Introduction
Metabolic dysfunction-associated fatty liver disease
(MAFLD), previously known as non-alcoholic fatty liver disease
(NAFLD), is the most globally prevalent liver disease linked to the
accumulation of fat in the liver (1–4). MAFLD
is characterized by histological lesions ranging from steatosis,
non-alcoholic steatohepatitis and fibrosis to cirrhosis and
hepatocellular carcinoma (5). MAFLD
develops as a consequence of improper lifestyle, such as unhealthy
diet, sedentary habits and excessive sugar intake, and threatens
the health of 25–30% of the global population (6,7). The
main treatment option for MAFLD is weight loss (8). There is no approved pharmacological
treatment for MAFLD, due to the complex and diverse pathogenesis of
this disease (9). Thus,
identification of new approaches for the treatment of MAFLD is
required.
Recently, traditional Chinese herbal extracts have
been shown to alleviate MAFLD (10). Parthenolide (PAR), an active
sesquiterpene lactone extracted from the feverfew plant, can
promote the apoptosis of various malignant cell types (11). In mice, PAR treatment can reduce
acute hepatitis and promote functional recovery of the liver
(12). PAR has also been reported
to protect against liver fibrosis (13). Furthermore, a previous study
indicated that PAR could represent a candidate agent for MAFLD
treatment (14). However, the
molecular mechanisms underlying the effects of PAR in MAFLD remain
unclear.
A previous study has demonstrated that the HIPPO
pathway regulates gene expression and that its mediators,
Yes-associated protein (YAP) and transcriptional coactivator with
PDZ-binding motif (TAZ), are associated with the development,
regeneration and tumorigenesis of the liver (15). The HIPPO pathway is associated with
hepatic fibrosis, due to the involvement of oxidative stress injury
and macrophage inflammation (16).
Additionally, the HIPPO pathway is associated with extracellular
matrix stiffness and hepatic fibrosis in hepatic stellate cells
(17). HIPPO/YAP signalling also
participates in the regulation of lipid metabolism (18). Moreover, the essential role of the
HIPPO pathway in the initiation and progression of MAFLD-associated
hepatocellular carcinoma has been demonstrated (19).
Therefore, considering the protective role of PAR
and involvement of the HIPPO pathway in several liver-associated
disorders, it was hypothesized that PAR could display similar
efficacy in MAFLD, possibly through the HIPPO pathway.
Consequently, histological and molecular experiments were carried
out to identify the regulatory mechanism governing the role of PAR
in MAFLD and to provide novel insight into therapeutic strategies
against MAFLD initiation and progression.
Materials and methods
Reagent preparation
PAR (Sigma-Aldrich; Merck KGaA) and verteporfin (VP)
(Selleck Chemicals) were dissolved in DMSO to prepare a 100-mM
solution, which was stored at −30°C and diluted to the required
concentration using 9% normal saline.
Animal treatment and grouping
C57BL/6 male mice (age, 8 weeks old; weight, 25–30
g) were purchased from Guangdong Medical Laboratory Animal Centre.
Mice were housed at 21±2°C and 50±5% relative humidity with a 12-h
day/night cycle and free access to food and drinking water. After
one week of adaptation, 144 mice were randomly allocated into 8
groups with 18 mice per group: i) Mice in the control group were
fed normal diet and intraperitoneally injected with 200 µl normal
saline containing 1.5% (v/v) DMSO three times a week; ii) mice in
the control + PAR group were fed normal diet and intraperitoneally
injected with 6 mg/kg PAR three times a week; iii) mice in the
model group were fed high-fat diet and intraperitoneally injected
with 200 µl normal saline containing 1.5% (v/v) DMSO three times a
week; iv) mice in the PAR-low (L), medium (M) and high (H) groups
were fed high-fat diet and intraperitoneally injected with 2, 4, or
6 mg/kg PAR three times a week (20,21);
v) mice in the PAR-VP group were fed high-fat diet and
intraperitoneally injected with 6 mg/kg PAR three times a week
followed by daily injections of 50 mg/kg VP five weeks later
(lasting for three weeks); and vi) mice in the PAR negative control
(PAR-NC) group were fed high-fat diet and intraperitoneally
injected with 6 mg/kg PAR three times a week followed by daily
injections of 200 µl normal saline containing 1.5% (v/v) DMSO five
weeks later (lasting for three weeks).
Normal mouse diet (cat. no. D12450H; Research Diets,
Inc.) and high-fat diet (cat. no. D12451; Research Diets, Inc.)
contained 12 and 45% kcal fat, respectively. Mice were fed for 8
weeks and weighed every week. At the end of the experiment, 1%
pentobarbital sodium (45 mg/kg) was intraperitoneally injected in
mice for anaesthesia. The mouse eyeballs were extracted for blood
collection, and the blood was centrifuged at 900 × g for 15 min at
4°C. The serum was stored at −20°C. The mice were then immediately
fixed and their thoracic cavity was opened for cardiac perfusion.
When the liver completely turned white, sterile surgical
instruments were used for liver resection. The liver of each mouse
was weighed, and the liver index was calculated (liver weight/body
weight) ×100%. The livers of 6 mice in each group were combined as
a tissue homogenate sample that was stored at −80°C. The same
regions of the liver of 6 other mice in each group were used to
prepare frozen sections (−20°C). The same regions of the liver of
the remaining 6 mice were fixed in 10% neutral formalin at room
temperature for 4 h to prepare paraffinized sections for tissue
staining.
The present study was approved by The Animals Ethics
Committee of Liaocheng People's Hospital (approval no. 2019036).
Significant efforts were made to minimize the number of animals
used and the pain they experienced. All procedures were conducted
strictly in accordance with the National Institutes of Health Guide
for the Care and Use of Laboratory Animals.
Biochemical analysis and ELISA
The alanine aminotransferase (ALT; cat. no.
C009-2-1) and aspartate aminotransferase (AST; cat. no. C010-2-1)
levels in the serum were detected according to the instructions of
the commercial kits. The supernatant was prepared by centrifugation
of the liver tissue homogenate at 16,000 × g for 10 min at 4°C.
Total cholesterol (TC; cat. no. A111-1-1), triglyceride (TG; cat.
no. A110-1-1) and malondialdehyde (MDA; cat. no. A003-1-2) levels
and superoxide dismutase (SOD; cat. no. A001-3-2) and glutathione
peroxidase (GSH-Px; cat. no. A005-1-2) activities were measured in
the liver tissue samples according to the manufacturer instruction
for the corresponding kits. All kits were purchased from Nanjing
Jiancheng Bioengineering Institute.
Haematoxylin and eosin (H&E)
staining and NAFLD activity score (NAS)
Paraffinized sections (6 µm) were dewaxed and
stained with haematoxylin (Beijing Solarbio Science &
Technology Co., Ltd.) for 3 min at room temperature. The sections
were then washed and placed in hydrochloric acid and ammonia for 2
sec followed by washes with distilled water. Then, the sections
were stained with eosin for 5 min at room temperature and washed
with distilled water. Finally, the sections were dehydrated in
gradient alcohol, cleared in xylene, and sealed using neutral gum.
Sections were observed under a CKX41SF inverted microscope (Olympus
Corporation). The NAS was used for histological scoring, liver
injury: i) Steatosis (0-3, 0 represents <5%, and 3 represents
>66%); ii) lobular inflammation (0-2, 0 represents no foci, and
2 represents 2–4 foci); and iii) hepatocellular ballooning (0-2, 0
represents none and 2 represents a high number of cells/prominent
ballooning) (22).
