Introduction
Ozone therapy is widely used in clinics to treat
herniated lumbar discs and osteoarthritis (1,2) due to
its effectiveness and convenient application. However, certain
studies have shown that ozone overdose may induce an
oxidant-antioxidant imbalance, resulting in damage to the nervous
system (3,4). The nervous system is sensitive to this
imbalance due to its high demand for oxygen. The spinal cord may be
preferentially damaged in cases where ozone overdose occurs during
ozone therapy for lumbar disc herniation (5). However, the mechanism by which ozone
overdose damages spinal cord neurons (SCNs) is poorly understood
(6). Nonetheless, it has been shown
that ozone-induced neurotoxicity is associated with its strong
oxidizing properties (7).
Liu et al (4)
reported that ozone activates endoplasmic reticulum stress-mediated
Ca2+ release and the calmodulin-dependent protein kinase
II/MAPK signaling pathway to produce neurotoxicity in SCNs. Both
autophagy and the nuclear factor (erythroid-derived-2)-related 2
factor/antioxidant response element (NRF2/ARE) signaling pathway
are key contributors to the antioxidant system. Kelch-like
ECH-associated protein 1 (KEAP1) associates with and negatively
regulates NRF2 (8,9). This interaction may be disrupted by
ubiquitin-binding protein p62, which promotes degradation of KEAP1
and maintains levels of NRF2. Through a positive feedback
mechanism, NRF2 activates p62 to maintain redox homeostasis
(10). Moreover, autophagy and the
NRF2/ARE antioxidant pathway influence formation, injury, repair,
degeneration and aging of the nervous system (11–17).
Tert-butylhydroquinone (tBHQ), an activator of NRF2, has been used
to investigate the function of NRF2 in inflammation, antioxidative
stress response, autophagy and neuroprotection (18–21).
However, there is a lack of evidence regarding the function of tBHQ
in SCN injury by ozone overdose. In the present study, the Cell
Counting Kit 8 (CCK8) was used to detect cell viability to identify
an appropriate concentration of medical ozone for subsequent
experiments. Reverse transcription-quantitative PCR (RT-qPCR) and
western blotting were performed to detect alterations in the mRNA
and protein expression levels of NRF2, heme oxygenase 1 (HO-1), P62
and LC3. Immunofluorescence assays were conducted to detect the
expression of nuclear NRF2. The Normal/Apoptotic/Necrotic Cell
Detection Kit (AO/EB) was used to detect the rate of live,
apoptotic and necrotic cells. In the present study, tBHQ was used
to activate NRF2 to investigate the role of the p62/NRF2/ARE
pathway in SCN injury caused by ozone overdose.
Materials and methods
Reagents, chemicals and
antibodies
Neonatal Wistar rats were purchased from the
Experimental Animal Center of Shandong University. All programs
involving the use of animals were performed in accordance with the
Guide for the Care and Use of Laboratory Animals of the National
Institutes of Health (NIH; olaw.nih.gov/sites/default/files/Guide-for-the-Care-and-Use-of-Laboratory-Animals.pdf)
and were approved by the Animal Care and Use Committee of the
School of Medicine of Shandong University (Shandong, China).
Poly-L-lysine hydrobromide (PLL; Beijing Solarbio Science &
Technology Co., Ltd.), FBS (Biological Industries), DMEM-High
Glucose (HG; Invitrogen; Thermo Fisher Scientific, Inc.), B27
(Invitrogen; Thermo Fisher Scientific, Inc.), L-glutamine (Gln;
Beijing Solarbio Science & Technology Co., Ltd.), Neurobasal-A
medium (Thermo Fisher Scientific, Inc.), penicillin-streptomycin
(Beijing Solarbio Science & Technology Co., Ltd.), PBS (Gibco;
Thermo Fisher Scientific, Inc.), Tyrisin (Gibco; Thermo Fisher
Scientific, Inc.), 4% paraformaldehyde (Beijing Solarbio Science
& Technology Co., Ltd.), and normal goat serum (Abbkine
Scientific Co., Ltd.) were used to extract and cultivate primary
neuronal cells. The antibodies included anti-MAP2 antibody (rabbit
anti-rat IgG; cat. no. 17490-1-AP; ProteinTech Group, Inc.),
Dylight 594 goat anti-rabbit IgG (H+L; cat. no. A23420; Abbkine
Scientific Co., Ltd.), Dylight 488 goat anti-rabbit IgG (H+L; cat.
no. A23220; Abbkine Scientific Co., Ltd.), anti-NRF2 NFE2L2 (rabbit
anti-rat; cat. no. 16396-1-AP; ProteinTech Group, Inc.), anti-GAPDH
(EPR16891 rabbit anti-rat; cat. no. ab181602; Abcam), anti-light
chain (LC)3B (EPR18709 rabbit anti-rat; cat. no. ab192890; Abcam),
anti-sequestosome (SQSTM)1/p62 (EPR4844 rabbit anti-rat; cat. no.
ab109012; Abcam), anti-HO-1 (EPR1390Y rabbit anti-rat; cat. no.
ab68477; Abcam) and horseradish peroxidase (HRP)-goat anti-rabbit
IgG (H+L; cat. no. SA00001-2; ProteinTech Group, Inc.). Other
reagents and kits included Fluorescent Mounting Media (cat. no.
S2100-5; Beijing Solarbio Science & Technology Co., Ltd.), CCK8
(Abcam), PMSF (cat. no. P1260-1; Beijing Solarbio Science &
Technology Co., Ltd.), AG RNAex Pro Reagent (Accurate
Biotechnology; cat. no. AG21101), Custom DNA/RNA oligos (BioSune;
cat. no. 5002), Pro Taq HS SYBR Green quantitative (q)PCR kit (AG,
cat. no. AG11701), Evo M-MLV RT Premix for qPCR (AG, cat. no.
AG11706), tBHQ (MedChemExpress; cat. no. HY-100489), bicinchoninic
acid (BCA) protein assay kit (cat. no. PC0020-500; Beijing Solarbio
Science & Technology Co., Ltd.), enhanced chemiluminescence
(ECL) substrate (cat. no. WBKLS0100; Merck KGaA) and the
Normal/Apoptotic/Necrotic Cell Detection kit (cat. no. KGA501;
Nanjing KeyGen Biotech Co., Ltd.).
