Introduction
Proliferative vitreoretinopathy (PVR), which is
characterized by the formation of contractile preretinal membranes
(PRMs) in the vitreous and on the inner and outer surfaces of the
detached retina, is one of the leading causes of blindness
(1–3). PVR is associated with recurrent
retinal detachment and is the main cause of failure of retinal
reattachment surgery (1). Despite
the development of advanced surgical techniques and devices, there
remain unresolved issues regarding the pathogenesis and treatment
of PVR (1). Therefore, the
identification of key molecules involved in PVR is required for
improving the clinical outcome of this disease.
PRMs result from cell dedifferentiation, migration,
adherence and proliferation and the secretion of extracellular
matrix (ECM) proteins, including collagen, following rhegmatogenous
retinal detachment (3,4). PVR represents an excessive
wound-healing response during tissue repair following eye injury.
Previous studies have demonstrated that retinal pigment epithelium
(RPE) cells, Müller glial cells and blood-derived lymphocytes are
present in the PRMs of patients with PVR (5–7). Among
these cell types, RPE cells appear to be the primary proliferative
cells that can change their phenotype and undergo the process of
epithelial-mesenchymal transition (EMT) (2,8). It
has been reported that EMT of RPE cells is triggered by a host of
growth factors, including TGF-β1, platelet-derived growth factor
and epidermal growth factor (2,9–11).
Activated RPE cells can proliferate and migrate into the vitreous
or the inner layer of the retina, producing ECM components and
being transformed from epithelial cells to fibroblast-like cells, a
biological phenomenon termed EMT, and ultimately forming a PRM
(2,12). When the PRM contracts it results in
traction to the retina (1). Apert
from stripping the PRM during surgery, no other target-oriented
interventions are available to decrease the EMT of RPE cells and
the production of ECM, thereby increasing the success rate of a PVR
operation (1,3).
Previous studies have found that growth factors are
necessary for the repair of damaged tissue; however, prolonged
production or dysregulated expression of growth factors can lead to
persistent EMT and excessive wound healing, as found in PVR
(4,13,14).
Among these growth factors, TGF-β is a key factor in EMT and matrix
remodeling (2). The role of TGF-β
is controversial in eye diseases and is associated with the
specific condition/system. Studies have demonstrated that topical
application of a TGF-β1-loaded liposomal suspension could protect
retinal tissue in rat models of age-related macular degeneration
(15,16). In PVR, TGF-β is a pivotal
contributor to tissue fibrosis (2,17).
Previous studies have focused on a downstream mediator of TGF-β
signaling, connective tissue growth factor (CTGF) (2,18–20).
CTGF, which is rich in cysteine residues, is a secreted growth
factor and was originally identified in a conditioned medium from
human umbilical vein endothelial cells using affinity
chromatography (21). CTGF is
identified as a possible key determinant of progressive tissue
fibrosis and excessive scarring, which also serves an important
role in wound repair, neoangiogenesis, tumor growth and embryonic
development (18,22,23). A
number of studies have demonstrated that CTGF can stimulate the
growth and adhesion of fibroblasts and vascular endothelial cells,
as well as promoting the secretion of ECM proteins (2,18,22,23).
Our previous study confirmed that CTGF increases the
migratory potential of RPE cells by stimulating the calcium
signaling system and serves an important role in the pathogenesis
of PVR (19). Previous studies have
also demonstrated that CTGF is a major mediator of retinal fibrosis
and potentially an effective therapeutic target for PVR (17,18).
However, the role of CTGF in the progress of PVR and the downstream
signaling mechanisms remains to be elucidated.
The present study aimed to investigate the
expression of CTGF, fibronectin and collagen type III in PVR
membranes, as well as the effects and mechanisms of CTGF and its
upstream regulator TGF-β1 in the process of EMT and ECM synthesis
by RPE cells in vitro. The findings may provide insights
into the mechanistic details of intraocular proliferative diseases
including PVR.
Materials and methods
PRM collection and preparation
A total of 26 PRM specimens were obtained from
patients who underwent a vitreoretinal surgery after having been
diagnosed with rhegmatogenous retinal detachment, accompanied by
PVR, at the Department of Ophthalmology, Xijing Hospital (Xian,
China) between March 2017 and March 2019. In accordance with the
tenets of the Declaration of Helsinki, informed consent of the
patients and the approval by the institutional review board were
obtained before sample collection. The patients ranged in age
between 19 and 67 years (12 female and 14 male) and their course of
disease ranged between 4 months and 2 years. Among the specimens
collected, six were classified as grade B membranes [moderate PVR:
Wrinkling of the inner retinal surface, a rolled edge(s) of a
retinal break(s), retinal stiffness and vascular tortuosity];
eleven were classified as grades C1-C3 (marked PVR: Full-thickness
retinal folds in 1–3 quadrants, respectively); and nine were
classified as grades D1-D3 (massive PVR: Fixed retinal folds in
four quadrants) according to the classification of PVR grades
established by the International Retinal Society in 1983 (24). All 26 specimens were fixed in 4%
(w/v) neutral formalin for 24 h at room temperature, and dehydrated
sequentially as follows: 1X into 70% ethanol (1 h at 4°C), 1X into
85% ethanol (1 h at 4°C), 2X into 95% ethanol (1 h each at 4°C),
and 2X into 100% ethanol (1 h each at 4°C). The tissues were
immersed 2X into the paraffin bath (2 h each at 60°C), then
transported to the mold with paraffin, and incubated at room
temperature until set. The tissues were prepared as 6-µm-thick
sections.
Immunohistochemical staining of
PRMs
Sections were deparaffinized in multiple changes of
xylene (Sangon Biotech Co., Ltd.) and rehydrated in a decreasing
graded ethanol series (Sangon Biotech Co., Ltd.). Endogenous
peroxidase was blocked by incubation of the sections in 3%
H2O2 (Beyotime Institute of Biotechnology)
for 10 min at room temperature. For heat-induced epitope retrieval,
the sections were immersed in 0.01 M citrate buffer, pH 6.0 (Sangon
Biotech Co., Ltd.) and heated for 10 min in a microwave, followed
by cooling for 10 min and reheating for 5 min. The sections were
then cooled for 30 min at room temperature and washed with water
and phosphate-buffered saline (PBS; Beyotime Institute of
Biotechnology). Following incubation with blocking serum (Beyotime
Institute of Biotechnology) for 30 min, the sections were incubated
with the following primary antibodies at 4°C overnight: CTGF
(1:100; cat. no. ab5097; Abcam), collagen type III (1:100; cat. no.
22734-1-AP; ProteinTech Group, Inc.) and fibronectin (1:100; cat.
no. 26836; Cell Signaling Technology, Inc.). A streptavidin/biotin
complex immunostaining kit was purchased from Wuhan Boster
Biological Technology, Ltd. Incubation with a biotinylated
secondary antibody (1:500; cat. no. ab205718; Abcam) was followed
by incubation with peroxidase-labeled streptavidin at room
temperature for 1 h (Sangon Biotech Co., Ltd.). Staining was
visualized via the reaction of samples with a diaminobenzidine
substrate-chromogen solution (Sangon Biotech Co., Ltd.). The
sections were additionally counterstained with hematoxylin at room
temperature for 2 min (Sangon Biotech Co., Ltd.), dehydrated and
mounted in Paramount (Agilent Technologies, Inc.). The primary
antibodies were omitted in negative controls and the sections were
incubated only in the diluent (PBS). All subsequent steps of the
control and experimental procedures were identical.
After the colorimetric reaction was completed, the
sections were observed under a light microscope (Eclipse E100,
Nikon Corporation). The area and intensity of staining for each
antibody was independently estimated by two researchers. A clear
background, yellow or pale brown in the cytoplasm, was a positive
signal. The number of positive cells per 100 cells was counted in
each field (magnification, ×400) and the average was calculated for
three fields in each specimen. i) The samples were scored according
to the presence of color and its darkness in the proliferative
membrane tissue as follows: 0 points for no coloration; 1 point for
light yellow; 2 points for yellow or pale brown; and 3 points for
brown. ii) The samples were scored according to the proportion of
positive cells: 0 points, <5%; 1 point, 5–30%; 2 points, 30–60%;
and 3 points, >60%. The final score for each specimen was (i +
ii)/2 and the samples were classified according to the scores as
follows: score 0, no labeling (−); score 0.5–1, weak, focal
staining (+); score 1.5–2, moderate to strong staining (++); and
score 2.5–3, intense staining (+++). No differences were found in
the data obtained by the two observers.
Immunofluorescence staining
PRMs were fixed with 4% paraformaldehyde (PFA;
Sangon Biotech Co., Ltd.) at 4°C for 1 h, incubated in 30% sucrose
at 4°C overnight, embedded in Tissue-Tek optimal cutting
temperature compound (Sakura Finetek USA, Inc.) and frozen to
prepare 8-µm-thick sections (cat. no. CM1800; Leica Microsystems
GmbH). The sections were dried at room temperature for 2 h and
blocked with PBS containing 1% bovine serum albumin (BSA, Zeta-life
Company) and 0.5% Triton X-100 for 1 h. The sections were incubated
with primary antibodies at 4°C overnight, followed by incubation
with secondary antibodies at room temperature for 1 h. Each step
was followed by washing three times with PBS for 5 min each. The
primary antibodies included CTGF (1:100; cat. no. ab5097; Abcam)
and retinal pigment epithelium-specific 65 kDa protein (RPE65;
1:100; cat. no. MA1-16578; Invitrogen; Thermo Fisher Scientific,
Inc.). The secondary antibodies included Cy3-conjugated goat
anti-rabbit IgG (H+L; 1:100; cat. no A10522; Invitrogen; Thermo
Fisher Scientific, Inc.) and Alexa Fluor 488-conjugated goat
anti-mouse IgG (H+L; 1:100; cat. no. ab150113; Abcam).