Oil Red O staining
Frozen sections (7 µm) of the liver were fixed in
10% neutral formaldehyde at room temperature for 30 min and washed
three times with deionized water. Oil Red O solution
(Sigma-Aldrich; Merck KGaA) was added at room temperature for 10
min. The sections were then washed with deionized water, stained
with haematoxylin at room temperature for 2 min, sealed with
neutral gum and observed under a light microscope (Olympus CKX51;
Olympus Corporation).
Masson's trichrome staining
Paraffinized sections were dewaxed, stained with 100
µl Masson mixture (Beijing Solarbio Science & Technology Co.,
Ltd.) for 5 min and washed using distilled water. The sections were
then stained with 100 µl phosphomolybdic acid for 5 min and dried.
Then, the sections were stained with 100 µl of aniline blue for 5
min and washed with distilled water. Sections were washed twice
continuously (each time for 40 sec) using 1% glacial acetic acid
(100 µl). Finally, the sections were dehydrated using 95% alcohol
and anhydrous alcohol, cleared, sealed using neutral gum and
examined under a light microscope (Olympus CKX51; Olympus
Corporation). All staining assays were performed at room
temperature.
Assay of liver hydroxyproline
content
Liver hydroxyproline content was detected using a
kit (cat. no. A030-2-1; Nanjing Jiancheng Bioengineering Institute)
according to the manufacturer's instructions. Liver samples (20 mg)
were weighed and hydrolysed using an alkaline solution at 95°C for
20 min. Then, 1 ml supernatant was added into R1 and allowed to
stand for 10 min, R2 was added for 5 min and R3 was added to water
bath at 60°C for 15 min. A spectrophotometer was used to measure
the optical density (OD) at 550 nm, and hydroxyproline content per
gram of liver tissue was calculated using a standard curve.
Reverse transcription-quantitative
(RT-q) PCR
Total RNA was extracted from liver samples using a
MiniBEST Universal RNA extraction kit (Takara Biotechnology Co.,
Ltd.), and the concentration of RNA was measured. Total RNA was
reverse transcribed into cDNA according to the instructions of a
SYBR Premix Ex Taq™ II kit (Takara Bio, Inc.). The reaction
conditions were as follows: 70°C for 5 min, 42°C for 60 min, and
75°C for 10 min. qPCR was carried out on a Roche LightCycler 480
instrument (Roche Diagnostics) using SYBR-Green reagent (Thermo
Fisher Scientific, Inc.). The reaction conditions were as follows:
Pre-denaturation at 95°C for 5 min, and 40 cycles of denaturation
at 95°C for 15 sec, annealing at 58°C for 35 sec, and extension at
72°C for 30 sec. β-actin was used as an internal reference, and
relative gene expression was calculated using the 2−ΔΔCq
method (23). Primer sequences are
listed in Table I.
 | Table I.Primer sequences for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences for reverse
transcription-quantitative PCR.
| Gene | Primer
sequence |
|---|
| PPAR-α | F:
5′-ATGGTGGACACAGAGAGCCCCAT-3′ |
|
| R:
5′-TCAGTACATGTCTCTGTAGAT-3′ |
| SREBP-1c | F:
5′-GTGAGCCTGACAAGCAATCA-3′ |
| | R:
5′-GGTGCCTACAGAGCAAGAG-3′ |
| FASN | F:
5′-GGAGGTGGTGATAGCCGGTAT-3′ |
| | R:
5′-TGGGTAATCCATAGAGCCCAG-3′ |
| ACC1 | F:
5′-ATTGGGCACCCCAGAGCTA-3′ |
| | R:
5′-CCCGCTCCTTCAACTTGCT-3′ |
| SCD1 | F:
5′-TTCTTGCGATACACTCTGGTGC-3′ |
| | R:
5′-CGGGATTGAATGTTCTTGTCGT-3′ |
| CPT1α | F:
5′-CACCAACGGGCTCATCTTCTA-3′ |
| | R:
5′-CAAAATGACCTAGCCTTCTATCGAA-3′ |
| ChREBP | R:
5′-ATGCGCGAATACCACAAGTGGAGA-3′ |
| | R:
5′-TTATAATGGTCTCCCCAGGGTGC-3′ |
| ACADS | F:
5′-GACTGGCGACGGTTACACA-3′ |
| | R:
5′-GGCAAAGTCACGGCATGTC-3′ |
| α-SMA | F:
5′-GTCCCAGACATCAGGGAGTAA-3′ |
| | R:
5′-TCGGATACTTCAGCGTCAGGA-3′ |
| TIMP-1 | F:
5′-TTCGTGGGGACACCAGAAGTC-3′ |
| | R:
5′-TATCTGGGACCGCAGGGACTG-3′ |
| TGF-β1 | F:
5′-ATTCCTGGCGTTACCTTGG-3′ |
| | R:
5′-AGCCCTGTATTCCGTCTCCT-3′ |
| TNF-α | F:
5′-CCTGTAGCCCACGTCGTAG-3′ |
| | R:
5′-GGGAGTAGACAAGGTACAACCC-3′ |
| IL-1β | F:
5′-ACGGACCCCAAAAGATGAAG-3′ |
| | R:
5′-TTCTCCACAGCCACAATGAG-3′ |
| IL-6 | F:
5′-CAAAGCCAGAGTCCTTCAGAG-3′ |
| | R:
5′-GTCCTTAGCCACTCCTTCTG-3′ |
| MST1 | F:
5′-CTTCCACTACAACATGAGCAGC-3′ |
| | R:
5′-TGCAGGTCCGCACATAATCTT-3′ |
| LATS1 | F:
5′-CGGAGTACTTCAGAAGTTAATCC-3′ |
| | R:
5′-TACTGACAGATGATCCTCCTC-3′ |
| YAP | F:
5′-CCCGACTCCTTCTTCAAGC-3′ |
| | R:
5′-CTCGAACATGCTGTGGAGTC-3′ |
| TAZ | F:
5′-TCTGGACCAAGTACATGAACC-3′ |
| | R:
5′-AGGACTGGTGATTGGACAC-3′ |
| β-actin | F:
5′-GGCATGGGTCAGAAGGATTCC-3′ |
| | R:
5′-ATGTCACGCACGATTTCCCGC-3′ |
Western blot analysis
Total protein was extracted from mouse liver tissue
using precooled RIPA buffer (Biosesang) containing protease
inhibitor (Sigma-Aldrich; Merck KGaA), and the concentration was
determined using a BCA kit (Beyotime Institute of Biotechnology).
Then, 10 µg protein was separated via SDS-PAGE on a 12% gel. The
proteins were then transferred to a PVDF membrane, which was
blocked with 5% non-fat milk for 1 h at room temperature. The
membranes were incubated with primary antibodies against mammalian
STE20-like protein kinase 1 (MST1; cat. no. ab79199; 1:500; 56
kDa), large tumour suppressor kinase 1 (LATS1; cat. no. ab70561;
1:5,000; 127 kDa), YAP (cat. no. ab52771; 1:5,000; 70 kDa) and TAZ
(cat. no. ab119373; 1:1,000; 44 kDa) overnight at 4°C. A
HRP-conjugated rabbit anti-mouse immunoglobulin G (cat. no. ab6728;
1:2,000) secondary antibody was added for 1 h at room temperature
followed by exposure, development and visualization using an ECL
working solution (EMD Millipore). β-actin (cat. no. ab8226;
1:1,000; 42 kDa) was used as an internal reference. All antibodies
were purchased from Abcam. Protein bands were analysed using
ImageJ2× software (version 2.1.4.7; National Institutes of
Health).
