Introduction
Osteoarthritis (OA) is caused by the imbalance of
the decomposition and anabolic metabolism among chondrocytes,
extracellular matrix (ECM) and subchondral bone. Moreover, it is
manifested as pain and inflammation in the affected joints, and in
severe cases, leads to restricted or even disabled joint activity
(1,2). OA is a common disease worldwide, and
with the aging of the population, the incidence of OA increases
year by year (3,4). OA not only harms the health of
patients and affects their quality of life, it also causes a heavy
economic burden on families and society (5).
The main pathological feature of OA is the
significantly increased apoptotic rate of articular chondrocytes
(6). Apoptosis is a type of
programmed cell death, which is initiated under the action of in
vivo or in vitro factors, such as hypoxia and nutrient
deprivation, eventually leading to increased expression levels of
apoptosis-related proteins and the occurrence of apoptosis
(7).
Chondrocytes, the only cell component existing in
articular cartilage, are mainly responsible for the synthesis and
renewal of ECM, but also serve an irreplaceable role in maintaining
the normal structure and physiological function of articular
cartilage (8). Since normal
chondrocytes are a necessary for maintaining the stability of ECM,
chondrocyte apoptosis is a key factor in the degeneration of
articular cartilage in OA, and the proportion of apoptosis is
highly consistent with the destruction of articular cartilage
(9). Therefore, it is particularly
important to promote chondrocyte proliferation and inhibit its
apoptosis, which is a key link in alleviating OA.
Our previous research has confirmed that Zhuanggu
huoxue Decoction can promote the anabolism of cartilage matrix in
rabbits with OA, including increasing the content of glucuronic
acid in cartilage, reducing the moisture content of cartilage and
preventing and slowing the degeneration of cartilage in OA
(10), indicating that Zhuanggu
huoxue Decoction has great application value in OA. However, due to
the multi-component, multi-target and integrated characteristics of
traditional Chinese medicine (TCM), its effective components and
specific mechanisms are unknown, which limits its wide application
to some extent (11). Therefore,
the present study will further analyze the possible effective
components of Zhuanggu huoxue Decoction and clarify its
mechanisms.
Zhuanggu huoxue Decoction is a clinical prescription
for the treatment of OA using Pingle Village Guo's bone setting,
which is composed of Radix rehmanniae preparata, Chinese
yam, Achyranthes bidentata, Eucommia ulmoides, Asiatic
cornelian cherry fruit, Salvia miltiorrhiza, Eupolyphaga
sinensis and Carthami flos, amongst other ingredients
(12). Among these factors,
cryptotanshinone (CTS) is the most effective water-soluble
component in the pharmacological effect of Salva
miltiorrhiza, which has been shown to have anti-inflammatory,
anti-tumor and cell activity protective effects (13,14).
CTS was confirmed to effectively inhibit IL-1β-induced secretion of
inflammatory factors (IL-6 and IL-8) in OA synovial cells and
reduce MMP-1 expression (15).
Moreover, CTS has been reported to alleviate the development of OA
in mice by reducing the expression levels of MMP-13 and
aggrecanases (such as ADAMTS like 5) that can induce chondrocyte
degradation (15). CTS has also
demonstrated protective effect on chondrocytes, but the specific
targets remain unknown, which is also the focus of the present
study.
Materials and methods
Bioinformatics analysis
The microarray GSE93008 dataset (16) was downloaded from the Gene
Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/pubmed). This dataset
comprised three normal controls and three osteoarthritis samples.
The relevant genes in the samples were analyzed using GEO2R
(http://www.ncbi.nlm.nih.gov/geo/geo2r) and R language
program (4.0.3). The screening conditions were adjusted P<0.05
and |logFC|≥1.
OA mouse model
A total of 12 male C57BL/6 mice (age, 12 weeks;
weight, 20–25 g) were provided by Junke Biological Co., Ltd., and
housed in ZK-SLJ mouse cages (Henan Zhike Hongrun Environmental
Protection Technology Co., Ltd.) at 20°C with 50% humidity and a
12-h light/dark cycle. The animals had unlimited access to a
standard rodent chow and water, and adapted for ≥7 days before the
experiment. The mice were randomly divided into three groups of 12
mice each: i) Sham group, the mice were injected with 20 µl 0.9%
normal saline; ii) OA model group, 20 µl 0.9% normal saline
containing 0.6 mg monosodium iodoacetate (MIA; AIKE Regent) was
injected into the knee joint and after 14 days, the mice were
gavaged with 0.5% sodium carboxymethyl cellulose; and iii) OA + CTS
group, CTS (MedChemExpress) was dissolved in 0.5% sodium
carboxymethyl cellulose (Sangon Biotech Co., Ltd.) to form an oral
suspension and then gavaged using a syringe after 14 days of MIA
injection (10 mg/kg/day) until 1 day before death. After performing
the Von Frey test, the mice were sacrificed by cervical
dislocation, and then the knee tissues were collected. The knee
tissues of 6 mice were stained, and the expression levels of
related genes in the other 6 mouse knee tissues were detected via
western blotting and reverse transcription-quantitative (RT-q) PCR,
amongst other techniques. All animal procedures were approved by
Luoyang Orthopedic-Traumatological Hospital of Henan Province
(Henan Provincial Orthopedic Hospital) and complied with the
Institutional Animal Care and Use Committee.
Cell cultures and treatments
Isolation and culture of chondrocytes was performed
as described previously (17). The
knee joints were collected from male C57BL/6 mice (age, 4 days;
weight, <12 g; Junke Biological Co., Ltd.; housing conditions as
previously described), cleaned with sterile PBS at 4°C and cut into
pieces. After digestion with 0.25% trypsin (Thermo Fisher
Scientific, Inc.) in an incubator at 37°C for 30 min, the knee
joints were incubated with 0.2% collagenase type II (Thermo Fisher
Scientific, Inc.) at 37°C for another 24 h. The residual
collagenase was removed with PBS. Next, 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.), 100 U/ml penicillin and 100 mg/ml
streptomycin were added into DMEM/Nutrient Mixture F-12 Ham
(Merck-KGaA), and then cultured at 37°C with 5% CO2
after inoculation. The chondrocytes used in this study were
passaged twice.
Chondrocytes were obtained and divided into four
groups according to different treatments. In the IL-1β group,
chondrocytes were treated with 0.5 ng/ml IL-1β (Sigma-Aldrich;
Merck KGaA) at 37°C for 24 h to simulate arthritis (18). In the IL-1β + CTS group,
chondrocytes were treated with 0.5 ng/ml IL-1β for 24 h and 10 µM
CTS for 8 h, both at 37°C (18–20).
In the IL-1β + CTS + microRNA (miRNA/miR)-574-5p mimic and IL-1β +
CTS + miR-574-5p mimic + pcDNA-YAF2 groups, chondrocytes were
treated with 0.5 ng/ml IL-1β for 24 h and 10 µM CTS for 8 h at
37°C. After that, cells were plated into 6-well plates and cultured
to 70–80% confluence, and then transfected with 100 pmol
miR-574-5p-mimic (Thermo Fisher Scientific, Inc.), 100 pmol NC
mimic and/or 1 µg/µl pcDNA-YY1-associated factor 2 (YAF2) at 37°C
for 48 h using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
After 48 h of cell transfection, the cells were collected for
subsequent experimentation. In the IL-1β + CTS +
5-aza-2′-deoxycytidine (5-aza-CdR; Sigma-Aldrich; Merck KGaA)
group, chondrocytes were treated with 0.5 ng/ml IL-1β for 24 h, 10
µM CTS for 8 h and 5 µM 5-aza-CdR (methylation inhibitor;
Sigma-Aldrich; Merck KGaA) for 24 h at 37°C.