Human RPE cells were fixed with 4% PFA at 4°C for 30
min and then blocked with PBS containing 1% BSA and 0.5% Triton
X-100 at room temperature for 1 h. The primary antibody was RPE65
(1:100; cat. no. MA1-16578, Invitrogen; Thermo Fisher Scientific,
Inc.) and the secondary antibody was Alexa Fluor 488-conjugated
goat anti-mouse IgG (H+L; 1:100; cat. no. ab150113; Abcam). Cell
nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI) at
room temperature for 5 min (Bioworld Technology, Inc.). The cells
were incubated with antibodies as described above.
All samples were observed and images captured under
a confocal scanning laser microscope (FV1000; Olympus Corporation,
magnification, ×400); three randomly selected fields from each
sample were examined. Red and green staining indicated the positive
reaction. ImageJ software (version 1.49p; National Institutes of
Health) was used for analysis.
Human RPE cell culture, small
interfering RNA (siRNA) transfection and treatment
The human RPE cell line ARPE19 was obtained from the
Chinese Academy of Sciences cell bank. Cells were routinely
cultured in low-glucose Dulbecco's modified Eagle's medium (DMEM;
Invitrogen; Thermo Fisher Scientific, Inc.) containing 10% fetal
bovine serum (Zeta-Life Inc.) and 1% penicillin/streptomycin
(Sigma-Aldrich; Merck KGaA) in a humidified atmosphere of 5%
CO2 at 37°C. ARPE19 cells were used between passages 4
and 6 and a specific marker (RPE65) was detected using
immunofluorescence (25) (Fig. S1). Cells were fixed with 4% PFA at
4°C for 30 min and then blocked with PBS containing 1% BSA and 0.5%
Triton X-100 at room temperature for 1 h. Cells were incubated the
primary and secondary antibodies as above. Cell nuclei were stained
with DAPI at room temperature for 5 min (Bioworld Technology,
Inc.).
RPE cells were cultured to 60–70% confluence and
then transfected with a negative control (NC) siRNA or siCTGF
(SIGS0002293-1; RiboBio, Guangzhou, China) at a concentration of 50
nM using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
Target sequences for siCTGF were as follows: si-1,
GCACCAGCATGAAGACATA; si-2, GTGCATCCGTACTCCCAAA; si-3,
CTCCAAGCCTATCAAGTTT. The sequences of negative control siRNA are as
follows: Sense 5′-3′UUCUCCGAACGUGUCACGUTT, antisense
5′-3′ACGUGACACGUUCGGAGAATT.
RPE cells were treated with 15, 30 and 60 ng/ml
recombinant human CTGF (cat. no. CM22; Shanghai Novoprotein
Technology Co., Ltd.) or 30 ng/ml TGF-β1 (cat. no. CA59; Shanghai
Novoprotein Technology Co., Ltd.) for 24 h to perform reverse
transcription- quantitative (RT-q) PCR and western blotting. Based
on the results of a preliminary experiment and the reference
concentration, the most suitable concentration of TGF-β1 (30 ng/ml)
was chosen for this experiment (20). Cells were used for subsequent
experimentation 24 h following transfection.
RT-qPCR
RPE cells were cultured to 95% confluence. Total RNA
was purified using TRIzol® reagent (Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
Complementary DNA (cDNA) was synthesized using a reverse
transcription kit (Takara Bio, Inc.) according to the
manufacturer's protocols. RT-qPCR was performed in a 20-µl
reaction, containing the cDNA and specific primers, using a PCR kit
(SYBR Premix Ex Taq; Takara Bio, Inc.) according to the
manufacturer's instructions. The thermocycling conditions were as
follows: 98°C for 3 min, followed by 40 cycles at 98°C for 15 sec,
55°C for 15 sec and 72°C for 15 sec. Gene expression levels were
determined using the 2−ΔΔCq method (26) with β-actin used as an internal
reference. The primer sequences are shown in Table I. Each experiment was performed
three times.
 | Table I.Primer sequences used in the
study. |
Table I.
Primer sequences used in the
study.
| Gene | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| β-actin |
TGTTACCAACTGGGACGACA |
CTTTTCACGGTTGGCCTTAG |
| CTGF |
CAGCATGGACGTTCGTCTG |
AACCACGGTTTGGTCCTTGG |
| Fibronectin |
CGGTGGCTGTCAGTCAAAG |
AAACCTCGGCTTCCTCCATAA |
| ZO-1 |
CAACATACAGTGACGCTTCACA |
CACTATTGACGTTTCCCCACTC |
| N-cadherin |
CAGAATCGTGTCTCAGGCTCCAAG |
CTGCGTTCCAGGCTGGTGTATG |
| E-cadherin |
TACAATGCCGCCATCGCTTACAC |
TGACGGTGGCTGTGGAGGTG |
| Collagen type
III |
GGAGCTGGCTACTTCTCGC |
GGGAACATCCTCCTTCAACAG |
| α-SMA |
CTATGAGGGCTATGCCTTGCC |
GCTCAGCAGTAGTAACGAAGGA |
Western blotting
RPE cells were lysed with RIPA buffer (Beyotime
Institute of Biotechnology) containing a complete protease
inhibitor cocktail (Roche Molecular Biochemicals). A bicinchoninic
acid protein assay kit (Beyotime Institute of Biotechnology) was
used to determine protein concentrations. Proteins (30 µg) were
separated by 8–12% sodium dodecyl sulfate polyacrylamide gel
electrophoresis (Bio-Rad Laboratories, Inc.) and transferred to
polyvinylidene fluoride membranes (EMD Millipore). The membranes
were blocked with 5% skimmed milk [diluted with Tris-buffered
saline containing 0.05% Tween-20 (TBST)] at room temperature for 1
h, followed by incubation with primary antibodies in TBST at 4°C
overnight. After three washes with TBST for 5 min each, the
membranes were incubated with a secondary antibody for 1 h at room
temperature, followed by three washes. The membranes were scanned
using an enhanced chemiluminescence assay (Beijing 4A Biotech Co.,
Ltd.). ImageJ software (version 1.49p; National Institutes of
Health) was used to calculate the gray value and analysis of the
protein bands.
The following primary antibodies were used:
Fibronectin (1:1,000; cat. no. 26836; Cell Signaling Technology,
Inc.); ZO-1 (1:1,000; cat. no. 13663S; Cell Signaling Technology,
Inc.); N-cadherin (1:1,000; cat. no. 13116T; Cell Signaling
Technology, Inc.); E-cadherin (1:1,000; cat. no. 3195T; Cell
Signaling Technology, Inc.); collagen type III (1:1,000; cat. no.
22734-1-AP, ProteinTech Group, Inc.); α-smooth muscle actin (α-SMA;
1:1,000; cat. no. ab5694, Abcam); phosphorylated (p)-PI3K (1:1,000;
cat. no. 4228; Cell Signaling Technology, Inc.); PI3K (1:1,000;
cat. no. 4257; Cell Signaling Technology, Inc.); p-AKT (1:1,000;
cat. no. 4060; Cell Signaling Technology, Inc.); AKT (1:1,000; cat.
no. 4691; Cell Signaling Technology, Inc.); CTGF (1:1,000; cat. no.
ab5097, Abcam); GAPDH (1:1,000; cat. no. 10494-1-AP, ProteinTech
Group, Inc.). Horseradish peroxidase-conjugated goat anti-rabbit
IgG (1:3,000; cat. no. SA00001-2; ProteinTech Group, Inc.) was used
as the secondary antibody.
[3H]proline incorporation
by RPE cells
The [3H]proline incorporation assay was
employed as an indicator of the rate of collagen synthesis
(27). RPE cells
(5×104/ml) were seeded into 96-well plates and grown to
85% confluence. The medium was replaced with a serum-free medium
and incubation continued for 24 h before the cells were treated
with CTGF (5, 15, 30 and 60 ng/ml), 8-Bromoadenosine (8-Br) cAMP
(0.1 mM; Sigma-Aldrich; Merck KGaA) and 8-Br-cAMP (0.1 mM) + CTGF
(60 ng/ml). 8-Br-cAMP can inhibit the expression of CTGF (28). The CTGF-oligonucleotide-transfected
cells were treated with 15 ng/ml TGF-β1 for 24 h, followed by
incubation with 2 µCi of [3H]proline (China Institute of
Atomic Energy) for an additional 24 h. Subsequently, the cells were
washed three times with PBS for 5 min each and lysed with trypsin
(Sigma-Aldrich; Merck KGaA) at room temperature for 2 min. The
incorporation of [3H]proline was determined by
scintillation counting. Each experimental condition was performed
in triplicate wells and the experiments were repeated three times.
The results are expressed as cpm/103 cells ± standard
error of the mean (SEM) after counting trypsinized cell monolayers;
the tested reagents caused only minor changes in the cell numbers
of these growth-arrested cultures over a 2-day assay period.
CTGF-oligonucleotide ODN (CTGF-ODN)
transfection of RPE cells
Oligonucleotide (ODN) is a polymer consisting of a
small number of nucleotides (29).
The ODNs used to bind to CTGF included CTGF sense
oligonucleotide (SODN; 5′-ATGACCGCCGCCAGTA-3′) and antisense
oligonucleotide (ASODN, to inhibit the expression of CTGF;
5′-TACTGGCGGCGGTCAT-3′), as previously designed (30). The ODNs included a phosphorothioate
modification synthesized by Shanghai Shenggong Biotech Co., Ltd.
