM rats, as determined by RT-qPCR.
(B) Transfection with miR-875-5p inhibitor significantly reduced
miR-875-5p expression in the serum of rats, as analyzed by RT-qPCR.
(C) miR-875-5p inhibitor significantly reduced FBG, FINS and
HOMA-IR blood levels. (D) Blood lipids levels were significantly
affected by transfection with the miR-875-5p inhibitor. n=10, data
are presented as the mean ± SD. *P<0.05, **P<0.01,
***P<0.001. miR, microRNA; GDM, gestational diabetes mellitus;
RT-qPCR, reverse transcription-quantitative PCR; FBG, fasting blood
glucose; FINS, serum fasting insulin; HOMA-IR, homeostatic model
assessment of insulin resistance; TG, triglyceride; TC, total
cholesterol; HDL, high-density lipoprotein. |
miR-875-5p inhibitor reduces
inflammation in the pancreatic islet tissues of GDM rats
CRP is a non-specific sensitive indicator of the
inflammatory response that is closely associated with IR (24). CRP can inhibit the activity of the
insulin receptor tyrosine kinase and induce the phosphorylation
reaction of insulin receptor substrates (25). Therefore, the insulin synthesis and
secretory mechanism are disrupted, which eventually lead to IR.
TNF-α can enhance IR by directly inhibiting the expression of the
glucose transporter T4 in adipocytes. IL-6 can reduce the
sensitivity of the body to insulin and promote the secretion of
growth hormones and glucocorticoids, which result in increased IR
and BG levels (26,27). Therefore, the present study analyzed
the expression levels of CRP, TNF-α and IL-6 in the pancreatic
islet tissues in order to assess the effects of miR-875-5p on
inflammation. The results indicated that miR-875-5p inhibition
significantly reduced CRP, TNF-α and IL-6 expression levels in GDM
rats (Fig. 2A and B). The induction
of oxidative stress in the pancreatic tissues was evaluated by
analysis of SOD, CAT and MDA levels (Fig. 2C). The results indicated that the
enzyme activity of SOD was significantly increased following
suppression of miR-875-5p levels in GDM rats compared with that of
the GDM group (Fig. 2C). The levels
of the anti-oxidative stress marker CAT were also significantly
