Introduction
Epithelial ovarian cancer (EOC) is the most lethal
gynecological malignancy for which chemotherapy is an indispensable
treatment option in patients of advanced cases. Despite significant
progress in treatment, ~70% of patients with ovarian cancer (OC)
relapse within 2 years after surgery (1). The main reason for the high mortality
rate of OC is drug resistance. Therefore, elucidation of the
underlying molecular mechanisms is critical for improving
therapeutic effects and prognosis.
Multiple drug resistance (multidrug resistance;
MDR), a phenomenon whereby human tumors that acquire resistance to
one type of therapy are found to be resistant to several other
drugs that are often quite different in both structure and mode of
action, has been recognized clinically for several decades
(2). One of the classical
mechanisms of MDR is increased expression of P-glycoprotein (P-gp),
a drug efflux pump on the cell membrane. It has been well
established that P-gp actively effluxes cytotoxic substrates such
as paclitaxel and adriamycin, the first-line OC chemotherapy drugs
(3,4). Moreover, P-gp may have side effects
that are independent of its drug efflux activity. Several authors
have described the influences of P-gp on the function of proteins
involved in regulatory pathways, including apoptotic progression
(such as p53, caspase-3 and Pokemon) (5). Gibalová et al (6) demonstrated that even though platinum
drugs are not substrates of P-gp, increased P-gp levels can induce
platinum resistance in tumor cells.
Human epididymis protein 4 (HE4), a secreted
glycoprotein, has been recently characterized as a more sensitive
OC biomarker than carbohydrate antigen 125 (CA125). The protein is
overexpressed in patients with OC and is positively correlated with
malignancy (7). Several recent
clinical studies have demonstrated that HE4 is closely associated
with prognosis and is effective in predicting recurrence and
chemotherapeutic resistance (8,9). A
recent study by Ribeiro et al (10) also reported that increased HE4
expression simultaneously induces resistance to paclitaxel and
carboplatin in OC cells in vitro.
ANXA2, a calcium-dependent phospholipid-binding
protein, acts as a fibrinolytic receptor to promote matrix
remodeling, protein hydrolysis, neovascularization, and invasion
and metastasis of malignant tumors (11–13).
Previously, it was demonstrated that HE4 and ANXA2 are interacting
proteins and HE4 requires ANXA2 to promote cancer cell invasion and
metastasis. Invasion/metastasis and drug resistance are important
biological characteristics of malignant tumors (14). Metastatic tumors are more resistant
to chemotherapeutic drugs, suggesting an association between drug
resistance and cancer invasion/metastasis. In breast cancer, ANXA2
interacts with P-gp and promotes the migration and invasion of
drug-resistant tumor cells (15).
Multiple studies have provided evidence of a significant
association of ANXA2 with drug resistance in diverse malignant
tumors (16–18). ANXA2 increases P-gp expression in
gastric cancer cells by activating the MAPK signaling pathway,
resulting in MDR of cancer cells (19).
Thus, HE4 could promote the malignant biological
behavior of OC cells through interactions with ANXA2, and ANXA2 is
closely associated with P-gp-mediated drug resistance. However, the
mechanisms underlying the potential involvement of HE4 in the
development of drug resistance in OC remain to be elucidated. In
the present study, in order to analyze the role of HE4, ANXA2 and
P-gp in drug resistance of OC, the expression of HE4, ANXA2 and
P-gp were firstly detected in three OC cell lines. Then, the
associations among HE4, ANXA2 and P-gp in ovarian cancer cells were
investigated via immunoprecipitation (IP) and immunofluorescence
(IF). To further explore whether HE4 affects P-gp-mediated drug
resistance in OC and whether the effects require the involvement of
ANXA2, stably transfected cell lines with low expression of HE4 and
ANXA2 were constructed, respectively. A variety of techniques,
including thiazolyl blue tetrazolium bromide (MTT) assay,
immunocytochemistry, western blotting (WB), reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) and
flow cytometry were employed to assess drug sensitivity, expression
and activity of P-gp before and after the transfection. The
expression of P-gp was additionally determined before and after the
addition of active HE4 or ANXA2 protein or antibody in ovarian
cancer cells. Besides experiments in vitro, the association
between HE4 and P-gp was further studied in human samples.
Expression patterns of HE4 and P-gp in drug-resistant and
drug-sensitive EOC tissues were assessed via IHC. The potential
associations between HE4 or P-gp expression with
clinical-pathological parameters and the prognosis of ovarian
cancer were confirmed. To further elucidate the underlying
mechanisms by which the HE4 regulates drug resistance in ovarian
cancer cells, microRNAs (miRNAs/miRs) that bound P-gp gene, ABCB1,
were predicted using bioinformatics methods and their expression
changes before and after the transfection and exposure to different
interventions. In addition, since HE4 may participate in the
development of multidrug resistance of OC at multiple levels, the
common signaling pathways shared by HE4, ANXA2 and P-gp were
selected using the network database. The present study provides a
strong foundation for establishing the role of HE4 in drug
resistance of human EOC.
Materials and methods
Specimen sources
Paraffin specimens of surgical resection from 52
cases of ovarian cancer (33 of low-grade serous carcinoma, 3 of
mucinous cystadenocarcinoma, 3 of ovarian endometrioid carcinoma, 2
of ovarian clear cell carcinoma, and 11 of high-grade serous
carcinoma), were obtained from the Department of Obstetrics and
Gynecology, Shengjing Hospital of China Medical University
(Shenyang, China), between 2006 and 2009. All patients with ovarian
cancer have not undergone chemotherapy before surgery, and
pathology sections were confirmed by experts after hematoxylin and
eosin (HE) staining. The classification of the cancer stage was
according to the International Federation of Gynecology and
Obstetrics (FIGO, 2009) (20).
Among the 52 patients with OC, 12 were at FIGO stage I, 5 cases
were at FIGO stage II, 34 were at FIGO stage III, and 1 was at FIGO
stage IV. All patients were given 6–8 cycles of paclitaxel plus
platinum (TC) systemic chemotherapy after surgery according to
National Comprehensive Cancer Network (NCCN) guidelines (21) and have accepted regular follow-ups
for more than one year after chemotherapy. According to the
guideline of NCCN (the chemotherapy resistant group includes
patients who had a clinical response to the initial TC chemotherapy
program but experienced subsequent relapse, either in the late
stage of chemotherapy or within 6 months after the completion of
chemotherapy, and the chemotherapy sensitive group includes
patients who maintained a clinical response for ≥12 months). The
patients were divided into chemotherapy resistant group (23 cases)
and sensitive group (29 cases). The age range of the chemosensitive
patients was 24 to 78 years (median age, 52 years); The age range
of the chemoresistant patients was 34 to 76 years (median age, 54
years). There was no statistically significant difference in age
between the chemosensitive and chemoresistant patients.
Cell culture and transfection
All ovarian cancer cell lines (CAOV3, SKOV3 and
OVCAR3) were purchased from American Type Culture Collection and
cultured in McCoy's 5A modified medium (Gibco; Thermo Fisher
Scientific, Inc.) with 10% fetal bovine serum (Thermo Fisher
Scientific, Inc.) at 37°C and 5% CO2. Construction of
expression vectors were prepared as previously described (14). The short hairpin RNA (shRNA)
expression vectors for human HE4 or ANXA2 were constructed using
the pSilencer 4.1-CMV neo vector (Ambion; Thermo Fisher Scientific,
Inc.). Cultured CAOV-3 cells were transfected with 3.0 µg plasmid
according to the manufacturer's instructions of the
Lipofectamine® LTX with Plus reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). Briefly, 15 µl Lipofectamine LTX reagent
was diluted in 150 µl Opti-MEM (Invitrogen; Thermo Fisher
Scientific, Inc.), and 16.8 µg plasmid was diluted in 700 µl
Opti-MEM. Then, 150 µl diluted Lipofectamine LTX reagent was mixed
with 150 µl diluted plasmid, and incubated for 5 min at room
temperature. Subsequently, 250 µl complex was added to the cells in
6-well plates and the cells were incubated for 48 h at 37°C. Stable
cell lines CAOV3-HE4-L (expressing HE4 shRNAs) and CAOV3-ANXA2-L
(expressing ANXA2 shRNAs) were screened for 14 days with 800 µg/ml
G418 (Invitrogen; Thermo Fisher Scientific, Inc.; cat. no.
11811023), and the mock group was transfected with mock plasmid
(empty plasmid). Cells were cultured for a further 10 days and the
stable cell lines were used for subsequent experiments. Reagents
and transfection procedures were the same as previously described
(14).
Western blot analysis (WB)
Western blots were prepared as previously described
(22). Primary antibodies included:
Rabbit anti-human HE4 (1:2,000; Abcam; cat. no. ab109298), mouse
anti-human ANXA2 (1:1,000; ProteinTech Group, Inc.; cat. no.
66035-1-Ig), rabbit anti-human P-gp (1:500; Santa Cruz
Biotechnology, Inc.; cat. no. sc-8313), and mouse anti-human
β-actin (1:5,000; Beijing Biosynthesis Biotechnology Co., Ltd.;
cat. no. bsm-33036M). Goat anti-rabbit or mouse secondary
antibodies were used (1:5,000; ZSGB-BIO; cat. nos. ZB-2301 and
ZB-2305).
Immunocytochemistry
Monolayer cell slides were prepared from three types
of ovarian cancer cells, CAOV3, SKOV3 and OVCAR3, during the
exponential growth phase. Reagents were prepared as described for
the immunohistochemistry assay (14), and the primary antibody used was
rabbit anti-human P-gp (1:200; Santa Cruz Biotechnology, Inc.; cat.
no. sc-8313). All procedures were carried out as described on
reagent kit instructions. The samples were visualized using an
inverted light optical microscope (magnification, ×400; Olympus
Corporation).
Co-immunoprecipitation
CAOV3 cells were cultured in McCoy's 5A (Hyclone;
Cytvia; cat. no. SH30270.01) medium with 10% fetal bovine serum
(Thermo Fisher Scientific, Inc.) albumin medium, and incubated in
an incubator at 37°C and 5% CO2. Rabbit anti-human HE4
antibodies (cat. no. ab109298) and rabbit anti-human ANXA2
antibodies (cat. no. ab235939) were purchased from Abcam.