The sequences of the mimics were as follows:
Mimic-negative control (NC), 5′-UGGGUUUGUGUGUGUGAGUGUGU-3′; and
miR-574-5p mimics, 5′-UGAGUGUGUGUGUGUGAGUGUGU-3′. Mimic-NC and
miR-574-5p mimics were synthesized by Shanghai GenePharma Co., Ltd.
The full-length sequence of YAF2 cDNA was amplified via PCR and
cloned into the pcDNA3.1 vector (Invitrogen; Thermo Fisher
Scientific, Inc.) to generate pcDNA-YAF2 constructs by Ke Lei
Biotechnology Co., Ltd. An empty vector was used as the
corresponding control.
293T cells were purchased from the Shanghai Cell
Bank of the Chinese Academy of Sciences. 293T cells were cultured
in high glucose-DMEM containing 10% (v/v) FBS and 1%
penicillin/streptomycin (Thermo Fisher Scientific, Inc.). The cells
were cultured at 37°C with 5% CO2.
RT-qPCR
Total RNAs from each sample were extracted using
TRIzol® reagent (Vigorous Biotechnology), and then
reverse transcribed using a cDNA Synthesis kit (Takara
Biotechnology Co., Ltd.) at 37°C for 15 min and 85°C for 5 sec. A
qPCR assay was then used to validate the expression levels of
miR-574-5p, miR-468-3p, miR-32-3p and miR-672-5p and YAF2, which
was performed on the ABI 7500 Fast Real-Time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The RT-qPCR reaction
was conducted in 20 µl reaction volumes containing cDNA, primers
and SYBR Green Real-time PCR Master mix (Shanghai Yeasen
Biotechnology Co., Ltd.). The following thermocycling conditions
were used: Initial denaturation at 95°C for 5 min; followed by 40
cycles at 95°C for 10 sec, 60°C for 30 sec and 72°C for 30 sec. The
2−ΔΔCq method was used to access the relative RNA
expression levels (21,22). GAPDH and U6 were used as internal
references, respectively. The primer sequences are listed in
Table I.
 | Table I.Primer sequences for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences for reverse
transcription-quantitative PCR.
| Gene | Primer sequence
(5′→3′) |
|---|
| YY1-associated
factor 2 | F:
TCGGATGAGGGTTACTGGGACTG |
|
| R:
AATCTTGGCCTGGTTTTCTTATGG |
| miR-574-5p | F:
CGCGTGAGTGTGTGTGTGTGA |
|
| R:
AGTGCAGGGTCCGAGGTATT |
| miR-468-3p | F:
UAUGACUGAUGUGCGUGUGUCUG |
|
| R:
GACUGAUGUACUGAUAAGAAACUCAGU |
| miR-32-3p | F:
TTTCTCTATCGATAGGTACCGGCAGTTACCATTTCACAC |
|
| R:
CACGCCGAATCAACATCAGTCTGATAA |
| miR-672-5p | F:
UGAGGUUGGUGUACUGUGUGUGA |
|
| R:
ACACACAGUCGCCAUCUUCGA |
| GAPDH | F:
TGCCCCCATGTTCGTCA |
|
| R:
TTGGCCAGGGGTGCTAAG |
| U6 | F:
CTCGCTTCGGCAGCACA |
|
| R:
AACGCTTCACGAATTTGCGT |
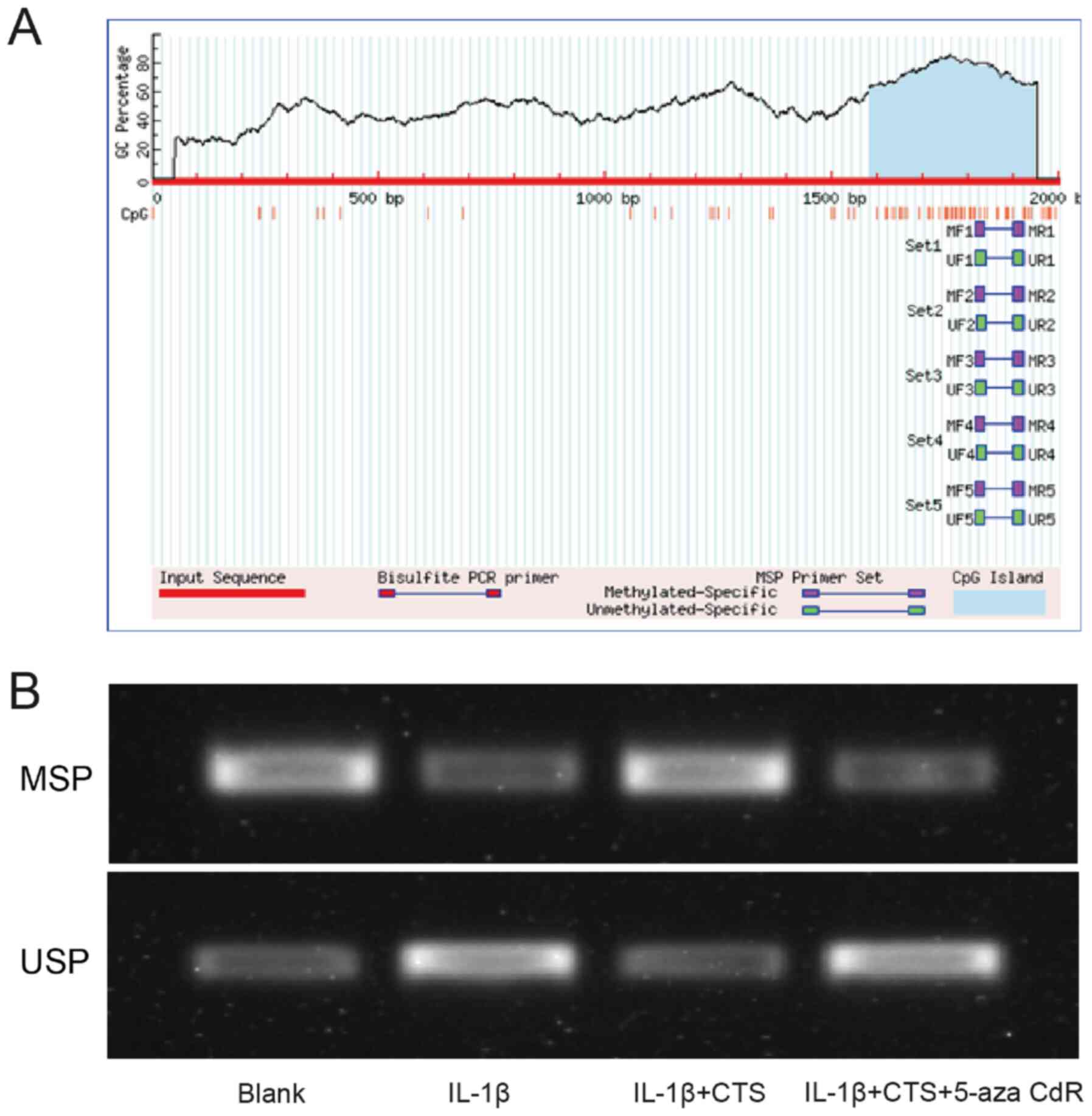
Methylation specific PCR (MSPCR)
The UCSC database (https://genome.ucsc.edu) was used to predict the
promoter sequence of miR-574-5p, and the upstream 2 kB region of
the promoter was extracted. MethPrimer (http://www.urogene.org/methprimer), an online analysis
site, was used to analyze and predict the CpG islands of miR-574-5p
promoters. The methylation status of miR-574-5p was detected via
MSPCR. For MSPCR, total genomic DNA was isolated from chondrocytes
in the different groups using a Genomic DNA Mini kit (Thermo Fisher
Scientific, Inc.) and then 1 µg DNA was treated with bisulfate
using the EZ DNA Methylation kit (Zymo Research Corp.) according to
the manufacturer's instructions. Specific primers in the form of
unmethylated or methylated were used: Methylated forward,
5′-TTAATTTAGAATCGGGAAATTTTAC-3′ and reverse,
5′-CACTACACCCTAACCTACTACGAA-3; and unmethylated forward,
5′-GAGATCCAAAACTATCAGTCTTGAATGGC-3 and reverse,
5′-GTGTCTGCTCACAGCAGTGAAACCCTAA-3′. The MSPCR products were
electrophoresed on 2% SYBR™ safe stained agarose gel.