RT-qPCR was used to detect the knockdown efficiency of CTGF-ASODN
(Fig. S2). RPE cells
(5×104/ml) were seeded in 96-well plates and incubated
in a 5% CO2 incubator at 37°C for 2 days until they
almost reached 85% confluence. Prior to transfection, the medium
was replaced with a serum-free medium without antibiotics. For
transfection, each ODN was diluted with serum-free DMEM to a final
concentration of 400 nM and Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) was diluted 1:25 with
serum-free DMEM. The solution of each ODN was mixed with diluted
Lipofectamine® and incubated at room temperature for 20
min, followed by aliquoting 50 µl of the resulting complexes into
each well. Cells mixed with the ODN/Lipofectamine® 2000
were incubated in a 5% CO2 incubator at 37°C for 12 h.
Cells with >90% viability, as determined using trypan blue
exclusion, were maintained in culture. The cells that took up the
trypan blue were considered non-viable. Cells (1 ml) were stained
with 0.1 ml of trypan stock solution (Sigma-Aldrich; Merck KGaA).
Cells (10 µl) were added to a hemocytometer (Paul Marienfeld GmbH
& Co. KG) and examined immediately under a light microscope
(Eclipse E100, Nikon Corporation; magnification, ×10). The number
of blue staining cells and the number of total cells were counted.
The transfection medium was replaced with a serum-free medium
containing 100 µg/ml vitamin C (Tianjin Aoran Fine Chemical
Research Institute) and 15 ng/ml TGF-β1 and the cells were cultured
for another 24 h.
Statistical analysis
Data analysis was performed using GraphPad Prism 8.0
(GraphPad Software, Inc.). All quantitative results are presented
as the mean ± SEM. An unpaired Student's t-test was used to
determine the statistical significance of differences between the
two groups. To determine the statistical significance of the
differences among multiple groups, one-way analysis of variance
(ANOVA) with Dunnett's or Tukey's post hoc test was employed.
P<0.05 was considered to indicate a statistically significant
difference.
Results
CTGF induces EMT and the ECM
synthesis
Immunohistochemical staining was employed to detect
the expression of CTGF, fibronectin and collagen type III in PRM
sections. These proteins were detected in all PRMs (Fig. 1A). CTGF-positive cells were
scattered or clustered in PRMs and were found in fibrotic regions
of the membranes (Fig. 1A).
CTGF-positive cells were mainly epithelioid cells with a long
elliptic or polygonal phenotype and abundant cytoplasm.
Immunofluorescence co-staining with a specific marker of RPE cells
demonstrated that CTGF-positive cells were also RPE65 positive,
which confirmed that these cells were RPE cells (Fig. 1B). However, a smaller population of
CTGF-positive cells included fibroblast-like cells, which were
spindle shaped and large bodied.
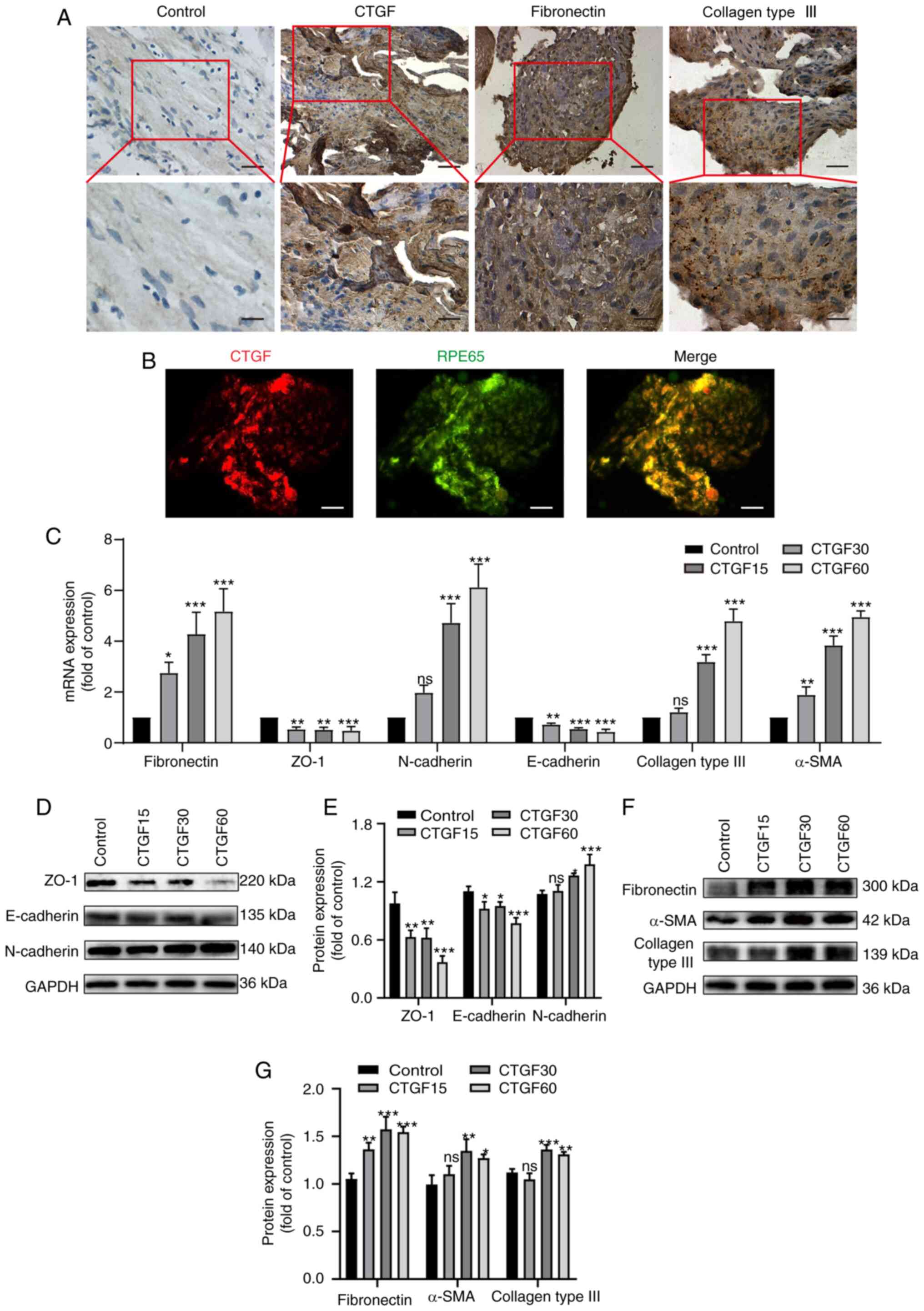 | Figure 1.Effects of CTGF on EMT and the ECM
synthesis. (A) Immunohistochemistry of surgically excised human PVR
specimens stained for CTGF, fibronectin and collagen III. Original
magnification, ×400 and ×800 (enlargement of boxed area). (B)
Immunofluorescence co-staining of PVR membranes for CTGF (red) and
RPE65 (green). Original magnification, ×400 (C). Relative mRNA
expression levels of biomarkers for EMT and the ECM synthesis in
ARPE19 cells treated with CTGF (15, 30 and 60 ng/ml), including
specific markers of epithelial cells (ZO-1 and E-cadherin),
mesenchymal cells (fibronectin, N-cadherin and α-SMA) and the ECM
(collagen type III). (D) Representative western blot analysis of
the expression levels of EMT markers in ARPE19 cells treated with
CTGF (15, 30 and 60 ng/ml). (E) Quantification of relative protein
expression of ZO-1, E-cadherin and N-cadherin. (F) Representative
western blot analysis of the expression levels of EMT and ECM
synthesis markers in ARPE19 cells treated with CTGF (15, 30 and 60
ng/ml). (G) Relative protein expression levels of fibronectin,
α-SMA and collagen type III; n=3. *P<0.05, **P<0.01,
***P<0.001 vs. control group; ns, not significant; CTGF,
connective tissue growth factor; EMT, epithelial-mesenchymal
transition; ECM, extracellular matrix; PVR, proliferative
vitreoretinopathy; RPE65, retinal pigment epithelium-specific 65
kDa protein; α-SMA, α-smooth muscle actin. |
Tables II and
III demonstrate that the intense
positive rate of CTGF expression in the grade B membranes (16.7%)
was lower compared with the grade C and D membranes (54.5 and
77.8%, respectively). ECM proteins, fibronectin and collagen type
III, were expressed in all PRMs and evidently expressed in CTGF
intense-positive PRMs. The positive rates of fibronectin expression
in the grade B and C membranes (66.7 and 36.4%, respectively) were
higher compared with the grade D membranes (22.2%). By contrast,
the positive rates of collagen type III expression in the grade B
and C membranes (33.3 and 54.5%, respectively) were significantly
lower compared with the grade D membranes (66.7%). These results
support the hypothesis that fibronectin is involved in scar
formation during the early phase of wound healing, while collagens
become the primary constituents of the ECM during the advanced
stages of PVR.
| Table II.CTGF, fibronectin and collagen type
III expression in the PRMs of PVR. |
Table II.