Statistical analysis
SPSS 21.0 (IBM Corp.) was used for statistical
analysis. The data were normally distributed, according to a
Kolmogorov-Smirnov test. Thus, the results are presented as the
mean ± SD. Unpaired Student's t-tests were used to compare the
differences between two groups. One-way or two-way ANOVA followed
by Sidak's post hoc test were used to compare multiple groups.
P-values were obtained from two-tailed tests. P<0.05 was
considered to indicate statistically significant difference.
Results
PAR alleviates liver injury in MAFLD
mice
Previous studies have suggested that PAR has
anti-inflammatory, antioxidant and antifibrotic properties
(24–26). Therefore, it was hypothesized that
PAR may have protective effects on MAFLD-induced liver injury. To
examine effect of PAR on MAFLD, male C57BL/6 mice were fed high-fat
diet for 8 weeks. The body weight and liver index (liver to body
weight ratio) of mice in the model group were significantly higher
than those in the control group (P<0.01; Fig. 1A and B). In addition, H&E
staining demonstrated that hepatocytes in the model group displayed
microbubbles and macrovesicular steatosis (Fig. 1C). The histological scores
(steatosis, lobular inflammation and hepatocyte ballooning) in the
model group were also significantly increased, compared with the
control group (all P<0.01; Fig.
1D). These results indicated that the MAFLD mouse model was
successfully generated.
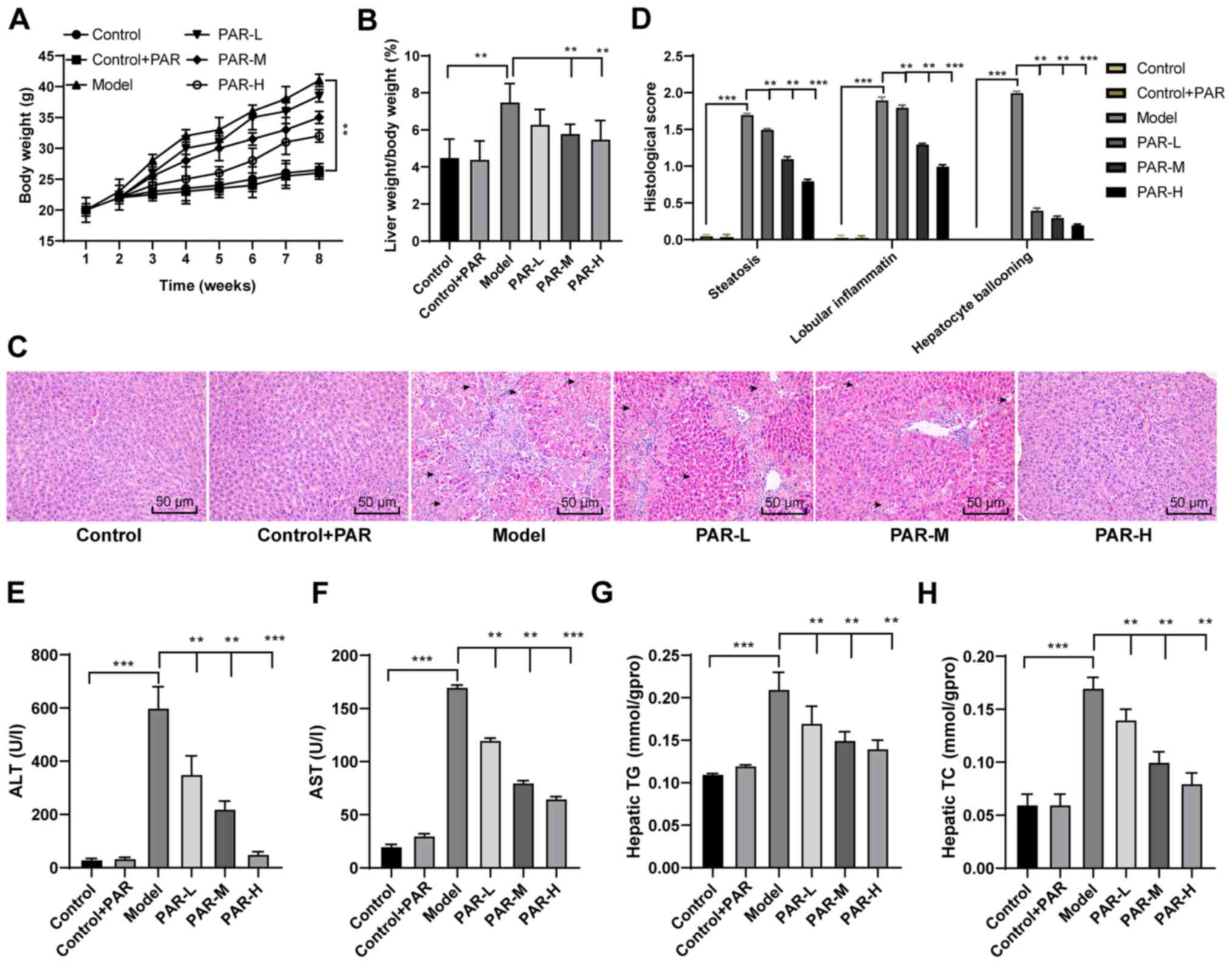 | Figure 1.PAR has beneficial effects on the
manifestations of MAFLD in mouse liver. (A) Changes in the body
weight of mice during the experiment (1-8 weeks). **P<0.01,
two-way ANOVA followed by Sidak's post hoc test. (B) Liver to body
weight ratio at week 8. (C) Representative images of H&E
staining of mouse livers (scale bar, 50 µm). (D) H&E-stained
sections were evaluated using a histological score. (E and F) ALT
and AST serum levels in mice. (G and H) Hepatic TC and TG levels in
mice. The data are presented as the mean ± SD. n=6. **P<0.01,
***P<0.001, one-way ANOVA followed by Sidak's post hoc test,
unless otherwise stated. PAR, parthenolide; MFALD, metabolic
dysfunction-associated fatty liver disease; H&E, haematoxylin
and eosin; ALT, alanine transaminase; AST, aspartate transaminase;
TC, total cholesterol; TG, triglyceride; L, low; M, medium; H,
high; gpro, gramme of protein. |
There were no significant differences in the body
weight, liver index and H&E staining in the liver sections
between the control + PAR and the control groups (Fig. 1A-D), indicating that PAR did not
damage normal mouse livers. Mice in the PAR-L, -M and -H treatment
groups exhibited significantly reduced body weight, liver index and
the histological score (all P<0.01; Fig. 1A-D). PAR treatment also
significantly reduced the serum levels of ALT and AST and decreased
the hepatic TC and TG contents in MAFLD mice (all P<0.01;
Fig. 1E-H).
PAR improves liver lipid metabolism
and fibrosis in MAFLD mice
Oil Red O staining was performed to determine
whether PAR could improve hepatic lipid metabolism. The volume and
number of lipid droplets were reduced in the PAR-H group, compared
with the model group (Fig. 2A).