Primary culture of newborn Wistar rat
neurons
A total of 400 Wistar rats (weight, 4–6 g; age,
<24 h) were euthanized by decapitation and dehydrated by soaking
in 75% alcohol for 3 min at 0°C. The spinal cord was removed and
sectioned into 0.5–1 mm3 pieces. The sections were
incubated in 3 ml 0.125% trypsin for 15 min at 37°C. The digestion
was terminated by incubating the sections in isopycnic FBS for 3
min at room temperature, after which the liquid supernatant was
removed. A temporary medium, composed of FBS, DMEM-HG and
penicillin-streptomycin, was added, followed by mixing and
agitating the solution 15 times with a pipette. The liquid
supernatant was aspirated into a new centrifuge tube following 2
min incubation at room temperature; this step was repeated twice.
The supernatant was filtered and the cells were counted using a
BX51 light microscope (magnification, ×400; Olympus
Corporation).
The cells were plated onto a culture plate
(1×107/cm2) that had been coated 3 times with
PLL. The cells were incubated for 4 h at 37°C, after which the
temporary medium was removed. PBS was used to wash the plate 2–3
times and medium was replaced with complete medium (97.5
Neurobasal-A medium, 1.0 B27, 0.5 Gln and 1.0%
penicillin-streptomycin) for incubation at 37°C. During the
culture, half of the culture medium was changed every 3 days; on
the seventh day, cells were harvested for subsequent
experiments.
Immunofluorescence assay
In order to authenticate the purity of cell culture,
cells were cultured for 7 days. The cells were washed 3 times (5
min each) with PBS and immobilized by incubation in 4%
paraformaldehyde for 30 min at room temperature. After
immobilization, the cells were washed 3 times (5 min each) with PBS
again and blocked with 10% goat serum for 1 h in an incubator at
37°C. Subsequently, cells were washed 3 times (5 min each) to
remove the goat serum. Anti-MAP2 monoclonal antibody (1:300)
attenuated by goat serum was added, followed by overnight
incubation in a refrigerator at 4°C. The following day, cells were
washed 3 times (5 min each) with PBS, then Dylight 594 Goat
anti-rabbit IgG (H+L; 1:200) attenuated by 100% goat serum was
added, followed by incubation at room temperature for 1 h. The
secondary antibody was removed by washing the cells 3 times (5 min
each) with PBS.DAPI (10 µg/ml) was added to the cells and incubated
for 10 min at room temperature, followed by washing with PBS to
remove excess DAPI. A fluorescent mounting media (cat. no. S2100-5;
Beijing Solarbio Science & Technology Co., Ltd.) was added, and
the cells were observed under a confocal immunofluorescence
microscope (magnification, ×400). The same procedure was performed
with anti-NRF2 NFE2L2 (1:300) and Dylight 488 Goat anti-rabbit IgG
(H+L) (1:200) to compare expression levels of NRF2 in the
nucleus.
CCK8 assay
In order to determine the appropriate concentration
of medical ozone for future assays, the cells were cultured in a
96-well plate (100 µl/well) for 7 days. Different concentrations of
medical ozone (0, 10, 20, 30 and 40 µg/ml) were administered to the
cells for 30 min at room temperature. Following the manufacturer's
protocol, 10 µl CCK8 solution was added to each well. The plate was
incubated at 37°C for 4 h and the absorbance was measured at 450
nm.
RT-qPCR analysis
Total RNA was extracted from SCNs using AG RNAex Pro
Reagent according to the manufacturer's protocol. The sample
concentration was subsequently measured and Evo M-MLV RT Premix for
qPCR (AG) was used to synthesize cDNA (500 ng total RNA per 10 µl
reaction mixture) according to the manufacturer's instructions. The
Pro Taq HS SYBR Green qPCR kit was used for RT-qPCR to quantify RNA
on a Roche LightCycler480 system. The following thermocycling
conditions were used for qPCR: 95°C for 30 sec; followed by 40
cycles of 95°C for 5 sec and 60°C for 30 sec. β-actin was used as
an internal control. The 2−ΔΔCq method (22) was used to calculate the fold changes
in gene expression levels. The primer sequences were as follows:
β-actin forward, 5′-CTCTGTGTGGATTGGTGGCT-3′ and reverse,
5′-CGCAGCTCAGTAACAGTCCG-3′; NRF2 forward,
5′-CCATTTACGGAGACCCACCG-3′ and reverse, 5′-TTTGACACTTCCAGGGGCAC-3′;
p62 forward, 5′-CTGAGAAGGACTCGCTCGAC-3′ and reverse,
5′-TCAGTACCCGCTCTTTCAGC-3′; HO-1 forward,
5′-GAGCGAAACAAGCAGAACCC-3′ and reverse, 5′-ACCTCGTGGAGACGCTTTAC-3′
and LC3, forward, 5′-TTGGTCAAGATCATCCGGCG-3′ and reverse,
5′-GTCAGCGATGGGTGTGGATA-3′.
Western blotting
SCNs were lysed using RIPA lysis buffer with PMSF
and phosphatase inhibitor (PI; RIPA:PMSF:PI=100:1:1), stored on ice
for 30 min and vortexed every 5 min. The supernatant was collected
following centrifugation at 16,000 × g for 30 min at 4°C. The BCA
protein assay kit was used to measure the total protein
concentration. Sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (12%) was used to separate the proteins (30
µg/lane). After loading the samples, the gel was pre-run at 75 V
for 45 min and then at 110 V for ~90 min. The protein was
transferred to a polyvinylidene fluoride membrane via
electrotransformation at 220 mA (1 min per molecular weight). The
membranes were blocked in 5% skimmed milk for 60 min at room
temperature, followed by 3 washes for 5 min each in TBST (1%
Tween-20). The membranes were then incubated overnight with primary
anti-NRF2 NFE2L2 (1:2,000), anti-GAPDH (1:5,000), anti-LC3B
(1:5,000), anti-SQSTM1/p62 (1:10,000) and anti-HO-1 antibody
(1:2,000) at 4°C. The primary antibody was removed by washing the
membranes three times with TBST (10 min each). The membranes were
then incubated for 1 h at room temperature with HRP-Goat
anti-rabbit IgG (H+L; 1:5,000) antibody, followed by three 10-min
washes. The membranes were developed using ECL substrates for
detection. The band densities were quantified by ImageJ software
(version 1.51; NIH).
Acridine orange/ethidium bromide
(AO/EB) assay
The Normal/Apoptotic/Necrotic Cell Detection kit was
used to calculate the necrosis and apoptosis rates. SCNs were
extracted and cultured for 7 days. SCNs were subsequently subjected
to digestion (0.125% trypsin for 10 min), centrifugation (72 × g
for 4 min at room temperature) and resuspension in PBS
(1×106 cells/ml). Mixed dye reagent (1 µl dye reagent
1:dye reagent 2=1:1) was added to 25 µl cells and mixed thoroughly.
A total of 10 µl mixture was dropped on glass slides and covered
with a cover slip. A total of 200 cells were observed under a
confocal immunofluorescence microscope at ×100 magnification and
necrosis and apoptosis rates were calculated.