CTGF, fibronectin and collagen type
III expression in the PRMs of PVR.
| No. | Sex (M/F) | Age (year) | Duration of
symptoms (month) | PVR (C/D) | CTGF | FN | Collagen III |
|---|
| 1 | M | 42 | 12 | D2 | +++ | ++ | +++ |
| 2 | F | 29 | 6 | B | +++ | +++ | +++ |
| 3 | M | 67 | 14 | C2 | ++ | ++ | + |
| 4 | M | 39 | 18 | D2 | +++ | ++ | +++ |
| 5 | F | 52 | 10 | C1 | +++ | ++ | +++ |
| 6 | M | 27 | 20 | C3 | +++ | +++ | +++ |
| 7 | F | 24 | 15 | D2 | ++ | + | ++ |
| 8 | M | 38 | 12 | D1 | +++ | +++ | ++ |
| 9 | F | 49 | 5 | B | ++ | +++ | ++ |
| 10 | F | 33 | 4 | B | + | ++ | + |
| 11 | M | 19 | 9 | D1 | ++ | ++ | ++ |
| 12 | F | 55 | 15 | D1 | +++ | +++ | +++ |
| 13 | M | 48 | 12 | C1 | + | + | + |
| 14 | M | 34 | 8 | B | ++ | +++ | ++ |
| 15 | F | 25 | 9 | C1 | ++ | ++ | ++ |
| 16 | M | 36 | 24 | D3 | +++ | + | +++ |
| 17 | F | 58 | 18 | D1 | +++ | ++ | +++ |
| 18 | M | 28 | 12 | C3 | +++ | +++ | +++ |
| 19 | M | 26 | 10 | C2 | +++ | ++ | +++ |
| 20 | F | 30 | 6 | C2 | ++ | ++ | ++ |
| 21 | M | 22 | 14 | C3 | +++ | +++ | +++ |
| 22 | M | 21 | 4 | B | + | + | + |
| 23 | F | 36 | 9 | C2 | ++ | ++ | ++ |
| 24 | F | 39 | 15 | C3 | +++ | +++ | +++ |
| 25 | M | 43 | 18 | D2 | +++ | + | +++ |
| 26 | F | 49 | 7 | B | ++ | +++ | +++ |
| Table III.Expression of CTGF, fibronectin,
collagen type III protein in the PRMs of PVR. |
Table III.
Expression of CTGF, fibronectin,
collagen type III protein in the PRMs of PVR.
|
| PVR B | PVR C1-C3 | PVR D1-D3 |
|---|
|
|
|
|
|
|---|
| n (%) | + (%) | ++ (%) | +++ (%) | + (%) | ++ (%) | +++ (%) | + (%) | ++ (%) | +++ (%) |
|---|
| CTGF | 2 (33.3) | 3 (50) | 1 (16.7) | 1 (9.1) | 4 (36.4) | 6 (54.5) | 0 | 2 (22.2) | 7 (77.8) |
| Fibronectin | 1 (16.7) | 1 (16.7) | 4 (66.7) | 1 (9.1) | 6 (54.5) | 4 (36.4) | 3 (33.3) | 4 (44.4) | 2 (22.2) |
| Collagen III | 2 (33.3) | 2 (33.3) | 2 (33.3) | 2 (18.2) | 3 (27.3) | 6 (54.5) | 0 | 3 (33.3) | 6 (66.7) |
The present study further investigated whether CTGF
promotes EMT, as well as ECM synthesis, by treating RPE cells with
different concentrations of CTGF (15, 30 and 60 ng/ml) using
RT-qPCR and western blotting (Fig.
1C-G). The results demonstrated that the mRNA (Fig. 1C) and protein (Fig. 1D and E) expression levels of the
epithelial markers ZO-1 and E-cadherin decreased in a CTGF
concentration-dependent manner. In addition, the mRNA (Fig. 1C) and protein (Fig. 1D-G) expression levels of the
mesenchymal markers fibronectin, N-cadherin and α-SMA were
elevated. Furthermore, the expression of the ECM protein collagen
type III was upregulated with the increased concentrations of CTGF
(Fig. 1C, F and G). These results
indicated that CTGF may promote EMT and the ECM synthesis by RPE
cells.
Knocking down CTGF reverses EMT and
the ECM synthesis by RPE cells
To verify the effect of CTGF on EMT and the ECM
synthesis by RPE cells, the present study screened for a siCTGF
with the highest knockdown efficiency using RT-qPCR (Fig. 2A). Thereafter, RPE cells were
treated with an NC siRNA and siCTGF and the expression of EMT and
ECM biomarkers was determined using RT-qPCR (Fig. 2B) and western blotting (Fig. 2C-F).
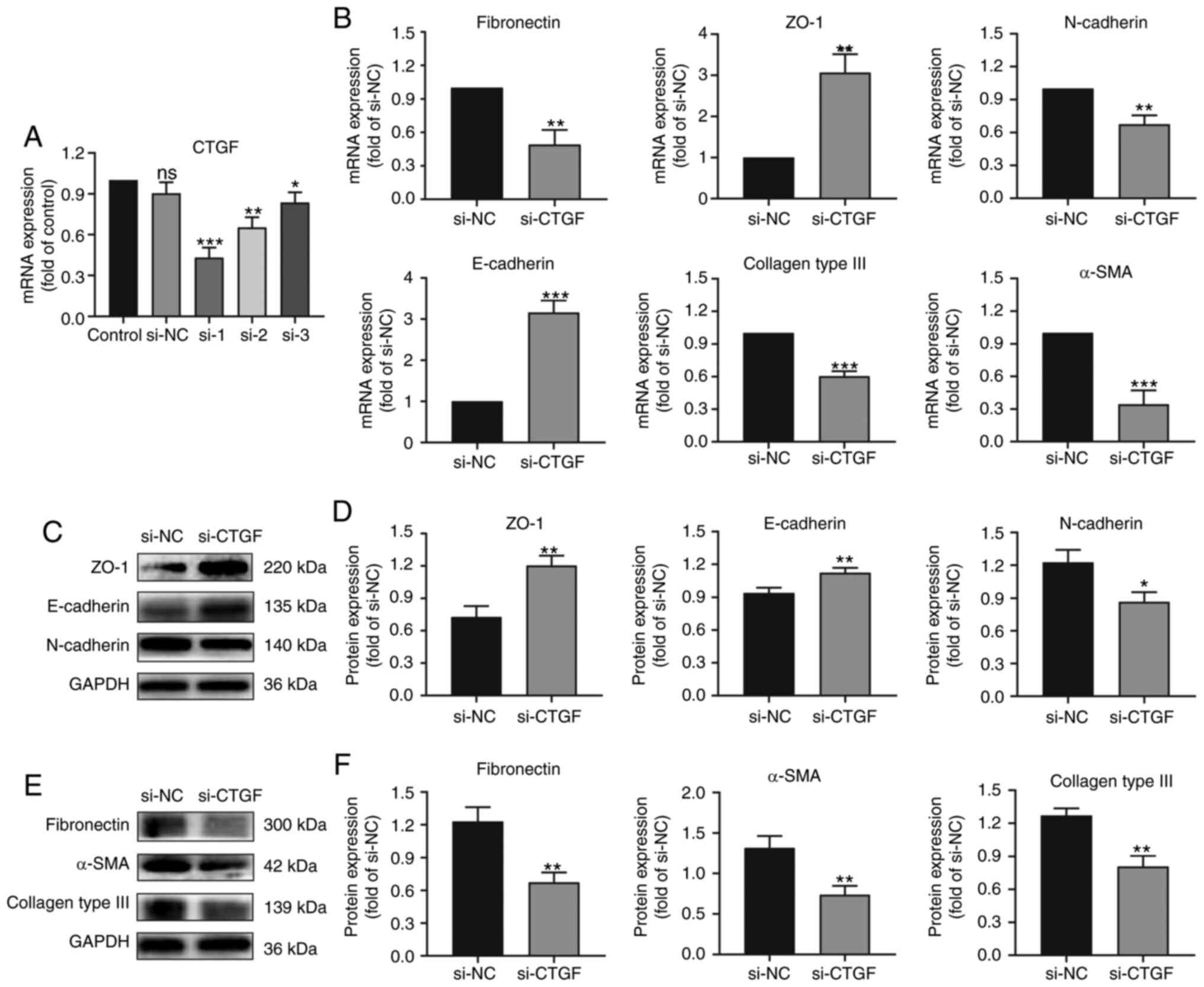 | Figure 2.Effects of CTGF knockdown on EMT and
the ECM synthesis by ARPE19 cells. (A) Screening for a siCTGF with
the highest knockdown efficiency by reverse
transcription-quantitative PCR. (B) Relative mRNA expression levels
of biomarkers for EMT and the ECM synthesis in ARPE19 cells
transfected with si-NC and siCTGF, including specific markers of
epithelial cells (ZO-1 and E-cadherin), mesenchymal cells
(fibronectin, N-cadherin and α-SMA) and the ECM (collagen type
III). (C) Representative western blot analysis of the expression
levels of EMT markers in ARPE19 cells transfected with si-NC and
siCTGF. (D) Quantification of relative expression levels of ZO-1,
E-cadherin and N-cadherin. (E) Representative western blot analysis
of the expression levels of EMT and ECM synthesis markers in ARPE19
cells transfected with si-NC and siCTGF. (F) Quantification of
relative expression levels of fibronectin, α-SMA and collagen type
III; n=3. *P<0.05, **P<0.01, ***P<0.001 vs. si-NC group;
ns, not significant; CTGF, connective tissue growth factor; EMT,
epithelial-mesenchymal transition; ECM, extracellular matrix;
siCTGF, CTGF-specific small interfering RNA (siRNA); si-NC,
negative control siRNA; α-SMA, α-smooth muscle actin. |
The results demonstrated that the expression of ZO-1
and E-cadherin (epithelial markers) increased after knocking down
CTGF compared with the NC group (Fig. 2B-D). By contrast, the expression of
fibronectin, N-cadherin and α-SMA (mesenchymal markers) decreased
after knocking down CTGF compared with the NC group
(Fig. 2B-F). Knockdown of
CTGF significantly downregulated the expression of collagen
type III compared with the NC group (Fig. 2B, E and F). These results indicate
that knocking down CTGF can reverse the EMT process and ECM
synthesis by RPE cells.