Western blotting
The expression levels of YAF2, Bcl-2 and Bax were
analyzed using western blot analysis. Cells were rinsed in
phosphate buffer, and then dissociated using RIPA lysis buffer (EMD
Millipore). The concentrations of protein extracts were determined
using the Bradford method. An equal amount of protein (80 µg) was
electrophoresed via 10% SDS-PAGE, and then transferred to PVDF
membranes (EMD Millipore). PVDF membranes were blocked in 5%
non-fat milk at room temperature for 1 h, and then incubated with
primary antibodies (anti-YAF2, 1:10,000, cat. no. ab177945;
anti-Bax, 1:1,000, cat. no. ab32503; anti-Bcl-2, 1:1,000, cat. no.
ab182858; all Abcam) overnight at 4°C. Next, all membranes were
incubated with HRP-labeled secondary antibody (1:2,000; cat. no.
ab6721; Abcam) at room temperature for 1 h. Immunoblots were
visualized using an ECL kit (Beyotime Institute of Biotechnology)
according to the manufacturer's instructions, and then the
chemiluminescent signals of protein bands were recorded. GAPDH was
used as an internal control.
Von Frey test
The Von Frey test was used to measure the mechanical
nociceptive threshold of mice at 0, 7, 14 and 21 days (19). Mice were placed on the test bench
with a mesh bottom and adapted for 30 min. Then, the hind paw
plantar of each mouse was vertically stimulated with calibrated von
Frey cilia (0.02–1.4 g; Stoelting Co.) to ensure that the cilia
continued to flex for ≥2 sec during stimulation. Each mouse was
stimulated with cilia of the same intensity for five times, and the
behavior of the mice was recorded. The occurrence of related
behavioral reactions (foot shrinking, leg swinging, licking, leg
lifting, etc.) were recorded as positive reactions, and the minimum
stimulus intensity of three positive reactions was recorded as the
paw withdrawal threshold (PWT) of the mouse.
H&E and Safranin O (SO
staining)
The cartilages of the knee joints of the mice in
each group were removed after surgery and rinsed with pre-cooled
PBS. After fixation with 4% paraformaldehyde at room temperature
for 48 h, routine paraffin embedding was performed and 5-µm
sections were created. A H&E Staining kit (Beyotime Institute
of Biotechnology) was used for H&E and SO staining. In H&E
staining, the sections were stained with hematoxylin dye solution
for 5 min, placed in 1% hydrochloric acid ethanol differentiation
solution for a few sec and rinsed with water. The sections were
then stained with eosin solution for 3 min at room temperature and
washed with water. The sections were subsequently dehydrated in
gradient alcohol (70, 80, 90 and 100%) for 5 min and washed with
xylene I and II for 5 min. For SO staining, after treatment with 1%
hydrochloric acid ethanol differentiation solution at room
temperature for 15 sec, sections were soaked in 1% dilute ammonia
for 5 sec. After washing with water, sections were stained with
Fast Green Stain for 3 min and quickly rinsed with 1% acetic acid
for 10 sec. Sections were stained with eosin for 3 min at room
temperature and the following steps were the same as the H&E
staining. Cartilages of knee joints in different groups of mice
were observed with a CKX41 phase-contrast microscope
(magnifications, ×200 and ×400; Olympus Corporation).
Alcian Blue staining
Chondrocytes at the logarithmic growth stage were
inoculated into 6-well plates at a density of 1×105
cells/well, and cultured in an incubator of 5% CO2 at
37°C. When the cells reached 90% fusion, they were washed once with
PBS, fixed with paraformaldehyde for 10 min at room temperature,
stained with 1% Alcian blue for 30 min at 20°C, and then washed
once with 0.1% HCl solution. The Alcian Blue Stain kit
(Sigma-Aldrich; Merck KGaA) was used for Alcian Blue staining.
Cells were visualized using a light microscope (magnification,
×1,000; YS110; Nikon Corporation).
TUNEL analysis
TUNEL staining was used to determine the apoptotic
status of chondrocytes in different groups, and the apoptotic
nuclei were stained brown. TUNEL staining was performed using TUNEL
Apoptosis Detection kit IV (FITC+POD; Wuhan Boster Biological
Technology, Ltd.) according to the manufacturer's instructions.
Briefly, cells were fixed with 4% paraformaldehyde for 30 min at
room temperature. Before adding TUNEL working solution for 1 h at
37°C, DNase I was used to digest the cells for 30 min to prepare
positive cell controls. After adding 3,3′-diaminobenzidine for 15
min at room temperature, the slides were mounted with neutral
balsam mounting media (Sangon Biotech Co., Ltd.) and the apoptotic
nuclei showed a yellow-brown color. Apoptotic cells were visualized
using a CKX41 phase-contrast microscope (magnifications, ×200 and
×400; Olympus Corporation).
Flow cytometry assay
The chondrocytes in the logarithmic growth phase
were digested with 0.25% trypsin (Thermo Fisher Scientific, Inc.)
and inoculated into 6-well cell culture plates at a density of
2×106 cells/well. The Annexin V FITC APOPTOSIS Detection
kit I 100TST (BD Pharmingen; BD Biosciences) was used according to
the manufacturer's instructions. Briefly, 5 µl Annexin V-FITC and
10 µl propidium iodide (PI) was added, and cells were incubated at
room temperature for 15 min in dark. A CytoFLEX flow cytometer
(Beckman Coulter, Inc.) was used to detect cell fluorescence
immediately after staining with PI solution at room temperature.