Furthermore, goat anti-human HE4 antibodies (cat. no. sc27570),
rabbit anti-human P-gp antibodies (cat. no. sc-8313) and mouse
anti-human P-gp antibodies (cat. no. sc13131) were purchased from
Santa Cruz Biotechnology, Inc. Mouse anti-human ANXA2 antibodies
(cat. no. 66035-1-Ig) were purchased from ProteinTech Group, Inc.
Details of the experimental procedures were carried out as
previously (22). Three
co-immunoprecipitations were performed: i) ANXA2 protein was
precipitated with mouse anti-human ANXA2 antibodies and detected
with rabbit anti-human ANXA2 antibodies, HE4 protein was detected
with rabbit anti-human HE4 antibodies, and P-gp protein was
detected with rabbit anti-human P-gp antibodies; ii) P-gp protein
was precipitated with rabbit anti-human P-gp antibodies and
detected with mouse anti-human P-gp antibodies, ANXA2 protein was
detected with mouse anti-human ANXA2 antibodies, and HE4 protein
was detected with goat anti-human HE4 antibodies; and iii) and HE4
protein was precipitated with rabbit anti-human HE4 antibodies and
detected with goat anti-human HE4 antibodies, ANXA2 protein was
detected with mouse anti-human ANXA2 antibodies, and P-gp protein
was detected with mouse anti-human P-gp antibodies. The primary
antibody was replaced by rabbit IgG (BIOSS; cat. no. bs-0295P) as a
negative control. An enhanced chemiluminescence reagent was used
(Amersham ECL Prime Western Blotting Detection reagent; Cytvia;
cat. no. RPN2232), and the experiment was repeated three times.
Confocal laser scanning
microscopy
Two immunofluorescence double-labeling experiments
were carried out. For both, protocols strictly followed the
instructions of the reagent suppliers. Firstly, rabbit anti-human
HE4 antibodies (1:50; Abcam; cat. no. ab109298) and mouse
anti-human P-gp antibodies (1:50; Santa Cruz Biotechnology, Inc.;
cat. no. sc13131) were simultaneously incubated on monolayered cell
slides prepared from CAOV3 cells. Then, the secondary antibodies
were applied: Fluorescein isothiocyanate (FITC) green
fluorescence-labeled rabbit IgG fluorescence (1:100; Beijing
Zhongshan Jinqiao Biotechnology Co., Ltd.; cat. no. ZF-0314) and
tetramethylrhodamine (TRITC) red fluorescence-labeled mouse IgG
(1:50; Beijing Zhongshan Jinqiao Biotechnology Co., Ltd.; cat. no.
ZF-0316). Cell nuclei were stained with
4′,6-diamidino-2-phenylindole (DAPI; Beyotime Institute of
Biotechnology; cat. no. C1002). Secondly, mouse anti-human ANXA2
antibodies (1:100; Abcam; cat. no. 66035-1-Ig) and rabbit
anti-human P-gp antibodies (1:50; Biotechnology, Inc.; cat. no.
sc-8313) were simultaneously incubated on monolayered cell slides
prepared from CAOV3 cells. After that, the secondary antibodies
were applied: FITC green fluorescence-labeled rabbit IgG
fluorescence (1:100) and TRITC red fluorescence-labeled mouse IgG
(1:50). Cell nuclei were stained with DAPI. Double-labeled
immunofluorescence samples were captured by fluorescence confocal
microscopy (magnification, ×400).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RNA was extracted from pretreated cells with RNAiso
Plus reagent (Takara Biotechnology Co., Ltd.) and
reverse-transcribed by a Prime-Script RT reagent kit (Takara
Biotechnology Co., Ltd.). The reaction conditions for mRNA were
37°C for 15 min, 85°C for 5 sec, 4°C for 5 min; and for miRNA were
37°C for 60 min, 85°C for 5 min. Real-time PCR and Ct value
analysis were performed with a Roche LightCycler system. The primer
sequences were as follows: HE4 forward, 5′-CCGACAACCTCAAGTGCTG-3′;
HE4 reverse, 5′-CGAGCTGGGGAAAGTTAATG-3′; ANXA2 forward,
5′-TTGCCATTAAACTCACATGAAGTAG-3′; ANXA2 reverse,
5′-GCACAGGATTTCGTGAACAGTAG-3′; P-gp forward,
5′-TGGGGCTGGACTTCCTCTCATGATGC-3′; P-gp reverse,
5′-GCAGCAACCAGCACCCCAGCACCAAT-3′; β-actin forward,
5′-ACTTAGTTGCGTTACACCCTTTCTTG-3′; β-actin reverse,
5′-TGTCACCTTCACCGTTCCAGTTTT-3′; hsa-miR-129-5p,
5′-CTTTTTGCGGTCTGGGCTTGC-3′; hsa-miR-4282,
5′-TAAAATTTGCATCCAGGA-3′; hsa-miR-4778-5p,
5′-AATTCTGTAAAGGAAGAAGAGG-3′; hsa-miR-4708-3p,
5′-AGCAAGGCGGCATCTCTCTGAT-3′; hsa-miR-4432,
5′-AAAGACTCTGCAAGATGCCT-3′; RNU6-2 forward,
5′-CACCACGTTTATACGCCGGGTG-3′; and RNU6-2 reverse,
5′-CTGGTGTCGTGGAGTCGGCAATTC-3′.
The qPCR amplification conditions were as follows:
denaturation at 95°C for 30 sec, extension at 95°C for 5 sec, and
annealing at 60°C for 31 sec, total of 40 cycles. The reaction
mixture contained SYBR Premix Ex Taq™ (X2) 10 µl, PCR Forward
Primer (10 µmol/l) 0.4 µl, PCR reverse primer (10 µmol/l) 0.4 µl,
cDNA 2 µl, dH2O 7.2 µl. β-actin or RNU6-2 was used as the internal
reference. Once the amplification was completed, the melting curve
was analyzed. The change of target gene expression level was
calculated using the 2−ΔΔCq method (23).
MTT assay
All cells were inoculated at 2,000 cells/well in
96-well plates, with a blank well set aside. Increasing doses of
adriamycin (Meilun Biotechnology Co., Ltd.; cat. no. MB1087-S) were
added after 6 h of cell culture, and this was regarded as the ‘0
time points’. After 24 h, cell viability assays were performed by
adding MTT solution (10 µl; 5 µg/ml; Beijing Solarbio Science &
Technology Co., Ltd.) and incubated at 37°C/5% CO2 for 4
h. The culture medium was removed and 150 µl DMSO (Beyotime
Institute of Biotechnology; cat. no. ST038) was added. After
shaking for 10 min, the OD was measured using a universal
microplate reader at a wavelength of 490 nm. For each treatment,
three parallel wells were settled for statistical analysis. The
experiment was repeated three times.
Assessment of P-gp expression on the
cell surface
To analyze the P-gp expression level on the cell
surface, CAOV3, CAOV3-HE4-L, CAOV3-A2-L, or CAOV3-Mock (empty
plasmid control group) cells were digested with 0.25% trypsin
(Gibco; Thermo Fisher Scientific, Inc.; cat. no. 25200-072) and
counted. Cells (2×105) were rinsed and dissolved with
ice-cold PBS containing 1% BSA (Biological Industries; cat. no.
04-011-1A/B) and incubated with 5 µl of the PE-labeled monoclonal
anti-(human P-gp) antibody UIC2 (eBioscience; Thermo Fisher
Scientific, Inc.; cat. no. 12-2439) at 4°C for 30 min in darkness.
Then, cells were washed and resuspended in cold PBS. The binding
affinity of the PE-labeled P-gp antibody was measured using flow
cytometer.
Measurement of adriamycin accumulation
by flow cytometry
CAOV3, CAOV3-HE4-L, CAOV3-A2-L, or CAOV3-Mock (empty
plasmid control group) cells were inoculated onto 6-well plates at
a density of 5×105 cells per well. After 24 h, the cells
were pretreated with complete medium containing or not containing
verapamil (50 µM; Nanjing SenBeiJia Biological Technology Co.,
Ltd.; cat. no. 100223) for 1 h. After that, cells were incubated
with 10 µM adriamycin in culture medium in the dark for 60 min at
37°C, 5% CO2. After incubation, the medium was removed,
and all monolayers were washed three times with ice-cold PBS. Cells
were re-suspended and the fluorescence intensity of intracellular
adriamycin was determined by flow cytometry.
Antibody blocking and exogenous active
protein supplementation tests
CAOV3 or CAOV3-A2-L cells during the log phase were
selected and single-cell suspensions were prepared. The dialysis
membrane with medium-length molecular retention of 10 kDa was used
to remove sodium azide from the antibodies. IgG (BIOSS; cat. no.
bs-0295P), HE4 exogenous active protein (100 ng/ml; Novoprotein;
cat. no. C550A), ANXA2 exogenous active protein (10 ng/ml;
Novoprotein; cat. no. C205A), HE4 antibodies (10 µg/ml; Abcam; cat.
no. ab109298), ANXA2 antibodies (10 µg/ml; ProteinTech Group, Inc.;
cat. no. 66035-1-Ig), or ANXA2 antibodies (10 µg/ml; ProteinTech
Group, Inc.; cat. no. 66035-1-Ig) with HE4 exogenous active
protein, was added to the adherent cells. The cells were incubated
at 37°C for 0, 12 and 24 h, and then the protein and mRNA were
extracted for analysis. The experiments were repeated three times
and the average value was calculated.
IHC
The expression of HE4 and P-gp was detected using
streptavidin-peroxidase (SP) and Strept Avidin-Biotin Complex
(SABC) IHC staining, using rabbit anti-human HE4 antibodies (1:40;
Abcam; cat. no. ab109298) and rabbit anti-human P-gp antigen
antibodies (1:50; Santa Cruz Biotechnology, Inc.; cat. no.
sc-8313). All other reagents were provided by the central
laboratory of Shengjing Hospital of China Medical University. Each
experiment included low-grade serous carcinoma, mucinous
cystadenocarcinoma, endometrioid carcinoma, clear cell carcinoma,
and high-grade serous carcinoma), as well as both positive and
negative control sections. The positive control sections for HE4
were epididymal carcinoma tissues, and for P-gp, were P-gp positive
colon carcinoma tissues. For negative controls, PBS was substituted
for primary antibodies. Staining protocols followed the
manufacturer's instructions for the reagent kit as previously
described (24).
