Introduction
Kidney cancer is a malignant tumor of the urinary
system, accounting for 2.2% of all adult malignancies worldwide
(1). The most common type of kidney
cancer is renal cell carcinoma (RCC), which accounts for 85–90% of
all cases (2). According to the
classification of International Society of Urological Pathology,
RCC is primarily divided into three subtypes: i) Clear cell
(ccRCC); ii) papillary; and iii) chromophobe (3), of which ccRCC is the most common
subtype (4). In the past two
decades, the incidence of kidney cancer has been increasing by 2%
each year worldwide (5). Although
progress has been made and the 5-year survival rate of patients
with kidney cancer has increased by ~30% in recent years due to the
early diagnosis of low-grade tumors, there were 403,262 new cases
of kidney cancer and 175,098 related deaths worldwide in 2018 based
on the GLOBOCAN 2018 estimates (1).
Therefore, the identification of novel prognostic candidate genes
for predicting the prognosis of patients with kidney cancer is
important.
SEC61 translocon subunit-γ (SEC61G) is a member of
the SEC61 translocon, which is composed of three subunits in
mammals, namely Sec61α, Sec61β and Sec61γ (6). SEC61 translocon is the central
component of the protein translocation apparatus of the endoplasmic
reticulum (ER) membrane, which mediates the integration of
ribosomally synthesized unfolded proteins into the ER membrane and
the translocation of nascent polypeptides into the ER lumen
(7). It has been also reported that
SEC61 directly interacts with SEC62 and SEC63 to promote protein
folding, modification and translocation (8).
Over the past few years, mutations in the SEC61A1
gene have been associated with autosomal dominant
tubulointerstitial kidney disease in humans, diabetes and
hepatosteatosis in mice, and professional secretory cell apoptosis
(9–11). In addition, SEC63 gene mutations
were associated with autosomal dominant polycystic liver disease
(12,13). Furthermore, amplification and
upregulation of SEC genes, primarily SEC62 and SEC63, was detected
in several types of human cancer, including gastric, colorectal,
cervical and non-small cell lung cancer, as well as small bowel
cancer associated with hereditary non-polyposis colorectal cancer
and hepatocellular carcinoma associated with Lynch syndrome
(14–19). Regarding SEC61G, a previous study
reported that SEC61G was remarkably overexpressed in 77% of
glioblastoma multiforme cases, whereas SEC61G knockdown attenuated
tumor cell proliferation and induced cell apoptosis (20). However, whether SEC61G is
upregulated in kidney cancer tissues and associated with tumor
progression is not completely understood.
In the present study, The Cancer Genome Atlas (TCGA)
database was screened to obtain the expression profile of SEC61G
and identify its association with kidney cancer prognosis.
Furthermore, SEC61G was knocked down in kidney cancer cells and the
in vitro effect of SEC61G knockdown on kidney cancer cell
proliferation, migration, invasion and apoptosis was investigated.
All these efforts aimed to investigate the role of SEC61G in kidney
cancer, which will provide a novel target or biomarker for the
treatment and prognosis of kidney cancer.
Materials and methods
Cell culture
A498 and 769-P cell lines were purchased from The
Cell Bank of Type Culture Collection of The Chinese Academy of
Sciences. A498 cells were cultured in DMEM (Hyclone; Cytiva)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.),
100 U/ml penicillin (Sigma-Aldrich; Merck KGaA) and 100 mg/µl
streptomycin (Sigma). 769-P cells were cultured in RPMI-1640
(Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% FBS,
100 U/ml penicillin and 100 mg/ml streptomycin. Cells were
maintained at 37°C with 5% CO2. Cells were mycoplasma
free and authenticated by STR assay. Cells were seeded into 6-well
plates and allowed to grow to 80% confluence prior to use in
subsequent experiments.
Bioinformatics analysis
The University of Alabama Cancer Database (UALCAN;
ualcan.path.uab.edu/index.html) (21) was used to assess the expression of
SEC61G in kidney cancer based on TCGA (https://cancergenome.nih.gov/) data. The Gene
Expression Profiling Interactive Analysis (GEPIA; gepia.cancer-pku.cn/index.html)
(22) was used to construct
survival curves based on SEC61G gene expression data and clinical
data in kidney cancer samples obtained from TCGA database, which
were analyzed using the log-rank test. The cut-off value for
overall survival (OS) was set to the median SEC61G gene expression
value. The expression profile of SEC61G was clearly defined in
human renal cancer using UALCAN and GEPIA.
Small interfering (si)RNA
transfection
A498 and 769-P cells were seeded into 6-well plates
at a density of 4–8×105 cell/well. At 2 h prior to
transfection, culture medium was replaced with fresh complete
medium (RPMI-1640 supplemented with 10% FBS, 100 U/ml penicillin
and 100 mg/ml streptomycin). Subsequently, cells were transfected
with 5 µl siRNAs against SEC61G (5′-CAUUGUUGGUGGCUGAAUATT-3′) or
scrambled NC siRNA (5′-TTCTCCGAACGTGTCACGT-3′) at room temperature
(~25°C) using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
Following incubation for 6 h, cells were supplemented with fresh
complete culture medium and cultured for an additional 48 h. Then,
cells were used for subsequent experiments. siRNAs were purchased
from Oligobio Biotechnology Co., Ltd. (http://www.oligobio.com.cn/).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from different transfected
A498 and 769-P cells using an Ultrapure RNA kit (CoWin Biosciences)
according to the manufacturer's protocol. Total RNA was reverse
transcribed into cDNA using HiFiScript cDNA synthesis kits (CoWin
Biosciences) following the manufacturer's protocol. Subsequently,
with the SYBR Master Mixture (CoWin Biosciences), qPCR was
performed to assess the effect of SEC61G knockdown on SEC61G mRNA
gene expression levels. The following primers were used for qPCR
(synthesized by Genewiz, Inc.): GAPDH forward,
5′-TGACTTCAACAGCGACACCCA-3′ and reverse,
5′-CACCCTGTTGCTGTAGCCAAA-3′; and Sec61G forward,
5′-TGGATCAGGTAATGCAGTTT-3′ and reverse, 5′-TCAGCCACCAACAATGATG-3′.