FlowJo Software 10 (FlowJo LLC) was used for data analysis. Early
and late apoptotic cells were counted to determine the levels of
cardiomyocyte apoptosis.
Dual-luciferase assay
First, the binding sites of miR-574-5p and YAF2 were
predicted via the miRDB database (http://www.mirdb.org), and the binding sites were
amplified by PCR. Then, the binding sites were inserted into the
pmirGLO reporter vector (Promega Corporation) to construct the YAF2
wild-type (WT) plasmid (Luc-WT-YAF2). The YAF2 mutant (MUT) plasmid
(Luc-MUT-YAF2) was constructed by mutating nucleotides using gene
mutation technology. After cells were plated into 6-well plates and
cultured to 70–80% confluence, they were co-transfected with 0.25
µg Luc-WT-YAF2, 0.25 µg Luc-MUT-YAF2 and 0.25 µg miR-574-5P-mimic
or 0.25 µg mimic-NC into 293T cells at 37°C for 24 h using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). The Luciferase Assay kit (Invitrogen; Thermo
Fisher Scientific, Inc.) was used to detect the luciferase activity
of each group. Normalized relative luciferase activity=Firefly
luciferase activity/Renilla luciferase activity.
Cell proliferation detection
Cell proliferation detection was conducted according
to the Cell Counting Kit-8 (CCK-8) instructions (MedChemExpress).
Chondrocytes at the logarithmic growth stage were inoculated into
96-well plates (1×105 cells/well) and incubated in an
incubator (37°C; 5% CO2) for 48 h. Next, 100 µl CCK-8
solution (10% concentration) was added for further culture for 1 h
at 37°C. The absorbance value (optical density value) at 450 nm was
determined using a microplate reader. All the experiments were
repeated three times.
Statistical analysis
SPSS 20.0 software (IBM Corp.) was used for
statistical analysis of data collected from ≥3 independent
experiments. The measurement data are presented as the mean ± SD.
An unpaired Student's t-test was used to analyze the difference
between the two groups. One-way ANOVA and Tukey's post hoc test
were used to analyze the data between multiple groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
Expression levels of miR-574-5p, YAF2,
Bax and Bcl-2 in the mouse model of OA
Differential expression analysis based on the
microarray data set GSE93008 (Sham vs. OA, n=3 vs. n=3) of OA mice
was performed. Taking the adjusted P-value <0.05 and |logFC|≥1
as screening conditions, four conservative miRNAs, namely
miR-574-5p, miR-468-3p, miR-32-3p and miR-672-5p, were identified
(data not shown). RT-qPCR was used to detect the expression levels
of four conserved miRNAs in the Sham group and OA group, and the
results demonstrated that miR-574-5p had the most significant
changes, indicating that miR-574-5p may be an important regulatory
factor affecting OA (Fig. 1A).
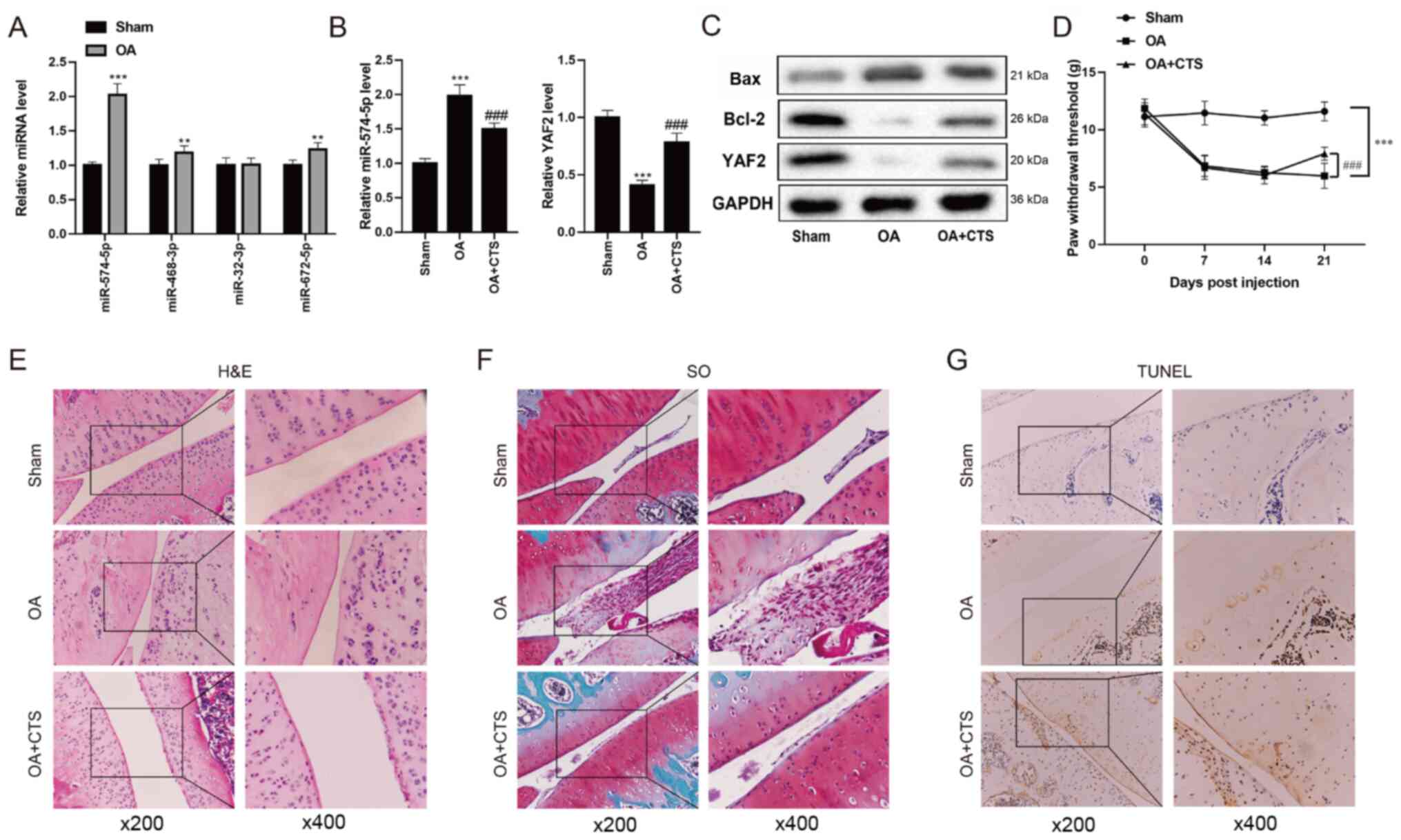 | Figure 1.Expression levels of miR-574-5p,
YAF2, Bax and Bcl-2 in the mouse model of OA. (A) Differential
expression analysis based on the microarray data set GSE93008 (Sham
vs. OA, n=3 vs. n=3) of OA mice was performed. RT-qPCR was used to
detect the expression levels of four conserved miRNAs in the Sham
group and OA group. (B) RT-qPCR was performed to detect the
expression levels of miR-574-5p and YAF2 in the Sham group, OA
group and OA + CTS group. (C) Western blot analysis was used to
detect the protein expression levels of YAF2, Bax and Bcl-2 in OA
mice. (D) Von Frey test was used to measure mechanical nociceptive
threshold of mice at 0, 7, 14 and 21 days. (E) H&E and (F) SO
staining were performed to reveal the morphology of mouse cartilage
between Sham group, OA group and OA + CTS group. (G) Chondrocyte
apoptosis was detected using a TUNEL assay. Magnifications, ×200
and ×400. **P<0.01, ***P<0.001 vs. Sham group;
###P<0.001 vs. OA group. OA, osteoarthritis; CTS,
Cryptotanshinone; miRNA/miR, microRNA; YAF2, YY1-associated factor
2; SO, Safranin O; RT-qPCR, reverse transcription-quantitative
PCR. |
Yang et al (23) reported that YAF2 has a protective
effect on chondrocytes by inhibiting cell apoptosis. Therefore,
RT-qPCR was performed to detect the expression levels of miR-574-5p
and YAF2 in the Sham group, OA group and OA + CTS group. The
results demonstrated that the expression level of miR-574-5P was
the highest in the OA group, and was downregulated in the OA + CTS
group (Fig. 1B). However, compared
with the Sham group, YAF2 expression in the OA group was
significantly decreased, while YAF2 expression in the OA + CTS
group was increased (Fig. 1B and
C). In addition, the expression levels of Bcl-2 and Bax, which
inhibit and promote cell apoptosis (24), respectively, were measured. The
western blotting results indicated that the protein expression
level of Bcl-2 in OA mice was markedly downregulated, but was
increased after the gavage of CTS suspension, while the protein
expression level of Bax exhibited the opposite effects (Fig. 1C).