increased, whereas the concentration of MDA was decreased.
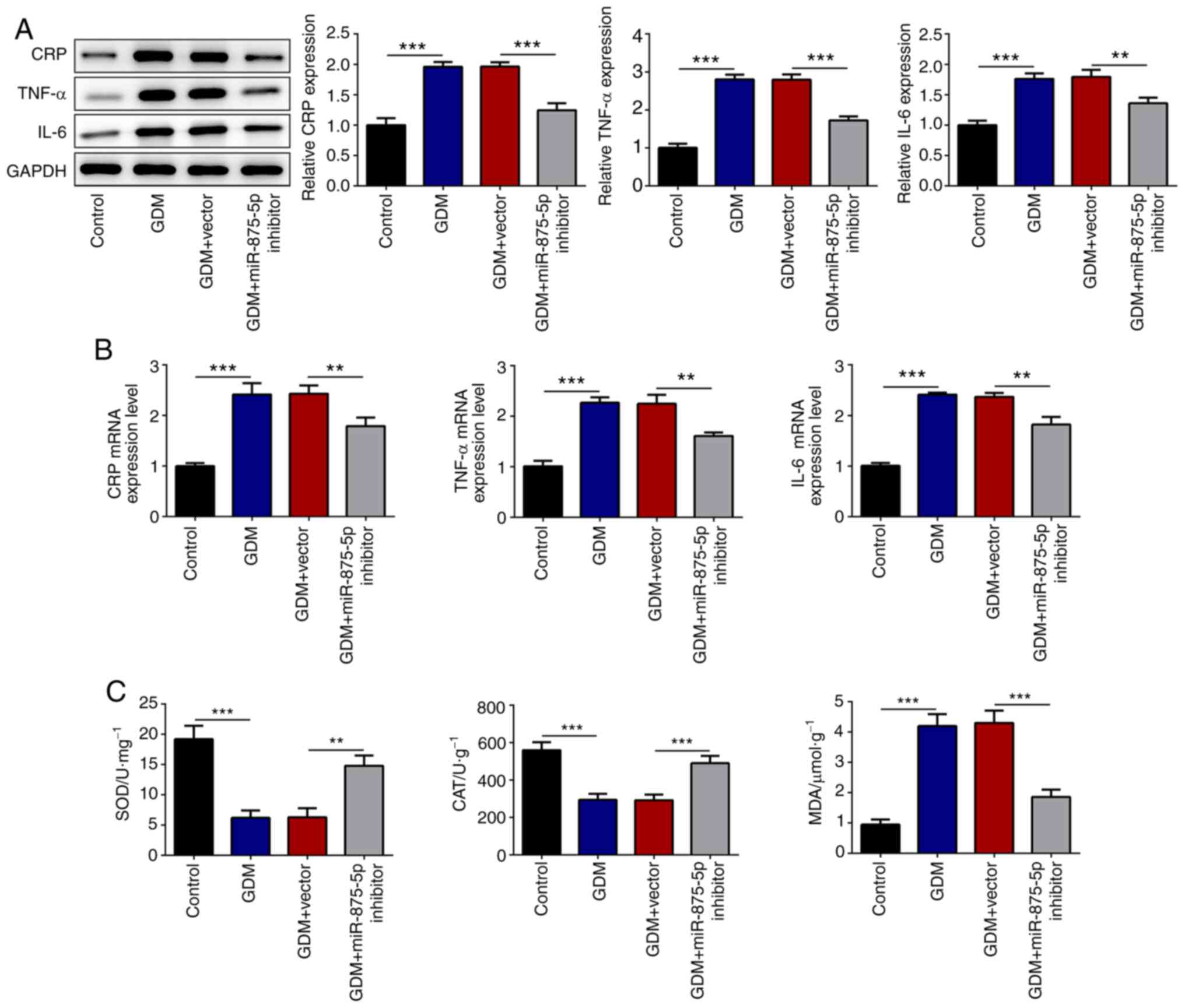 | Figure 2.miR-875-5p inhibitor significantly
reduces the levels of inflammatory markers and alters the levels of
oxidative stress mediator. (A) Transfection with miR-875-5p
inhibitor significantly reduced the protein expression of
pro-inflammatory markers in the pancreatic islet tissues, as
determined by western blotting. (B) Transfection with miR-875-5p
inhibitor significantly reduced the mRNA expression of
pro-inflammatory markers in the pancreatic islet tissues, as
determined by reverse transcription-quantitative PCR. (C) Levels of
oxidative stress markers were significantly altered following
transfection with miR-875-5p inhibitor. n=10, data are presented as
the mean ± SD. **P<0.01, ***P<0.001. miR, microRNA; GDM,
gestational diabetes mellitus; CRP, C reactive protein; SOD,
superoxide dismutase; CAT, catalase; MDA, malondialdehyde. |
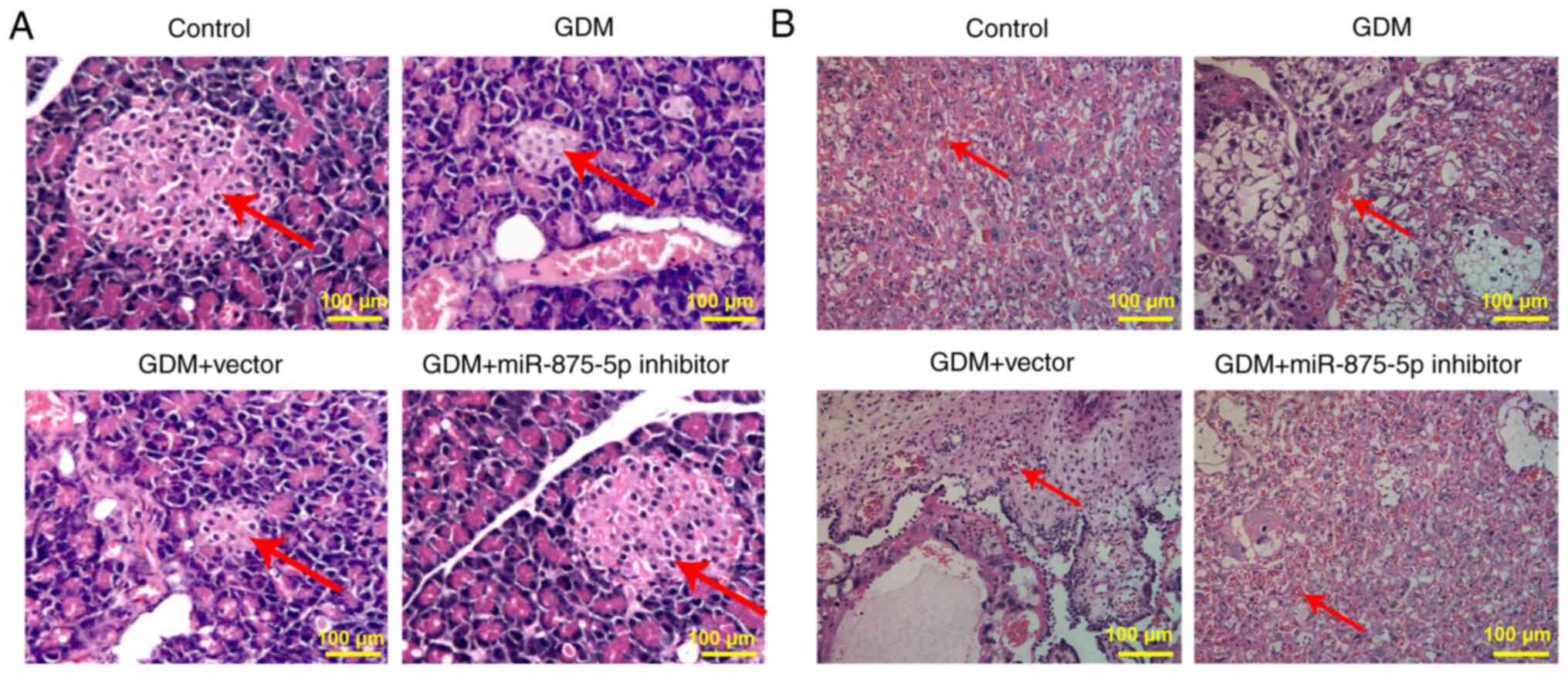
miR-875-5p inhibitor alleviates the
pathological damage on the pancreatic and placental tissues of GDM
rats
The pancreatic tissue presented normal morphology in
the control group, whereas in the GDM group it demonstrated notable
pathological changes, accompanied by varying degrees of
inflammatory cell infiltration. The islets were markedly atrophic,
with a reduced number of cells, sparse distribution and disordered
arrangement. Following treatment of the animals with the miR-875-5p
inhibitor, the pancreatic tissue lesions were notably decreased
(Fig. 3A). In the control group,
maturation of placental cell differentiation was noted, whereas the
distribution was uniform and dense and the size of intercellular
space was uniform and regular. The capillary blood vessels were
evenly distributed and a large number of red blood cells were
observed in the blood vessels. The cells in the placenta of the
rats in the GDM group (Fig. 3B)
were loosely distributed and disordered and the intercellular space
was increased. Capillary distribution was reduced and the presence
of red blood cells was rare. However, miR-875-5p inhibitor
treatment markedly reduced placental injury in GDM rats.