The PCR reaction was as follows: Pre-incubation at 95°C for 10 min
and incubation at 95°C for 30 sec, followed by 40 cycles of 95°C
for 15 sec, 60°C for 1 min and a final dissociation stage (95°C for
15 sec, 60°C for 1 min and 95°C for 15 sec). mRNA expression levels
were quantified using the 2−ΔΔCq method (23) and normalized to the internal
reference gene GAPDH.
Western blotting
Total protein was isolated from A498 and 769-P cells
using RIPA lysis buffer. The concentration of total protein was
detected using the BCA method. Proteins (20 µg per lane) were
separated via 10% SDS-PAGE and transferred to PVDF membranes (EMD
Millipore). After blocking with TBS containing 0.1% Tween-20
containing 5% skimmed milk at room temperature for 1 h, the
membranes were incubated with primary antibodies targeted against:
SEC61G (Rabbit; 1:500; cat. no. 11147-2-AP; ProteinTech Group,
Inc.), E-cadherin (Rabbit; 1:1,000; cat. no. 20874-1-AP;
ProteinTech Group, Inc.), N-cadherin (Rabbit; 1:2,000; cat. no.
22018-1-AP; ProteinTech Group, Inc.), Bax (Rabbit; 1:2,000; cat.
no. 50599-2-lg; ProteinTech Group, Inc.), GAPDH (Mouse; 1:5,000;
cat. no. 60004-1-Ig; ProteinTech Group, Inc.), β-catenin (Rabbit;
1:1,000; cat. no. AF6266; Affinity Biosciences), BCL-2 (Rabbit;
1:1,000; cat. no. AF6139; Affinity Biosciences), Cleaved Caspase-3
(Rabbit; 1:1,000; cat. no. AF7022; Affinity Biosciences), AKT
(Rabbit; 1:1,000; cat. no. AF6261; Affinity Biosciences), and p-AKT
(Rabbit; 1:1,000; cat. no. AF0016; Affinity Biosciences) overnight
at 4°C. Following primary antibody incubation, the membranes were
incubated with corresponding HRP-conjugated anti-rabbit IgG (Goat;
1:5,000; cat. no. S0001; Affinity Biosciences) or anti-mouse IgG
(Goat; 1:5,000; cat. no. S0002; Affinity Biosciences) secondary
antibodies at room temperature for 2 h. Protein bands were
visualized using an ECL detection kit (Pierce; Thermo Fisher
Scientific, Inc.). Protein expression levels were semi-quantified
using Quantity One Software (version 4.2.2; Bio-Rad Laboratories,
Inc.) with GAPDH as the loading control.
Cell proliferation assay
A498 and 769-P cell proliferation was evaluated by
performing a Cell Counting Kit-8 (CCK-8) assay. Briefly, cells were
harvested, resuspended in 100 µl medium (1.5×104
cells/ml), seeded into 96-well plates (2,000 cells/well) and
incubated at 37°C. At 24 h intervals, 10 µl CCK-8 reagent (Beijing
TransGen Biotech Co., Ltd.) was added to each well and incubated at
37°C for an additional 1.5 h. Absorbance was measured at a
wavelength of 450 nm using a microplate reader.
Cell apoptosis assay
Flow cytometry was performed to determine A498 and
769-P cell apoptosis using the FITC-Annexin V Apoptosis Detection
kit (BD Biosciences) according to the manufacturer's protocol. At
24 h post-transfection, cells (1–5×105 cells/ml) were
supplemented in serum-free medium and cultured for an additional 24
h. Subsequently, cells were harvested, resuspended, incubated with
5 µl FITC-conjugated Annexin V at room temperature for 5 min and
then incubated with 10 µl PI in the dark at room temperature for 5
min. Cell apoptosis was analyzed using FACSCalibur flow cytometer
(BD Biosciences) and FlowJo software (version 7.6.1; FlowJo LLC).
The percentage of total apoptotic events was defined as the sum of
the apoptotic cells in the early stage (Annexin V positive/PI
negative) and late stage (Annexin V positive/PI positive).
Cell invasion and migration assay
A498 and 769-P cell migration and invasion were
assessed using 24-well Transwell chambers. To assess cell invasion,
100 µl Matrigel was added to the upper chamber and incubated for 2
h. Subsequently, 500 µl serum-free medium was plated into a 24-well
Transwell chamber for 30 min. Then, 100 µl cell suspension
(5×104 cells) was plated into the upper chamber, and 600
µl complete medium containing 10% FBS was plated into the lower
chamber. Following incubation at 37°C for 24 h, cells on the upper
surface of the filter membrane were removed using cotton swabs.
Invading cells were fixed with 4% paraformaldehyde at room
temperature for 15 min and stained with 0.1% Giemsa at room
temperature for 5 min. Stained cells were quantified in five
randomly selected fields of view per filter under a fluorescence
microscope (CKX 51; Olympus Corporation). To assess cell migration,
the aforementioned protocol was performed, but the upper chamber
was not precoated with Matrigel.
Wound healing assay
Cells were seeded (5×105 cells/well) into
6-well plates containing RPMI-1640 with 10% FBS to form a confluent
cell monolayer. The following day, an artificial wound was created
using a sterile 200-µl pipette tip. Following washing with PBS,
cells were cultured for 24 h. Wounds were observed at 0 and 24 h
using an inverted fluorescence microscope (CKX51; Olympus
Corporation). The wound closure area of the migrating monolayer of
cells was quantified using ImageJ software (version 1.49; National
Institutes of Health).
Statistical analysis
All experiments were repeated three times.
Statistical analyses were performed using SPSS software (version
18.0; SPSS, Inc.). Comparisons between groups were analyzed using
the unpaired Student's t-test. Data are presented as the mean ± SD.
P<0.05 was considered to indicate a statistically significant
difference.