Effect of CTS on chondrocyte apoptosis
in vivo
To investigate the effect of OA on pain sensation in
mice, the present study compared the differences in PWT between
Sham group, OA group and OA + CTS group. The bilateral PWT in the
OA group was detected using a Von Frey test, and the results
demonstrated that there was no significant change between the Sham
group and the OA-contralateral group at day 21, while the PWT in
the OA-ipsilateral group was markedly decreased compared with the
Sham and OA-contralateral groups (Fig.
S1). Thus, this study only compared the unilateral PWT between
the Sham group, OA group and OA + CTS group. As presented in
Fig. 1D, when compared with the
Sham group, the PWT of mice in the OA group at day 21 was
significantly reduced, and the PWT was significantly increased at
day 21 in the OA + CTS group compared with the OA group, indicating
that CTS could relieve the pain of mice with OA.
Next, HE and SO staining were conducted to reveal
the morphology of mouse cartilage between the Sham group, OA group
and OA + CTS group. In the Sham group, it was found that the
cartilage remained intact and the chondrocytes (blue) were ordered
neatly, while the cartilage was thinner and the number of
chondrocytes was decreased in the OA group (Fig. 1E). After the addition of CTS, the
apoptosis of chondrocytes was alleviated (Fig. 1E). SO staining of the cytoplasmic
matrix in the Sham group was normal, while the degree of the
cytoplasmic matrix in the OA group was increased and was recovered
in the OA + CTS group (Fig. 1F). In
addition, TUNEL staining results identified that there was no
positive nuclear staining in the Sham group, and that the number of
apoptotic cells in the OA group was markedly increased, while the
number of apoptotic cells was significantly decreased after adding
CTS (Fig. 1G).
Expression levels of miR-574-5p, YAF2,
Bax and Bcl-2 in chondrocytes
Next, it was verified whether the expression levels
of miR-574-5p, YAF2, Bax and Bcl-2 in chondrocytes were consistent
with those in the mouse model of OA. The successful differentiation
of chondrocytes was first determined using Alcian Blue staining
(Fig. 2A), and then the
chondrocytes were divided into control group, IL-1β group and IL-1β
+ CTS group after different treatments. The differential expression
levels of miR-574-5p and YAF2 in chondrocytes were detected via
RT-qPCR, and the results indicated that the expression level of
miR-574-5p in the IL-1β group was increased, but was decreased in
the IL-1β + CTS group (Fig. 2B). By
contrast, YAF2 mRNA and protein expression levels were both
decreased in the IL-1β group compared with control group, and were
increased after the addition of CTS (Fig. 2B and C).
 | Figure 2.Expression levels of miR-574-5p,
YAF2, Bax and Bcl-2 in chondrocytes. Chondrocytes were divided into
Con group, IL-1β group and IL-1β + CTS group after different
treatments. (A) Alcian Blue staining was used to detect the
differentiation of chondrocytes. Magnification, ×1,000 (B)
Differential expression levels of miR-574-5p and YAF2 in
chondrocytes were detected via reverse transcription-quantitative
PCR. (C) Western blot analysis was used to detect the protein
expression levels of YAF2, Bax and Bcl-2 in chondrocytes. (D)
Apoptotic rate of chondrocytes was detected via flow cytometry, and
(E) the results were quantified. **P<0.01, ***P<0.001 vs.
control group; #P<0.05, ##P<0.01,
###P<0.001 vs. IL-1β group. Con, control; CTS,
Cryptotanshinone; miR, microRNA; YAF2, YY1-associated factor 2. |
Western blot analysis revealed that, compared with
control group, the protein level of Bax in the IL-1β group was
markedly increased, while the expression of Bcl-2 was decreased.
After adding CTS, the protein expression level of Bax was
downregulated, while the expression level of Bcl-2 was upregulated
(Fig. 2C).
As shown in Fig. 2D and
E, the apoptotic rate of chondrocytes in the IL-1β group
detected by flow cytometry was higher compared with that in the
control group, while the apoptotic rate in the IL-1β + CTS group
was decreased. All these findings suggested that the expression
levels of miR-574-5p and the pro-apoptotic gene Bax were
upregulated, while the expression levels of YAF2 and the
anti-apoptotic gene Bcl-2 were downregulated in arthritic
chondrocytes, indicating that CTS could alleviate the apoptosis of
chondrocytes.
CTS affects chondrocyte apoptosis by
regulating the expression of miR-574-5p and then interfering with
YAF2
Since the biogenic analysis found that miR-574-5p
may target binding to YAF2 (Fig.
3A), a dual-luciferase reporter assay was performed to
determine whether miR-574-5p could interact with the YAF2 promoter.
The 293T cells were co-transfected with the YAF2
promoter-luciferase reporter plasmid and mimic-NC or miR-574-5p
mimic. As presented in Fig. 3B, in
the Luc-WT-YAF2 group, the luciferase activity of the miR-574-5p
mimic was significantly decreased compared with that in 293T cells
transfected with mimic-NC, indicating that miR-574-5p had a
regulating effect on YAF2. The mRNA expression levels of YAF2 were
validated in Fig. S2.