Immunohistochemistry results were interpreted as
previously described (21). Samples
were marked positive when the cell membrane or cytoplasm appeared
brown or yellow. The 52 OC samples were separated into high
expression (++/+++) and low expression (−/+) groups according to
levels of HE4 and P-gp staining. Each sample was independently
assessed by two people, and by a third person when inconsistencies
arose.
Bioinformatics methods of predicting
miRNAs that bound to the P-gp gene, ABCB1
Four major miRNA predictive analysis tools, miRanda
(http://www.microrna.org/microrna/home.do), TargetScan
(http://www.targetscan.org), DIANA
(http://www.microrna.gr) and miRDB (http://www.mirdb.org/miRDB) were used to predict the
miRNAs that bound to the P-gp gene, ABCB1.
Data collection from The Cancer Genome
Atlas (TCGA) database and Gene Set Enrichment Analysis (GSEA)
Ovarian cancer data was downloaded from GDC
Application Programming Interface (API; http://gdc.cancer.gov/developers/gdc-application-programming-interface-api)
and prescreened, and 374 tumor samples were included in the present
study. Samples were sorted according to the expression level of
HE4, ANXA2 or ABCB1, from low to high, and equally aliquoted into
four parts: The first 25% of the samples were the low expression
group; the last 25% of samples were the high expression group. GSEA
3.0 (https://www.gsea-msigdb.org/gsea/msigdb/index.jsp) was
used to analyze data. C2.cp.kegg.v6.1.symbols.gmt data cluster was
downloaded from the Molecular Signatures Database data bank on the
GSEA website. Enrichment analysis was performed on the sorted
samples using default weighted enrichment statistics. Random
assortment times were set to 1,000. For GSEA analysis, gene
clusters with false discovery rates (FDR) <0.25 and P<0.05
were considered as significantly enriched genes.
Statistical analysis
SPSS 21.0 software (IBM Corp.) was used for
statistical analysis, with all data represented by the mean ±
standard deviation. χ2 test, the one-way ANOVA and
two-way ANOVA tests followed by a Tukey's post hoc test were
employed. Correlated factors were analyzed with multivariate
logistic regression and their associations were analyzed with
Fisher's exact test. Prognosis and overall survival were analyzed
with the multivariate logistic regression, Kaplan-Meier survival
analysis and a log-rank test. P<0.05 was considered to indicate
a statistically significant difference.
Results
Expression patterns of HE4, ANXA2 and
P-gp in different OC cell lines
In order to study the role of HE4, ANXA2 and P-gp in
drug resistance of OC, the expression of HE4, ANXA2 and P-gp were
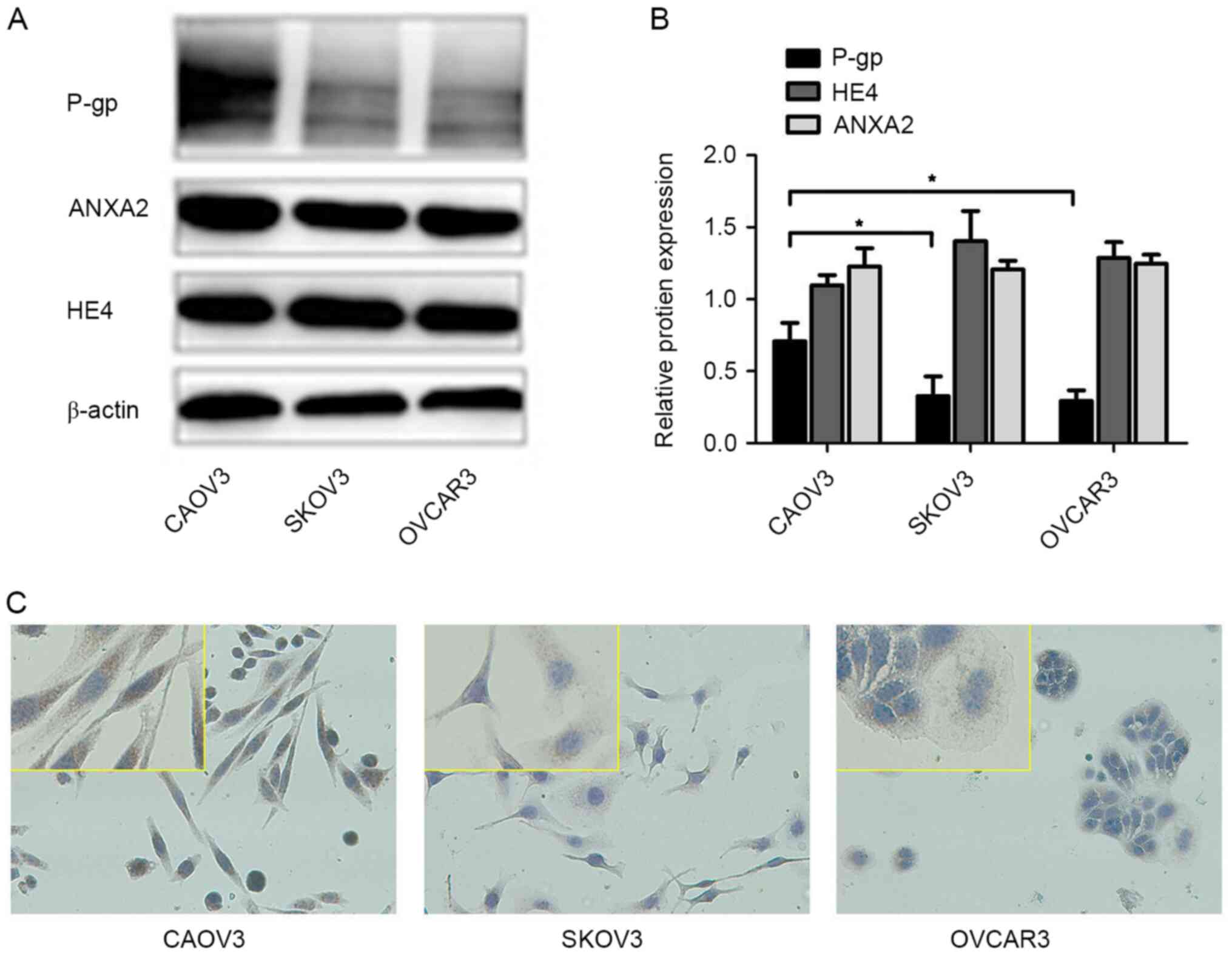
firstly detected in three OC cell lines. The results of WB showed
that HE4 and ANXA2 were highly expressed and there was no
significant difference among the CAOV3, SKOV3 and OVCAR3 cell lines
(Fig. 1A and B). Both WB and
immunocytochemistry experiments suggested greater P-gp expression
in CAOV3 compared with OVCAR3 and SKOV3 cells (Fig. 1). The average optical densities of
ICC were measured under a microscope with image processing, and are
presented as the mean ± standard deviation of three separate
experiments, and the results were statistically significant
(P<0.05; Table I). The
forementioned results suggested that HE4, ANXA2 and P-gp were
widely expressed in OC cell lines, and CAOV3 with the highest P-gp
expression was used for further experiments.
 | Table I.Average optical densities of
immunocytochemistry in different groups. |
Table I.
Average optical densities of
immunocytochemistry in different groups.
| Cell lines | Integrated optical
density |
|---|
| CAOV3 |
15.261±0.421a,b |
| SKOV3 | 5.698±0.499 |
| OVCAR3 | 7.619±0.787 |
Interaction of HE4, ANXA2 and P-gp in
CAOV3 cells
The associations among HE4, ANXA2 and P-gp were
investigated in CAOV3. Co-expression of HE4 and ANXA2, as well as
P-gp and ANXA2, were detected via immunoprecipitation analysis. No
direct interactions between HE4 and P-gp were detected (Fig. 2A). Colocalization patterns of HE4,
ANXA2 and P-gp in CAOV3 cells were assessed via double-labeling
immunofluorescence confocal experiment. HE4 labeled with green
fluorescence was mainly localized in the cell membrane and
cytoplasm, similar to red fluorescence-labeled P-gp, red
fluorescence-labeled ANXA2 and green fluorescence-labeled P-gp.
Blue fluorescence represents DAPI staining of nuclei. Yellow
fluorescence from the overlapping of red and green fluorescence
confirmed the colocalization of HE4 or ANXA2 with P-gp (Fig. 2B). Although no direct interactions
were detected between HE4 and P-gp via immunoprecipitation
analyses, colocalization of HE4/P-gp and ANXA2/P-gp was determined
via immunofluorescent staining, indicative of interactions between
HE4-ANXA2 complex and P-gp.