TGF-β1-stimulated synthesis of
collagen is partially mediated by CTGF
[3H]proline incorporation assay was used
to determine the effects of CTGF on collagen synthesis. The results
demonstrated that the incorporation of [3H]proline by
RPE cells increased in a CTGF concentration-dependent manner
(Fig. 3A). CTGF also significantly
enhanced the synthesis of collagen by RPE cells compared with that
by control cells. In combination with CTGF, 8-Br-cAMP inhibited the
CTGF-stimulated incorporation of [3H]proline by RPE
cells (Fig. 3B). Compared with CTGF
treatment (60 ng/ml), 8-Br-cAMP reduced the [3H]proline
incorporation, but no significant difference in collagen production
was observed between cells exposed to 8-Br-cAMP and control
cells.
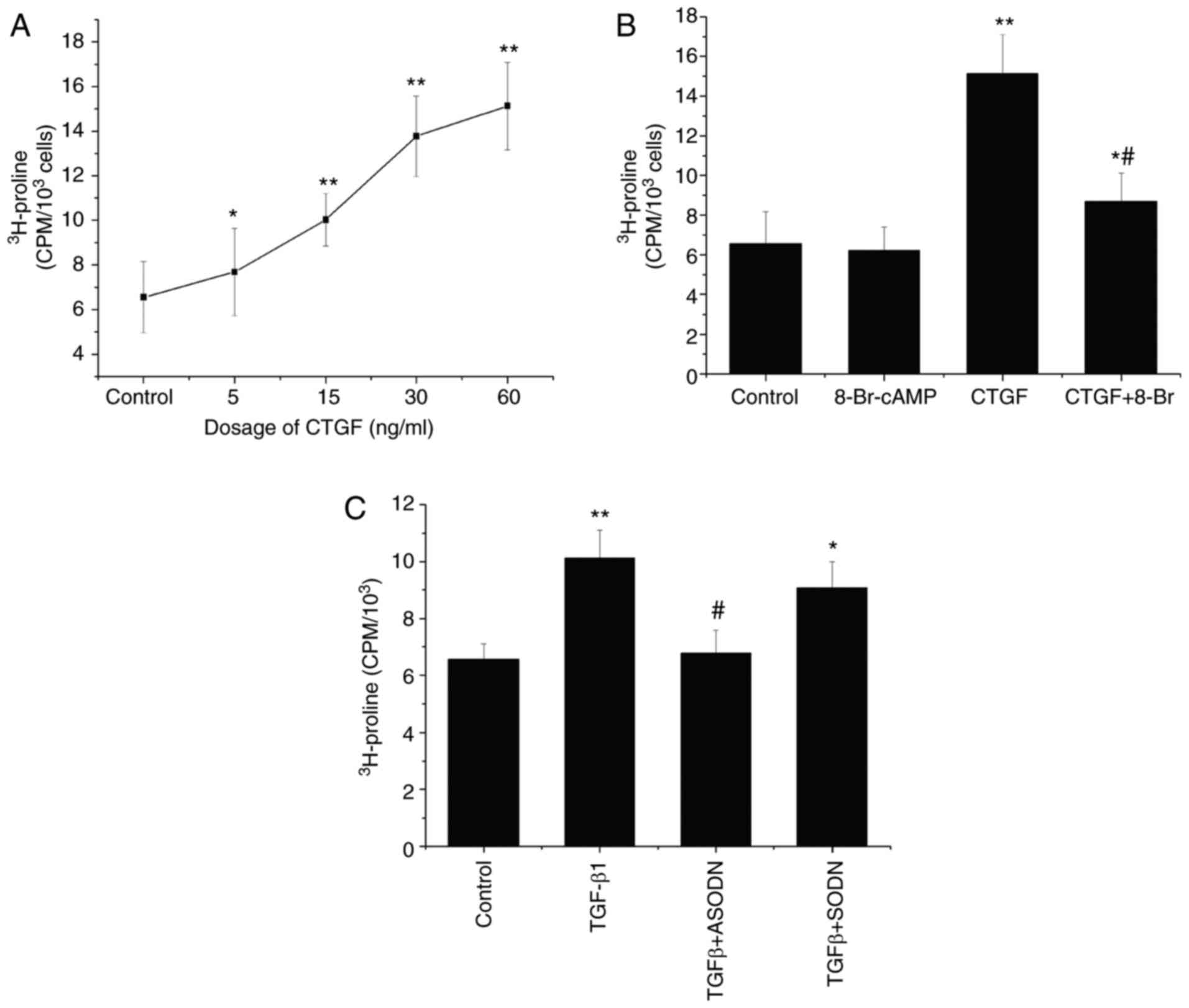 | Figure 3.Mediation of TGF-β1-stimulated
collagen synthesis by CTGF. (A) Incorporation of
[3H]proline by ARPE19 cells treated with CTGF (5, 15, 30
and 60 ng/ml). (B) Incorporation of [3H]proline by
untreated ARPE19 cells and cells treated with 8-Br-cAMP (0.1 mM),
CTGF (60 ng/ml) and CTGF (60 ng/ml) + 8-Br-cAMP (0.1 mM).
*P<0.05, **P<0.01 vs. control; #P<0.05 vs. the
CTGF group. (C) Incorporation of [3H]proline by
CTGF-ODN-transfected ARPE19 cells treated with 15 ng/ml TGF-β1. The
results are expressed as cpm/103 cells ± standard error
of the mean; n=3. *P<0.05, **P<0.01 vs. control;
#P<0.05 vs. the TGF-β1 group. CTGF, connective tissue
growth factor; 8-Br, 8-Bromoadenosine; ODN, oligonucleotide; S,
sense; A, antisense; cpm, counts per min. |
The incorporation of [3H]proline
significantly increased following treatment of RPE cells with 15
ng/ml TGF-β1 compared with control cells. By contrast, RPE cells
transfected with the CTGF antisense oligonucleotide and then
treated with TGF-β1 (30 ng/ml) exhibited a decrease in the rate of
[3H]proline incorporation, which indicated that the
CTGF antisense oligonucleotide could partially block the
TGF-β1-induced synthesis of collagen (Fig. 3C). By contrast, the CTGF
sense oligonucleotide demonstrated no such effect under the same
experimental conditions, resulting in no significant difference in
[3H]proline incorporation compared with the TGF-β1 group
(Fig. 3C). Taken together, these
results indicate that the TGF-β1-stimulated production of ECM
proteins is partially mediated by CTGF.
The PI3K/AKT pathway is regulated by
CTGF
To explore the potential pathway involved in the
CTGF-mediated EMT process and ECM synthesis by RPE cells, the
phosphorylation levels of PI3K and AKT in CTGF-treated RPE cells
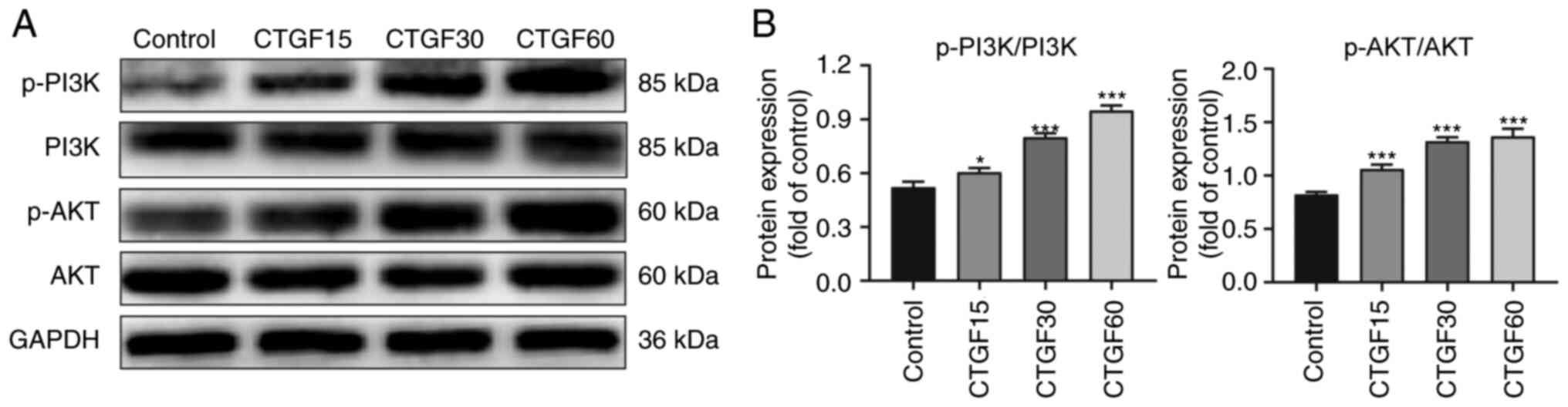
were determined using western blotting (Fig. 4). The results revealed marked
phosphorylation of PI3K and AKT in a CTGF concentration-dependent
manner, whereas the total PI3K and AKT levels did not significantly
change (Fig. 4). These data
suggested that the observed regulation of EMT and the ECM synthesis
by CTGF was associated with PI3K/AKT signaling in RPE cells.