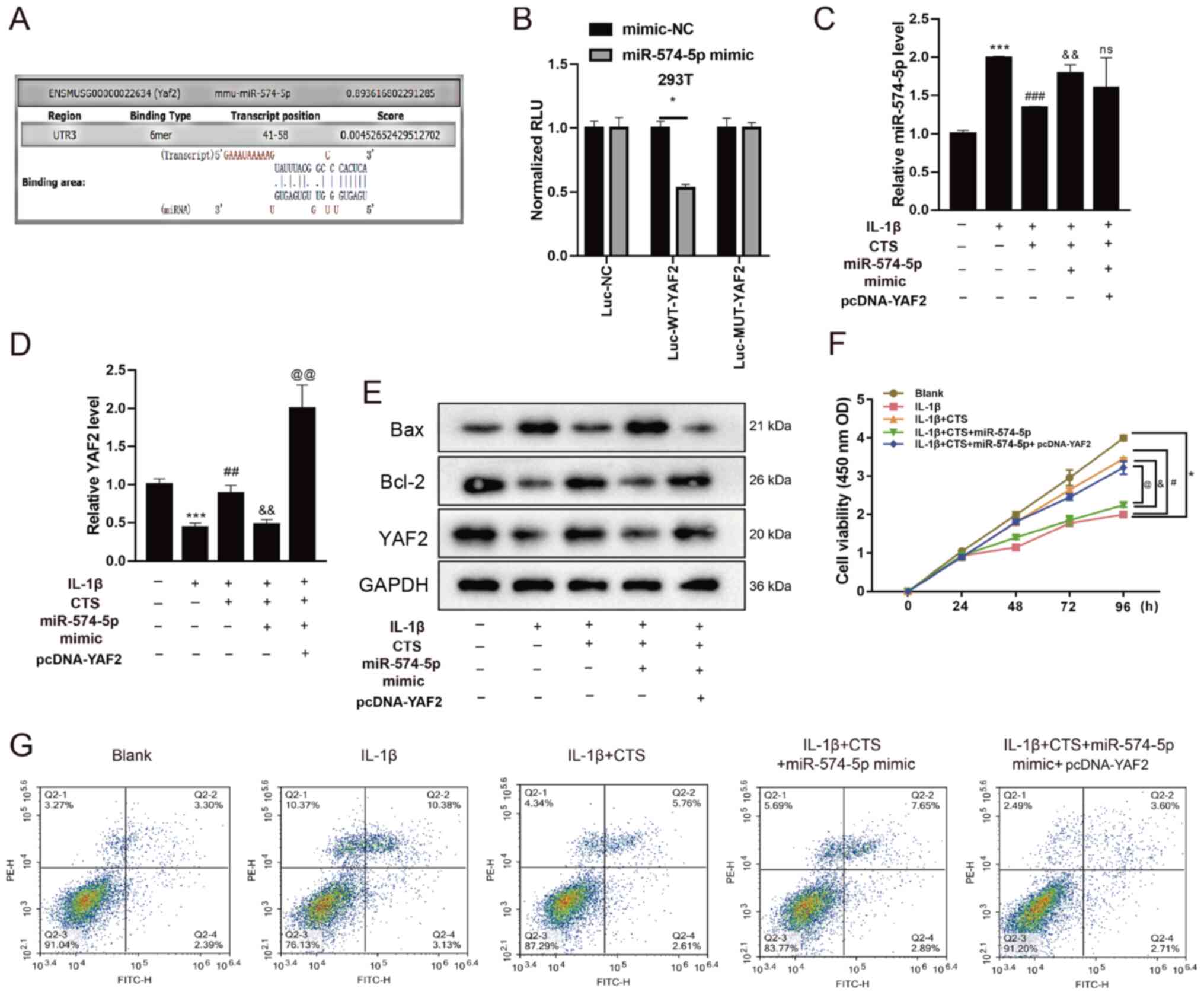 | Figure 3.CTS affects chondrocyte apoptosis by
regulating the expression of miR-574-5p and then interfering with
YAF2. (A) Biogenic analysis and (B) dual-luciferase reporter assay
was preformed to verify the interaction between miR-574-5p and
YAF2. Chondrocytes were divided into Blank group, IL-1β group,
IL-1β + CTS group, IL-1β + CTS + miR-574-5p-mimic group and IL-1β +
CTS + miR-574-5p-mimic + pcDNA-YAF2 group. *P<0.05. Differential
expression levels of (C) miR-574-5p and (D) YAF2 in chondrocytes
were detected via reverse transcription-quantitative PCR. (E)
Western blot analysis was used to detect the protein expression
levels of YAF2, Bax and Bcl-2 in chondrocytes. Cell proliferation
and apoptosis of chondrocytes were detected using a (F) Cell
Counting Kit-8 assay and (G) flow cytometry. *P<0.05,
***P<0.001 vs. Blank group; #P<0.05,
##P<0.01, ###P<0.001 vs. IL-1β group;
&P<0.05, &&P<0.01 vs. IL-1β
+ CTS group; @P<0.05, @@P<0.001 vs.
IL-1β + CTS + miR-574-5p-mimic group. CTS, Cryptotanshinone; miR,
microRNA; YAF2, YY1-associated factor 2; OD, optical density; Luc,
luciferase; NC, negative control; RLU, relative luciferase
activity; WT, wild-type; MUT, mutant. |
After treatment with IL-1β and CTS, chondrocytes
were transfected with miR-574-5p-mimics and then co-transfected
with pcDNA-YAF2. The mRNA expression levels of miR-574-5p and YAF2
after cell transfection with miR-574-5p mimic and pcDNA-YAF2 were
detected via RT-qPCR (Fig. S3).
The expression level of miR-574-5p in arthritic chondrocytes with
addition of IL-1β was increased, and then decreased after the
addition of CTS (Fig. 3C),
indicating that CTS could reduce the expression of miR-574-5p in
arthritic chondrocytes. After chondrocytes treated with IL-1β and
CTS, and then transfected with miR-574-5p-mimics, miR-574-5p
expression was found to be upregulated (Fig. 3C). In addition, there was no
significant change in the expression level of miR-574-5p after
overexpressing YAF2 (Fig. 3C).
Opposite results were found for the mRNA and protein expression
levels of YAF2, indicating that overexpression of miR-574-5p could
decrease the expression of YAF2 (Fig.
3D and E). Transfection with pcDNA-YAF2 reversed the effect of
miR-574-5p mimic, as YAF2 expression was significantly increased
(Fig. 3D and E).
Bax expression was upregulated in arthritic
chondrocytes and was downregulated after the addition of CTS, which
was upregulated after IL-1β + CTS-treated cells were transfected
with miR-574-5p-mimic. Moreover, after transfection with
pcDNA-YAF2, the expression of Bax was decreased (Fig. 3E). By contrast, the protein
expression level of Bcl-2 showed an opposite trend, indicating that
miR-574-5p could increase Bax expression and decrease Bcl-2
expression (Fig. 3E).
Next, a CCK-8 assay was performed to detect the
effects of CTS, miR-574-5p and YAF2 on chondrocytes proliferation.
The results demonstrated that the proliferative ability of
chondrocytes in the IL-1β group was significantly decreased, but
this was restored after the addition of CTS. However, the
proliferative ability of chondrocytes after transfection with
miR-574-5p-mimic was decreased to a level similar to that of the
IL-1β group (Fig. 3F). Furthermore,
transfection with pcDNA-YAF2 could reverse the effect of
miR-574-5p, and the proliferative ability was significantly
increased.