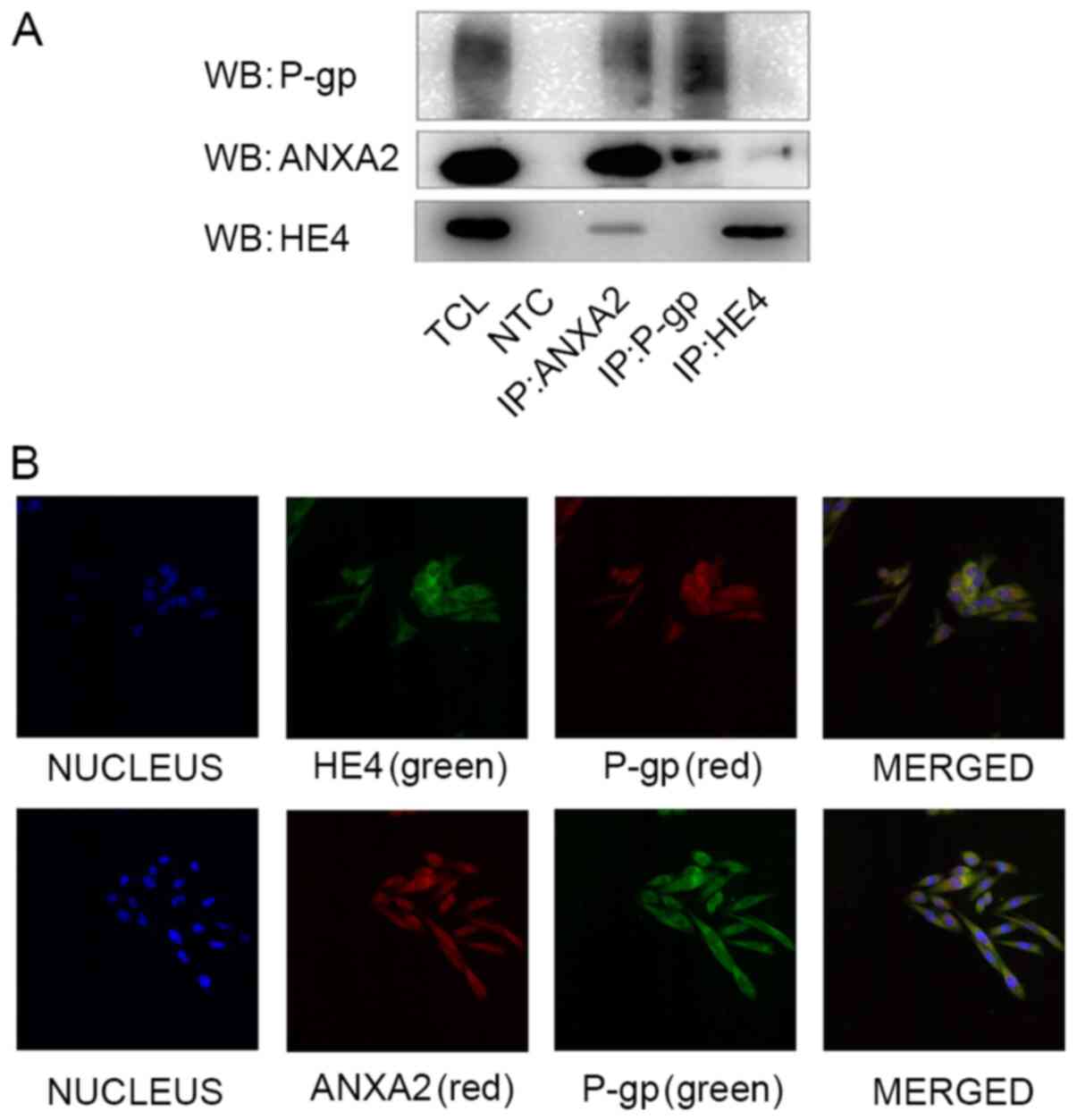 | Figure 2.Interaction of HE4, ANXA2 and P-gp in
CAOV3 cells. (A) Protein lysates of CAOV3 cells were precipitated
using HE4, ANXA2 and P-gp-specific antibodies. WB revealed
co-expression of HE4 with ANXA2, and ANXA2 with P-gp, respectively.
(B) Double-labeling immunofluorescence showed the colocalization of
HE4 or ANXA2 with P-gp in CAOV3 cells. Magnification, ×400. HE4;
human epididymis protein 4; P-gp, P-glycoprotein; WB, western
blotting; TCL, total cell lysate; NTC, negative control, the
primary antibody was replaced by rabbit IgG; IP,
immunoprecipitate. |
Interactions between HE4 and ANXA2
enhances P-gp expression and promotes resistance to adriamycin in
CAOV3 cells
In order to further explore whether HE4 affects
P-gp-mediated drug resistance in OC and whether the effects require
the involvement of ANXA2, stably transfected cell lines with low
expression of HE4 (CAOV3-HE4-L) and ANXA2 (CAOV3-A2-L),
respectively, were constructed. WB and RT-qPCR assays confirmed
decreased HE4 expression in CAOV3-HE4-L, as well as decreased ANXA2
expression in CAOV3-A2-L cells, both accompanied with decreased
P-gp expression in protein and mRNA levels (Fig. 3A-C). Compared with the untreated
(CAOV3) and mock groups (CAOV3-Mock; empty plasmid control group),
MTT assay showed a significant increase in the proliferation
inhibition rate and decrease in IC50 value in
CAOV3-HE4-L and CAOV3-A2-L cells 24 h after adriamycin treatment
(P<0.05; Fig. 3D; Table II). Flow cytometry data showed a
significant decrease in P-gp expression on the cell membrane in
CAOV3-HE4-L and CAOV3-A2-L cells (P<0.05; Fig. 3E). Simultaneously, intracellular
drug accumulation after 1 h adriamycin treatment was greater in
CAOV3-HE4-L and CAOV3-A2-L compared with CAOV3 and CAOV3-Mock
(empty plasmid control group) cells (Fig. 3G). Upon pre-treatment of cells with
50 µM verapamil (VER), a specific P-gp inhibitor, intracellular
adriamycin accumulation in CAOV3 and CAOV3-Mock (empty plasmid
control group) cells was markedly increased, compared with that in
untreated cells (all P<0.05; Fig. 3F
and G). However, VER exerted no significant effects on
adriamycin accumulation in CAOV3-HE4-L and CAOV3-A2-L cells
(Fig. 3F and G). The aforementioned
results suggest that drug resistance to adriamycin and the
expression and activity of P-gp were significantly decreased
accompanied by decreased HE4 or ANXA2 expression after transfection
in CAOV3 cells (Fig. 3A-G). WB and
RT-qPCR assays confirmed increased P-gp protein and mRNA expression
in CAOV3 cells treated with HE4 active protein (group 2) or ANXA2
active protein (group 3), and decreased P-gp expression in CAOV3
cells treated with HE4 antibody (group 4) or ANXA2 antibody (group
5) within 24 h (Fig. 3H-J). In
addition, combined treatment with ANXA2 antibody and active HE4
protein (group 6) resulted in a significant decrease in P-gp
protein and mRNA levels in CAOV3 cells within 24 h (all P<0.05)
However, P-gp protein and mRNA expression patterns in CAOV3-A2-L
cells treated with exogenous active HE4 protein were not rescued
within 24 h (group 8) (P>0.05). P-gp protein and mRNA expression
in untreated CAOV3 cells (group 1), untreated CAOV3-A2-L cells
(group 7) and cells treated with IgG (group 9) were not
significantly altered across the 24 h time point (P>0.05;
Fig. 3H-J). These results showed
that the expression and activity of P-gp were associated with HE4,
which may require the involvement of ANXA2.
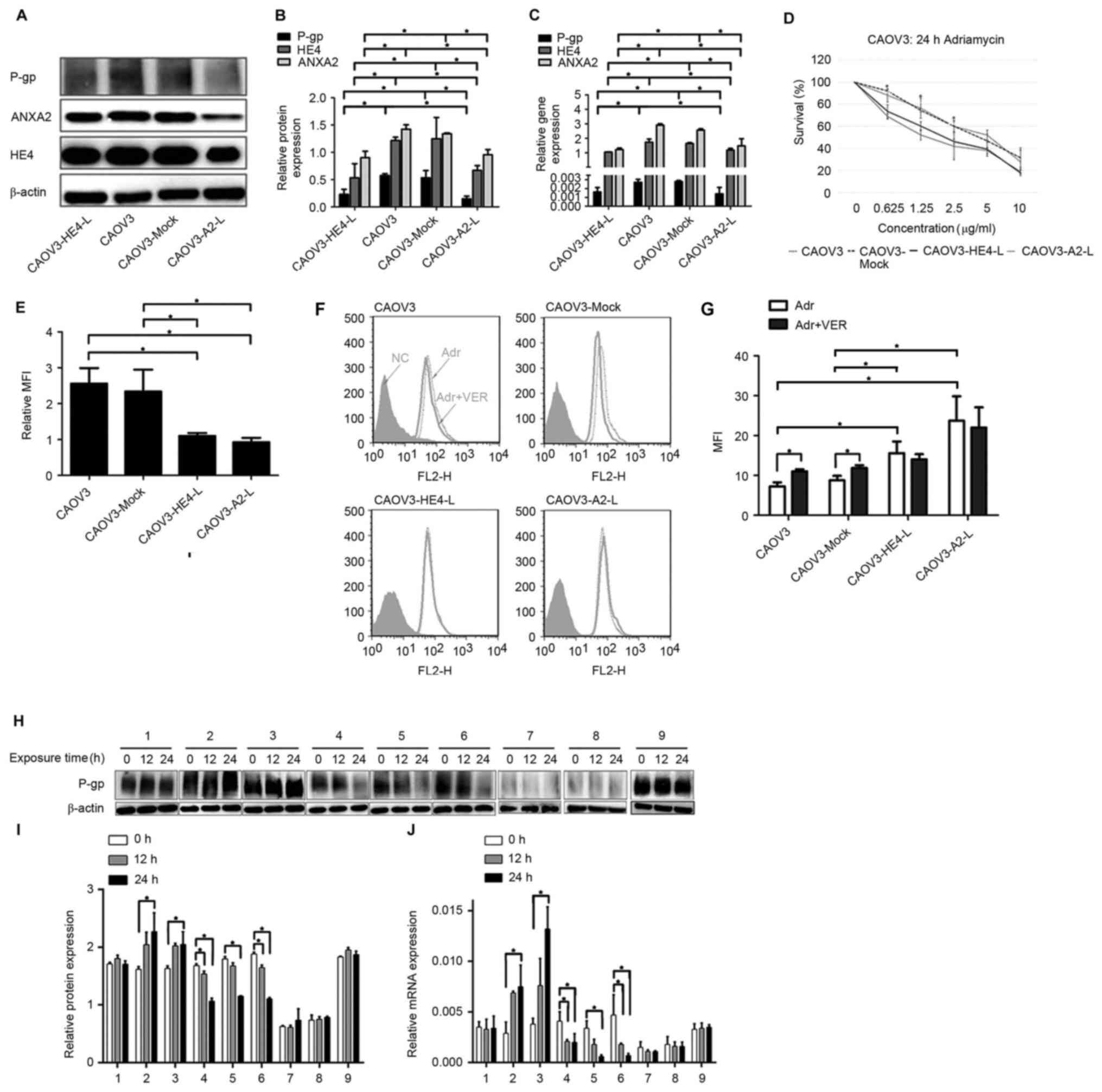 | Figure 3.Interaction of HE4 with ANXA2
enhances P-gp expression and promotes resistance to adriamycin in
CAOV3 cells. (A) Expression of HE4, ANXA2 and P-gp before and after
transfection, detected by western blotting. (B) Comparison of HE4,
ANXA2 and P-gp protein levels before and after transfection. (C)
Expression of HE4, ANXA2 and P-gp before and after transfection,
detected by RT-qPCR assays. (D) Cells were treated with 0–10 µg/ml
adriamycin for 24 h before and after transfection, at which time
the cells were subjected to MTT assay to measure viability. Results
are displayed as percent survival of vehicle-treated cells. Error
bars represent the standard deviation of biological replicates. (E)
Flow cytometry detection of the expression of P-gp on the cell
membrane before and after transfection. (F) Measurement of
adriamycin accumulation by flow cytometry before and after
transfection, cells of different groups were pretreated with
complete medium containing or not containing verapamil (50 µM) for
1 h. After pretreatment, cells were incubated with adriamycin in
culture medium. (G) Measurement of MFI of adriamycin by flow
cytometry in cells before and after transfection. (H) Expression of
P-gp in cells before and after treatment by different antibodies or
active protein at different time points, detected by western
blotting. Group 1, CAOV3 cells; group 2, CAOV3 cells treated with
HE4 active protein; group 3, CAOV3 cells treated with ANXA2 active
protein; group 4, CAOV3 cells treated with HE4 antibody; group 5,
CAOV3 cells treated with ANXA2 antibody; group 6, CAOV3 cells
treated with HE4 active protein and ANXA2 antibody; group 7,
CAOV3-A2-L cells; group 8, CAOV3-A2-L cells treated with HE4 active
protein; and group 9, CAOV3 cells treated with IgG. (I) Comparison
of P-gp protein levels before and after treatment by different
antibodies or active protein at different time points. The cells of
each group were treated as same as (H). (J) Expression of P-gp in
cells before and after treatment by different antibodies or active
protein at different time points, detected by RT-qPCR assays. The
cells of each group were treated as same as (H). *P<0.05. HE4;
human epididymis protein 4; P-gp, P-glycoprotein; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; MFI, mean
fluorescence intensity; VER, verapamil. |
| Table II.IC50 of adriamycin in
different groups. |
Table II.