Results
SEC61G expression and its association
with survival in patients with kidney cancer
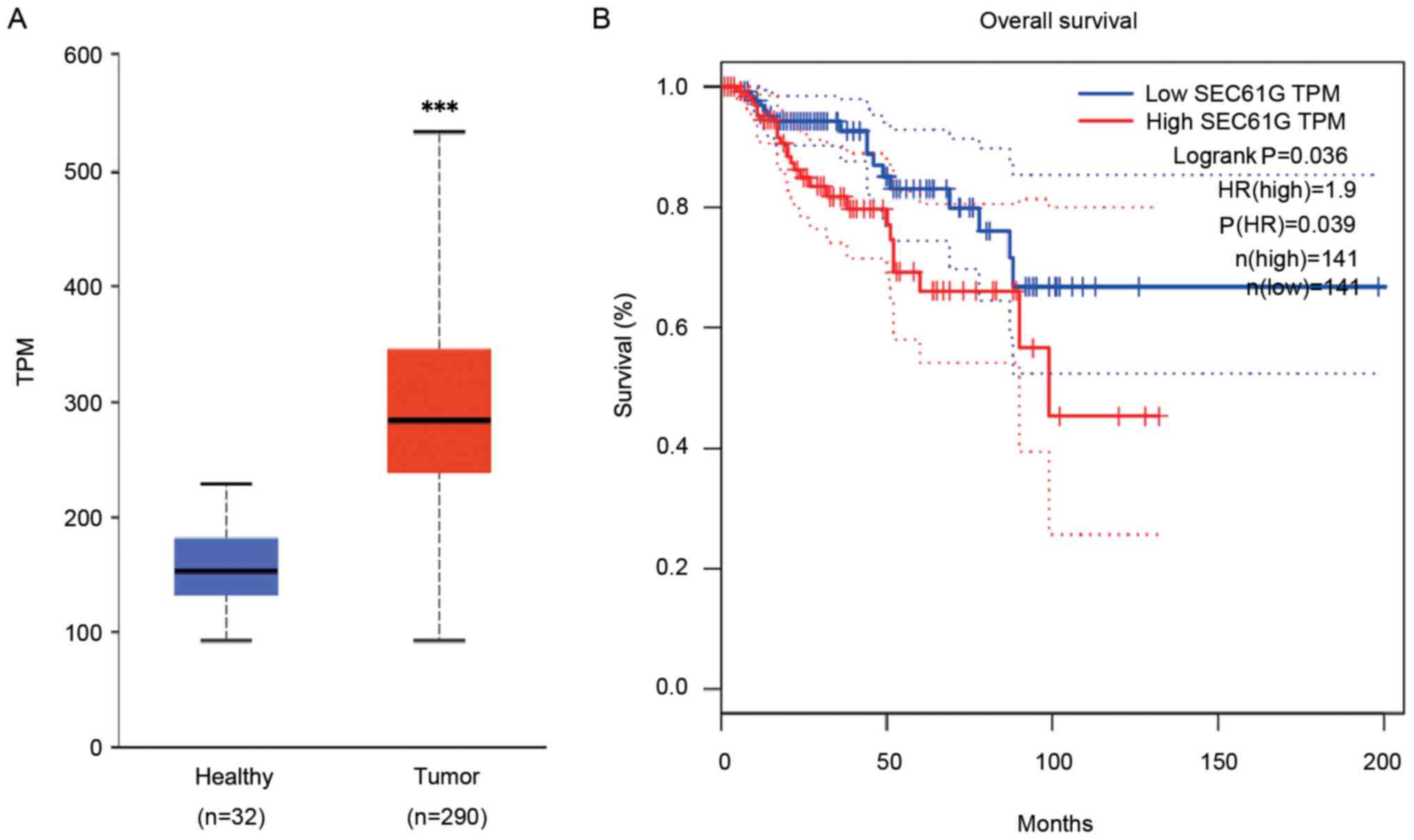
To investigate whether SEC61G served as a novel and
efficient candidate gene for predicting kidney cancer, the
expression profile of SEC61G was obtained using the UALCAN web-tool
based on TCGA database. The results demonstrated that SEC61G
expression was significantly increased in human kidney tumor
tissues compared with healthy tissues (P<0.001; Fig. 1A). Subsequently, the association
between increased expression levels of SEC61G and the survival of
patients with kidney cancer was investigated. Kaplan-Meier curve
analysis revealed that high SEC61G expression levels were
significantly associated with poor OS in patients with kidney
cancer compared with low SEC61G expression levels (Fig. 1B). The results indicated that SEC61G
might serve as a biomarker for kidney cancer.
SEC61G knockdown attenuates human
kidney cancer cell proliferation
Tumor progression is a complex process, which is
characterized by tumor cell proliferation, migration, invasion,
metastasis, colony formation and adhesion (24,25).
Therefore, the present study investigated the effects of SEC61G on
cell proliferation, migration, invasion and survival.