Flow cytometry results identified that the apoptotic
rate of OA chondrocytes was markedly increased compared with the
Blank group, while the apoptotic rate in the IL-1β + CTS group was
decreased and was increased after transfection with
miR-574-5p-mimic (Fig. 3G).
Moreover, the addition of YAF2 decreased the apoptotic rate of
chondrocytes. These results suggested that CTS could affect
chondrocyte proliferation and apoptosis by regulating the
expression of miR-574-5p and then interfering with YAF2.
CTS regulates the methylation of the
miR-574-5p promoter
Since it was found that the CTS/miR-574-5p/YAF2
signaling pathway could affect the proliferation and apoptosis of
arthritic chondrocytes, this study further aimed to investigate how
CTS regulates the expression of miR-574-5p and thus affects YAF2
expression. It has been reported that CTS can upregulate the
methylation levels of some molecular promoters in prostate cancer
(25). Therefore, the 2 kb region
upstream of miR-574-5p was examined, and the presence of CpG island
was found (Fig. 4A), indicating
that CTS may regulate miR-574-5p expression by affecting its
methylation. The methylation levels of the miR-574-5p promoter in
the Blank group, IL-1β group, IL-1β + CTS group and IL-1β + CTS +
5-aza-CdR group were measured via MSPCR. The methylation degree of
the miR-574-5p promoter was reduced in arthritic chondrocytes, but
was restored after the addition of CTS. After the addition of the
methylation inhibitor 5-aza-CdR, the methylation degree of the
miR-574-5p promoter was reduced, indicating that CTS could regulate
the methylation of the miR-574-5p promoter (Fig. 4B).
Discussion
Non-coding RNA (ncRNA) is a type of RNA that does
not encode proteins. According to the length of ncRNA fragments,
they can be divided into short ncRNA, such as small interfering
RNA, piwi-interacting RNA, miRNA and long non-coding RNA (lncRNA)
(26). ncRNAs are involved in
almost every step of the life cycle, from gene transcription to
mRNA splicing to RNA decay and translation (27).
miRNAs are a class of small molecular ncRNAs with a
length of 20–23 nucleotides. Previous studies have reported that
miRNAs, as an important regulatory factor, are often observed in
various diseases, and ~1/2 of the mRNAs are regulated by miRNAs
(28,29). Moreover, studies have confirmed that
miRNAs and lncRNAs are involved in regulating OA progression by
regulating chondrocyte activity and function (27–29).
In order to identify key miRNAs that can affect OA, the present
study established an OA mouse model. The C57BL/6 mice used in this
study are easy to feed and are often used as experimental animal
models of physiology and pathology, such as obesity, type 2
diabetes, atherosclerosis and osteoporosis, and also used as an OA
model (15,30). Next, the current study conducted
bioinformation analysis on the OA mouse data set and found four
conserved miRNAs, among which the expression level of miR-574-5p
was the highest in the knee joint of mice with OA. Based on the
protective effect of CTS on chondrocytes and the differential
expression of miR-574-5p, it was suggested that CTS may promote
chondrocyte proliferation and inhibit its apoptosis by regulating
the expression of miR-574-5p, thus alleviating OA. However, the
specific signal pathway remains unknown.
YAF2, a zinc finger protein, was first identified
due to its interaction with the transcription factor Yin Yang 1,
which has an effect on the proliferation of tumor cells (31). In genotoxic stress reactions, YAF2
can positively regulate the function of programmed cell death 5 in
the human lung adenocarcinoma cell line A549 and human melanoma
cell line HCT116 (32). Other
studies have shown that the interaction between YAF2 and polycomb
group protein can inhibit the apoptosis induced by caspase 8 during
the early embryonic development of zebrafish (33). Based on the protective effect of
YAF2 on chondrocytes by regulating apoptosis, the present study
found that expression levels of YAF2 and the anti-apoptotic gene
Bcl-2 were significantly decreased in mice with OA, while the
expression levels of miR-574-5p and the pro-apoptotic gene Bax
showed opposite changes. The in vitro experiments
demonstrated that the expression levels of miR-574-5p, YAF2, Bax
and Bcl-2 in chondrocytes were consistent with the in vivo
findings. Furthermore, the addition of CTS could alter the effect,
which verified that CTS could alleviate OA.
Next, the present study aimed to determine how CTS
regulates the expression levels of miR-574-5P and YAF2, to
ultimately affect chondrocyte apoptosis. Using biogenic analysis
and verification experiments, it was identified that miR-574-5p
could interact with the YAF2 promoter, and exerted a regulating
effect on YAF2. The miR-574-5P/YAF2 pathway has been confirmed, but
it is necessary to further study how CTS regulates the expression
of miR-574-5p. As a potential active ingredient in the treatment of
prostate cancer, CTS could regulate the methylation of the key
factor histone H3 lysine 9 (25).
After the analysis and prediction, the present study further found
that the methylation degree of the miR-574-5p promoter was
increased by the effect of CTS, and was reduced after the addition
of the methylation inhibitor.
In conclusion, the present study provided novel
targets for the treatment of OA with CTS and validated that
miR-574-5p is an important regulatory factor affecting OA.
Furthermore, the present study investigated the mechanism via which
CTS affects chondrocyte apoptosis by regulating the methylation of
miR-574-5P, thus interfering with YAF2.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by Special project of
Scientific Research on Traditional Chinese Medicine of Henan
Province (grant no. 2018ZY2019).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SY and MG designed the study. SY and XS wrote the
manuscript. MG supervised the study. JT and JW analyzed the data.
SY and JW prepared the figures. The experiments were performed by
XS, JT and JW. SY and MG confirm the authenticity of all the raw
data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Ethics Committee of Orthopedic-Traumatological Hospital of Henan
Province (Henan Provincial Orthopedic Hospital) (approval no.
KY201902046).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Loeser RF, Collins JA and Diekman BO:
Ageing and the pathogenesis of osteoarthritis. Nat Rev Rheumatol.
12:412–420. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Keen HI, Hensor EMA, Wakefield RJ, Mease
PJ, Bingham CO III and Conaghan PG: Ultrasound assessment of
response to intra-articular therapy in osteoarthritis of the knee.
Rheumatology (Oxford). 54:1385–1391. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bellot G, Garcia-Medina R, Gounon P,
Chiche J, Roux D, Pouysségur J and Mazure NM: Hypoxia-induced
autophagy is mediated through hypoxia-inducible factor induction of
BNIP3 and BNIP3L via their BH3 domains. Mol Cell Biol.
29:2570–2581. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cleveland RJ, Alvarez C, Schwartz TA,
Losina E, Renner JB, Jordan JM and Callahan LF: The impact of
painful knee osteoarthritis on mortality: A community-based cohort
study with over 24 years of follow-up. Osteoarthritis Cartilage.
27:593–602. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nho SJ, Kymes SM, Callaghan JJ and Felson
DT: The burden of hip osteoarthritis in the United States:
Epidemiologic and economic considerations. J Am Acad Orthop Surg.