IC50 of adriamycin in
different groups.
| Cell lines | IC50,
µg/ml |
|---|
| CAOV3 |
4.36±0.433a,b |
| CAOV3-Mock |
4.29±0.366a,b |
| CAOV3-HE4-L | 2.18±0.166 |
| CAOV3-A2-L | 1.72±0.238 |
Expression of HE4 and P-gp in OC
tissues from different groups
Besides experiments in vitro the association
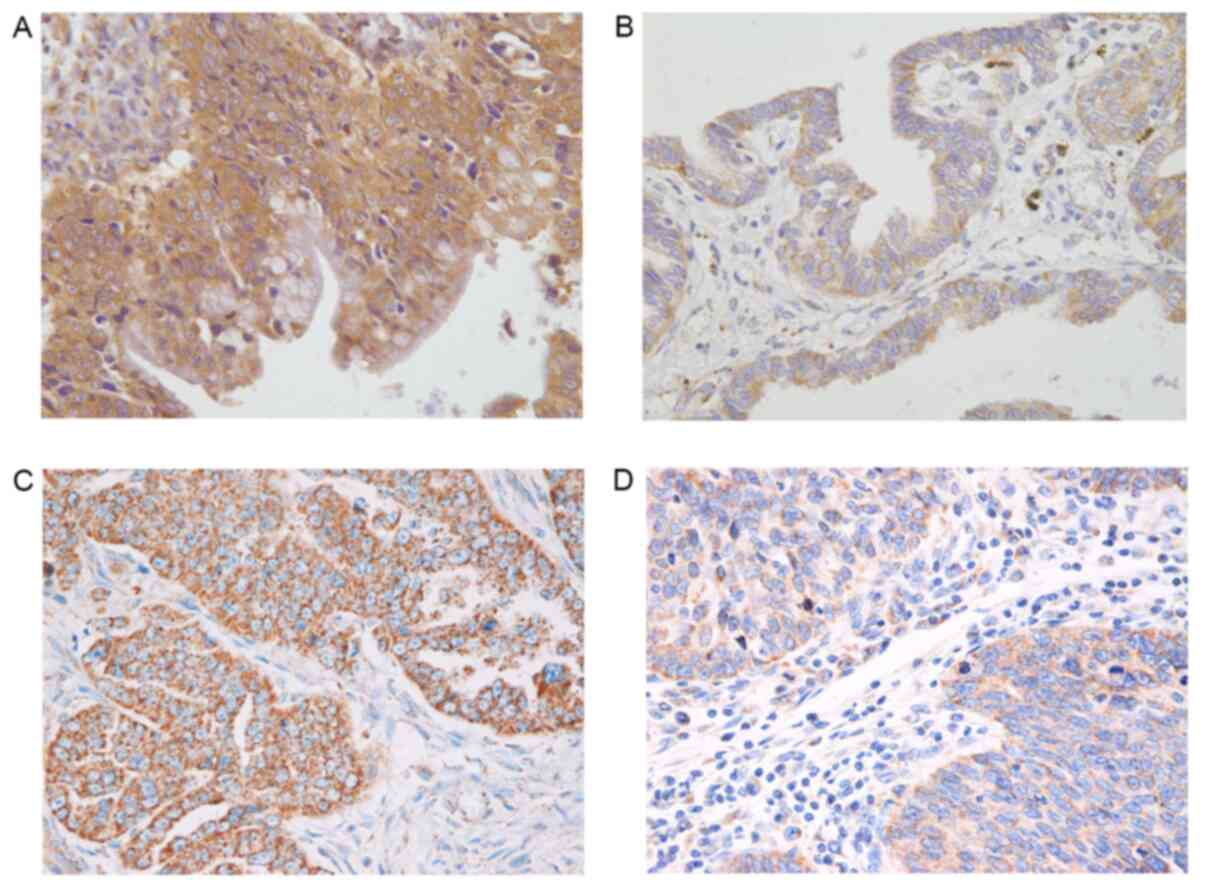
between HE4 and P-gp was further studied in human samples. HE4
staining was mainly detected in the cell membrane and cytoplasm in
all 52 cases of malignant ovarian tumors. The positive rate of HE4
from the drug-resistant group was 95.65%, which was significantly
higher than that (65.51%) of the drug-sensitive group.
Additionally, the expression level intensity of HE4 of the
drug-resistant group was significantly higher compared with that of
the drug-sensitive group. In the drug-resistant group, 17 cases
showed strong positive HE4 staining (73.91%), while there were only
8 cases with strong positive HE4 staining in the drug-sensitive
group (All P<0.05; Fig. 4A and
B; Tables III and IV).
| Table III.Expression intensity of HE4 and P-gp
in different groups. |
Table III.
Expression intensity of HE4 and P-gp
in different groups.
| Groups | HE4 | P-gp |
|---|
| Resistance
group | 0.2642±1.2220 |
0.2608±0.09394 |
| Sensitive
group | 0.1920±0.9621 | 0.2088±0.8579 |
| P-value | 0.025a | 0.043a |
| Table IV.Expression of HE4 in different
groups. |
Table IV.
Expression of HE4 in different
groups.
| Groups | Cases | − | + | ++ | +++ | High expression
cases, n | High expression
rate, % | Positive cases,
n | Positive
rate,% |
|---|
| Resistance
group | 23 | 1 | 5 | 5 | 12 | 17 | 73.91 | 22 | 95.65 |
| Sensitive
group | 29 | 10 | 11 | 5 | 3 | 8 | 27.59 | 19 | 65.51 |
P-gp staining was also mainly detected in the cell
membrane and cytoplasm. The percentage of positively stained
samples was significantly higher in the drug-resistant group
compared with that of drug-sensitive group (95.65 vs. 75.86%).
Moreover, the expression intensity of P-gp in the drug-resistant
group was significantly higher compared with that of the
drug-sensitive group. In the drug-resistant group, 18 patients
showed strong positive staining (78.26%). In the drug-sensitive
group, strong positive staining was only detected in 12 cases (All
P<0.05; Fig. 4C and D; Tables III and V).
| Table V.Expression of P-glycoprotein in
different groups. |
Table V.
Expression of P-glycoprotein in
different groups.
| Groups | Cases | − | + | ++ | +++ | High expression
cases, n | High expression
rate, % | Positive cases,
n | Positive
rate,% |
|---|
| Resistance
group | 23 | 1 | 4 | 8 | 10 | 18 | 78.26 | 22 | 95.65 |
| Sensitive
group | 29 | 7 | 10 | 9 | 3 | 12 | 41.37 | 22 | 75.86 |
Association between expression of HE4
and P-gp with clinical pathological-parameters
Expression patterns of HE4 were similar to those of
P-gp. The high expression rate of HE4 in the late stages group of
FIGO was 57.14%, which was higher than that of the early stages
(29.41%); and the lymph node metastasis group (60%) was higher than
that of the non-lymph node metastasis group (36%) (P>0.05). The
high expression rate of HE4 in the postoperative residual tumor ≥1
cm group (52.63%) was higher than that of the residual tumor <1
cm group (45.45%), although this difference was not statistically
significant (P>0.05; Table
VI).
| Table VI.Expression of HE4 and P-gp in ovarian
cancer tissue. |
Table VI.
Expression of HE4 and P-gp in ovarian
cancer tissue.
|
|
| HE4 |
| P-gp |
|
|---|
|
|
|
|
|
|
|
|---|
|
Characteristics | Cases | High, n | % | P-value | High, n | % | P-value |
|---|
| FIGO stage |
|
|
| 0.060 |
|
| 0.093 |
|
I–II | 17 | 5 | 29.41 |
| 7 | 41.18 |
|
|
III–IV | 35 | 20 | 57.14 |
| 23 | 65.71 |
|
| Lymph node
metastasis |
|
|
| 0.317 |
|
| 0.317 |
|
Yes | 5 | 3 | 60 |
| 4 | 80 |
|
| No | 25 | 9 | 36 |
| 14 | 56 |
|
| Histologic
subtype |
|
|
| 0.051 |
|
| 0.904 |
|
Low-grade serous
carcinoma | 33 | 20 | 60.61 |
| 20 | 60.61 |
|
|
Mucinous
cystadenocarcinoma | 3 | 1 | 33.33 |
| 1 | 33.33 |
|
|
Endometrioid carcinoma | 3 | 0 | 0 |
| 2 | 66.67 |
|
| Clear
cell carcinoma | 2 | 0 | 0 |
| 1 | 50 |
|
|
High-grade serous
carcinoma | 11 | 3 | 27.27 |
| 6 | 54.55 |
|
| Postoperative
residual lesions |
|
|
| 0.618 |
|
| 0.235 |
| <1
cm | 33 | 15 | 45.45 |
| 17 | 51.51 |
|
| ≥1
cm | 19 | 10 | 52.63 |
| 13 | 68.42 |
|
The high expression rate of P-gp was 65.71% in the
late stages (FIGO stage III/IV) relative to 41.18% in the early
stages (FIGO stage I/II), 80.0% (4/5) in the lymph node metastasis
group relative to the non-lymph node metastasis group (56%), and
68.42% in the residual tumor ≥1 cm group relative to the residual
tumor <1 cm group (51.51%). However, these differences were not
statistically significant (all P>0.05; Table VI).