miR-875-5p regulates the expression
levels of TXNRD1
Subsequently, the effects of miR-875-5p on TXNRD1
expression were evaluated. First, the expression of miR-875-5p was
determined in INS-1 cells following transfection with a miR-875-5p
mimic. A significant increase in the expression of miR-875-5p was
observed in INS-1 cells with miR-875-5p mimic (Fig. 4A). Compared with the GDM+vector
group, miR-875-5p inhibitor significantly upregulated the levels of
TXNRD1 in the serum of GDM rats (Fig.
4B), indicating that miR-875-5p could regulate the expression
of TXNRD1 in GDM rats. The StarBase database predicted that
miR-875-5p could bind to the 3′UTR of TXNRD1, which was further
confirmed using a dual-luciferase reporter gene assay. The
luciferase activity in the TXNRD1 WT vector co-transfected with
miR-875-5p mimic was significantly reduced compared with that of
the vector group, whereas no differences were observed in the
TXNRD1 MUT vectors co-transfected with miR-875-5p mimic or
scrambled miR-875-5p (Fig. 4C).
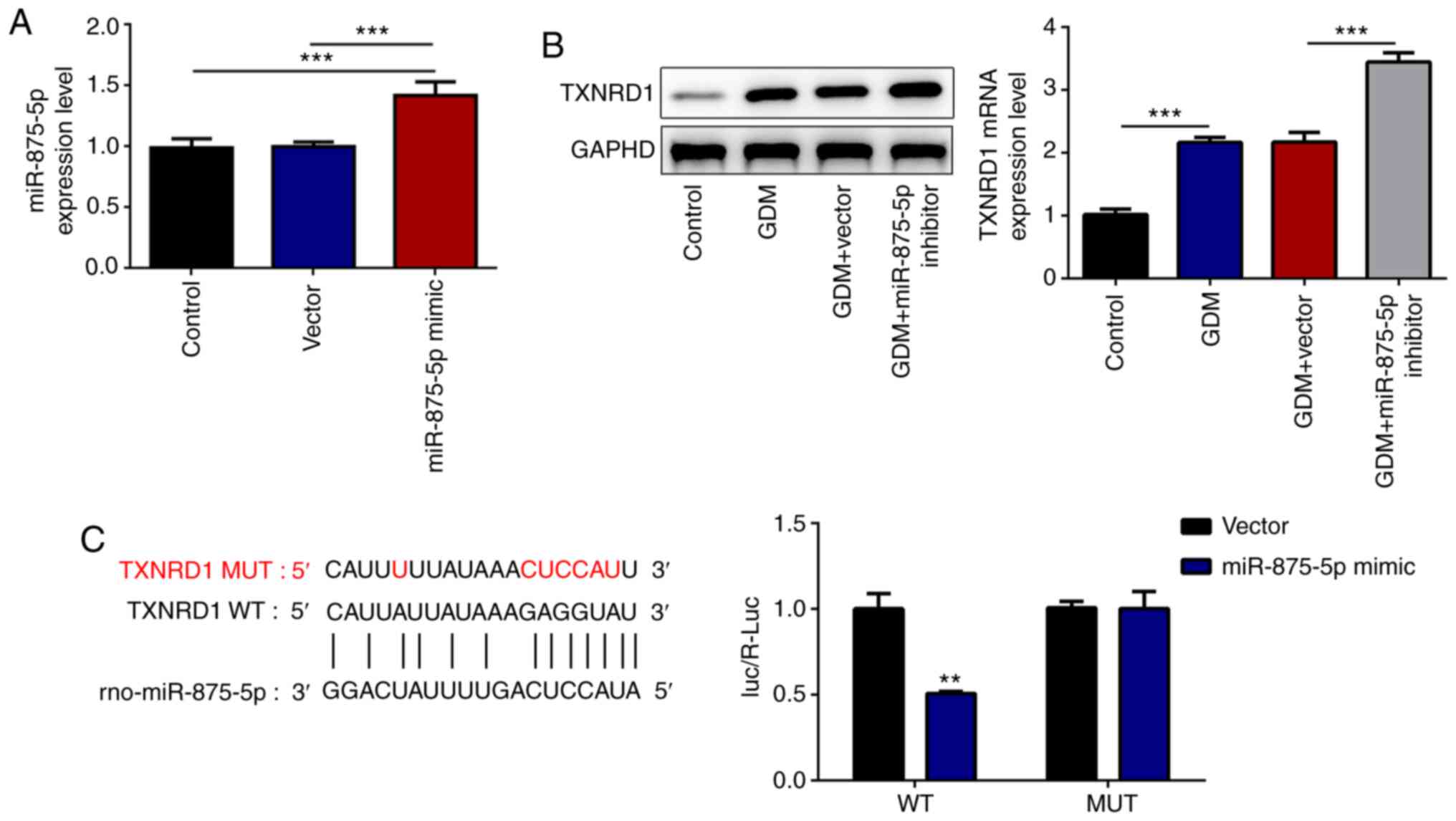 | Figure 4.miR-875-5p knockdown upregulates the
transcription and translation levels of TXNRD1. (A) Following
transfection of INS-1 cells with miR-875-5p mimic, miR-875-5p
expression significantly increased. (B) Transfection with
miR-875-5p inhibitor significantly increased the TXNRD1 expression
in the serum of GDM rats. (C) StarBase was used to predict that
miR-875-5p can bind to the 3′ untranslated region of TXNRD1, and
the luciferase activity was significantly reduced by transfection
with the miR-875-5p mimic in the WT group. n=10, data are presented
as the mean ± SD. **P<0.01, ***P<0.001 vs. miR-875-5p-NC.