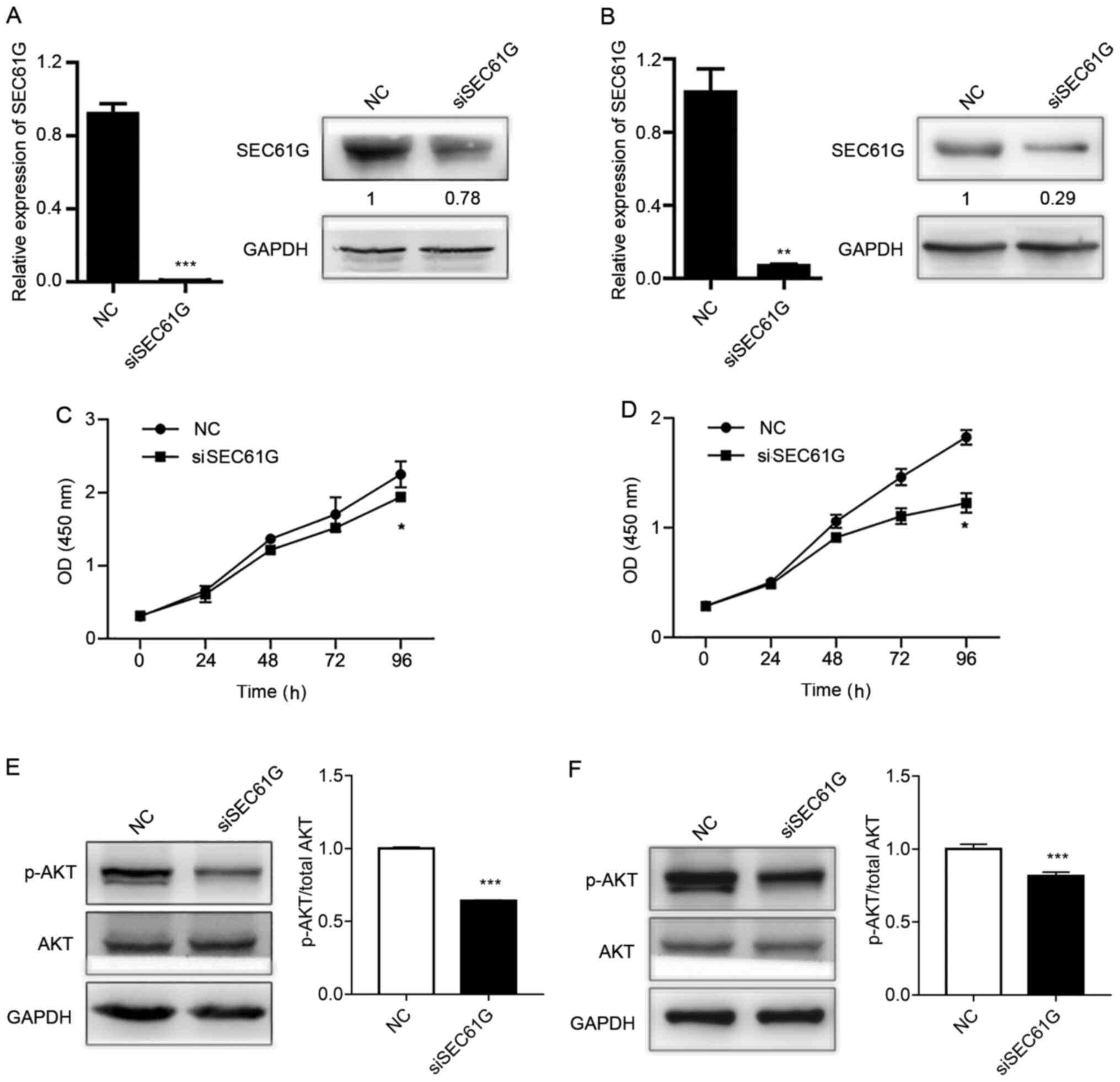
To determine the role of SEC61G in human kidney
cancer cell proliferation, A498 and 769-P cells were transfected
with SEC61G siRNA. Following transfection with SEC61G siRNA, SEC61G
expression levels were dramatically decreased at the mRNA and
protein levels in A498 (Fig. 2A)
and 769-P (Fig. 2B) cells compared
with the NC group. Subsequently, the present study examined whether
SEC61G was essential for cell proliferation. The CCK-8 assay
results demonstrated that SEC61G knockdown significantly inhibited
A498 (Fig. 2C) and 769-P (Fig. 2D) cell proliferation compared with
the NC group.
The PI3K/AKT signaling pathway promotes cell
proliferation in the majority of cell types, including kidney
cancer cells (26,27). In addition, it has been reported
that the PI3K/AKT signaling pathway is activated in ~50% of RCC
cases (28–30). Therefore, the effect of SEC61G
knockdown on the activation status of AKT in A498 and 769-P cells
was assessed. The results revealed that AKT phosphorylation was
significantly decreased by SEC61G knockdown in both kidney cancer
cell lines compared with the NC group (Fig. 2E and F). The results indicated that
SEC61G affected human kidney cancer cell proliferation via the
PI3K/AKT signaling pathway.
SEC61G knockdown inhibits human kidney
cancer cell migration
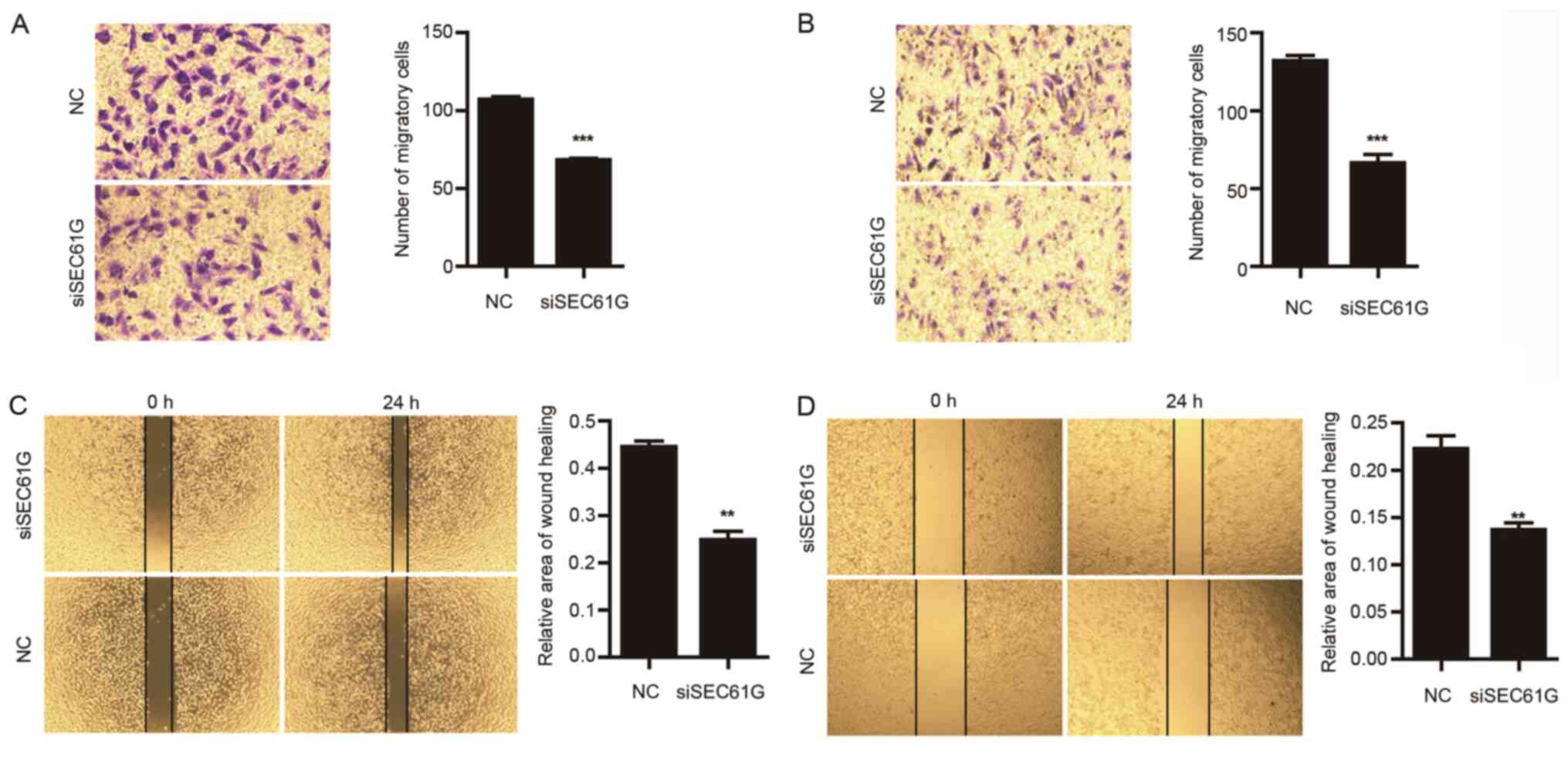
Subsequently, the effect of SEC61G on cell migration
was investigated. Compared with the NC group, SEC61G knockdown
significantly decreased A498 (Fig.