Associations between HE4 and P-gp
expression in OC tissues
Among 52 OC tissue cases, 38 showed double-positive
staining for HE4 and P-gp, while 5 cases showed that neither
existed. Cases with highly expressed HE4 concomitantly showed high
P-gp expression, suggesting associations between the two proteins
(P=0.008; Table VII).
| Table VII.Association between HE4 and P-gp
expression. |
Table VII.
Association between HE4 and P-gp
expression.
|
|
| P-gp |
|---|
|
|
|
|
|---|
| HE4 | Cases, n | Negative, n | Positive, n |
|---|
| Negative | 10 | 5 | 5 |
| Positive | 42 | 4 | 38 |
| Total cases | 52 | 9 | 43 |
Associations of P-gp and HE4
expression with prognosis
Multiple factors logistic regression analysis was
carried out. FIGO stage, residual tumor volume, drug resistance and
expression of HE4 and P-gp were used as dependent variables. The
analysis revealed that FIGO stage III–IV and high expression of HE4
and P-gp were independent risk factors for poor prognosis of
epithelial ovarian cancer (Table
VIII).
| Table VIII.Ovarian cancer prognostic
multi-factor analysis. |
Table VIII.
Ovarian cancer prognostic
multi-factor analysis.
| Variables | B | SE | Wald | df | P-value | OR | 95.0%
CI |
|---|
| HE4 | 1.022 | 0.360 | 8.058 | 1 | 0.005 | 2.780 | 1.372~5.632 |
| P-gp | 0.945 | 0.357 | 7.003 | 1 | 0.008 | 2.572 | 1.278~5.177 |
| Surgical stage | 1.085 | 0.451 | 5.774 | 1 | 0.016 | 2.958 | 1.221~7.164 |
| Postoperative
residual lesions | 0.024 | 0.420 | 0.003 | 1 | 0.954 | 1.024 | 0.450~2.333 |
| Drug
resistance | 0.598 | 0.531 | 1.268 | 1 | 0.260 | 1.819 | 0.642~5.150 |
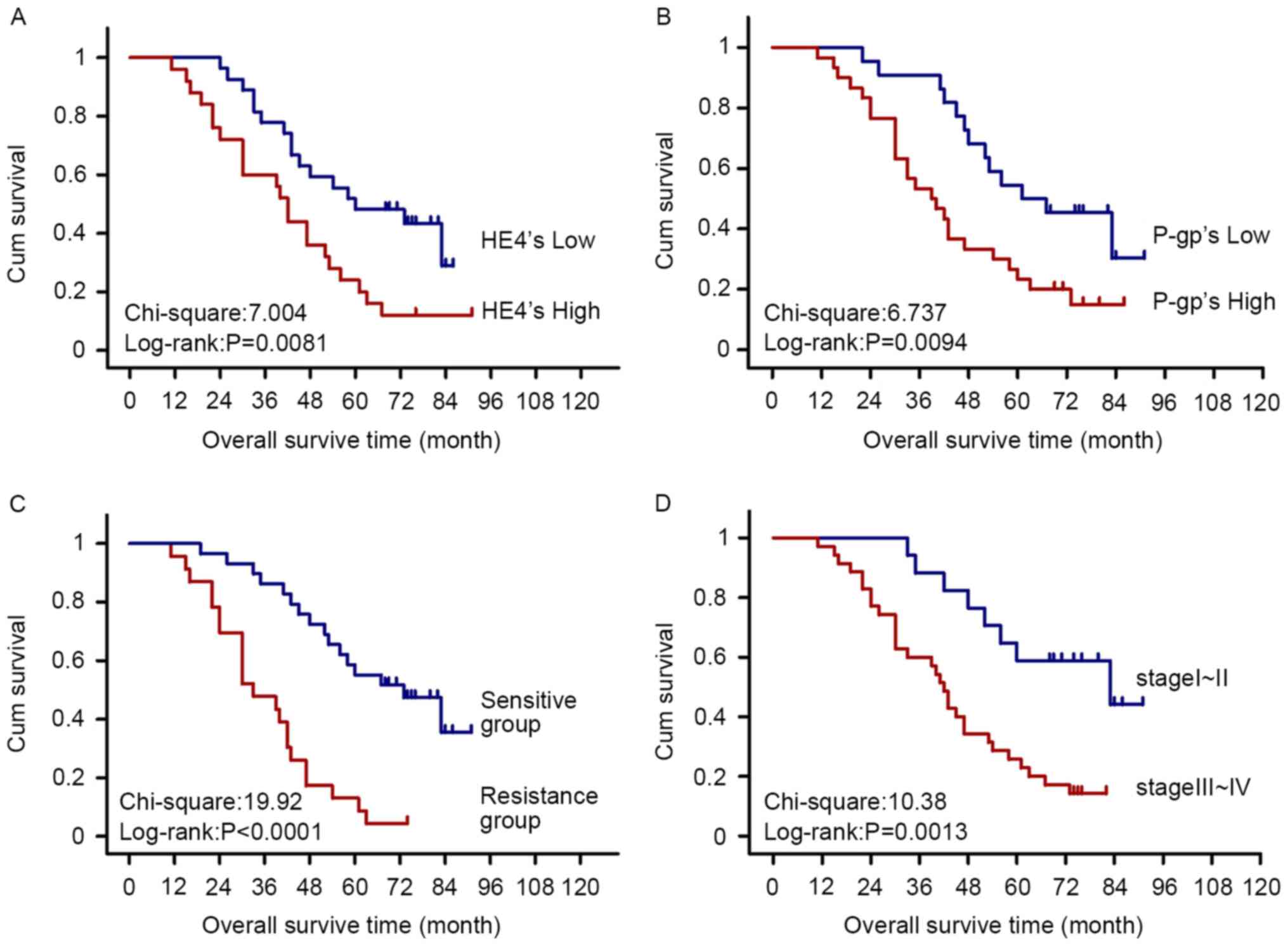
Survival analysis
Among the 52 cases of ovarian cancer, 43 were
followed up and 9 were lost (until May 2015). The results of
Kaplan-Meier and log-rank tests showed significantly higher
mortality in HE4 and P-gp high expression, late FIGO stage (III–IV)
and drug-resistant groups compared with that in HE4 and P-gp low
expression, early FIGO stage (I–II) and chemotherapy-sensitive
groups, respectively (Fig. 5).
Prediction of miRNAs regulating the
3′-UTR of ABCB1
Furthermore, in order to elucidate the detailed
mechanisms by which the HE4-ANXA2 complex regulates P-gp-mediated
drug resistance in OC cells, miRNAs that bound the P-gp gene,
ABCB1, were predicted using bioinformatics methods. The number of
miRNAs with the potential to bind the 3′-UTR region of ABCB1 was
14, 376, 26 and 13, respectively, as predicted by different
algorithms of the four databases. Five miRNAs were intersected by
three of the four websites, specifically, hsa-miR-129-5p,
hsa-miR-4282, hsa-miR-4778-5p, hsa-miR-4708-3p and hsa-miR-4432
(Fig. 6).
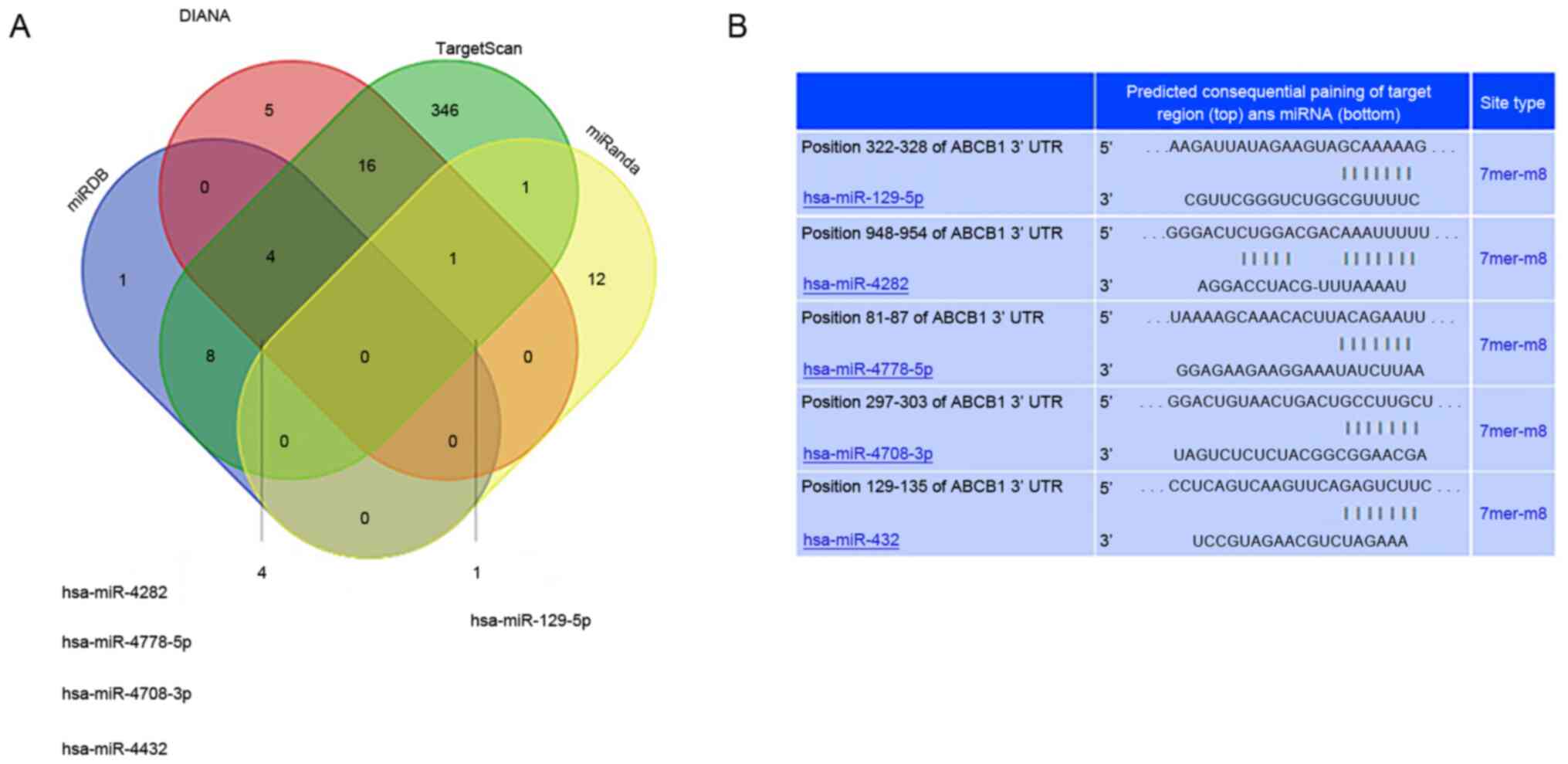 | Figure 6.Prediction of miRNAs regulating the
3′URT region of ABCB1. (A) Venn diagram depicting the number of
predicted miRNAs of ABCB1 from miRanda, TargetScan, DIANA and
miRDB. The overlaps indicate the numbers of miRNAs predicted by
more than one algorithm. miRanda, TargetScan, DIANA and miRDB
predicted a total of 14, 376, 26, and 13 microRNAs, respectively.