Additionally, the expression levels of lipogenesis- and
metabolism-related genes were measured using RT-qPCR. The mRNA
expression levels of liposynthesis-related genes, such as sterol
regulatory element binding protein (SREBP)-1c, fatty acid synthase
(FASN), acetyl CoA carboxylase 1 (ACC1), stearoyl CoA desaturase 1
(SCD1) and carbohydrate response element-binding protein (ChREBP),
were significantly lower in the PAR-H group than in the model group
(all P<0.01; Fig. 2B). Moreover,
PAR-H treatment also significantly increased the mRNA expression
levels of genes associated with fatty acid β-oxidation, including
peroxisome proliferator-activated receptor α (PPAR-α), carnitine
palmitoyl transferase 1α (CPT1α) and acyl-CoA dehydrogenase short
chain (ACADS) (all P<0.01; Fig.
2C).
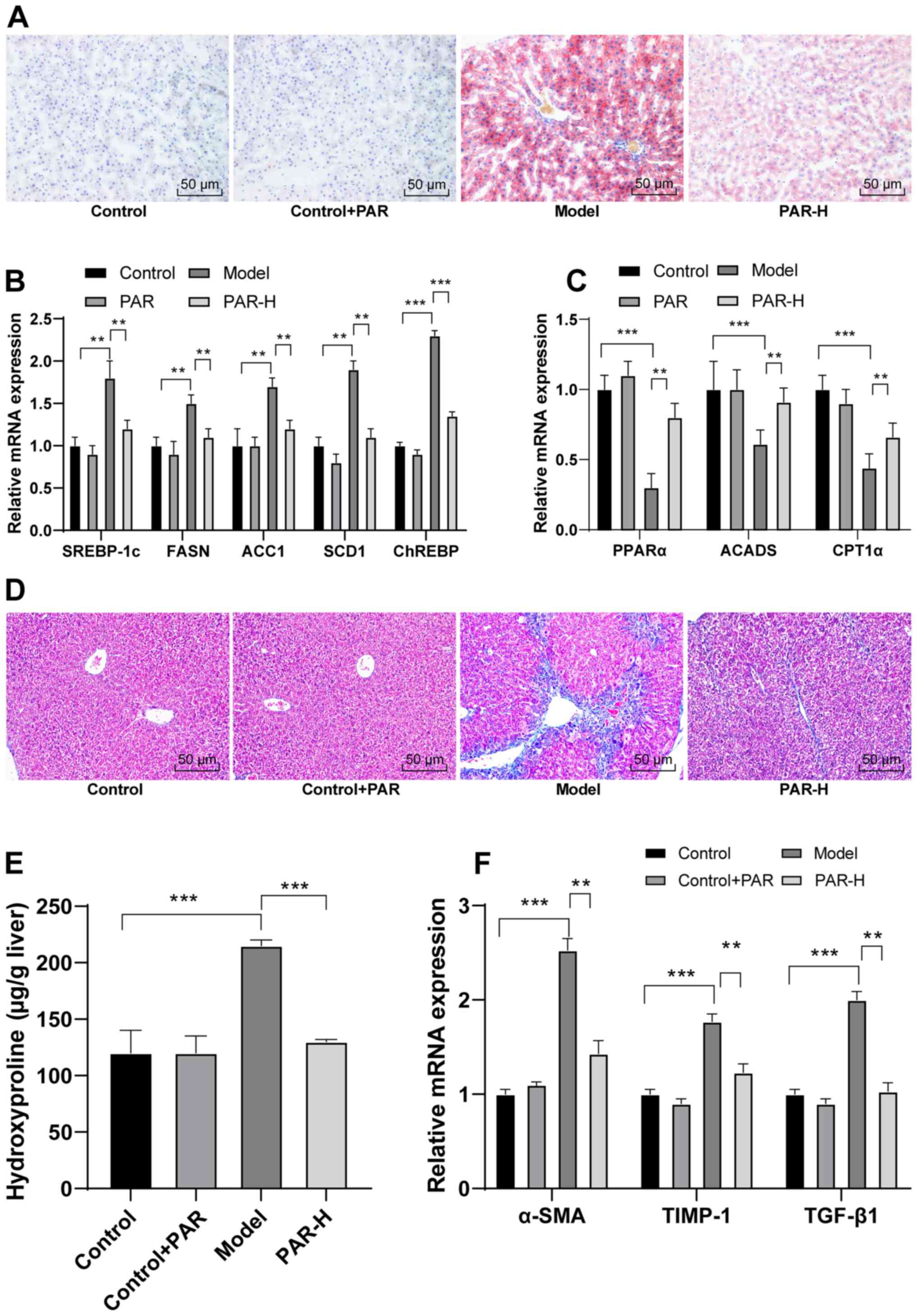 | Figure 2.PAR improves hepatic lipid metabolism
and fibrosis in mice with MAFLD. (A) Representative images of Oil
Red O staining of mouse liver sections (scale bar, 50 µm). (B) mRNA
expression levels of the liposynthesis-related genes SREBP-1c,
FASN, ACC1, SCD1 and ChREBP. (C) mRNA expression levels of the
fatty acid β-oxidation-related genes PPARα, ACADS and CPT1α. (D)
Representative images of Masson's trichrome staining of mouse liver
sections (scale bar, 50 µm). (E) Liver hydroxyproline content.
***P<0.001, one-way ANOVA followed by Sidak's post hoc test. (F)
mRNA expression levels of hepatic fibrosis-related genes α-SMA,
TIMP-1 and TGF-β1. The data are presented as the mean ± SD. n=6.
**P<0.01, ***P<0.001, two-way ANOVA followed by Sidak's post
hoc test, unless otherwise stated. PAR, parthenolide; MFALD,
metabolic dysfunction-associated fatty liver disease; SREBP-1c,
sterol regulatory element binding protein-1c; FASN, fatty acid
synthase; ACC1, acetyl CoA carboxylase 1; SCD1, stearoyl CoA
desaturase 1; ChREBP, carbohydrate response element-binding
protein; PPAR-α; peroxisome proliferator-activated receptor α;
ACADS, acyl-CoA dehydrogenase short chain; CPT1α, carnitine
palmitoyl transferase 1α; α-SMA, α-smooth muscle actin; TIMP-1,
tissue inhibitor of metalloproteinase 1; L, low; M, medium; H,
high. |
Liver fibrosis was assessed in all mice using
Masson's trichrome staining, and liver hydroxyproline content was
measured in order to determine whether PAR could reduce hepatic
fibrosis in mice with MAFLD. Mice in the PAR-H group displayed
improved liver fibrosis (Fig. 2D)
and significantly reduced liver hydroxyproline content, compared
with the model group (P<0.001; Fig.
2E). In addition, PAR-H treatment significantly reduced the
mRNA expression levels of liver fibrosis-related genes, such as
α-smooth muscle actin (α-SMA), tissue inhibitor of
metalloproteinase 1 (TIMP-1) and TGF-β1 (all P<0.01; Fig. 2F).
PAR reduces liver inflammation and
oxidative stress in MAFLD mice
The mRNA expression levels of pro-inflammatory
cytokines (TNF-α, IL-1β and IL-6) were measured using RT-qPCR to
determine the effect of PAR on liver inflammation in MAFLD. PAR-H
treatment significantly reduced the expression levels of these
pro-inflammatory factors in mice with MAFLD (all P<0.01;
Fig. 3A).