3A) and 769-P (Fig. 3B) cell
migration. The effect of SEC61G on cell migration was further
assessed by performing wound healing assays. Consistent with the
previous findings, SEC61G knockdown significantly decreased the
area of wound healing compared with the NC group (Fig. 3C and D). Therefore, the results
suggested that SEC61G knockdown inhibited kidney cancer cell
migration compared with the NC group.
SEC61G knockdown inhibits human kidney
cancer cell invasion
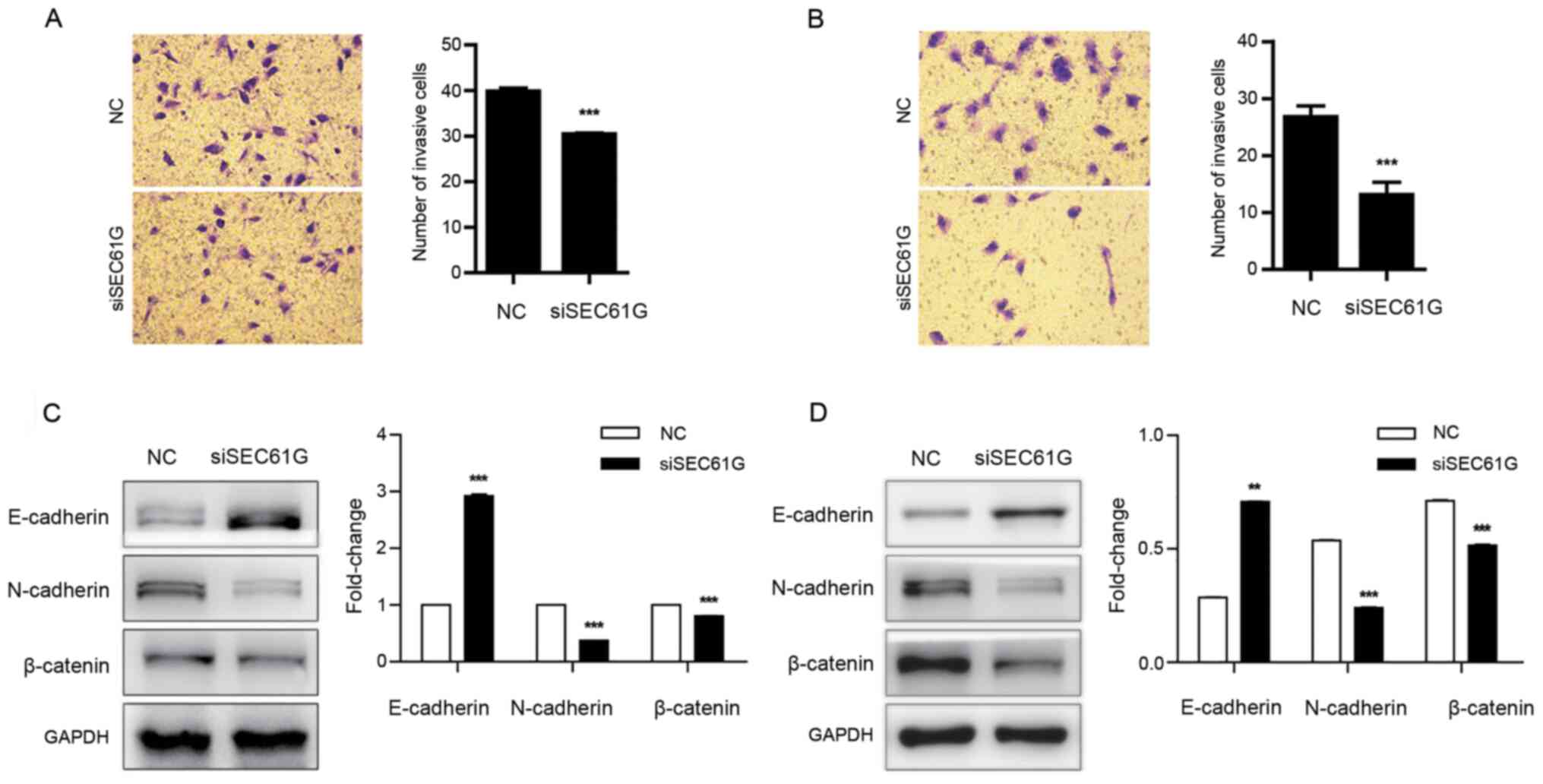
To evaluate the effect of SEC61G on kidney cancer
cell invasion, a cell invasion assay was performed. The results
demonstrated that SEC61G knockdown significantly inhibited A498
(Fig. 4A) and 769-P (Fig. 4B) cell invasion compared with the NC
group.
Several transcriptional factors are involved in
cancer cell invasion and migration, including E-cadherin,
N-cadherin and β-catenin (31,32).