Statistical analysis
SPSS (version 23.0; IBM Corp.) was used for
statistical analysis. The data are expressed as the mean ± SD
(n≥3). Data were analyzed by one-way ANOVA, followed by Tukey's
post hoc test for multiple comparisons. P<0.05 was considered to
indicate a statistically significant difference.
Results
Primary neuron authentication
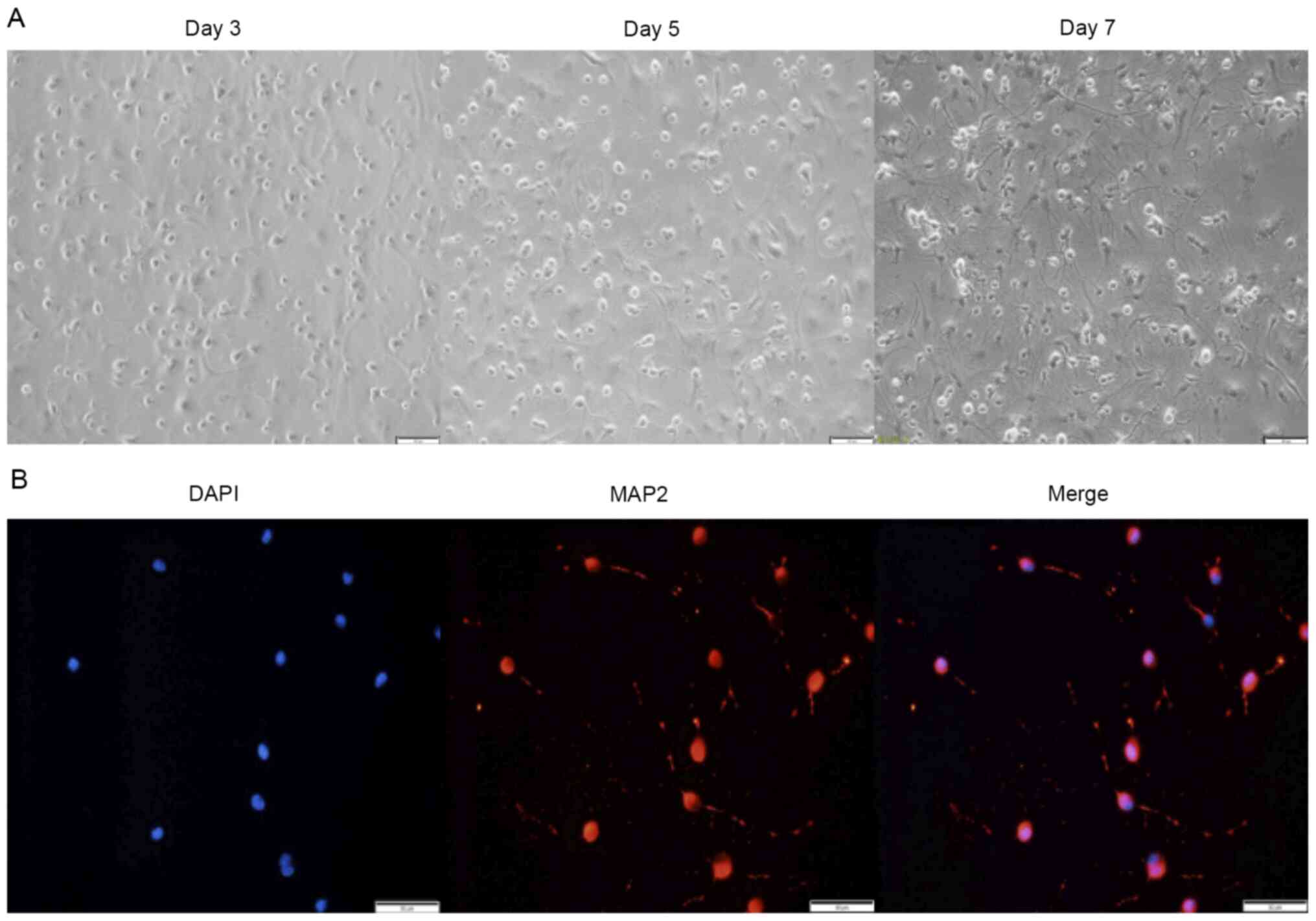
MAP2 was used as a specific neuronal marker to
verify the purity of the primary neuron extraction (Fig. 1A and B). The results showed purity
>90% (calculated according to the formula: MAP2 staining/DAPI
staining), which was considered acceptable for use in subsequent
experiments.
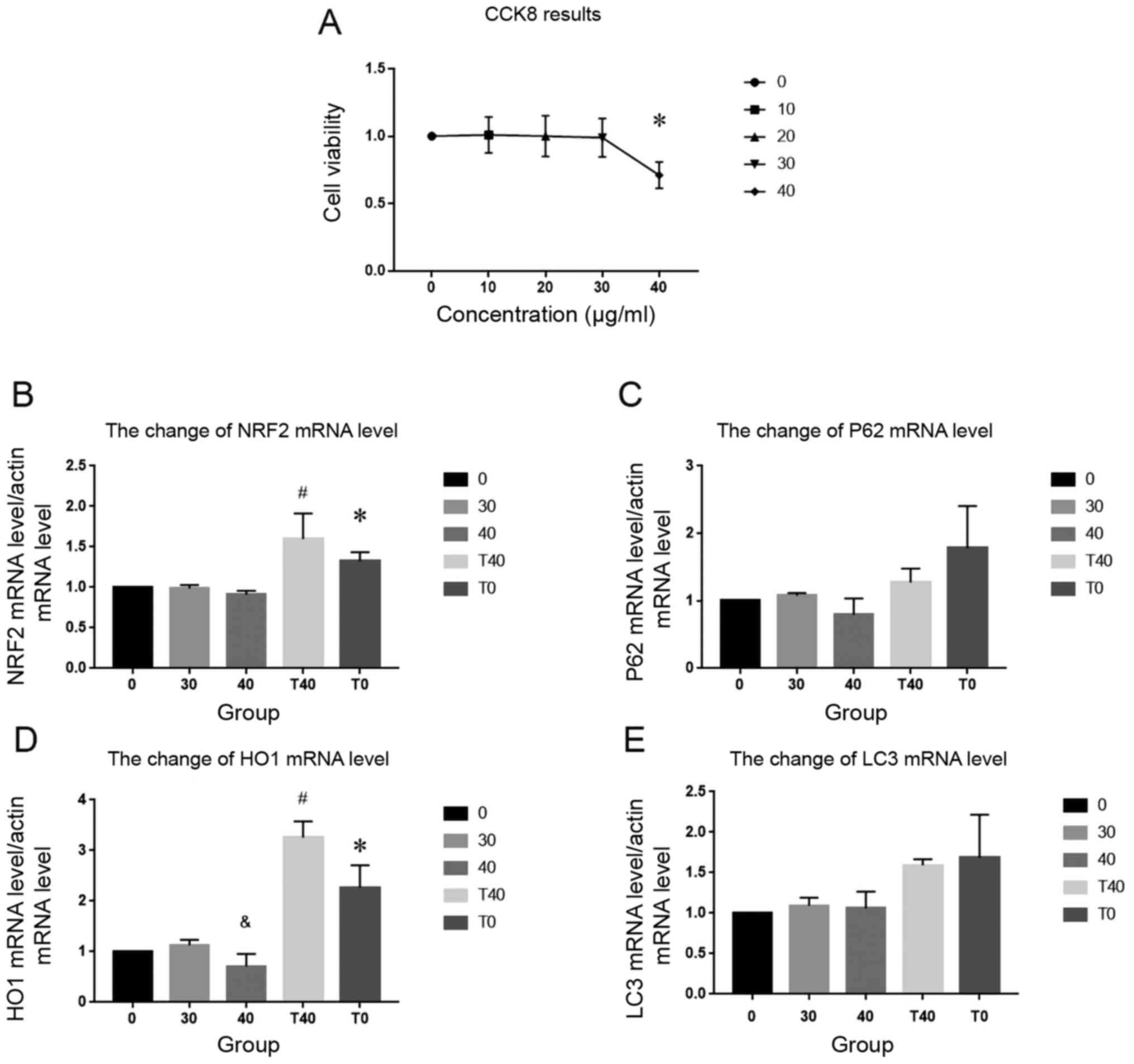
Medical ozone at 40 µg/ml impacts cell
viability
CCK8 assay was performed to determine assess cell
viability and determine an appropriate experimental concentration
of ozone for subsequent experiments. No notable change in
absorbance at 520 nm was observed in cells treated with 10–30 µg/ml
ozone. However, at 40 µg/ml, absorbance at 520 nm decreased
significantly (Fig. 2A). Thus, 40
µg/ml ozone was selected for subsequent experiments.
Changes in mRNA levels of NRF2 and
HO-1 following ozone and tBHQ treatment
RT-qPCR was used to assess the relative changes in
mRNA levels of genes involved in redox homeostasis. No
statistically significant difference was observed in the mRNA
expression levels of p62 and LC3II following treatment with 40