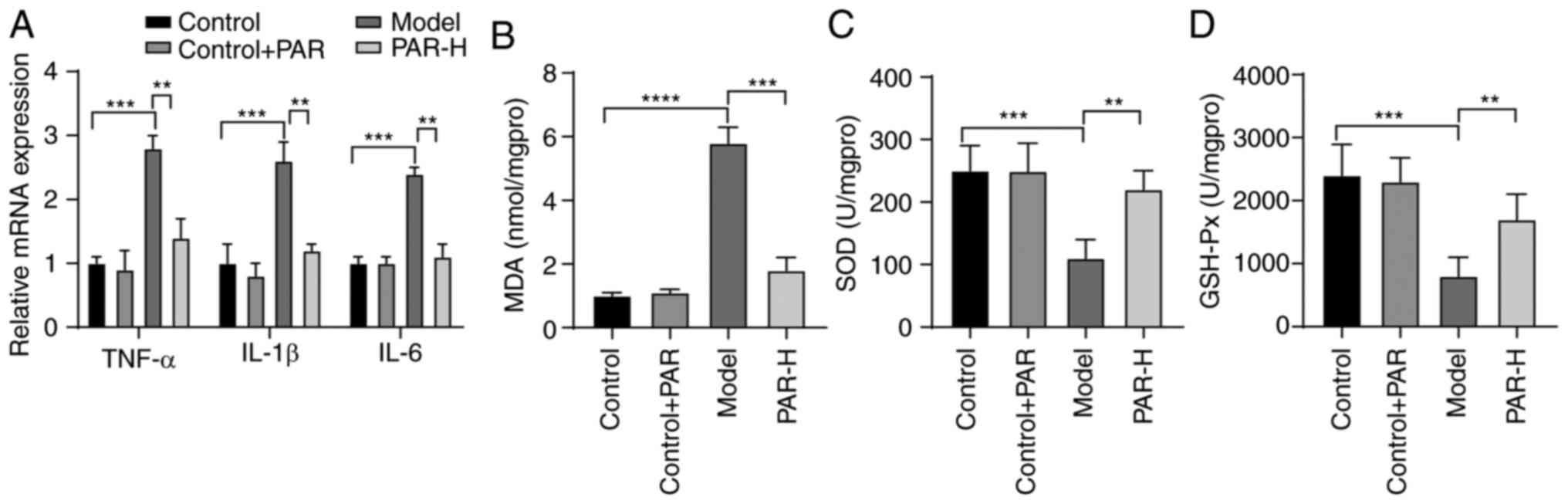 | Figure 3.PAR reduces liver inflammation and
oxidative stress in mice with MAFLD. (A) mRNA expression levels of
pro-inflammatory cytokines TNF-α, IL-1β and IL-6. **P<0.01,
***P<0.001, two-way ANOVA followed by Sidak's post hoc test.
(B-D) MDA, SOD and GSH-Px levels in the liver. The data are
presented as the mean ± SD. n=6. **P<0.01, ***P<0.001,
two-way ANOVA followed by Sidak's post hoc test, unless otherwise
stated. PAR, parthenolide; MFALD, metabolic dysfunction-associated
fatty liver disease; MDA, malondialdehyde; SOD, superoxide
dismutase; GSH-Px, glutathione peroxidase; H, high. |
MDA levels and SOD and GSH-Px activities were
measured in the liver in order to evaluate the role of PAR in
hepatic oxidative stress in mice with MAFLD. Mice in the model
group displayed a significant increase in MDA levels and a decrease
in SOD and GSH-Px activities, compared with the control group (all
P<0.001; Fig. 3B-D). However,
PAR-H treatment reversed these changes in mice with MAFLD (all
P<0.01; Fig. 3B-D).
PAR activates the HIPPO pathway in the
liver of mice with MAFLD
YAP is a key mediator of the HIPPO pathway. Liver
dysfunction is accompanied by an increase in the YAP expression in
MAFLD and HIPPO pathway activation can reduce liver damage
(27,28). Therefore, it was hypothesized that
PAR could play a protective role in the liver of mice with MAFLD
through the activation of the HIPPO pathway. The mRNA and protein
expression levels of several mediators of the HIPPO pathway (MST1,
LATS1, YAP and TAZ) were measured. The expression levels of MST1
and LATS1 in the liver of PAR-H-treated mice were significantly
higher than those in the model mice. Moreover, the expression
levels of YAP and TAZ were reduced (all P<0.01; Fig. 4A and B). These results indicated
that PAR could activate the HIPPO pathway in the liver of mice with
MAFLD.
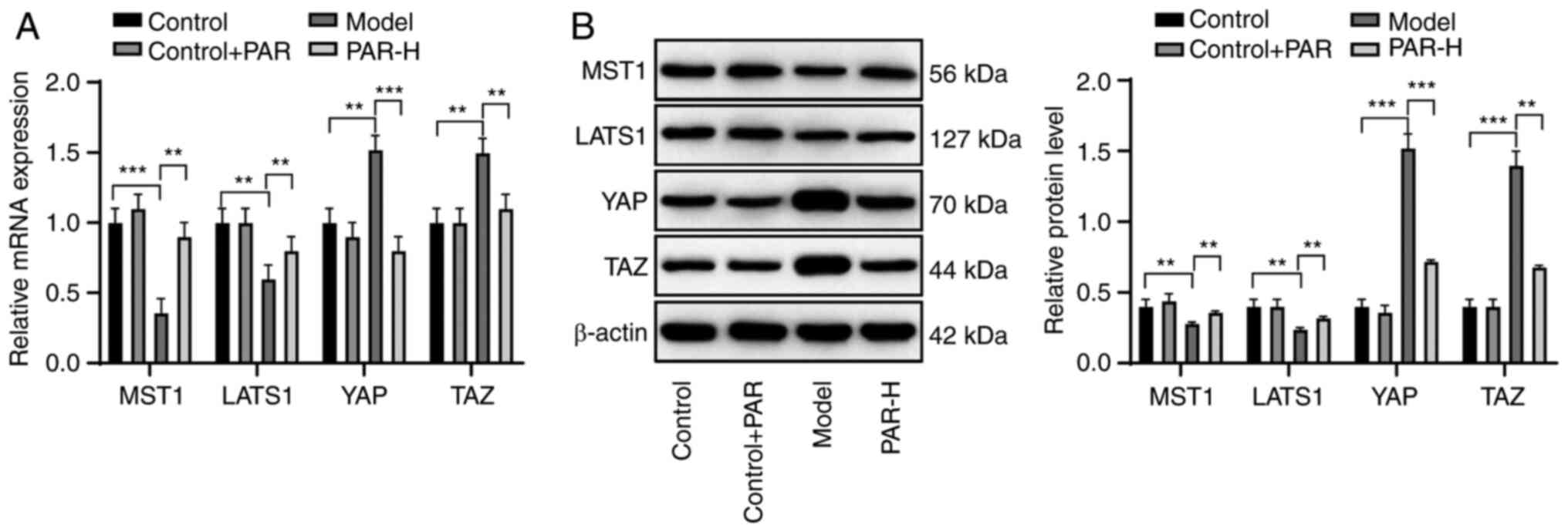 | Figure 4.PAR activates the HIPPO pathway in
the liver of mice with MAFLD. (A) mRNA and (B) protein expression
levels of MST1, LATS1, YAP and TAZ were detected using reverse
transcription-quantitative PCR and western blot analysis,
respectively. The data are presented as the mean ± SD. n=6.