(B) TargetScan was used to predict the interaction between miRNAs
and 3′URT region of ABCB1. microRNA/miR, microRNAs; UTR,
untranslated region. |
Inhibitory regulation of
hsa-miR-129-5p by HE4 via interactions with ANXA2
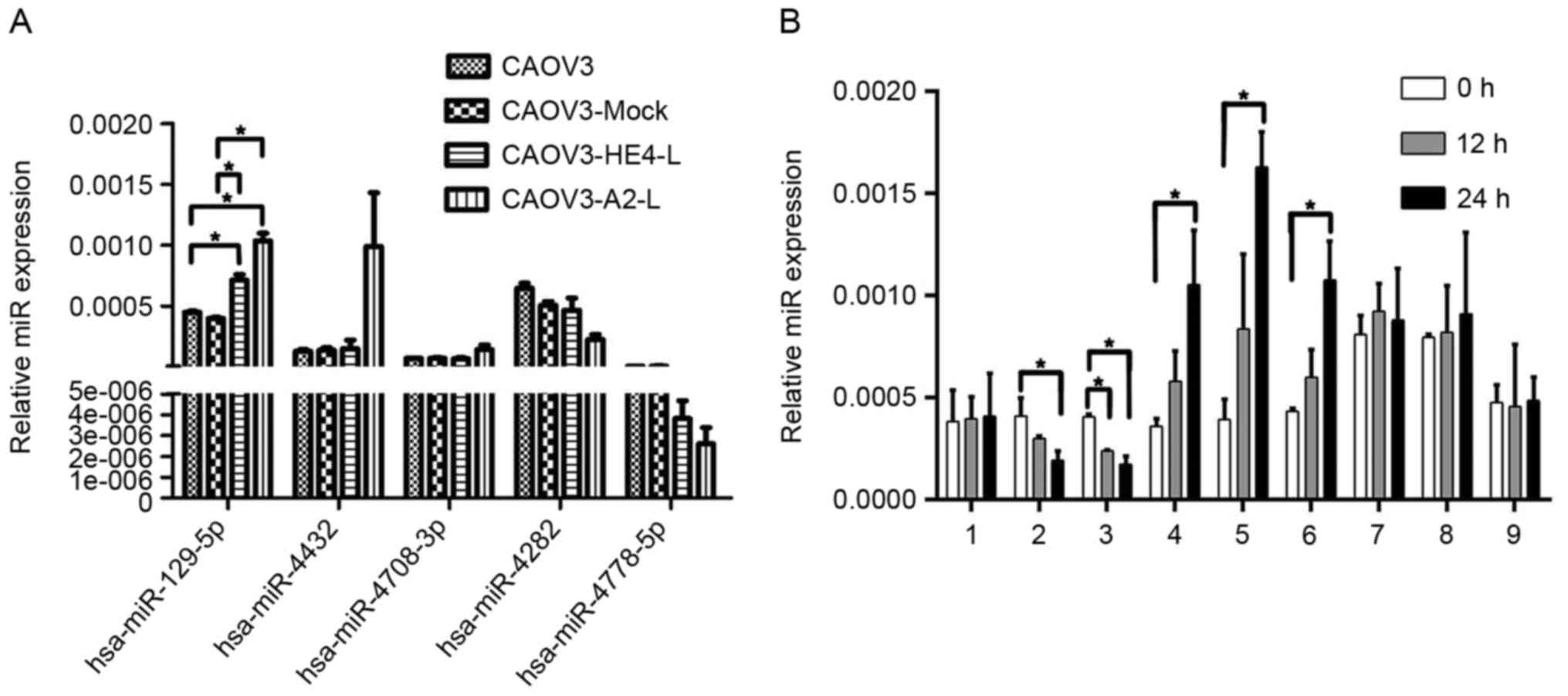
RT-qPCR was then employed to investigate the
associations among HE4, ANXA2 and the five miRNAs in CAOV3-HE4-L
and CAOV3-A2-L cells. Data from a RT-qPCR assay showed increased
expression of hsa-miR-129-5p in both CAOV3-HE4-L and CAOV3-A2-L
cells, compared with CAOV3 and CAOV3-Mock group (empty plasmid
control group), while no significant differences in hsa-miR-4708-3p
expression were observed. Levels of hsa-miR-4282 and
hsa-miR-4778-5p were slightly decreased in CAOV3-HE4-L and
CAOV3-A2-L cells, but these differences were not significant. The
expression of hsa-miR-4432 remained constant in CAOV3-HE4-L cells
but increased in CAOV3-A2-L cells, albeit not to a significant
extent (Fig. 7A). Among the five
miRNAs, only hsa-miR-129-5p was significantly increased,
accompanied by decreased HE4 or ANXA2 expression and P-gp
expression in CAOV3-HE4-L and CAOV3-A2-L cells, indicating that the
expression of hsa-miR-129-5p is consistent with those of HE4, ANXA2
and P-gp. CAOV3 cells were then treated with IgG, exogenous active
HE4, exogenous active ANXA2, HE4 antibody, ANXA2 antibody or
exogenous active HE4 protein with ANXA2 antibody. CAOV3-A2-L cells
were additionally treated with exogenous active HE4 protein for 0,
12 and 24 h. Treatment with exogenous active HE4 or ANXA2 led to a
significant decrease in hsa-miR-129-5p expression in CAOV3 cells
within 24 h, while treatment with HE4 antibody, ANXA2 antibody and
ANXA2 antibody with HE4 active protein induced a significant
increase in hsa-miR-129-5p. In CAOV3-A2-L cells, high expression of
hsa-miR-129-5p was not reversed within 24 h after treatment with
exogenous active HE4 protein. There was no significant difference
in hsa-miR-129-5p between the untreated group and treated group by
IgG in CAOV3 cells (P>0.05; Fig.
7B). Data from these tests further verified that the expression
change in hsa-miR-129-5p was associated with HE4, which may require
the involvement of ANXA2.
GSEA of HE4, ANXA2 and P-gp
In addition, since HE4 may participate in the
development of multidrug resistance of OC at different levels, the
GSEA network database was used to search for the common signaling
pathways shared by HE4, ANXA2 and P-gp. GSEA results showed
enrichment of both HE4 and ANXA2 genes in the signaling pathways
regulating the actin cytoskeleton and focal adhesion, and ANXA2 and
P-gp in the regulation of actin cytoskeleton, Toll-like receptor,
chemokine and JAK-STAT signaling pathways. Notably, all three genes
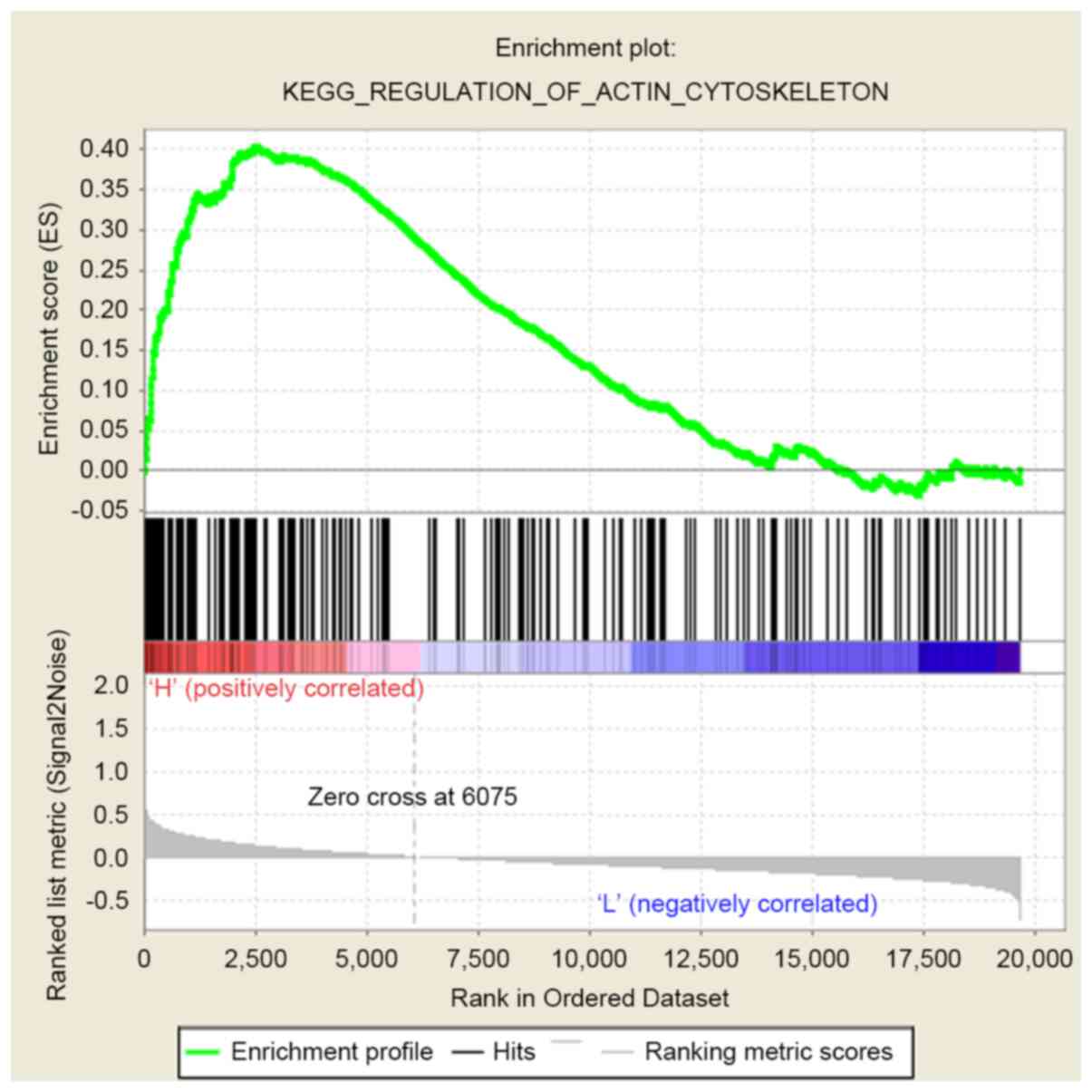
were commonly enriched in the signaling pathway involved in the
regulation of actin cytoskeleton (Fig.