It has been reported that during cell invasion and migration,
E-cadherin is downregulated, whereas N-cadherin and β-catenin are
upregulated (33). Therefore, the
expression levels of E-cadherin, N-cadherin and β-catenin were
assessed following SEC61G knockdown in A498 and 769-P cells.
Compared with the NC group, SEC61G knockdown significantly
increased E-cadherin expression levels, and significantly decreased
N-cadherin and β-catenin expression levels (Fig. 4C and D). The aforementioned results
indicated that SEC61G enhanced kidney cancer cell invasion.
SEC61G knockdown promotes human kidney
cancer cell apoptosis
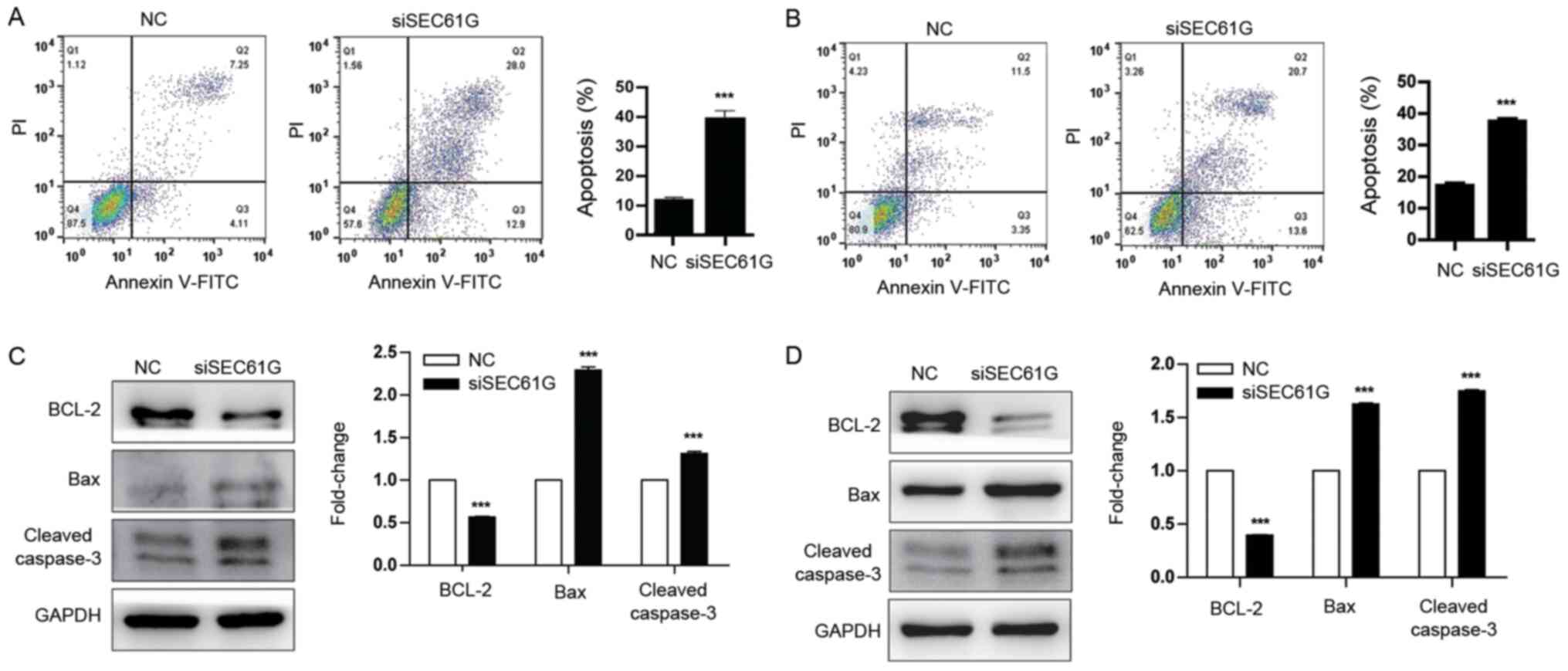
To further explore the role of SEC61G in kidney
cancer cell apoptosis, Annexin V/PI staining was performed.
Compared with the NC group, SEC61G knockdown significantly
increased A498 and 769-P cell apoptosis (Fig. 5A and B). In addition, the expression
levels of the apoptosis-related proteins BCL-2, Bax and Cleaved
caspase-3 were also determined. The western blotting results
demonstrated that compared with the NC group, SEC61G knockdown
significantly downregulated the expression of the antiapoptotic
protein BCL-2 and significantly upregulated the expression of the
proapoptotic protein Bax (Fig. 5C and
D). In addition, the protein expression levels of the
apoptosis-related marker Cleaved caspase-3 were also significantly
increased in SEC61G-knockdown A498 and 769-P cells compared with
the NC group. Collectively, the results suggested that SEC61G
promoted human kidney cancer cell survival.
Discussion
By using TCGA database, it was found that SEC61G was
upregulated in kidney cancer tissues compared with healthy tissues.
Additionally, high SEC61G expression levels were associated with
poor survival rates in patients with kidney cancer. Moreover,
SEC61G knockdown significantly inhibited kidney cancer cell
proliferation, migration and invasion, and enhanced cell apoptosis
via a caspase-dependent signaling pathway. Taken together, the
present study demonstrated that SEC61G might be a proto-oncogene in
human kidney cancer. The results suggested that SEC61G might serve
as a novel candidate gene for predicting the prognosis of patients
with kidney cancer.
The SEC61 complex, composed of SEC61 α, β and γ, is
the central component of the protein translocation apparatus for
the translocation of nascent polypeptides into the ER lumen and the
integration of transmembrane proteins into the ER bilayer (6,7).
SEC61G is frequently amplified and overexpressed in several types
of tumors, such as glioblastoma (20) and hepatocellular carcinoma (34). The present study demonstrated that
SEC61G was upregulated in kidney cancer tissues compared with
healthy tissues. High expression levels of SEC61G were associated
with poor overall survival in patients with kidney cancer,
suggesting that SEC61G might serve as a marker of poor prognosis
for patients with kidney cancer.
The uncontrolled proliferation of cancer cells is
considered as a hallmark of cancer (35). Previous studies have reported that
SEC61β is involved in the trafficking of EGFR and EGFR-mediated
activation of the PI3K/AKT signaling pathway (36,37).