**P<0.01, ***P<0.001, two-way ANOVA followed by Sidak's post
hoc test. PAR, parthenolide; MFALD, metabolic
dysfunction-associated fatty liver disease; MST1,
macrophage-stimulating 1; LATS1, large tumour suppressor kinase 1;
YAP, Yes-associated protein; TAZ, transcriptional coactivator with
PDZ-binding motif; low; M, medium. |
Inhibition of the HIPPO pathway
reverses the protective effect of PAR in the liver of mice with
MAFLD
To confirm whether PAR treatment exerted beneficial
effects on liver injury in mice with MAFLD through activation of
the HIPPO pathway, mice were intraperitoneally injected with VP, an
inhibitor of the HIPPO pathway. VP treatment in the PAR-VP group
significantly reduced MST1 expression in the liver and increased
YAP expression, compared with PAR-NC (all P<0.01; Fig. 5A and B), indicating that VP
successfully inhibited the HIPPO pathway in vivo. H&E
staining demonstrated that hepatocytes in PAR-VP mice displayed
microbubbles (Fig. 5C). In
addition, the histological scores (steatosis, lobular inflammation
and hepatocyte ballooning) in the PAR-VP group were significantly
increased, compared with PAR-NC (P<0.001; Fig. 5D). Moreover, PAR-VP treatment
increased the number of lipid droplets (Fig. 5E) and aggravated hepatic fibrosis in
mice (Fig. 5F).
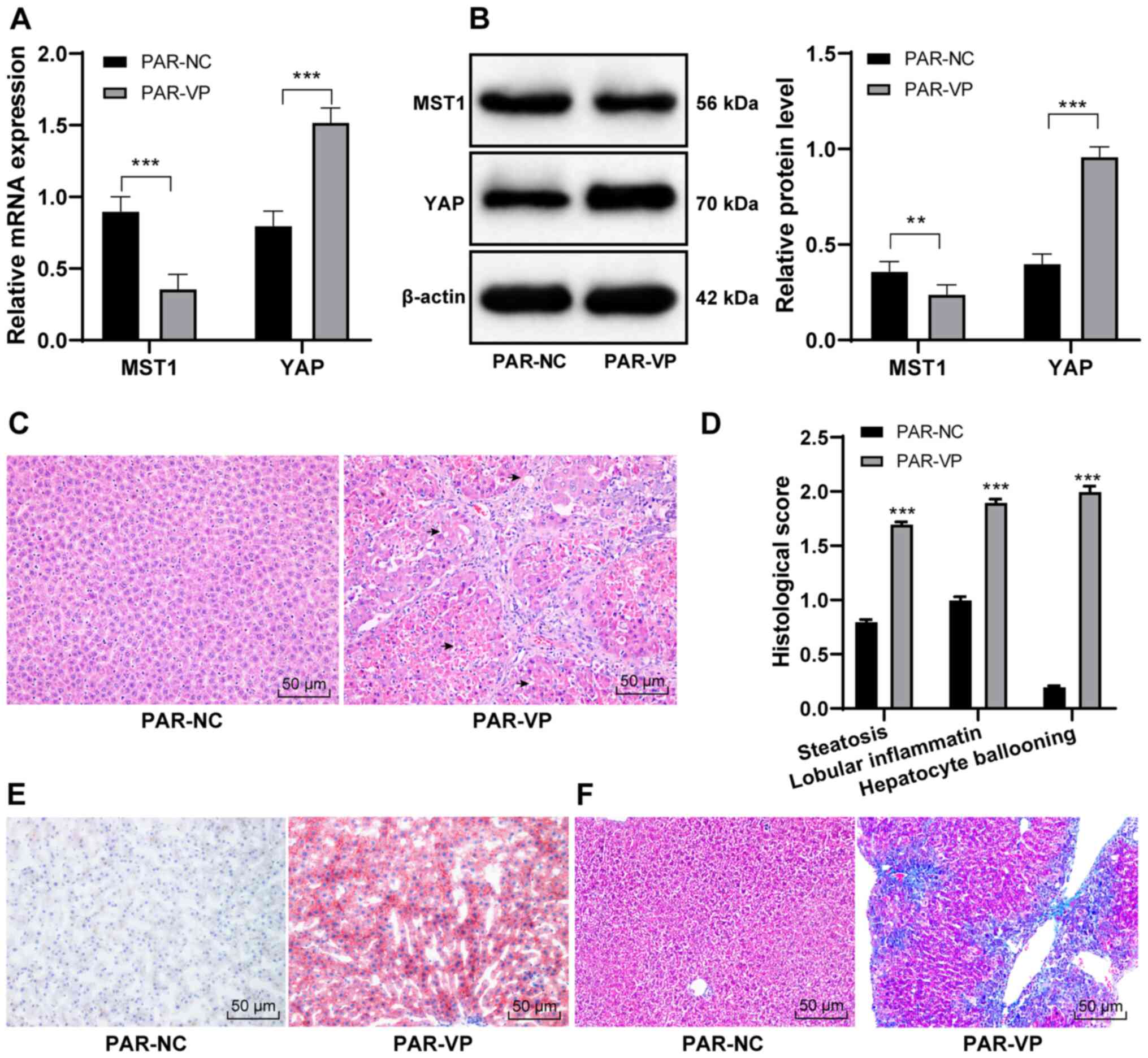 | Figure 5.Inhibition of the HIPPO pathway
reverses the protective effect of PAR on the livers of mice with
MAFLD. (A) mRNA and (B) protein expression levels of MST1 and YAP.
(C) Representative images of H&E staining of mouse livers
(scale bar, 50 µm). (D) H&E-stained sections were evaluated
using a histological score. ***P<0.001, Student's t-test. (E)
Representative images of Oil Red O staining of mouse liver sections
(scale bar, 50 µm). (F) Representative images of Masson's trichrome
staining of mouse liver sections. The data are presented as the
mean ± SD. n=6. **P<0.01, ***P<0.001 vs. PAR-NC group,
two-way ANOVA followed by Sidak's post hoc test, unless otherwise
stated. PAR, parthenolide; MFALD, metabolic dysfunction-associated
fatty liver disease; H&E, haematoxylin-eosin; VP, verteporfin;
NC, negative control; MST1, macrophage-stimulating 1; YAP,
Yes-associated protein. |
Discussion
MAFLD is the leading cause of liver disease globally
and affects both children and adults (29). PAR, a natural compound present in
the feverfew plant, has various biological and therapeutic
activities, including anti-inflammatory effects (30). In the present study, PAR attenuated
the pathological symptoms of MAFLD in mice by activating the HIPPO
pathway to suppress liver injury, hepatic lipid metabolism,
fibrosis, inflammation and oxidative stress.
An increase in the serum levels of the biochemical
markers AST, ALT, TC and TG may indicate acute hepatic failure
(31). In the present study, PAR
treatment improved histological scores and reduced the serum levels
of ALT and AST, as well as TC and TG content in the liver of mice
with MAFLD. Moreover, PAR administration reduced the body weight
and the levels of TG, ALT and AST in the liver of MAFLD model mice,
which was similar to the findings of a previous study (14). These results indicated that PAR
exerted beneficial effects on liver injury in the MAFLD model.