µg/ml ozone compared with the control group (Fig. 2C and E). However, mRNA levels of
NRF2 and HO-1 were increased in the T0 group [40 µmol/l tBHQ
treatment] compared with the control group, which indicated that
tBHQ activated the NRF2 signaling pathway. The concentration of
tBHQ selected in the present study was based on previous studies
(23,24). Furthermore, the levels of NRF2
increased more in the T40 than in the 40 µg/ml ozone group; levels
of HO-1 were lower in the 40 µg/ml ozone group compared with those
in the 30 µg/ml ozone group. The addition of tBHQ in the T40 group
did not result in a similar decrease in HO-1 (Fig. 2B and D).
High ozone dose alters expression
levels of proteins in the NRF2/ARE pathway
Protein levels of NRF2 and HO-1 were assessed to
determine changes in the NRF2/ARE pathway following ozone
administration. Protein levels of NRF2 and HO-1 were increased in
the 30 µg/ml ozone group compared with the 0 µg/ml ozone group, and
HO-1 levels were decreased in the 40 and 30 µg/ml ozone groups.
Conversely, treatment with tBHQ in the T40 group reversed the
changes in protein expression levels observed in the 40 µg/ml ozone
group (Fig. 3A and C).
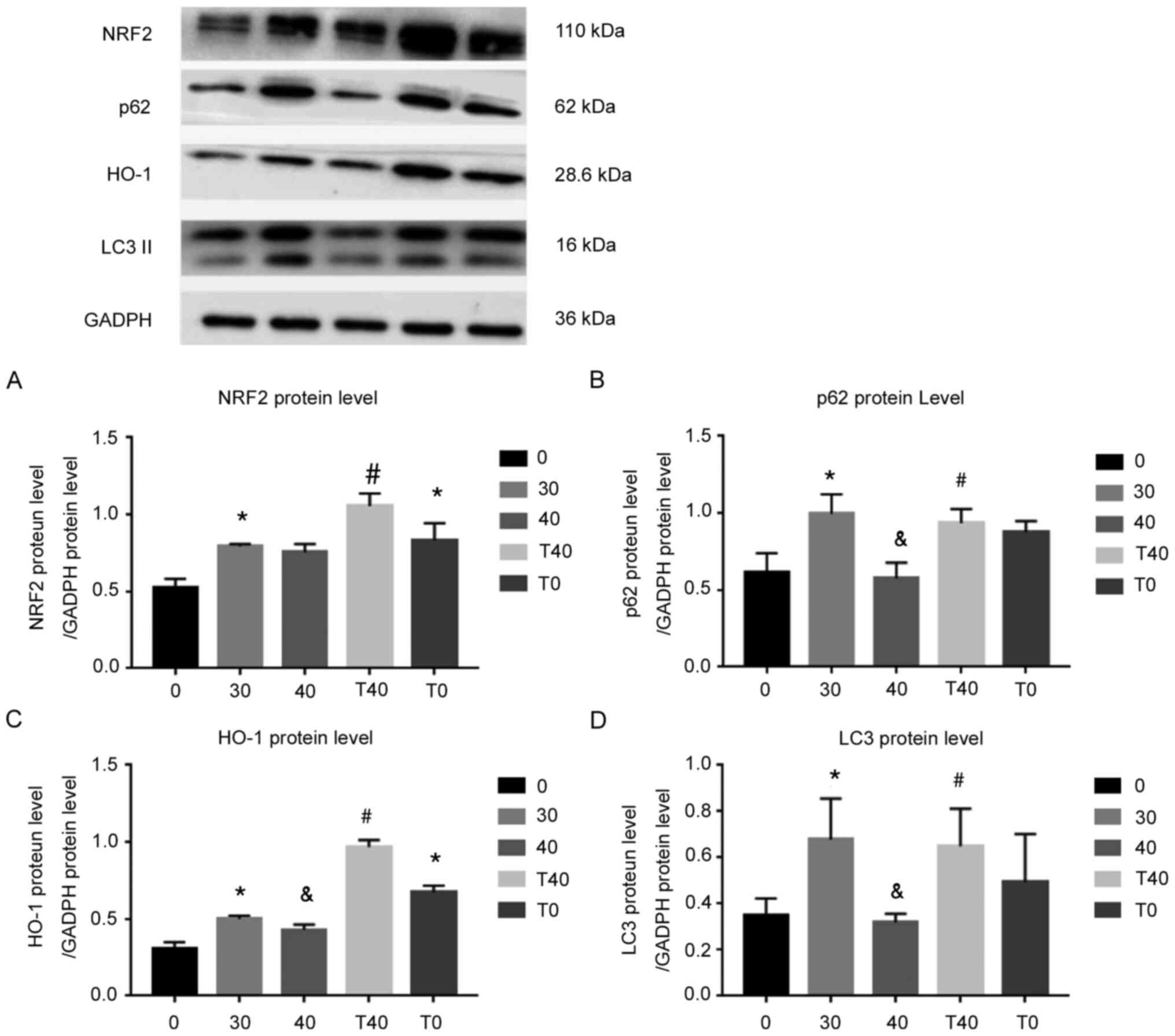 | Figure 3.Changes in protein levels of NRF2,
p62, HO-1 and LC3. Changes in (A) NRF2, (B) p62, (C) HO-1 and (D)
LC3 protein levels following different treatments. GADPH was used