The PI3K/AKT signaling pathway promotes cell proliferation and
inhibits apoptosis in the majority of cell types, including kidney
cancer cells (26,27). In addition, AKT is a
serine-threonine kinase downstream of the PTEN/PI3K signaling
pathway (38). The PI3K/AKT
signaling pathway was activated in ~50% of RCC cases (28–30).
To reveal the roles and mechanism underlying SEC61G in kidney
cancer cell proliferation, cell proliferation, apoptosis and AKT
phosphorylation were assessed in SEC61G-knockdown kidney cancer
cell lines. The results demonstrated that SEC61G knockdown
significantly inhibited kidney cancer cell proliferation and AKT
phosphorylation, and promoted cell apoptosis compared with the NC
group. Therefore, the results suggested that SEC61G promoted kidney
cancer cell proliferation via the PI3K/AKT signaling pathway.
Emerging evidence has indicated that increased tumor
cell migration and invasion are hallmarks of cancer, resulting in
tumor dissemination and aggressiveness (35,39,40).
However, the effect of SEC61G on kidney tumor cell migration and
invasion is not completely understood. In the present study, the
Transwell and wound healing assay results demonstrated that SEC61G
knockdown significantly suppressed kidney cancer cell migration and
invasion compared with the NC group. In addition, the expression
levels of E-cadherin were significantly upregulated, and the
expression levels of N-cadherin and β-catenin were significantly
decreased following SEC61G knockdown in human kidney cancer cells
compared with the NC group. Overall, the results indicated that
SEC61G knockdown inhibited kidney cancer cell migration and
invasion compared with the NC group.
siRNA technology is widely used to inhibit the
expression of cancer-specific genes for cancer treatment, and is
considered a feasible strategy (41). The present study demonstrated that
SEC61G knockdown significantly attenuated cell proliferation,
migration and invasion, and induced cell apoptosis in human kidney
cancer cells compared with the NC group. Therefore, the development
of novel drugs containing SEC61G siRNA or inhibitor may be
considered as a promising therapeutic strategy for patients with
kidney cancer.
Tumor progression is a complex process, which is
characterized by cell proliferation, migration, invasion,
metastasis, colony formation and adhesion (24,25). A
study reported that SEC61G was required for tumor cell survival in
glioblastoma multiforme (20). In
addition, the present study demonstrated that compared with healthy
tissues, SEC61G was upregulated in kidney tumor tissues, which
promoted tumor progression and predicted poor prognosis in patients
with kidney cancer. However, the molecular signaling pathways
underlying the effects of SEC61G are not completely understood.
Furthermore, the expression of SEC61G in other types of cancer,
including breast, lung and gastric cancer, as well as its role in
cancer cell migration, invasion, metastasis and colony formation
require further investigation. Therefore, further studies are
required to reveal the effects of SEC61G in tumor progression.
Collectively, the present study demonstrated that
compared with healthy tissues, SEC61G was upregulated in human
renal tumor tissues, which was associated with poor overall
survival in patients with kidney cancer based on data obtained from
TCGA database. In addition, compared with the NC group, SEC61G
knockdown significantly inhibited kidney cancer cell proliferation,
migration and invasion, and induced cell apoptosis. Therefore,
SEC61G may serve as a novel candidate gene for predicting the
progression of patients with kidney cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HM and YW were responsible for the conception and
design of the research, and drafting the manuscript or revising it
critically for important intellectual content. LG and GY confirmed
the authenticity of all the raw data, made substantial
contributions to acquisition of data and performed the experiments.
XJ and ZS performed the data analysis and interpretation. JW and YW
participated in the design of the study and performed the
statistical analysis. All authors have read and approved the
manuscript and given final approval of the version to be
published.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Choueiri TK and Motzer RJ: Systemic
therapy for metastatic renal-cell carcinoma. N Engl J Med.
376:354–366. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Srigley JR, Delahunt B, Eble JN, Egevad L,
Epstein JI, Grignon D, Hes O, Moch H, Montironi R, Tickoo SK, et
al: The international society of urological pathology (ISUP)
vancouver classification of renal neoplasia. Am J Surg Pathol.
37:1469–1489. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nickerson ML, Jaeger E, Shi Y, Durocher
JA, Mahurkar S, Zaridze D, Matveev V, Janout V, Kollarova H, Bencko
V, et al: Improved identification of von Hippel-Lindau gene
alterations in clear cell renal tumors. Clin Cancer Res.
14:4726–4734. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ljungberg B, Bensalah K, Canfield S,
Dabestani S, Hofmann F, Hora M, Kuczyk MA, Lam T, Marconi L,
Merseburger AS, et al: EAU guidelines on renal cell carcinoma: 2014
update. Eur Urol. 67:913–924. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Greenfield JJ and High S: The Sec61
complex is located in both the ER and the ER-Golgi intermediate
compartment. J Cell Sci. 112:1477–1486. 1999.PubMed/NCBI
|
|
7
|
Osborne AR, Rapoport TA and van den Berg
B: Protein translocation by the Sec61/SecY channel. Annu Rev Cell
Dev Biol. 21:529–550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zimmermann R, Muller L and Wullich B:
Protein transport into the endoplasmic reticulum: Mechanisms and
pathologies. Trends Mol Med. 12:567–573. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schauble N, Lang S, Jung M, Cappel S,
Schorr S, Ulucan O, Linxweiler J, Dudek J, Blum R, Helms V, et al:
BiP-mediated closing of the Sec61 channel limits Ca2+
leakage from the ER. EMBO J. 31:3282–3296. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bolar NA, Golzio C, Zivna M, Hayot G, Van
Hemelrijk C, Schepers D, Vandeweyer G, Hoischen A, Huyghe JR, Raes
A, et al: Heterozygous loss-of-function SEC61A1 mutations cause
autosomal-dominant tubulo-interstitial and glomerulocystic kidney
disease with anemia. Am J Hum Genet. 99:174–187. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lloyd DJ, Wheeler MC and Gekakis N: A
point mutation in Sec61alpha1 leads to diabetes and hepatosteatosis
in mice. Diabetes. 59:460–470. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Davila S, Furu L, Gharavi AG, Tian X, Onoe
T, Qian Q, Li A, Cai Y, Kamath PS, King BF, et al: Mutations in
SEC63 cause autosomal dominant polycystic liver disease. Nat Genet.