miR, microRNA; GDM, gestational diabetes mellitus; TXNRD1,
thioredoxin reductase 1 cytoplasmic; WT, wild-type; MUT, mutant;
NC, negative control. |
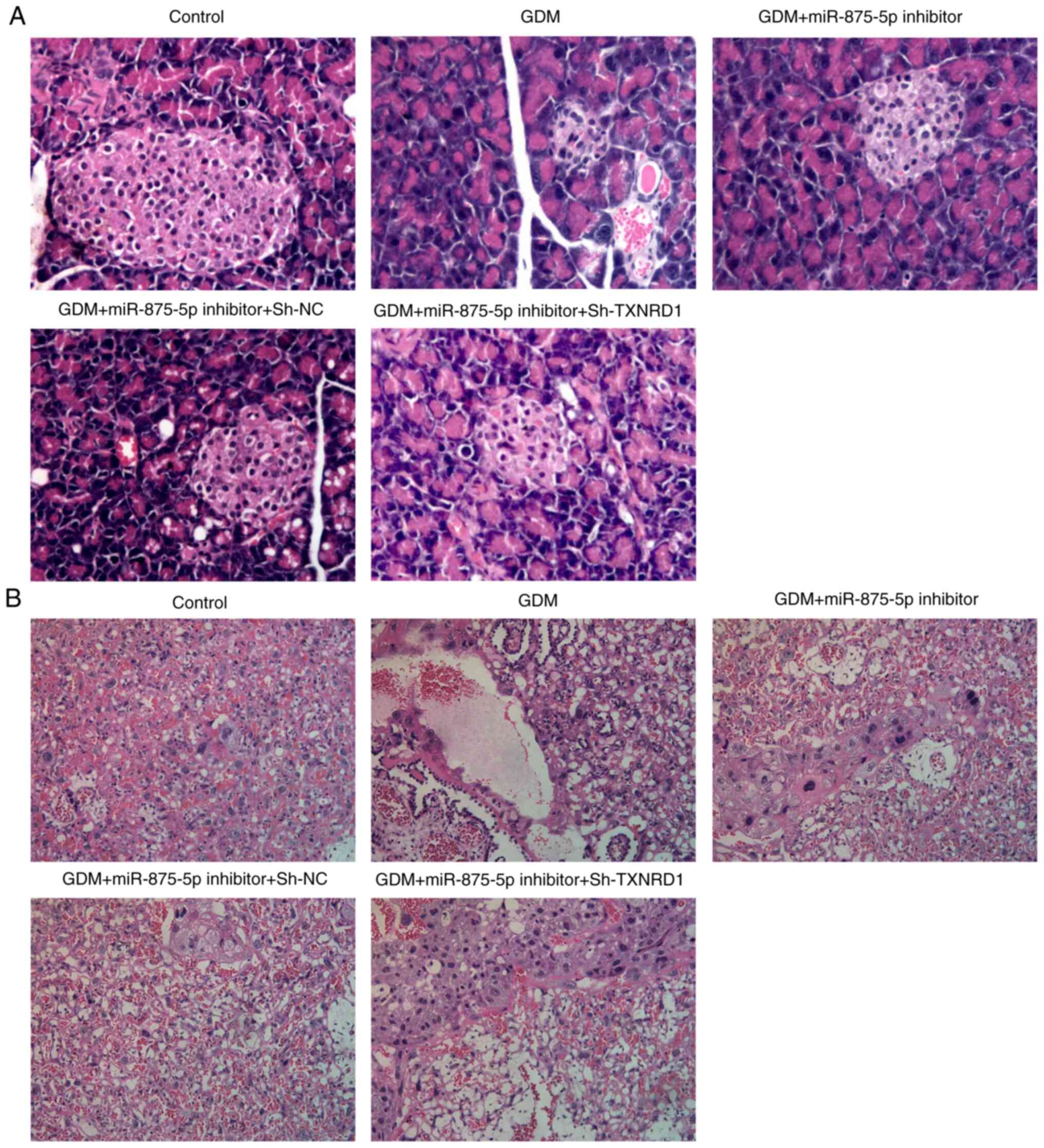
TXNRD1 inhibitor reverses the effects
of miR-875-5p on GDM rats
Cells were transfected with sh-TXNRD1, which
significantly suppressed the expression of TXNRD1 in the serum of
GDM rats (Fig. 5A and B).