TGF-β1 regulates CTGF expression and
the activation of the PI3K/AKT pathway
To study upstream factors involved in the regulation
by CTGF of EMT and ECM synthesis, RPE cells were treated with
TGF-β1 and the expression of CTGF and phosphorylation levels on the
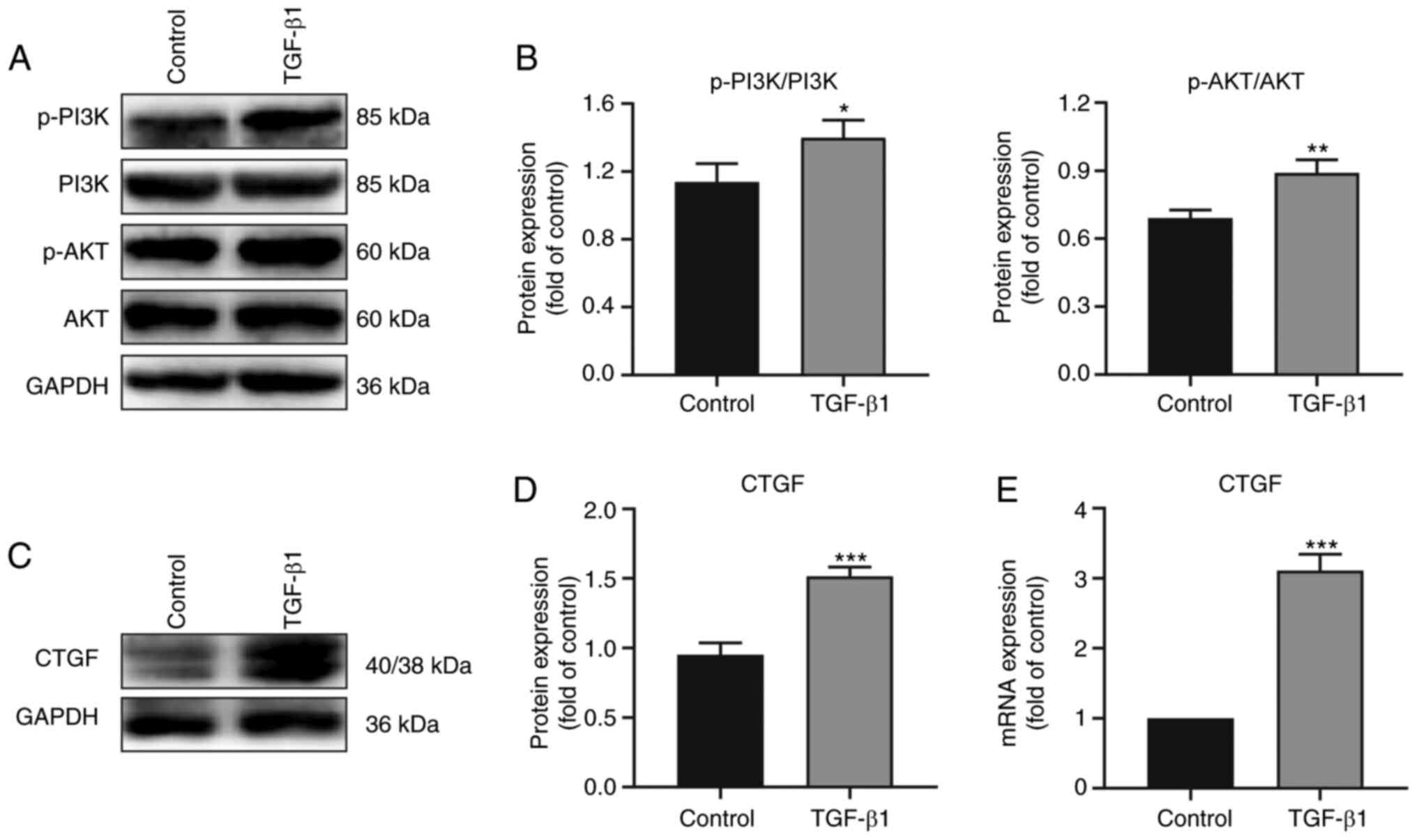
signaling pathway proteins determined by western blotting (Fig. 5). Notably, the expression of CTGF
and the phosphorylation of PI3K and AKT were higher in the TGF-β1
group compared with the control group (Fig. 5A-D). RT-qPCR analysis also
demonstrated that the mRNA expression of CTGF was higher in
the TGF-β1 group (Fig. 5E). Taken
together, the data indicated that TGF-β1 regulated CTGF expression
and the activation of the PI3K/AKT pathway, thereby further
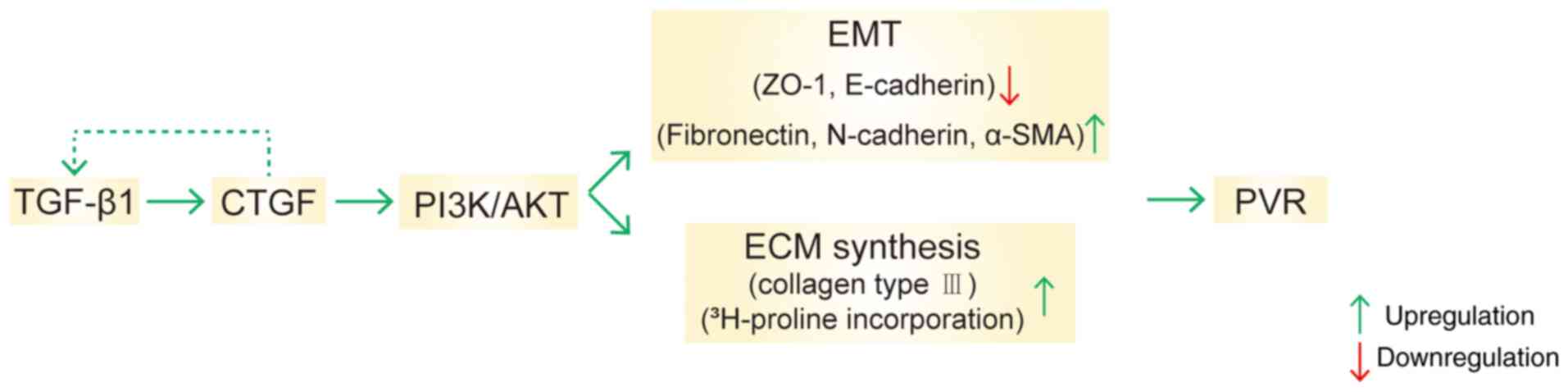
promoting EMT and the ECM synthesis in PVR (Fig. 6).
Discussion
PVR is a common disease, which occurs as a result of
a dysregulated healing process of an intraocular wound and is one
of the leading causes of blindness (1,14).
Prevention of the formation of contractile PRMs in the vitreous and
on the inner and outer surfaces of a detached retina contributes to
the clinical treatment of PVR (2).
CTGF is a member of a new immediate early gene family and
encodes a protein that has been classified into a group of
structurally related molecules, termed CCN (CTGF/Cyr61/Nov)
(31). CTGF is a growth factor that
has been demonstrated to stimulate the cell adhesion, chemotactic
activity and proliferation of multiple cell types; it also induces
angiogenesis and promotes EMT as well as the synthesis of ECM
components (2). CTGF is highly
expressed in fibrotic lesions of the lung and kidney, in the
cardiac muscle during atherosclerosis and in tumors (18,31,32).
In the present study, CTGF was found to be highly
expressed in PRMs. In addition, it was found that the expression of
fibronectin and collagen type III was upregulated compared with the
control group. The results supported the hypothesis that
fibronectin is involved in scar formation during the early phase of
wound healing, whereas collagens become the primary constituents of
the ECM in the advanced stages of PVR. Analysis of the cell
morphology and immunofluorescence staining demonstrated that
CTGF-expressing cells were RPE and fibroblast-like cells. In
previous studies, double staining of specimens demonstrate that
CTGF-positive cells are also positive for keratin expression,
indicating that the cells are derived from RPE cells (13,18).
Therefore, the present study hypothesized that CTGF-positive cells
are mainly RPE cells. Previous studies have demonstrated that RPE
cells can undergo changes in differentiation signaling and the
phenotype, acquiring resemblance to mesenchymal or fibroblast-like
cells (EMT), which are involved in the generation of a PVR lesion
(2,8,14).
Furthermore, the spindle cells positive for CTGF expression were
determined to be transformed RPE cells (33). CTGF immunoreactivity of RPE cells
has been demonstrated to increase from early- to late-stage PVR and
CTGF expression is the highest during late-stage PVR (13,18,34).
The results of the present study are consistent with these previous
results. It is hypothesized that at the later stage, the
inflammatory period of PVR development and upregulation of CTGF can
further aggravate RPE cell proliferation, adherence, EMT and
collagen secretion to form a PRM.
Previous studies have demonstrated that the EMT
process of RPE cells is a major pathological characteristic of PVR
(2,35). EMT refers to morphological changes
that involve the transformation of epithelial cells into
fibroblasts or mesenchymal cells and is associated with tissue
regeneration, damage repair and organ fibrosis (36,37).
The ability of fibrotic RPE cells to proliferate, migrate and
synthesize the ECM is significantly increased (1,2). The
present study found that CTGF was highly expressed in the
proliferative membranes of patients with PVR.