36:575–577. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Drenth JP, Martina JA, van de Kerkhof R,
Bonifacino JS and Jansen JB: Polycystic liver disease is a disorder
of cotranslational protein processing. Trends Mol Med. 11:37–42.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mori Y, Sato F, Selaru FM, Olaru A, Perry
K, Kimos MC, Tamura G, Matsubara N, Wang S, Xu Y, et al:
Instabilotyping reveals unique mutational spectra in
microsatellite-unstable gastric cancers. Cancer Res. 62:3641–3645.
2002.PubMed/NCBI
|
|
15
|
Schulmann K, Brasch FE, Kunstmann E, Engel
C, Pagenstecher C, Vogelsang H, Krüger S, Vogel T, Knaebel HP,
Rüschoff J, et al: HNPCC-associated small bowel cancer: Clinical
and molecular characteristics. Gastroenterology. 128:590–599. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Casper M, Weber SN, Kloor M, Müllenbach R,
Grobholz R, Lammert F and Zimmer V: Hepatocellular carcinoma as
extracolonic manifestation of Lynch syndrome indicates SEC63 as
potential target gene in hepatocarcinogenesis. Scand J
Gastroenterol. 48:344–351. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Heselmeyer K, Macville M, Schrock E,
Blegen H, Hellström AC, Shah K, Auer G and Ried T: Advanced-stage
cervical carcinomas are defined by a recurrent pattern of
chromosomal aberrations revealing high genetic instability and a
consistent gain of chromosome arm 3q. Genes Chromosomes Cancer.
19:233–240. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Allen DG, White DJ, Hutchins AM, Scurry
JP, Tabrizi SN, Garland SM and Armes JE: Progressive genetic
aberrations detected by comparative genomic hybridization in
squamous cell cervical cancer. Br J Cancer. 83:1659–1663. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dehan E, Ben-Dor A, Liao W, Lipson D,
Frimer H, Rienstein S, Simansky D, Krupsky M, Yaron P, Friedman E,
et al: Chromosomal aberrations and gene expression profiles in
non-small cell lung cancer. Lung Cancer. 56:175–184. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu Z, Zhou L, Killela P, Rasheed AB, Di C,
Poe WE, McLendon RE, Bigner DD, Nicchitta C and Yan H: Glioblastoma
proto-oncogene SEC61gamma is required for tumor cell survival and
response to endoplasmic reticulum stress. Cancer Res. 69:9105–9111.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chandrashekar DS, Bashel B, Balasubramanya
SA, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BV and Varambally
S: UALCAN: A portal for facilitating tumor subgroup gene expression
and survival analyses. Neoplasia. 19:649–658. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45:W98–W102.
2017. View Article : Google Scholar
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Steeg PS: Tumor metastasis: Mechanistic
insights and clinical challenges. Nat Med. 12:895–904. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chambers AF, Groom AC and MacDonald IC:
Dissemination and growth of cancer cells in metastatic sites. Nat
Rev Cancer. 2:563–572. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Martini M, De Santis MC, Braccini L,
Gulluni F and Hirsch E: PI3K/AKT signaling pathway and cancer: An
updated review. Ann Med. 46:372–383. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Porta C and Figlin RA:
Phosphatidylinositol-3-kinase/Akt signaling pathway and kidney
cancer, and the therapeutic potential of
phosphatidylinositol-3-kinase/Akt inhibitors. J Urol.
182:2569–2577. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hu H, Jiang C, Li G and Lu J: PKB/AKT and
ERK regulation of caspase-mediated apoptosis by methylseleninic
acid in LNCaP prostate cancer cells. Carcinogenesis. 26:1374–1381.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang C, Jette N, Moussienko D, Bebb DG and
Lees-Miller SP: ATM-deficient colorectal cancer cells are sensitive
to the PARP inhibitor olaparib. Transl Oncol. 10:190–196. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gross-Goupil M, Massard C and Ravaud A:
Targeted therapies in metastatic renal cell carcinoma: Overview of
the past year. Curr Urol Rep. 13:16–23. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mrozik KM, Blaschuk OW, Cheong CM,
Zannettino AC and Vandyke K: N-cadherin in cancer metastasis, its
emerging role in haematological malignancies and potential as a
therapeutic target in cancer. BMC Cancer. 18:9392018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pal I, Rajesh Y, Banik P, Dey G, Dey KK,
Bharti R, Naskar D, Chakraborty S, Ghosh SK, Das SK, et al:
Prevention of epithelial to mesenchymal transition in colorectal
carcinoma by regulation of the E-cadherin-β-catenin-vinculin axis.
Cancer Lett. 452:254–263. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tania M, Khan MA and Fu J: Epithelial to
mesenchymal transition inducing transcription factors and
metastatic cancer. Tumour Biol. 35:7335–7342. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gao H, Niu W, He Z, Gao C, Peng C and Niu
J: SEC61G plays an oncogenic role in hepatocellular carcinoma
cells. Cell Cycle. 19:3348–3361. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liao HJ and Carpenter G: Role of the Sec61
translocon in EGF receptor trafficking to the nucleus and gene
expression. Mol Biol Cell. 18:1064–1072. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sorkin A: Internalization of the epidermal
growth factor receptor: Role in signalling. Biochem Soc Trans.
29:480–484. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hu M, Zhu S, Xiong S, Xue X and Zhou X:
MicroRNAs and the PTEN/PI3K/Akt pathway in gastric cancer (Review).
Oncol Rep. 41:1439–1454. 2019.PubMed/NCBI
|
|
39
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Friedl P and Wolf K: Tumor-cell invasion
and migration: Diversity and escape mechanisms. Nat Rev Cancer.
3:362–374. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yagi N, Manabe I, Tottori T, Ishihara A,
Ogata F, Kim JH, Nishimura S, Fujiu K, Oishi Y, Itaka K, et al: A
nanoparticle system specifically designed to deliver short
interfering RNA inhibits tumor growth in vivo. Cancer Res.
69:6531–6538. 2009. View Article : Google Scholar : PubMed/NCBI
|