A previous study has demonstrated that the lipid
metabolism-associated genes SREBP-1c, ACC1, FASN, SCD1 and CPT1α
are markers of lipid accumulation and hepatic steatosis in MAFLD
(32). ChREBP is an important
transcription factor controlling hepatic energy metabolism and
lipid metabolism (33). Reduced
levels of hydroxyproline, as well as TGF-β1, α-SMA and TIMP-1 mRNA
expression levels are closely associated with the suppression of
hepatic fibrosis (34). The present
findings demonstrated that PAR treatment downregulated the
expression levels of liposynthesis markers (SREBP-1c, FASN, ACC1,
SCD1 and ChREBP) and upregulated the expression of genes associated
with fatty acid β-oxidation (PPARα, CPT1α and ACADS). Similarly,
PAR plays an inhibitory role in lipid accumulation and adipogenesis
(30).
PAR normalizes the aberrant lipid metabolism and
fatty acid β-oxidation in MAFLD rats (11). Additionally, a previous study has
demonstrated that increased hydroxyproline content in the liver and
hepatic collagen deposition are associated with aggravated liver
fibrosis (35). The results of the
present study indicated that PAR treatment could reduce the
expression levels of liver fibrosis-associated markers (α-SMA,
TIMP-1 and TGF-β1), as well as liver hydroxyproline content and
collagen deposition. These conclusions are supported by a study
that demonstrated the protective effect of PAR on hepatic fibrosis
(13). Overall, PAR alleviates the
changes in hepatic lipid metabolism and fibrosis in mice with
MAFLD.
Additionally, PAR has been reported to have potent
anti-inflammatory and antioxidative stress activities (14,36).
TNF-α, IL-1β and IL-6 are proinflammatory cytokines (37). Decreased SOD and GSH-Px in
combination with increased MDA levels are closely associated with
oxidative stress (38). PAR has
been shown to regulate obesity-induced inflammatory/oxidant
responses and proposed as an effective agent for the treatment of
obesity-related diseases (39). In
agreement with these previous findings, PAR-treated MAFLD model
mice in the present study displayed decreased TNF-α, IL-1β, IL-6
and MDA levels, together with elevated SOD and GSH-Px activities.
Thus, PAR alleviated liver injury, changes in hepatic metabolism
and fibrosis, inflammation and oxidative stress in the MAFLD
model.
However, the mechanism of action of PAR in MAFLD is
incompletely understood. The HIPPO pathway has been suggested to
play a role in metabolic diseases through the regulation of key
molecules, including MST1/2, LATS1/2 and the downstream YAP/TAZ
transcriptional coactivators (40).
The HIPPO pathway, as a novel pathway associated with growth
control and cancer inhibition, plays a significantly regulatory
role in liver development, injury and disease (41). The HIPPO pathway contributes to the
prevention of MAFLD and liver cancer progression (42). Furthermore, a previous study has
suggested that the HIPPO pathway is implicated in hepatic fibrosis
(16). YAP is closely associated
with the regulation of MAFLD-associated lipid metabolism and
hepatic fibrosis (28). In the
present study, PAR treatment activated the HIPPO pathway in MAFLD
model mice, as evidenced by increased levels of MST1 and LATS1
expression and a reduction in the levels of YAP and TAZ expression.
Additionally, inhibition of the HIPPO pathway reversed the
protective effect of PAR in MAFLD model mice. However, previous
studies have not reported that the protective effects of PAR in
MAFLD are mediated by the HIPPO pathway, which was therefore a
novel finding of the present study.
In conclusion, the present study demonstrated that
PAR alleviated MAFLD through the activation of the HIPPO pathway in
mice. However, only the protective effect of PAR on fatty liver in
MAFLD was evaluated, and the role of PAR in other hepatic lesions
was not studied. In the future, the role and mechanism of action of
PAR in other types of liver injury will be further assessed.
Altogether, the present study provided novel insight for the
treatment of MAFLD; however, the clinical applications of these
findings remain to be determined. Promising PAR-based therapeutic
approaches involving the HIPPO pathway may also be investigated for
the purpose of MAFLD treatment.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WW, YH and QL conceived and designed the study. WW
and QL performed the analysis and interpretation of the data and
drafted the manuscript. YH and QL critically revised the article
for important intellectual content and assisted in the literature
search for this article. All authors agree to be accountable for
all aspects of the work in ensuring that questions related to the
accuracy or integrity of any part of the work are appropriately
investigated. WW and YH confirm the authenticity of all the raw
data. All authors read and approval the final manuscript.
Ethics approval and consent to
participate
The present study was approved by The Animals Ethics
Committee of Liaocheng People's Hospital (approval no. 2019036).
Significant efforts were made to minimize the number of animals
used and the pain they experienced. All procedures were conducted
strictly in accordance with the National Institutes of Health Guide
for the Care and Use of Laboratory Animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tilg H and Effenberger M: From NAFLD to
MAFLD: When pathophysiology succeeds. Nat Rev Gastroenterol
Hepatol. 17:387–388. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Farzanegi P, Dana A, Ebrahimpoor Z, Asadi
M and Azarbayjani MA: Mechanisms of beneficial effects of exercise
training on non-alcoholic fatty liver disease (NAFLD): Roles of
oxidative stress and inflammation. Eur J Sport Sci. 19:994–1003.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Eslam M, Newsome PN, Sarin SK, Anstee QM,
Targher G, Romero-Gomez M, Zelber-Sagi S, Wai-Sun Wong V, Dufour
JF, Schattenberg JM, et al: A new definition for metabolic
dysfunction-associated fatty liver disease: An international expert
consensus statement. J Hepatol. 73:202–209. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Eslam M and George J: Reply to:
Correspondence on ‘A new definition for metabolic associated fatty
liver disease: An international expert consensus statement’: MAFLD:
Moving from a concept to practice. J Hepatol. 73:1268–1269. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Eslam M, Valenti L and Romeo S: Genetics
and epigenetics of NAFLD and NASH: Clinical impact. J Hepatol.
68:268–279. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jegatheesan P and De Bandt JP: Fructose
and NAFLD: The multifaceted aspects of fructose metabolism.
Nutrients. 9:2302017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bellentani S: The epidemiology of
non-alcoholic fatty liver disease. Liver Int. 37 (Suppl 1):81–84.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rotman Y and Sanyal AJ: Current and
upcoming pharmacotherapy for non-alcoholic fatty liver disease.
Gut. 66:180–190. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Van De Wier B, Koek GH, Bast A and Haenen
GR: The potential of flavonoids in the treatment of non-alcoholic
fatty liver disease. Crit Rev Food Sci Nutr. 57:834–855. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu C, Liao JZ and Li PY: Traditional
Chinese herbal extracts inducing autophagy as a novel approach in
therapy of nonalcoholic fatty liver disease. World J Gastroenterol.
23:1964–1973. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Duan D, Zhang J, Yao J, Liu Y and Fang J:
Targeting thioredoxin reductase by parthenolide contributes to
inducing apoptosis of HeLa cells. J Biol Chem. 291:10021–10031.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang D, Wang H, Fu S, Cheng X, Yang F,
Zhang Q, Li Y, Xue Z, Zhang L, Huang W, et al: Parthenolide
ameliorates Concanavalin A-induced acute hepatitis in mice and
modulates the macrophages to an anti-inflammatory state. Int
Immunopharmacol. 38:132–138. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Barbier-Torres L, Beraza N,
Fernández-Tussy P, Lopitz-Otsoa F, Fernández-Ramos D,
Zubiete-Franco I, Varela-Rey M, Delgado TC, Gutiérrez V, Anguita J,
et al: Histone deacetylase 4 promotes cholestatic liver injury in
the absence of prohibitin-1. Hepatology. 62:1237–1248. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bahabadi M, Mohammadalipour A, Karimi J,
Sheikh N, Solgi G, Goudarzi F, Hashemnia M and Khodadadi I:
Hepatoprotective effect of parthenolide in rat model of
nonalcoholic fatty liver disease. Immunopharmacol Immunotoxicol.