It has been reported that CTGF is closely associated
with the process of EMT and the synthesis of an ECM (2). However, whether CTGF induces EMT of
RPE cells has not been thoroughly investigated. There are two types
of molecular markers associated with the process of EMT; one
includes the markers of epithelial-like cells, including
E-cadherin, ZO-1 and β-catenin, and the other type includes
mesenchymal-like cell markers, including fibronectin, N-cadherin,
α-SMA and vimentin (36,38). The present study further
demonstrated that CTGF significantly decreased the expression of
epithelial markers, including ZO-1 and E-cadherin, and increased
that of mesenchymal markers, including fibronectin, N-cadherin and
α-SMA, in a concentration-dependent manner. Meanwhile, the
expression trends of the above markers were reversed after knocking
down CTGF.
In the microenvironment wherein cells survive, a
number of factors, including the ECM, hypoxia and macrophages,
serve a significant role in inducing EMT (36,39).
The ECM is not only a cytoskeleton for maintaining the structure
and function, but it also mediates processes including cell growth,
differentiation, migration and wound healing (40,41).
Studies of PRMs have demonstrated that the primary ECM constituents
are fibronectin and collagen subtypes I, III and IV, which are most
consistently associated with RPE and fibroblast-like cells
(42–44).
The present study also found that the expression of
the ECM protein collagen type III was upregulated in RPE cells. In
addition, the incorporation of [3H]proline by RPE cells
increased in a CTGF concentration-dependent manner, while 8-Br-cAMP
inhibited the CTGF-stimulated collagen synthesis. Proline residues
account for almost 21% of the total amino acid content of
collagens, whereas they are rarely found in other proteins
(27). Consequently,
[3H]proline incorporation by cells can be used as an
indirect measure of the rate of collagen synthesis (27). An increase in the synthesis of
collagen can reflect an increase in the cell proliferation
(45,46). Accordingly, the present study
suggested that CTGF may promote EMT and the ECM synthesis by RPE
cells, thus contributing to the formation and contraction of
proliferative membranes in PVR.
A number of studies have focused on the possible
signaling pathways of EMT (2,38,47).
The precise regulatory mechanism of the EMT process involves
extracellular signal transduction to the cell via specific binding
to a receptor on the membrane, leading to the initiation of a
variety of signaling pathways to activate transcription factors in
the nucleus and finally regulate the expression of EMT-related
genes (36,37). EMT involves a number of regulatory
signaling pathways, including PI3K/AKT, TGF-β, Wnt and Notch
pathways, and there is also a complex crosstalk between pathways
during the EMT process (37,38).
However, the signaling pathway by which CTGF regulates EMT of RPE
cells remains to be elucidated. The present study demonstrated that
the phosphorylation of PI3K and AKT significantly increased with
the increase of the CTGF concentration. Therefore, the data
suggested that the regulation of EMT and the ECM synthesis by CTGF
is associated with the PI3K/AKT signaling pathway in RPE cells.
Several studies have indicated that TGF-β is a
strong inducer of CTGF expression in tubular epithelial cells and
in a variety of other cells, including fibroblasts, mesangial
cells, endothelial cells and chondrocytes, and that CTGF serves an
important role in TGF-β signaling (2,17,18,20).
Previous studies have demonstrated that TGF-β and CTGF are highly
expressed in the vitreous humor and subretinal fluid of patients
with PVR and exert stimulatory effects on the contraction of gels
seeded with RPE cells (18,34,48,49).
TGF-β is also considered to be an important factor in the
phenotypic transformation of RPE cells and in the fibrosis of PRMs
(2,50). CTGF has been demonstrated to
stimulate the synthesis of collagen by mediating the TGF-β or
autocrine signaling pathways (17).
Transient transfection has been employed to express a CTGF
antisense oligonucleotide and block CTGF gene expression in
RPE cells (51). The CTGF
antisense oligonucleotide, but not the CTGF sense
oligonucleotide, was found to block an increase in the
TGF-β1-induced [3H]proline incorporation in the present
study. The results suggested that the TGF-β1-stimulated production
of ECM proteins is partially mediated by CTGF. In addition, the
present study found that the expression of CTGF and the
phosphorylation of PI3K and AKT were higher in the TGF-β1 group
compared with the control group. It has been proposed that TGF-β
induces EMT of RPE cells via classical, Smad-dependent and
non-classical, Smad-independent, signaling (2,52).
Gressner et al (53)
demonstrated that CTGF acts as a Smad2-dependent sensitizer of
TGF-B actions in hepatocytes, not a direct activator of Smad2. Our
unpublished data indicated that CTGF can induce the phosphorylation
of Smad2/3, which needs further exploration.
There are some limitations to the present study. The
potential role of microRNAs in preventing PVR by regulating EMT in
RPE cells has been extensively studied and this has been helpful in
developing new biological markers and therapeutic targets (6,54,55).
The present study only found some indirect mechanisms; thus, it is
necessary to further explore novel direct mechanisms of CTGF
regulation of PVR, including the regulation of CTGF by
microRNAs.
In summary, the present study demonstrated that CTGF
promoted EMT and the ECM synthesis in PVR via the PI3K/AKT
signaling pathway. Its findings suggested that the inhibition of
CTGF expression may represent a possible target for the prevention
of an excessive wound-healing response by RPE cells, thereby
preventing PVR. Further detailed research on the role of CTGF in
PVR, along with the latest mechanism including microRNAs and more
in vivo studies, will enable the use of CTGF in future
clinical applications to alleviate the sufferings of patients with
PVR.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Professor Chaowei
Tian (Department of Ophthalmology, Xijing Hospital, Fourth Military
Medical University, Xi'an, Shaanxi, China) for his technical
assistance in collecting PVR membranes.
Funding
This project was supported by the National Natural
Science Foundation of China (grant nos. 81470655, 81570856,
81670863, 81770936 and 81970814).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
CG, YH and YW conceived and designed the study. YW,
TC, TW and HD performed experiments. YW, TC, TW, WY and GD analyzed
the data and wrote the manuscript. CG and YW reviewed and edited
the manuscript. GD, YW, and YH supervised the project, provided
thoughtful comments and suggestions during this study. YW and TC
confirmed the authenticity of all the raw data. All authors have
read and approved the final manuscript.
Ethics approval and consent to
participate
All donors provided written informed consent prior
to tissue donation. The study was performed in accordance with the
tenets of the Declaration of Helsinki and the protocol was approved
by the institutional research ethics committee of the Fourth
Military Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
α-SMA
|
α-smooth muscle actin
|
|
BSA
|
bovine serum albumin
|
|
cDNA
|
complementary DNA
|
|
CTGF
|
connective tissue growth factor
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
ECM
|
extracellular matrix
|
|
EMT
|
epithelial-mesenchymal transition
|
|
NC
|
negative control
|
|
PBS
|
phosphate-buffered saline
|
|
PFA
|
polyformaldehyde
|
|
PRM
|
preretinal membrane
|
|
PVR
|
proliferative vitreoretinopathy
|
|
RPE
|
retinal pigment epithelium
|
|
RT-qPCR
|
reverse transcription-quantitative
PCR
|
|
SEM
|
standard error of the mean
|
|
siRNA
|
small interfering RNA
|
|
TBST
|
Tris-buffered saline with
Tween-20
|
References
|
1
|
Tosi GM, Marigliani D, Romeo N and Toti P:
Disease pathways in proliferative vitreoretinopathy: An ongoing
challenge. J Cell Physiol. 229:1577–1583. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang S, Li H, Li M and Wang F: Mechanisms
of epithelial-mesenchymal transition in proliferative
vitreoretinopathy. Discov Med. 20:207–217. 2015.PubMed/NCBI
|
|
3
|
Campochiaro PA: Pathogenic mechanisms in
proliferative vitreoretinopathy. Arch Ophthalmol. 115:237–241.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Abu El-Asrar AM, Van den Steen PE, Al-Amro
SA, Missotten L, Opdenakker G and Geboes K: Expression of
angiogenic and fibrogenic factors in proliferative vitreoretinal
disorders. Int Ophthalmol. 27:11–22. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sethi CS, Lewis GP, Fisher SK, Leitner WP,
Mann DL, Luthert PJ and Charteris DG: Glial remodeling and neural
plasticity in human retinal detachment with proliferative
vitreoretinopathy. Invest Ophthalmol Vis Sci. 46:329–342. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kaneko H and Terasaki H: Biological
involvement of MicroRNAs in proliferative vitreoretinopathy. Transl
Vis Sci Technol. 6:52017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pastor JC: Proliferative
vitreoretinopathy: An overview. Surv Ophthalmol. 43:3–18. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee SC, Kwon OW, Seong GJ, Kim SH, Ahn JE
and Kay ED: Epitheliomesenchymal transdifferentiation of cultured
RPE cells. Ophthalmic Res. 33:80–86. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dvashi Z, Goldberg M, Adir O, Shapira M
and Pollack A: TGF-β1 induced transdifferentiation of rpe cells is
mediated by TAK1. PLoS One. 10:e01222292015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li M, Li H, Liu X, Xu D and Wang F:
MicroRNA-29b regulates TGF-β1-mediated epithelial-mesenchymal
transition of retinal pigment epithelial cells by targeting AKT2.
Exp Cell Res. 345:115–124. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pennock S and Kazlauskas A: Vascular
endothelial growth factor A competitively inhibits platelet-derived
growth factor (PDGF)-dependent activation of PDGF receptor and
subsequent signaling events and cellular responses. Mol Cell Biol.
32:1955–1966. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Umazume K, Barak Y, McDonald K, Liu L,
Kaplan HJ and Tamiya S: Proliferative vitreoretinopathy in the
Swine-a new model. Invest Ophthalmol Vis Sci. 53:4910–4916. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cui JZ, Chiu A, Maberley D, Ma P, Samad A
and Matsubara JA: Stage specificity of novel growth factor
expression during development of proliferative vitreoretinopathy.