as the internal control. All experiments were performed in
triplicate (n=3). *P<0.05 vs. 0 µg/ml ozone;
&P<0.05 vs. 30 µg/ml ozone; #P<0.05
vs. 40 µg/ml ozone. T40, 40 µmol/l tBHQ + 40 µg/ml ozone; T0, 40
µmol/l tBHQ; NRF2, nuclear factor (erythroid-derived-2)-related 2;
HO-1, heme oxygenase 1; LC3, light chain 3; tBHQ,
tert-butylhydroquinone. |
Protein levels of both p62 and LC3II increased in
the 30 µg/ml ozone group, compared with the 0 µg/ml ozone group,
and decreased in the 40 µg/ml ozone group compared with the 30
µg/ml ozone group. Moreover, tBHQ reversed the downregulation of
p62 and LC3II in the 40 µg/ml ozone group (Fig. 3B and D). Certain reports have shown
that p62 can form a positive feedback loop with KEAP1 and NRF2
(9,25), and that expression levels of p62 and
LC3II responds to the level of autophagy.
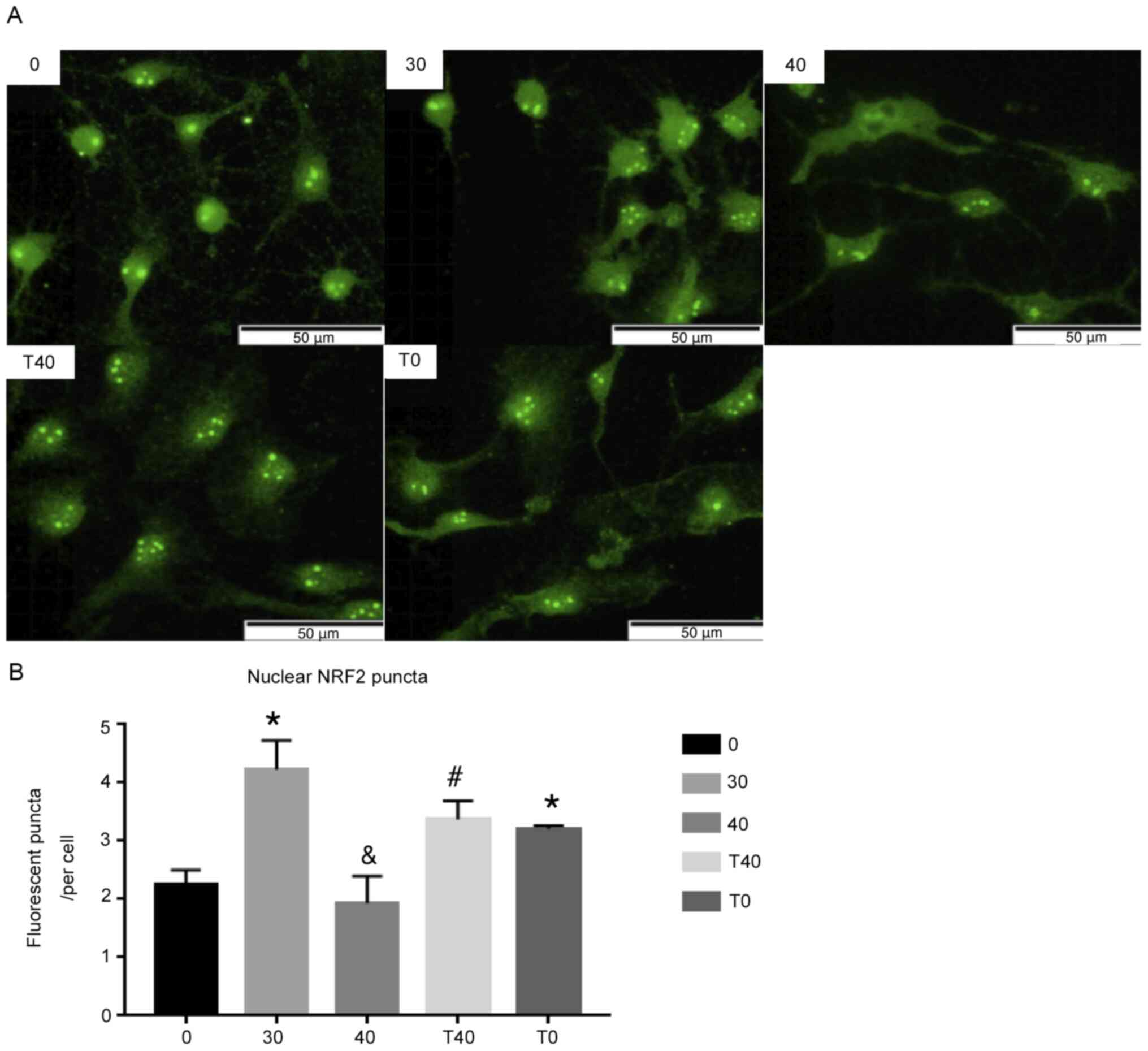
NRF2 localizes to the nucleus and
increases in the 30 µg/ml ozone, T40 and T0 groups
Cytoplasmic NRF2 localizes to the nucleus,
activating ARE and increasing antioxidant levels (26). NRF2 levels in the nucleus were
detected by immunofluorescence, which showed that nuclear NRF2
increased in the 30 µg/ml ozone group compared with the 0 µg/ml
ozone group. However, the 40 µg/ml ozone group exhibited less
nuclear NRF2 than the 30 µg/ml ozone group. Treatment with tBHQ in
the T40 group reversed this decrease in nuclear NRF2 (Fig. 4A and B).