Transfection with sh-TXNRD1 significantly reversed the effects of
miR-875-5p inhibitor on glucose, FINS and HOMA-IR (Fig. 5C), inflammatory indicators (CRP,
TNF-α and IL-6; Fig. 5D and E), and
oxidative stress levels (SOD, CAT and MDA; Fig. 5F). In addition, TXNRD1 knockdown
markedly altered the effects of miR-875-5p inhibitor on pancreatic
and placental injury in GDM rats (Fig.
6A and B).
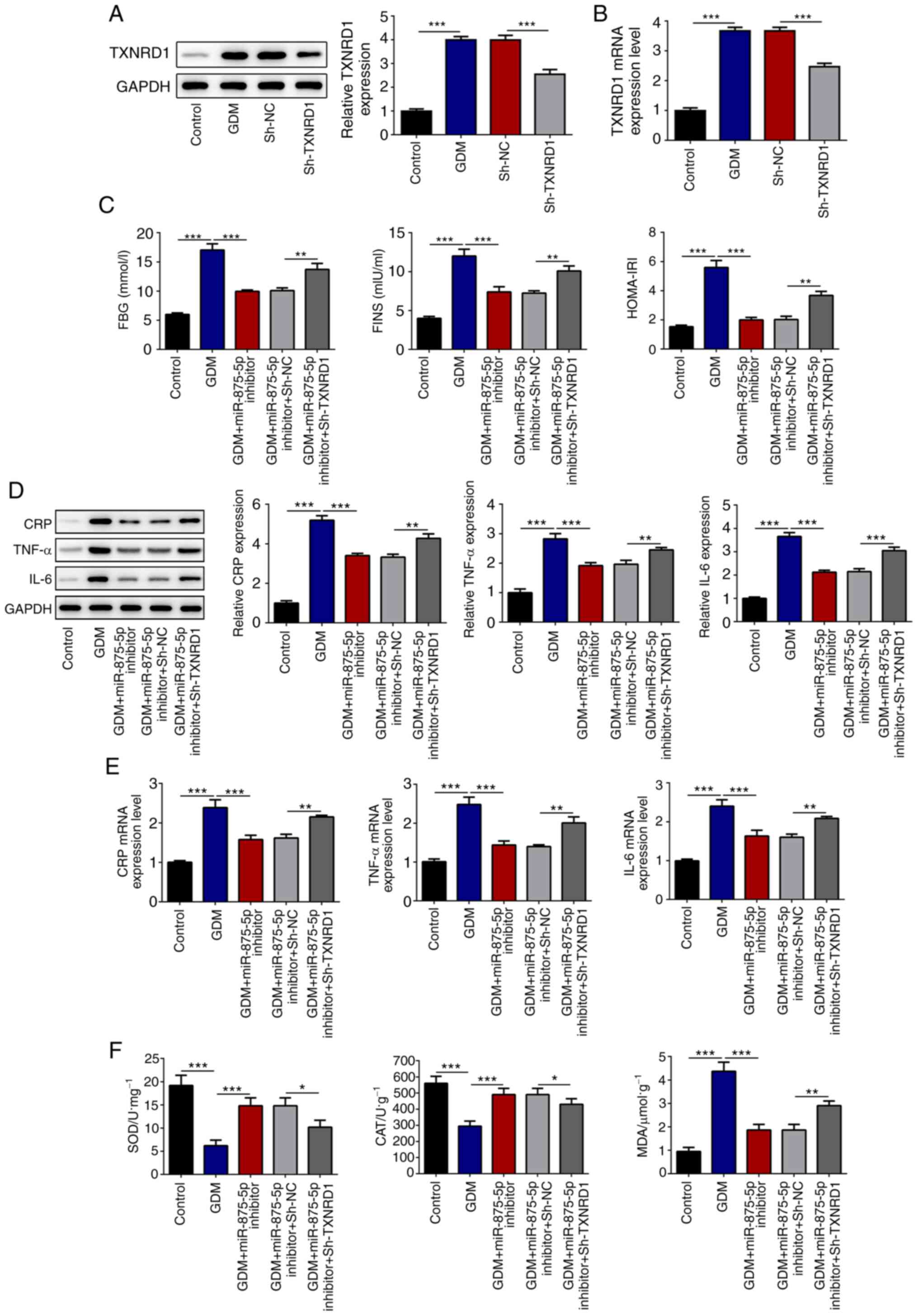 | Figure 5.TXNRD1 knockdown reverses the effects
of miR-875-5p inhibitor. (A and B) Transfection with sh-TXNRD1
suppressed the expression of TXNRD1 in the serum of GDM rats. (C)
TXNRD1 knockdown increased FBG and IR in the blood of GDM rats. (D
and E) TXNRD1 knockdown increased expression levels of
pro-inflammatory markers in the pancreatic islet tissues. (F)
Oxidative stress levels in the pancreatic islet tissues were
altered by TXNRD1 knockdown. n=10, data are presented as the mean ±
SD. *P<0.05, **P<0.01, ***P<0.001. miR, microRNA; GDM,
gestational diabetes mellitus; sh-, short hairpin RNA; TXNRD1,
thioredoxin reductase 1 cytoplasmic; FBG, fasting blood glucose;
FINS, serum fasting insulin; HOMA-IR, homeostatic model assessment
of insulin resistance; CRP, C reactive protein; SOD, superoxide
dismutase; CAT, catalase; MDA, malondialdehyde; NC, negative
control. |
Discussion
GDM is a serious life-threatening disease affecting
the mother and fetus that displays an increasing incidence annually
in China (2). High BG contributes
to an increase in the incidence of premature delivery and abortion
(28). The present study indicated
that transfection with miR-875-5p inhibitor played protective
effects by reducing FBG, IR and blood lipid levels, along with
improved pancreatic and placental injury in GDM rats. In addition,
the levels of the pro-inflammatory and oxidative stress markers
were significantly reduced, which implied that the miR-875-5p
inhibitor could reduce inflammation in pancreatic islets and
oxidative stress in the liver of GDM rats. The data indicated that
the effects of miR-875-5p inhibitor could be significantly reversed
by TXNRD1 inhibition. Therefore, TXNRD1 played a protective role in
GDM rats. Due to the low expression of miR-875-5p noted in the
plasma of GDM rats under normal physiological conditions, the
expression of TXNRD1 may be maintained at a relatively low level.