Eye (Lond). 21:200–208. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ciprian D: The pathogeny of proliferative
vitreoretinopathy. Rom J Ophthalmol. 59:88–92. 2015.PubMed/NCBI
|
|
15
|
Platania CBM, Fisichella V, Fidilio A,
Geraci F, Lazzara F, Leggio GM, Salomone S, Drago F, Pignatello R,
Caraci F and Bucolo C: Topical ocular delivery of TGF-β1 to the
back of the eye: Implications in age-related neurodegenerative
diseases. Int J Mol Sci. 18:20762017. View Article : Google Scholar
|
|
16
|
Fisichella V, Giurdanella G, Platania CB,
Romano GL, Leggio GM, Salomone S, Drago F, Caraci F and Bucolo C:
TGF-β1 prevents rat retinal insult induced by amyloid-β (1–42)
oligomers. Eur J Pharmacol. 787:72–77. 2016. View Article : Google Scholar : PubMed/NCBIPubMed/NCBI
|
|
17
|
Zhu J, Nguyen D, Ouyang H, Zhang XH, Chen
XM and Zhang K: Inhibition of RhoA/Rho-kinase pathway suppresses
the expression of extracellular matrix induced by CTGF or TGF-beta
in ARPE-19. Int J Ophthalmol. 6:8–14. 2013.PubMed/NCBI
|
|
18
|
He S, Chen Y, Khankan R, Barron E, Burton
R, Zhu D, Ryan SJ, Oliver N and Hinton DR: Connective tissue growth
factor as a mediator of intraocular fibrosis. Invest Ophthalmol Vis
Sci. 49:4078–4088. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guo CM, Wang YS, Hu D, Han QH, Wang JB,
Hou X and Hui YN: Modulation of migration and Ca2+
signaling in retinal pigment epithelium cells by recombinant human
CTGF. Curr Eye Res. 34:852–862. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Khankan R, Oliver N, He S, Ryan SJ and
Hinton DR: Regulation of fibronectin-EDA through CTGF
domain-specific interactions with TGFbeta2 and its receptor
TGFβRII. Invest Ophthalmol Vis Sci. 52:5068–5078. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bradham DM, Igarashi A, Potter RL and
Grotendorst GR: Connective tissue growth factor: A cysteine-rich
mitogen secreted by human vascular endothelial cells is related to
the SRC-induced immediate early gene product CEF-10. J Cell Biol.
114:1285–1294. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Blom IE, Goldschmeding R and Leask A: Gene
regulation of connective tissue growth factor: New targets for
antifibrotic therapy? Matrix Biol. 21:473–482. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Moussad EE and Brigstock DR: Connective
tissue growth factor: What's in a name? Mol Genet Metab.
71:276–292. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hilton G, Machemer R, Michels R, Okun E,
Schepens C and Schwartz A: The classification of retinal detachment
with proliferative vitreoretinopathy. Ophthalmology. 90:121–125.
1983. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dunn KC, Aotaki-Keen AE, Putkey FR and
Hjelmeland LM: ARPE-19, a human retinal pigment epithelial cell
line with differentiated properties. Exp Eye Res. 62:155–169. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Duncan MR, Frazier KS, Abramson S,
Williams S, Klapper H, Huang X and Grotendorst GR: Connective
tissue growth factor mediates transforming growth factor
beta-induced collagen synthesis: Down-regulation by cAMP. FASEB J.
13:1774–1786. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kothapalli D, Hayashi N and Grotendorst
GR: Inhibition of TGF-beta-stimulated CTGF gene expression and
anchorage-independent growth by cAMP identifies a CTGF-dependent
restriction point in the cell cycle. FASEB J. 12:1151–1161. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Scherer LJ and Rossi JJ: Approaches for
the sequence-specific knockdown of mRNA. Nat Biotechnol.
21:1457–1465. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hishikawa K, Oemar BS, Tanner FC, Nakaki
T, Lüscher TF and Fujii T: Connective tissue growth factor induces
apoptosis in human breast cancer cell line MCF-7. J Biol Chem.
274:37461–37466. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Daniels JT, Schultz GS, Blalock TD,
Garrett Q, Grotendorst GR, Dean NM and Khaw PT: Mediation of
transforming growth factor-beta(1)-stimulated matrix contraction by
fibroblasts: A role for connective tissue growth factor in
contractile scarring. Am J Pathol. 163:2043–2052. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Davis JT and Foster WJ: Substrate
stiffness influences the time dependence of CTGF protein expression
in muller cells. Int Physiol J. 1:12018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Meyer P, Wunderlich K, Kain HL, Prunte C
and Flammer J: Human connective tissue growth factor mRNA
expression of epiretinal and subretinal fibrovascular membranes: A
report of three cases. Ophthalmologica. 216:284–291. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hinton DR, He S, Jin ML, Barron E and Ryan
SJ: Novel growth factors involved in the pathogenesis of
proliferative vitreoretinopathy. Eye (Lond). 16:422–428. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Takayama K, Kaneko H, Hwang SJ, Ye F,
Higuchi A, Tsunekawa T, Matsuura T, Iwase T, Asami T, Ito Y, et al:
Increased ocular levels of microRNA-148a in cases of retinal
detachment promote epithelial-mesenchymal transition. Invest
Ophthalmol Vis Sci. 57:2699–2705. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tam WL and Weinberg RA: The epigenetics of
epithelial-mesenchymal plasticity in cancer. Nat Med. 19:1438–1449.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gonzalez DM and Medici D: Signaling
mechanisms of the epithelial-mesenchymal transition. Sci Signal.
7:re82014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Theocharis AD, Manou D and Karamanos NK:
The extracellular matrix as a multitasking player in disease. FEBS
J. 286:2830–2869. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Polanco TO, Xylas J and Lantis JC II:
Extracellular matrices (ECM) for tissue repair. Surg Technol Int.
28:43–57. 2016.PubMed/NCBI
|
|
42
|
Miller CG, Budoff G, Prenner JL and
Schwarzbauer JE: Minireview: Fibronectin in retinal disease. Exp
Biol Med (Maywood). 242:1–7. 2017.
|
|
43
|
Morino I, Hiscott P, McKechnie N and
Grierson I: Variation in epiretinal membrane components with
clinical duration of the proliferative tissue. Br J Ophthalmol.
74:393–399. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Priglinger SG, Alge CS, Kreutzer TC,
Neubauer AS, Haritoglou C, Kampik A and Welge-Luessen U:
Keratinocyte transglutaminase in proliferative vitreoretinopathy.
Invest Ophthalmol Vis Sci. 47:4990–4997. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kita T, Hata Y, Miura M, Kawahara S, Nakao
S and Ishibashi T: Functional characteristics of connective tissue
growth factor on vitreoretinal cells. Diabetes. 56:1421–1428. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhao X, Zhang LK, Zhang CY, Zeng XJ, Yan
H, Jin HF, Tang CS and Du JB: Regulatory effect of hydrogen sulfide
on vascular collagen content in spontaneously hypertensive rats.
Hypertens Res. 31:1619–1630. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jia P, Hu Y, Li G, Sun Y, Zhao J, Fu J, Lu
C and Liu B: Roles of the ERK1/2 and PI3K/PKB signaling pathways in
regulating the expression of extracellular matrix genes in rat
pulmonary artery smooth muscle cells. Acta Cir Bras. 32:350–358.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chen XF, Du M, Wang XH and Yan H: Effect
of etanercept on post-traumatic proliferative vitreoretinopathy.
Int J Ophthalmol. 12:731–738. 2019.PubMed/NCBI
|
|
49
|
Abu El-Asrar AM, Imtiaz Nawaz M, Kangave
D, Siddiquei MM and Geboes K: Osteopontin and other regulators of
angiogenesis and fibrogenesis in the vitreous from patients with
proliferative vitreoretinal disorders. Mediators Inflamm.
2012:4930432012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Gamulescu MA, Chen Y, He S, Spee C, Jin M,
Ryan SJ and Hinton DR: Transforming growth factor beta2-induced
myofibroblastic differentiation of human retinal pigment epithelial
cells: Regulation by extracellular matrix proteins and hepatocyte
growth factor. Exp Eye Res. 83:212–222. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yamanaka O, Saika S, Ikeda K, Miyazaki K,
Kitano A and Ohnishi Y: Connective tissue growth factor modulates
extracellular matrix production in human subconjunctival
fibroblasts and their proliferation and migration in vitro. Jpn J
Ophthalmol. 52:8–15. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Derynck R and Budi EH: Specificity,
versatility, and control of TGF-β family signaling. Sci Signal.
12:eaav51832019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Gressner OA, Lahme B, Siluschek M, Rehbein
K, Weiskirchen R and Gressner AM: Connective tissue growth factor
is a Smad2 regulated amplifier of transforming growth factor beta
actions in hepatocytes-but without modulating bone morphogenetic
protein 7 signaling. Hepatology. 49:2021–2030. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Usui-Ouchi A, Ouchi Y, Kiyokawa M, Sakuma
T, Ito R and Ebihara N: Upregulation of Mir-21 levels in the
vitreous humor is associated with development of proliferative
vitreoretinal disease. PLoS One. 11:e01580432016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chen X, Ye S, Xiao W, Luo L and Liu Y:
Differentially expressed microRNAs in TGFβ2-induced
epithelial-mesenchymal transition in retinal pigment epithelium
cells. Int J Mol Med. 33:1195–1200. 2014. View Article : Google Scholar : PubMed/NCBI
|