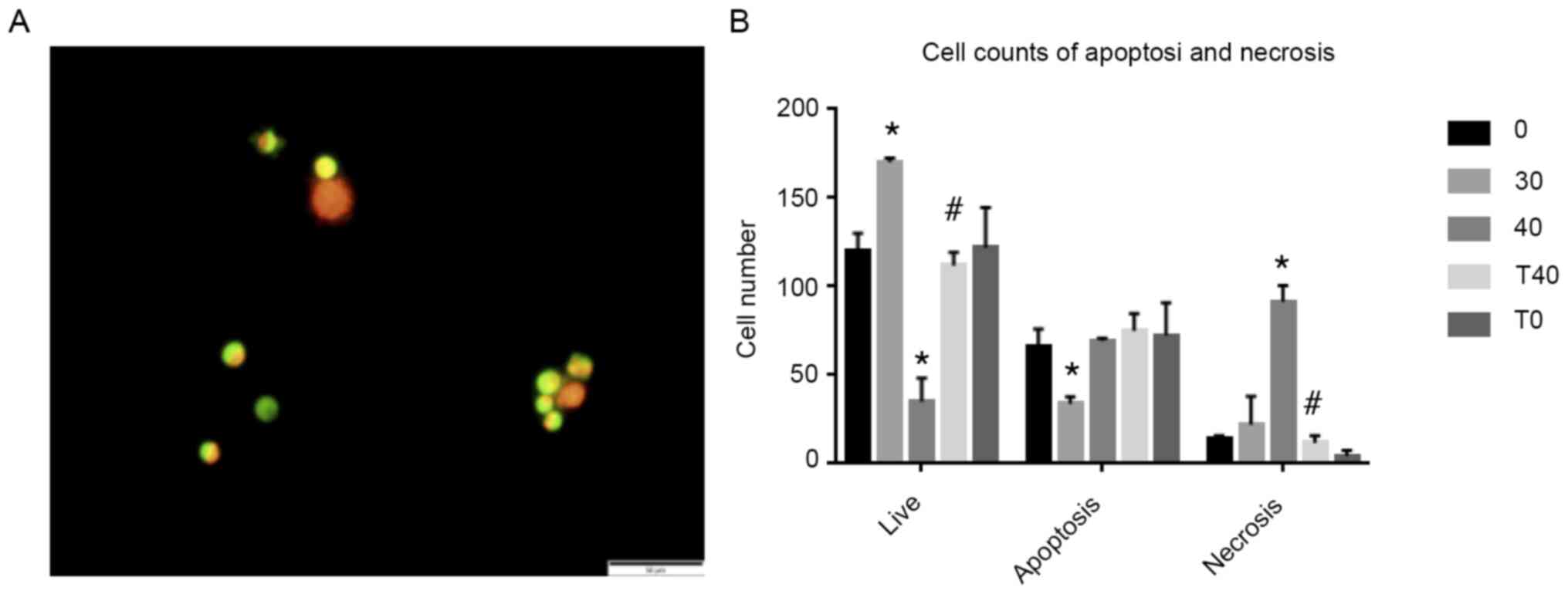
tBHQ reverses ozone-induced cell
death
CCK8 assay showed that 30 min ozone therapy at 40
µg/ml decreased cell viability by 71%. AO/EB assay was used to
verify whether tBHQ prevent and SCN death caused by 40 µg/ml ozone
treatment (Fig. 5A). Confirming the
findings of the CCK8 assay, the results showed a decrease in live
cells and an increase in necrotic cells in the 40 µg/ml ozone
(Fig. 5B). Conversely, a decrease
in necrosis and increase in live cells was observed in the T40
group compared with the 40 µg/ml ozone group.
Discussion
The effects of ozone, a gas with strong oxidizing
properties, are concentration-dependent (4). Overuse of ozone can affect redox
homeostasis and the central nervous system is easily influenced by
ozone due to its high oxygen requirement. SCNs were here extracted
from newborn Wistar rats and subjected to different ozone
concentrations. Similar to a previous report, 40 µg/ml ozone
injured the neurons in vitro (4).
Here, 40 µg/ml ozone induced death of SCNs in
vitro, and mRNA and protein levels of HO-1 decreased with
higher concentrations of ozone (30 and 40 µg/ml). Moreover, protein
levels of NRF2, p62 and LC3II and expression of nuclear NRF2
decreased in the 40 µg/ml ozone group compared with the 30 µg/ml
ozone group. Furthermore, tBHQ prevented this decrease in the 40
µg/ml ozone group.
Ozone has a strong oxygen-saturation capacity, and
medical ozone is a mixture of ozone and oxygen with over 100 years
of clinical use. It can interact with organic macromolecules to
induce the production of reactive oxygen species (ROS) and lipid
oxidation products (LOPS) to treat disease (27). However, the antioxidant system is
damaged if excessive ozone concentrations imbalance ROS and LOPS
(28). Previously, several pathways
have been implicated in redox homeostasis (29,30).
Reports have shown that the KEAP1/NRF2/ARE pathway is important to
counteracting oxidative stress (31) and that autophagy is sensitive to
oxidative stress (32). p62, a key
protein in autophagy, also regulates the activation of the
KEAP1/NRF2/ARE pathway (33). Based
on these studies, tBHQ (an activator of NRF2), was used to
investigate the role of the p62/KEAP1/NRF2/ARE pathway in
responding to the damage caused to SCNs by ozone overdose.
The NRF2/ARE pathway is a key pathway in
antioxidative stress signaling (31). It participates in all processes of
the neuron system (13–15) and is critical in supporting the
normal activity of the nervous system. In the present study, 40
µg/ml ozone decreased the mRNA levels of HO-1, and similar
decreases in protein expression levels of NRF2 and HO-1 were
observed. Similarly, p62 protein levels decreased with ozone
treatment. Moreover, NRF2 localizes to the nucleus to activate ARE
and enhance antioxidant capacity (33); the present study showed that
treatment with ozone increased expression levels of nuclear NRF2.
tBHQ reversed the changes in protein and mRNA expression levels
observed due to administration of 40 µg/ml ozone. Following
treatment with tBHQ and 40 µg/ml ozone, nuclear NRF2 expression
levels and necrosis rate decreased. These results showed that ozone
overdose induced autophagy in SCNs by inhibiting the activity of
NRF2/ARE, and that tBHQ activated NRF2 to protect SCNs.