However, in GDM, the decreased expression of miR-875-5p alleviates
its inhibition to TXNRD1 mRNA, which may be one of the underlying
mechanisms for the increased expression of reactive TXNRD1 to exert
antioxidation.
TrxR has been demonstrated to play a key role in
redox regulation and antioxidant protection and is considered a
potential target in managing metabolic syndromes (29). IR is closely associated with reduced
oxidative stress in GMD. Oxidative stress is considered to be
involved in the induction of IR (30–32).
Therefore, TrxR1 can decrease IR by suppressing oxidative stress.
TrxR is also involved in the regulation of inflammatory factors,
cell proliferation, apoptosis and embryonic development (33–36).
The results indicated that the expression levels of TRXR1 mRNA and
protein were significantly increased in GDM rats. The increase in
TRXR1 mRNA levels induced by GDM may be caused by upregulation of
TrxR enzyme activity, which can activate compensatory antioxidant
defense systems of the body to protect cells from ROS-induced
damage (37,38). A previous study indicated that
increased TrxR activity was positively associated with the
upregulation of oxidative stress markers (39). The enhanced activity of the Trx/TrxR
system is considered to play protective roles in preventing further
complications of diabetes. Trx1 can reduce β-cell damage in
non-obese diabetic subjects (40).
TrxR exerts anti-oxidative functions mainly by preventing
stress-induced oxidative damage (29). Previous studies demonstrated that
the transcription factor responsive element and the 3′UTR of TRXR1
could regulate its transcriptional activation or translation
(41,42). In addition, previous studies have
shown that transcription factors, such as nuclear factor erythroid
2-related factor 2 and AP-1 regulate TRXR1 expression. In the
present study, bioinformatic prediction and molecular biology
experiments confirmed that miR-875-5p can specifically bind to
TRXR1 3′UTR and inhibit the expression of endogenous TRXR1 at the
post-transcriptional level to suppress its translation or induce
degradation.
In conlcusion, the present study showed that
miR-875-5p regulates IR and inflammation via targeting TXNRD1 in
GDM model rats. Thus, these results indicated that miR-875-5p and
TXNRD1 could be potential targets for treating GDM.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Research Fund
Project of the First Hospital of Lanzhou University (grant no.
ldyyyn2018-59), the Research Fund Project of the Science and
Technology Development Guiding Plan of Lanzhou City (grant no.
2019-ZD-38), Construction of Gansu Province Clinical Research
Center for Endocrine disease (grant no. 20JR10FA667) and the
Special Funds of Science and Technology Development of the Chinese
Central Government to Guide Local in 2020 (an innovation platform
for improving the prevention and treatment of frequently-occurring
diseases in Gansu Province).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
Songbo F made substantial contributions to the
conception and design of the study, the acquisition, analysis and
interpretation of data, the acquisition of funding, the collection
of data and general supervision of the research group. Songquan F,
XM, XY and JL made substantial contributions to the acquisition,
analysis and interpretation of data. SongboF and SongquanF confirm
the authenticity of all the raw data. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
All the experimental procedures were approved by the
Animal Experimental Ethics Committee of The First Hospital of
Lanzhou University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Metzger BE and Coustan DR; The Organizing
Committee, : Summary and recommendations of the Fourth
International Workshop-Conference on Gestational Diabetes Mellitus.
Diabetes Care. 21 (Suppl 2):B161–B167. 1998.PubMed/NCBI
|
|
2
|
Johns EC, Denison FC, Norman JE and
Reynolds RM: Gestational Diabetes Mellitus: Mechanisms, Treatment,
and Complications. Trends Endocrinol Metab. 29:743–754. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hajifaraji M, Jahanjou F, Abbasalizadeh F,
Aghamohammadzadeh N, Abbasi MM and Dolatkhah N: Effect of probiotic
supplements in women with gestational diabetes mellitus on
inflammation and oxidative stress biomarkers: A randomized clinical
trial. Asia Pac J Clin Nutr. 27:581–591. 2018.PubMed/NCBI
|
|
4
|
Sha H, Zeng H, Zhao J and Jin H:
Mangiferin ameliorates gestational diabetes mellitus-induced
placental oxidative stress, inflammation and endoplasmic reticulum
stress and improves fetal outcomes in mice. Eur J Pharmacol.