39:233–242. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Patel SH, Camargo FD and Yimlamai D: Hippo
signaling in the liver regulates organ size, cell fate, and
carcinogenesis. Gastroenterology. 152:533–545. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou Y, Lv X, Qu H, Zhao K, Fu L, Zhu L,
Ye G and Guo J: Differential expression of circular RNAs in hepatic
tissue in a model of liver fibrosis and functional analysis of
their target genes. Hepatol Res. 49:324–334. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fujisawa K, Takami T, Sasai N, Matsumoto
T, Yamamoto N and Sakaida I: Metabolic alterations in
spheroid-cultured hepatic stellate cells. Int J Mol Sci.
21:34512020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yi M, Li J, Chen S, Cai J, Ban Y, Peng Q,
Zhou Y, Zeng Z, Peng S, Li X, et al: Correction to: Emerging role
of lipid metabolism alterations in Cancer stem cells. J Exp Clin
Cancer Res. 37:1552018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kodama T, Yi J, Newberg JY, Tien JC, Wu H,
Finegold MJ, Kodama M, Wei Z, Tamura T, Takehara T, et al:
Molecular profiling of nonalcoholic fatty liver disease-associated
hepatocellular carcinoma using SB transposon mutagenesis. Proc Natl
Acad Sci USA. 115:E10417–E10426. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lopez-Franco O, Hernández-Vargas P,
Ortiz-Muñoz G, Sanjuán G, Suzuki Y, Ortega L, Blanco J, Egido J and
Gómez-Guerrero C: Parthenolide modulates the NF-kappaB-mediated
inflammatory responses in experimental atherosclerosis.
Arterioscler Thromb Vasc Biol. 26:1864–1870. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim SL, Liu YC, Seo SY, Kim SH, Kim IH,
Lee SO, Lee ST, Kim DG and Kim SW: Parthenolide induces apoptosis
in colitis-associated colon cancer, inhibiting NF-κB signaling.
Oncol Lett. 9:2135–2142. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kleiner DE, Brunt EM, Van Natta M, Behling
C, Contos MJ, Cummings OW, Ferrell LD, Liu YC, Torbenson MS,
Unalp-Arida A, et al: Design and validation of a histological
scoring system for nonalcoholic fatty liver disease. Hepatology.
41:1313–1321. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang M and Li Q: Parthenolide could become
a promising and stable drug with anti-inflammatory effects. Nat
Prod Res. 29:1092–1101. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liao K, Xia B, Zhuang QY, Hou MJ, Zhang
YJ, Luo B, Qiu Y, Gao YF, Li XJ, Chen HF, et al: Parthenolide
inhibits cancer stem-like side population of nasopharyngeal
carcinoma cells via suppression of the NF-κB/COX-2 pathway.
Theranostics. 5:302–321. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li XH, Xiao T, Yang JH, Qin Y, Gao JJ, Liu
HJ and Zhou HG: Parthenolide attenuated bleomycin-induced pulmonary
fibrosis via the NF-κB/Snail signaling pathway. Respir Res.
19:1112018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Machado MV, Michelotti GA, Pereira TA, Xie
G, Premont R, Cortez-Pinto H and Diehl AM: Accumulation of duct
cells with activated YAP parallels fibrosis progression in
non-alcoholic fatty liver disease. J Hepatol. 63:962–970. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen P, Luo Q, Huang C, Gao Q, Li L, Chen
J, Chen B, Liu W, Zeng W and Chen Z: Pathogenesis of non-alcoholic
fatty liver disease mediated by YAP. Hepatol Int. 12:26–36. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Younossi Z, Anstee QM, Marietti M, Hardy
T, Henry L, Eslam M, George J and Bugianesi E: Global burden of
NAFLD and NASH: Trends, predictions, risk factors and prevention.
Nat Rev Gastroenterol Hepatol. 15:11–20. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Berdan CA, Ho R, Lehtola HS, To M, Hu X,
Huffman TR, Petri Y, Altobelli CR, Demeulenaere SG, Olzmann JA, et
al: Parthenolide covalently targets and inhibits focal adhesion
kinase in breast cancer cells. Cell Chem Biol. 26:1027–1035 e22.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu CT, Deng JS, Huang WC, Shieh PC, Chung
MI and Huang GJ: Salvianolic acid C against acetaminophen-induced
acute liver injury by attenuating inflammation, oxidative stress,
and apoptosis through inhibition of the Keap1/Nrf2/HO-1 signaling.
Oxid Med Cell Longev. 2019:90568452019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xia SF, Shao J, Zhao SY, Qiu YY, Teng LP,
Huang W, Wang SS, Cheng XR and Jiang YY: Niga-ichigoside F1
ameliorates high-fat diet-induced hepatic steatosis in male mice by
Nrf2 activation. Food Funct. 9:906–916. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rui L: Energy metabolism in the liver.
Compr Physiol. 4:177–197. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Makled MN, Sharawy MH and El-Awady MS: The
dual PPAR-α/γ agonist saroglitazar ameliorates
thioacetamide-induced liver fibrosis in rats through regulating
leptin. Naunyn Schmiedebergs Arch Pharmacol. 392:1569–1576. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Y, Miao H, Guan H, Wang C, Wang Z
and Ji L: Long-term diosbulbin B treatment induced liver fibrosis
in mice. Chem Biol Interact. 298:15–23. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li S, Gao X, Wu X, Wu Z, Cheng L, Zhu L,
Shen D and Tong X: Parthenolide inhibits LPS-induced inflammatory
cytokines through the toll-like receptor 4 signal pathway in THP-1
cells. Acta Biochim Biophys Sin (Shanghai). 47:368–375. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Paolucci EM, Loukov D, Bowdish DME and
Heisz JJ: Exercise reduces depression and inflammation but
intensity matters. Biol Psychol. 133:79–84. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Min YN, Niu ZY, Sun TT, Wang ZP, Jiao PX,
Zi BB, Chen PP, Tian DL and Liu FZ: Vitamin E and vitamin C
supplementation improves antioxidant status and immune function in
oxidative-stressed breeder roosters by up-regulating expression of
GSH-Px gene. Poult Sci. 97:1238–1244. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kim CY, Jung YW and Park JS: Parthenolide,
a feverfew-derived phytochemical, ameliorates obesity and
obesity-induced inflammatory responses via the Nrf2/Keap1 pathway.
Pharmacol Res. 145:1042592019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ardestani A, Lupse B and Maedler K: Hippo
signaling: Key emerging pathway in cellular and whole-body
metabolism. Trends Endocrinol Metab. 29:492–509. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Johnson RL: Hippo signaling and epithelial
cell plasticity in mammalian liver development, homeostasis, injury
and disease. Sci China Life Sci. 62:1609–1616. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jeong SH, Kim HB, Kim MC, Lee JM, Lee JH,
Kim JH, Kim JW, Park WY, Kim SY, Kim JB, et al: Hippo-mediated
suppression of IRS2/AKT signaling prevents hepatic steatosis and
liver cancer. J Clin Invest. 128:1010–1025. 2018. View Article : Google Scholar : PubMed/NCBI
|