Autophagy is the major intracellular degradation
system that is crucial in protecting cells from injury (34) and serves a key role in nervous
system disease (9–12). p62 is a key protein in autophagy
and, together with LC3II, reflects autophagy in cells (35). Conversely, p62 and NRF2 form a
positive feedback loop to enhance antioxidant capacity to protect
cells (8). On this basis, changes
in p62 and LC3II were measured. Previous studies have demonstrated
that autophagic capacity is enhanced by increased LC3II and
decreased p62 (36–38). Conversely, autophagic capacity
decreases if the reverse result is obtained (39). No changes were observed in the mRNA
levels of p62 and LC3II, and decreases in p62 and LC3II protein
levels were only observed following 40 µg/ml ozone treatment; tBHQ
reversed this decrease in p62 and LC3II protein levels. Moreover,
tBHQ increased the expression levels of LC3II, thus showing that
autophagic flux was activated. However, expression of p62 also
increased. This may have been due to simultaneous activation of
autophagy (40) and the NRF2/ARE
pathway (19) by ozone in
combination with tBHQ treatment. This positive feedback increased
the expression of p62 observed in the present study. Thus, it was
concluded that tBHQ activates autophagy and the p62/NRF2/ARE
pathway to increase nuclear NRF2 localization and enhance the
antioxidant system, which may protect cells from injury caused by
ozone overdose.
Certain limitations exist in the present study. For
example, the mechanism proposed should be further scrutinized by
arresting the rise in NRF2 following tBHQ treatment. Additionally,
other autophagy signaling pathways, such as PI3K/Akt/mTOR, should
be investigated for potential interactions with p62 and LC3II.
Other apoptotic proteins, such as cytochrome c, activating
transcription factor, Bax, Bid and caspase-3/9, should also be
assessed to clarify the mechanism underlying ozone and the
protective effect of tBHQ. Finally, the mechanism proposed here
needs to be tested in vivo to identify clinical
applications.
The present study proposed a potential mechanism of
SCN injury caused by ozone overdose. The results suggest that 40
µg/ml ozone injured SCNs in vitro. Furthermore, 40 µg/ml
ozone disrupted the p62/NRF2/ARE pathway and disturbed NRF2 nuclear
transfer to injure SCNs. While this study presents in vitro
data about the protective role of nuclear NRF2 localization, data
from in vivo studies are still required.
Acknowledgements
Not applicable.
Funding
The present study was funded by the National Natural
Science Foundation of China (grant nos. 81271346 and 81771199).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZF, CZ and XZ conceptualized and designed the
experiments. CZ and XZ performed the experiments. BW, PS and SM
performed data analysis. CZ and BW wrote the manuscript. CZ and ZF
confirm the authenticity of all the raw data. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Care and Use Committee of Shandong Provincial Hospital Affiliated
to Shandong University (approval no. 2020-663).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ARE
|
antioxidant response element
|
|
NRF2
|
nuclear factor
(erythroid-derived-2)-related 2 factor
|
|
KEAP1
|
Kelch-like ECH-associated protein
1
|
|
tBHQ
|
Tert-butylhydroquinone
|
|
HG
|
High Glucose
|
|
Gln
|
L-glutamine
|
|
PLL
|
Poly-L-lysine hydrobromide
|
|
SCN
|
Spinal cord neuron
|
|
HO-1
|
heme oxygenase1
|
|
CCK8
|
Cell Counting Kit-8
|
|
PI
|
phosphatase inhibitor
|
References
|
1
|
Anuj B, Peter M, Donald L, Elias G and
Murphy K: Percutaneous ozone treatment for herniated lumbar discs:
1-year follow-up of a multicenter pilot study of a handheld
disposable ozone-generating device. J Vasc Interv Radiol.
30:752–760. 2019. View Article : Google Scholar
|
|
2
|
Silva Júnior JIS, Rahal SC, Santos IFC,
Martins DJC, Michelon F, Mamprim MJ, Tomacheuski RM and Correia
LECS: Use of reticulated hyaluronic acid alone or associated with
ozone gas in the treatment of osteoarthritis due to hip dysplasia
in dogs. Front Vet Sci. 7:2652020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ginanneschi F, Cervelli C, Milani P and
Rossi A: Ventral and dorsal root injury after oxygen-ozone therapy
for lumbar disk herniation. Surg Neurol. 66:619–620. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu H, Wang Y, An J, Williams J and Cope
D: Thunderclap headache caused by an inadvertent epidural puncture
during oxygen-ozone therapy for patient with cervical disc
herniation. Chin Med J (Engl). 129:498–499. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Özugur S, Kunz L and Straka H:
Relationship between oxygen consumption and neuronal activity in a
defined neural circuit. BMC Biol. 18:762020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li Y, Lin X, Zhao X, Xie J, JunNan W, Sun
T and Fu Z: Ozone (O3) elicits neurotoxicity in spinal cord neurons
(SCNs) by inducing ER Ca(2+) release and activating the CaMKII/MAPK
signaling pathway. Toxicol Appl Pharmacol. 280:493–501. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rivas-Arancibia S, Guevara-Guzmán R,
López-Vidal Y, Rodríguez-Martínez E, Zanardo-Gomes M, Angoa-Pérez M
and Raisman-Vozari R: Oxidative stress caused by ozone exposure
induces loss of brain repair in the hippocampus of adult rats.
Toxicol Sci. 113:187–197. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jiang G, Xue L, Huang Y, Lan Z, Zhang Z,
Su Z, Fang Z, Lai Y, Yao W, Ting L, et al: p62 promotes
proliferation, apoptosis-resistance and invasion of prostate cancer
cells through the Keap1/Nrf2/ARE axis. Oncol Rep. 43:1547–1557.
2020.PubMed/NCBI
|
|
9
|
Li T, Jiang D and Wu K: p62 promotes
bladder cancer cell growth by activating KEAP1/NRF2-dependent
antioxidative response. Cancer Sci. 111:1156–1164. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liao W, Wang Z, Fu Z, Ma H, Jiang M, Xu A
and Zhang W: p62/SQSTM1 protects against cisplatin-induced
oxidative stress in kidneys by mediating the cross talk between
autophagy and the Keap1-Nrf2 signalling pathway. Free Radic Res.
53:800–814. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yuan B, Shen H, Lin L, Su T, Zhong L and
Yang ZJ: Autophagy promotes microglia activation through
beclin-1-atg5 pathway in intracerebral hemorrhage. Mol Neurobiol.