859:1725222019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Baig S, Rizi EP, Shabeer M and Agrawal M:
Heredity of type 2 diabetes confers increased susceptibility to
oxidative stress and inflammation. BMJ Open Diabetes Res Care.
8:e0009452020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nazem MR, Asadi M, Jabbari N and Allameh
A: Effects of zinc supplementation on superoxide dismutase activity
and gene expression, and metabolic parameters in overweight type 2
diabetes patients: A randomized, double-blind, controlled trial.
Clin Biochem. 69:15–20. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sudharshana Murthy KA, Bhandiwada A,
Chandan SL, Gowda SL and Sindhusree G: Evaluation of Oxidative
Stress and Proinflammatory Cytokines in Gestational Diabetes
Mellitus and Their Correlation with Pregnancy Outcome. Indian J
Endocrinol Metab. 22:79–84. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Elliott HR, Sharp GC, Relton CL and Lawlor
DA: Epigenetics and gestational diabetes: a review of epigenetic
epidemiology studies and their use to explore epigenetic mediation
and improve prediction. Diabetologia. 62:2171–2178. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vasu S, Kumano K, Darden CM, Rahman I,
Lawrence MC and Naziruddin B: MicroRNA Signatures as Future
Biomarkers for Diagnosis of Diabetes States. Cells. 8:E15332019.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao H and Tao S: miRNA-221 protects islet
beta cell function in gestational diabetes mellitus by targeting
PAK1. Biochem Biophys Res Commun. 520:218–224. 2008. View Article : Google Scholar
|
|
11
|
Zamanian Azodi M, Rezaei-Tavirani M,
Rezaei-Tavirani M and Robati RM: Gestational Diabetes Mellitus
Regulatory Network Identifies hsa-miR-145-5p and hsa-miR-875-5p as
Potential Biomarkers. Int J Endocrinol Metab. 17:e866402019.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu Q, Zhu Q, Zhou Z, Wang Y, Liu X, Yin G,
Tong X and Tu K: MicroRNA-876-5p inhibits epithelial-mesenchymal
transition and metastasis of hepatocellular carcinoma by targeting
BCL6 corepressor like 1. Biomed Pharmacother. 103:645–652. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang T, Cai X, Li Q, Xue P, Chen Z, Dong
X and Xue Y: Hsa-miR-875-5p exerts tumor suppressor function
through down-regulation of EGFR in colorectal carcinoma (CRC).
Oncotarget. 7:42225–42240. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cebula M, Schmidt EE and Arnér ES: TrxR1
as a potent regulator of the Nrf2-Keap1 response system. Antioxid
Redox Signal. 23:823–853. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kabuyama Y, Kitamura T, Yamaki J, Homma
MK, Kikuchi SI and Homma Y: Involvement of thioredoxin reductase 1
in the regulation of redox balance and viability of rheumatoid
synovial cells. Biochem Biophys Res Commun. 367:491–496. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chang E, Kim DH, Yang H, Lee DH, Bae SH
and Park CY: CB1 receptor blockade ameliorates hepatic fat
infiltration and inflammation and increases Nrf2-AMPK pathway in a
rat model of severely uncontrolled diabetes. PLoS One.
13:e02061522018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Park HR, Lee SE, Yang H, Son GW, Jin YH
and Park YS: Induction of Thioredoxin Reductase 1 by Korean Red
Ginseng Water Extract Regulates Cytoprotective Effects on Human
Endothelial Cells. Evid Based Complement Alternat Med.
2015:9720402015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Raninga PV, Di Trapani G, Vuckovic S and
Tonissen KF: TrxR1 inhibition overcomes both hypoxia-induced and
acquired bortezomib resistance in multiple myeloma through NF-кβ
inhibition. Cell Cycle. 15:559–572. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Matsui M, Oshima M, Oshima H, Takaku K,
Maruyama T, Yodoi J and Taketo MM: Early embryonic lethality caused
by targeted disruption of the mouse thioredoxin gene. Dev Biol.