21 (Suppl 1):S1–S6. 2013. View Article : Google Scholar
|
|
6
|
Hosseinzadeh A, Kamrava SK, Joghataei MT,
Darabi R, Shakeri-Zadeh A, Shahriari M, Reiter RJ, Ghaznavi H and
Mehrzadi S: Apoptosis signaling pathways in osteoarthritis and
possible protective role of melatonin. J Pineal Res. 61:411–425.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vuppalapati KK, Bouderlique T, Newton PT,
Kaminskyy VO, Wehtje H, Ohlsson C, Zhivotovsky B and Chagin AS:
Targeted deletion of autophagy genes Atg5 or Atg7 in the
chondrocytes promotes caspase-dependent cell death and leads to
mild growth retardation. J Bone Miner Res. 30:2249–2261. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Varela-Eirin M, Loureiro J, Fonseca E,
Corrochano S, Caeiro JR, Collado M and Mayan MD: Cartilage
regeneration and ageing: Targeting cellular plasticity in
osteoarthritis. Ageing Res Rev. 42:56–71. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Charlier E, Relic B, Deroyer C, Malaise O,
Neuville S, Collée J, Malaise MG and De Seny D: Insights on
molecular mechanisms of chondrocytes death in osteoarthritis. Int J
Mol Sci. 17:21462016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guo M, Guo J, Cui H and Guo Y and Guo Y:
Effects of Zhuanggu huoxue tang on gucuronic acid and water
contents in cartilage of rabbits with osteoarthritis. Chin J Tradit
Med Sci Technol. 23:538–539. 2016.
|
|
11
|
Tao J, Wang S, Chen Y, Zheng H, Jiang M
and Yuan B: Review on the research in network pharmacology of
traditional Chinese medicine compound. China J Tradit Chin Med
Pharm. 34:3903–3907. 2019.
|
|
12
|
Liu L, Liu HP, Xie JR, Yang LY, Wang MY,
Peng T, Liu PA and Zhang GM: Association analysis on effective
components in Zhuanggu Zhitong prescription. Chin Tradit Herbal
Drugs. 49:1451–1460. 2018.
|
|
13
|
Ashrafizadeh M, Zarrabi A, Orouei S,
Saberifar S, Salami S, Hushmandi K and Najafi M: Recent advances
and future directions in anti-tumor activity of cryptotanshinone: A
mechanistic review. Phytother Res. 35:155–179. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu YH, Wu YR, Li B and Yan ZY:
Cryptotanshinone: A review of its pharmacology activities and
molecular mechanisms. Fitoterapia. 145:1046332020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Feng Z, Zheng W, Li X, Lin J, Xie C, Li H,
Cheng L, Wu A and Ni W: Cryptotanshinone protects against
IL-1β-induced inflammation in human osteoarthritis chondrocytes and
ameliorates the progression of osteoarthritis in mice. Int
Immunopharmacol. 50:161–167. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kung LHW, Ravi V, Rowley L, Angelucci C,
Fosang AJ, Bell KM, Little CB and Bateman JF: Cartilage MicroRNA
dysregulation during the onset and progression of mouse
osteoarthritis is independent of aggrecanolysis and overlaps with
candidates from end-stage human disease. Arthritis Rheumatol.
70:383–395. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ouyang J, Jiang H, Fang H, Cui W and Cai
D: Isoimperatorin ameliorates osteoarthritis by downregulating the
mammalian target of rapamycin C1 signaling pathway. Mol Med Rep.
16:9636–9644. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pirozzi C, Francisco V, Guida FD, Gómez R,
Lago F, Pino J, Meli R and Gualillo O: Butyrate modulates
inflammation in chondrocytes via GPR43 receptor. Cell Physiol
Biochem. 51:228–243. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Na HS, Park JS, Cho KH, Kwon JY, Choi J,
Jhun J, Kim SJ, Park SH and Cho ML: Interleukin-1-interleukin-17
signaling axis induces cartilage destruction and promotes
experimental osteoarthritis. Front Immunol. 11:7302020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ji Q, Qi D, Xu X, Xu Y, Goodman SB, Kang
L, Song Q, Fan Z, Maloney WJ and Wang Y: Cryptotanshinone protects
cartilage against developing osteoarthritis through the
miR-106a-5p/GLIS3 axis. Mol Ther Nucleic Acids. 11:170–179. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Barra GB, Santa Rita TH, Almeida ALSC,
Jácomo RH and Nery LFA: Serum has higher proportion of janus kinase
2 V617F mutation compared to paired EDTA-whole blood sample: A
model for somatic mutation quantification using qPCR and the 2-∆∆Cq
method. Diagnostics (Basel). 10:1532020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang Q, Li X, Zhou Y, Fu W, Wang J and Wei
Q: A LINC00341-mediated regulatory pathway supports chondrocyte
survival and may prevent osteoarthritis progression. J Cell
Biochem. 120:10812–10820. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Knudson CM and Korsmeyer SJ: Bcl-2 and Bax
function independently to regulate cell death. Nat Genet.
16:358–363. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu CY, Hsieh CY, Huang KE, Chang C and
Kang HY: Cryptotanshinone down-regulates androgen receptor
signaling by modulating lysine-specific demethylase 1 function. Int
J Cancer. 131:1423–1434. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dong X, Dong X, Gao F, Liu N, Liang T,
Zhang F, Fu X, Pu L and Chen J: Non-coding RNAs in cardiomyocyte
proliferation and cardiac regeneration: Dissecting their
therapeutic values. J Cell Mol Med. 25:2315–2332. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Razmara E, Bitaraf A, Yousefi H, Nguyen
TH, Garshasbi M, Cho WC and Babashah S: Non-coding RNAs in
cartilage development: An updated review. Int J Mol Sci.
20:44752019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang Y, Wang G, Ma L, Wang C, Wang L, Guo
Y and Zhao X: miR-137 suppresses cell growth and extracellular
matrixdegradation through regulating ADAMTS-5 in chondrocytes. Am J
Transl Res. 11:7027–7034. 2019.PubMed/NCBI
|
|
29
|
Swingler TE, Niu L, Smith P, Paddy P, Le
L, Barter MJ, Young DA and Clark IM: The function of microRNAs in
cartilage and osteoarthritis. Clin Exp Rheumatol. 37 (Suppl
120):S40–S47. 2019.
|
|
30
|
Monteagudo S, Cornelis FMF, Aznar-Lopez C,
Yibmantasiri P, Guns LA, Carmeliet P, Cailotto F and Lories RJ:
DOT1L safeguards cartilage homeostasis and protects against
osteoarthritis. Nat Commun. 8:158892017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kalenik JL, Chen D, Bradley ME, Chen SJ
and Lee TC: Yeast two-hybrid cloning of a novel zinc finger protein
that interacts with the multifunctional transcription factor YY1.
Nucleic Acids Res. 25:843–849. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Park SY, Choi HK, Jo SH, Seo J, Han EJ,
Choi KC, Jeong JW, Choi Y and Yoon HG: YAF2 promotes TP53-mediated
genotoxic stress response via stabilization of PDCD5. Biochim
Biophys Acta. 1853:1060–1072. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Stanton SE, McReynolds LJ, Evans T and
Schreiber-Agus N: Yaf2 inhibits caspase 8-mediated apoptosis and
regulates cell survival during zebrafish embryogenesis. J Biol
Chem. 281:28782–28793. 2006. View Article : Google Scholar : PubMed/NCBI
|