54:115–124. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lipton J and Sahin MJN: The neurology of
mTOR. Neuron. 84:275–291. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sepe S, Nardacci R, Fanelli F, Rosso P,
Bernardi C, Cecconi F, Mastroberardino PG, Piacentini M and Moreno
S: Expression of Ambra1 in mouse brain during physiological and
Alzheimer type aging. Neurobiol Aging. 35:96–108. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou Z, Chen S, Zhao H, Wang C, Gao K, Guo
Y, Shen Z, Wang Y, Wang H and Mei X: Probucol inhibits neural cell
apoptosis via inhibition of mTOR signaling pathway after spinal
cord injury. Neuroscience. 329:193–200. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li D, Tian H, Li X, Mao L, Zhao X, Lin J,
Lin S, Xu C, Liu Y, Guo Y and Mei X: Zinc promotes functional
recovery after spinal cord injury by activating Nrf2/HO-1 defense
pathway and inhibiting inflammation of NLRP3 in nerve cells. Life
Sci. 245:1173512020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wen C, Huang C, Yang M, Fan C, Li Q, Zhao
J, Gan D, Li A, Zhu L and Lu D: The secretion from bone marrow
mesenchymal stem cells pretreated with berberine rescues neurons
with oxidative damage through activation of the keap1-nrf2-ho-1
signaling pathway. Neurotox Res. 38:59–73. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu C, Hou B, He P, Ma P, Yang X, Yang X,
Zhang L, Qiang G, Li W and Du G: Neuroprotective Effect of
salvianolic acid a against diabetic peripheral neuropathy through
modulation of nrf2. Oxid Med Cell Longev. 2020:64314592020.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nna VU, Ujah GA, Suleiman JB, Mohamed M,
Nwokocha C, Akpan TJ, Ekuma HC, Fubara VV, Kekung-Asu CB and Osim
EE: Tert-butylhydroquinone preserve testicular steroidogenesis and
spermatogenesis in cisplatin-intoxicated rats by targeting
oxidative stress, inflammation and apoptosis. Toxicology.
441:1525282020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Meng X, Zhang C, Guo Y, Han Y, Wang C, Chu
H, Kong L, Ma H and Saso L: TBHQ attenuates neurotoxicity induced
by methamphetamine in the VTA through the Nrf2/HO-1 and PI3K/AKT
signaling pathways. Oxid Med Cell Longev. 2020:87871562020.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ratliff WA, Delic V, Pick CG and Citron
BA: Dendritic arbor complexity and spine density changes after
repetitive mild traumatic brain injury and neuroprotective
treatments. Brain Res. 1746:1470192020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Muri J, Wolleb H, Broz P, Carreira EM and
Kopf M: Electrophilic Nrf2 activators and itaconate inhibit
inflammation at low dose and promote IL-1β production and
inflammatory apoptosis at high dose. Redox Biol. 36:1016472020.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li H, Wu S, Shi N, Lin W, You J and Zhou
W: NF-E2-related factor 2 activation in PC12 cells: Its protective
role in manganese-induced damage. Arch Toxicol. 85:901–910. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu W, Li F, Xu Z, Sun B, Cao J and Liu Y:
Tert-butylhydroquinone protects PC12 cells against ferrous
sulfate-induced oxidative and inflammatory injury via the Nrf2_ARE
pathway. Chem Biol Interact. 273:28–36. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gureev A, Sadovnikova I, Starkov N,
Starkov A and Popov V: p62-Nrf2-p62 mitophagy regulatory loop as a
target for preventive therapy of neurodegenerative diseases. Brain
Sci. 10:8472020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jena KK, Kolapalli SP, Mehto S, Nath P,
Das B, Sahoo PK, Ahad A, Syed GH, Raghav SK, Senapati S, et al:
TRIM16 controls assembly and degradation of protein aggregates by
modulating the p62-NRF2 axis and autophagy. EMBO J. 37:e983582018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bocci V, Valacchi G, Corradeschi F,
Aldinucci C, Silvestri S, Paccagnini E and Gerli R: Studies on the
biological effects of ozone: 7. Generation of reactive oxygen
species (ROS) after exposure of human blood to ozone. J Biol Regul
Homeost Agents. 12:67–75. 1998.PubMed/NCBI
|
|
28
|
Sagai M and Bocci V: Mechanisms of action
involved in ozone therapy: Is healing induced via a mild oxidative
stress? Med Gas Res. 1:292011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jimenez-Moreno N and Lane JD: Autophagy
and redox homeostasis in Parkinson's: A crucial balancing Act. Oxid
Med Cell Longev. 2020:88656112020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ferino A, Rapozzi V and Xodo LE: The
ROS-KRAS-Nrf2 axis in the control of the redox homeostasis and the
intersection with survival-apoptosis pathways: Implications for
photodynamic therapy. J Photochem Photobiol B. 202:1116722020.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kensler TW, Wakabayashi N and Biswal S:
Cell survival responses to environmental stresses via the
Keap1-Nrf2-ARE pathway. Annu Rev Pharmacol Toxicol. 47:89–116.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Filomeni G, Zio DD and Cecconi F:
Oxidative stress and autophagy: The clash between damage and
metabolic needs. Cell Death Differ. 22:377–388. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu Q, Tan Y, Qu T, Zhang J, Duan X, Xu H,
Mu Y, Ma H and Wang F: Therapeutic mechanism of human neural stem
cell-derived extracellular vesicles against hypoxia-reperfusion
injury in vitro. Life Sci. 254:1177722020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mizushima N and Komatsu MJC: Autophagy:
Renovation of cells and tissues. Cell. 147:728–741. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim T, Lee S, Kim M, Cheon C and Ko SG:
Kaempferol induces autophagic cell death via IRE1-JNK-CHOP pathway
and inhibition of G9a in gastric cancer cells. Cell Death Dis.
9:8752018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kadowaki M and Karim MR: Cytosolic LC3
ratio as a quantitative index of macroautophagy. Methods Enzymol.
452:199–213. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Min Z, Ting Y, Mingtao G, Xiaofei T, Dong
Y, Chenguang Z and Wei D: Monitoring autophagic flux using
p62/SQSTM1 based luciferase reporters in glioma cells. Exp Cell
Res. 363:84–94. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kimura S, Fujita N, Noda T and Yoshimori
T: Monitoring autophagy in mammalian cultured cells through the
dynamics of LC3. Methods Enzymol. 452:1–12. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sheng Y, Chen X, Hou X, Yuan X, Yuan BS,
Yuan YQ, Zhang QL, Cao X, Liu CF, Luo WF and Hu LF: Urate promotes
SNCA/α-synuclein clearance via regulating mTOR-dependent
macroautophagy. Exp Neurol. 297:138–147. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li S, Li J, Shen C, Zhang X, Sun S, Cho M,
Sun C and Song Z: Tert-Butylhydroquinone (tBHQ) protects
hepatocytes against lipotoxicity via inducing autophagy
independently of Nrf2 activation. Biochim Biophys Acta. 1841:22–33.
2014. View Article : Google Scholar : PubMed/NCBI
|