178:179–185. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shi X, Wang W, Zheng S, Zhang Q and Xu S:
Selenomethionine relieves inflammation in the chicken trachea
caused by LPS though inhibiting the NF-κB pathway. Biol Trace Elem
Res. 194:525–535. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ingram S, Mengozzi M, Heikal L, Mullen L
and Ghezzi P: Inflammation-induced reactive nitrogen species cause
proteasomal degradation of dimeric peroxiredoxin-1 in a mouse
macrophage cell line. Free Radic Res. 53:875–881. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
John CM, Ramasamy R, Al Naqeeb G,
Al-Nuaimi AH and Adam A: Nicotinamide supplementation protects
gestational diabetic rats by reducing oxidative stress and
enhancing immune responses. Curr Med Chem. 19:5181–5186. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bateman RM, Sharpe MD, Jagger JE, Ellis
CG, Solé-Violán J, López-Rodríguez M, Herrera-Ramos E,
Ruíz-Hernández J, Borderías L, Horcajada J, et al: 36th
International Symposium on Intensive Care and Emergency Medicine:
Brussels, Belgium. 15–18 March 2016. Crit Care. 20:942016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kaushik SV, Plaisance EP, Kim T, Huang EY,
Mahurin AJ, Grandjean PW and Mathews ST: Extended-release niacin
decreases serum fetuin-A concentrations in individuals with
metabolic syndrome. Diabetes Metab Res Rev. 25:427–434. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Skórzyńska-Dziduszko KE, Kimber-Trojnar Ż,
Patro-Małysza J, Olszewska A, Zaborowski T and Małecka-Massalska T:
An Interplay between Obesity and Inflammation in Gestational
Diabetes Mellitus. Curr Pharm Biotechnol. 17:603–613. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Roca-Rodríguez MDM, López-Tinoco C,
Fernández-Deudero Á, Murri M, García-Palacios MV, García-Valero
MDA, Tinahones FJ and Aguilar-Diosdado M: Unfavorable cytokine and
adhesion molecule profiles during and after pregnancy, in women
with gestational diabetes mellitus. Endocrinol Diabetes Nutr.
64:18–25. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Poston L, Caleyachetty R, Cnattingius S,
Corvalán C, Uauy R, Herring S and Gillman MW: Preconceptional and
maternal obesity: epidemiology and health consequences. Lancet
Diabetes Endocrinol. 12:1025–1036. 2016. View Article : Google Scholar
|
|
29
|
Tinkov AA, Bjørklund G, Skalny AV,
Holmgren A, Skalnaya MG, Chirumbolo S and Aaseth J: The role of the
thioredoxin/thioredoxin reductase system in the metabolic syndrome:
Towards a possible prognostic marker? Cell Mol Life Sci.
75:1567–1586. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Feng Y, Feng Q, Qu H, Song X, Hu J, Xu X,
Zhang L and Yin S: Stress adaptation is associated with insulin
resistance in women with gestational diabetes mellitus. Nutr Diab.
10:42020. View Article : Google Scholar
|
|
31
|
Rueangdetnarong H, Sekararithi R,
Jaiwongkam T, Kumfu S, Chattipakorn N, Tongsong T and Jatavan P:
Comparisons of the oxidative stress biomarkers levels in
gestational diabetes mellitus (GDM) and non-GDM among Thai
population: cohort study. Endocrine Connect. 7:681–687. 2018.
View Article : Google Scholar
|
|
32
|
Zygula A, Kosinski P, Zwierzchowska A,
Sochacka M, Wroczynski P, Makarewicz-Wujec M, Pietrzak B, Wielgos
M, Rzentala M and Giebultowicz J: Oxidative stress markers in
saliva and plasma differ between diet-controlled and
insulin-controlled gestational diabetes mellitus. Diabetes Res Clin
Pract. 148:72–80. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ren X, Zou L, Lu J and Holmgren A:
Selenocysteine in mammalian thioredoxin reductase and application
of ebselen as a therapeutic. Free Radic Biol Med. 127:238–247.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lu J and Holmgren A: The thioredoxin
antioxidant system. Free Radic Biol Med. 66:75–87. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Arnér ESJ: Focus on mammalian thioredoxin
reductases - Important selenoproteins with versatile functions.
Biochim Biophys Acta. 1790:495–526. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bobba A, Casalino E, Petragallo VA and
Atlante A: Thioredoxin/thioredoxin reductase system involvement in
cerebellar granule cell apoptosis. Apoptosis. 19:1497–1508. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sun QA, Wu Y, Zappacosta F, Jeang KT, Lee
BJ, Hatfield DL and Gladyshev VN: Redox Regulation of Cell
Signaling by Selenocysteine in Mammalian Thioredoxin Reductases. J
Biol Chem. 274:24522–24530. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
He L, He T, Farrar S, Ji L, Liu T and Ma
X: Antioxidants Maintain Cellular Redox Homeostasis by Elimination
of Reactive Oxygen Species. Cell Physiol Biochem. 44:532–553. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Calabrese V, Mancuso C, Sapienza M, Puleo
E, Calafato S, Cornelius C, Finocchiaro M, Mangiameli A, Di Mauro
M, Stella AM and Castellino P: Oxidative stress and cellular stress
response in diabetic nephropathy. Cell Stress Chaperones.
12:299–306. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chernatynskaya AV, Looney B, Hu H, Zhu X
and Xia CQ: Administration of recombinant human thioredoxin-1
significantly delays and prevents autoimmune diabetes in nonobese
diabetic mice through modulation of autoimmunity. Diabetes Metab
Res Rev. 27:809–812. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gasdaska JR, Harney JW, Gasdaska PY, Powis
G and Berry MJ: Regulation of Human Thioredoxin Reductase
Expression and Activity by 3′-Untranslated Region Selenocysteine
Insertion Sequence and mRNA Instability Elements. J Biol Chem.
274:25379–25385. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Rundlöf AK, Carlsten M and Arnér ESJ: The
core promoter of human thioredoxin reductase 1: Cloning,
transcriptional activity, and Oct-1, Sp1, and Sp3 binding reveal a
housekeeping-type promoter for the AU-rich element-regulated gene.
J Biol Chem. 276:30542–30551. 2001. View Article : Google Scholar : PubMed/NCBI
|