8).
Discussion
Drug resistance of OC cells is a complex process
involving multiple factors, levels and genes. Recent studies have
indicated a close association of HE4 with drug resistance of OC
cells (8,9). Ribeiro et al (10) reported that increased HE4 expression
in OC cells simultaneously induces resistance to both paclitaxel
and carboplatin in vitro. Moreover, the MAPK signaling
pathway and changes in the intracellular microtubule content and
stability may be involved in this process (10). Another study suggests that HE4
affects the tumor microenvironment by interacting with epidermal
growth factor receptor, insulin growth factor receptor and
hypoxia-inducible factor-1α, thereby enhancing proliferative
ability and inducing resistance to cisplatin in subcutaneous
xenografts of nude mice (25). The
collective results suggest that HE4 participates in the progress of
multidrug resistance of OC at multiple levels.
It was shown that HE4 and ANXA2 were interacting
proteins in OC cells. ANXA2 was required for HE4-mediated promotion
of invasion and metastasis. The interactions between HE4 and ANXA2
facilitated the invasion and metastasis of OC cells, which was
proposed to be attributed to ANXA2-assisted transfer of HE4 into
the cell nucleus and consequent induction of
invasion/metastasis-related genes (14). Several clinical and basic studies
have confirmed that drug resistance and invasion/metastasis of
malignant tumors are not two isolated processes but closely
associated biological behaviors. In human drug-resistant human
breast cancer cells, P-gp was proposed to promote migration and
invasion through its associations with ANXA2 (15). Another study on gastric cancer cells
reported that ANXA2 stimulates the expression of P-gp and the
development of drug resistance by activating MAPK signaling pathway
(19). These findings support the
theory that HE4, ANXA2 and P-gp act in concert to play significant
roles in the malignant biological behavior of cancer cells.
In the present study, both HE4 and ANXA2 were highly
expressed in all three OC cell lines examined (CAOV3, OVCAR3 and
SKOV3), while the expression of P-gp was slightly higher in CAOV3
compared with OVCAR3 and SKOV3 cells, which suggested HE4, ANXA2
and P-gp are widely expressed in OC cell lines.
Co-immunoprecipitation experiments demonstrated co-expression
between HE4 and ANXA2, as well as ANXA2 and P-gp, in CAOV3 cells.
Colocalization of HE4/P-gp and ANXA2/P-gp was determined via
immunofluorescent staining, indicating interactions among the three
proteins. These results confirmed an association among HE4, ANXA2
and P-gp. In further experiments, it was found that drug resistance
to adriamycin and the expression and activity of P-gp was
significantly decreased after transfection in CAOV3 cells
(CAOV3-HE4-L with decreased HE4 expression or CAOV3-A2-L with
decreased ANXA2 expression). Experiments with paclitaxel were not
performed in the present study, since other members of the team
have used CAOV3 cells to perform experiments with paclitaxel (data
unpublished). In addition, DC (liposomal adriamycin + carboplatin)
has been recommended for the first-line chemotherapy regimen for
ovarian cancer in 2019 NCCN guidelines (26). Treatment with exogenous active HE4
or ANXA2 protein induced a significant increase in P-gp in CAOV3
cells within 24 h, while treatment with HE4 or ANXA2 antibody
significantly decreased the expression of P-gp. Co-treatment with
ANXA2 antibody and active HE4 protein induced a decrease in P-gp
expression in CAOV3 cells. However, this expression pattern was not
significantly reversed within 24 h after the treatment of
CAOV3-A2-L cells with exogenous HE4 active protein. Therefore the
present results have demonstrated that both HE4 and ANXA2 could
regulate P-gp expression levels, thus resulting in P-gp-mediated
drug resistance in OC cells. Furthermore, this effect may be
associated with the interactions between HE4 and ANXA2, and so, it
is speculated that HE4 promotes P-gp-mediated drug resistance in OC
cells through a mechanism requiring the participation of ANXA2.
Besides experiments in vitro, the
relationship between HE4 and P-gp were further studied in human
samples and confirmed that HE4 and P-gp are highly expressed and
was positively associated in the tissues of drug-resistant patients
with OC. Both HE4 and P-gp expression levels were associated with
FIGO staging and chemoresistance. High expression of HE4 or P-gp
was an independent risk factor for poor prognosis of OC,
highlighting the significance of histopathological examination of
HE4 and P-gp expression in predicting drug resistance and prognosis
for patients with OC.
To further elucidate the detailed mechanisms by
which the interactions between HE4 and ANXA2 regulate P-gp
expression and lead to drug resistance in ovarian cancer cells,
five miRNAs that potentially bind the P-gp gene, ABCB1, were
predicted using bioinformatics and their associations with
HE4/ANXA2 were explored. miRNAs are single-stranded small RNA
molecules ~21-23 bases in length. In plants and animals, miRNAs
directly formed RNA-induced silencing complexes to degrade mRNA or
repress translation through base-pairing with the 3′-UTR region of
target mRNAs, thereby participating in post-transcriptional
regulation (27). Among the five
miRNAs, expression of hsa-miR-129-5p was particularly increased
after transfection in CAOV3 cells, indicating that the expression
of hsa-miR-129-5p is consistent with those of HE4, ANXA2 and P-gp.
The addition of exogenous active HE4 or ANXA2 protein significantly
suppressed the expression of hsa-miR-129-5p in CAOV3 cells within
24 h while treatment with HE4 antibody, ANXA2 antibody or ANXA2
antibody with HE4 active protein led to a remarkable increase in
hsa-miR-129-5p expression. Treatment of CAOV3-A2-L cells with
exogenous active HE4 protein did not lead to a significant reversal
of the hsa-miR-129-5p expression pattern within 24 h, indicating
that the expression of hsa-miR-129-5p was associated with HE4,
which may require the involvement of ANXA2. The increased
expression of miR-129-5p in human drug-resistant gastric cancer
cells was reported to decrease drug resistance, and data from the
luciferase reporter system further demonstrated that three
multidrug resistance genes, including ABCB1, are target genes of
miR-129-5p (28). Consistently, it
was proposed that HE4, through its interactions with ANXA2,
regulates hsa-miR-129-5p, then regulates P-gp, thereby enhancing
drug resistance of OC cells. In addition to the mechanism involving
miRNA, utilization of the TCGA database for GSEA of HE4, ANXA2 and
P-gp revealed that all three genes are involved in the regulation
of the actin cytoskeleton signaling, which is closely associated
with the development of drug resistance in tumor cells (29). Furthermore, knockout or knockdown or
point mutation of P-gp will be conducted in order to study the
biological behaviors of CAOV3 in future studies.
In summary, HE4 and P-gp are associated with OC
development, both displaying high expression in the tissues of
drug-resistant patients with OC. The present study has shown for
the first time that ANXA2 interacts and colocalizes with P-gp in OC
cells, suggesting that HE4 promotes P-gp-mediated OC resistance
through interactions with ANXA2, which led to poor prognosis.
Furthermore, this mechanism may be associated with decreased
expression of hsa-miR-129-5p and dysregulation of the actin
cytoskeleton signaling pathway. The present study results highlight
novel targets for the clinical treatment of drug-resistant OC and
provide a theoretical basis for developing effective new
chemotherapy regimens. However, the specific molecular pathway by
which HE4 regulates the expression and activity of P-gp requires
further study.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural
Science Foundation of China (grant nos. 81672590, 81472437,
30872757 and 81602304) and the Outstanding Scientific Fund of
Shengjing Hospital (grant no. 201303).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
QL and BL confirm the authenticity of all the raw
data. QL performed the experiments, acquired the data and drafted
the manuscript. DWL, HMW and SJ collected human OC tissues, and
performed the immunohistochemistry and western blotting
experiments. MJZ analyzed and interpreted the GSEA data. JJL, YYH,
LCZ and LD were involved in the in vitro cell experiments and
drafting the manuscript. BL participated in the conception and
experimental design of the project. All authors read and approved
the final manuscript, and agreed to be accountable for all aspects
of the research in ensuring that the accuracy or integrity of any
part of the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
Samples were fully encoded to protect patient
confidentially. The study and its protocols were approved by the
Research Ethics committees of Shengjing Hospital Affiliated with
China Medical University (IACUC permit no. 2020PS277K). Since all
the samples used in the study were discarded, the informed consents
were not needed, which was approved by the ethics committees.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kipps E, Tan DS and Kaye SB: Meeting the
challenge of ascites in ovarian cancer: New avenues for therapy and
research. Nat Rev Cancer. 13:273–282. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Baguley BC: Multiple drug resistance
mechanisms in cancer. Mol Biotechnol. 46:308–316. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Baguley BC: Multidrug resistance in
cancer. Methods Mol Biol. 596:1–14. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mechetner EB and Roninson IB: Efficient
inhibition of P-glycoprotein-mediated multidrug resistance with a
monoclonal antibody. Proc Natl Acad Sci USA. 89:5824–5828. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Breier A, Gibalova L, Seres M, Barancik M
and Sulova Z: New insight into p-glycoprotein as a drug target.
Anticancer Agents Med Chem. 13:159–170. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gibalová L, Sereš M, Rusnák A, Ditte P,
Labudová M, Uhrík B, Pastorek J, Sedlák J, Breier A and Sulová Z:
P-glycoprotein depresses cisplatin sensitivity in L1210 cells by
inhibiting cisplatin-induced caspase-3 activation. Toxicol In
Vitro. 26:435–444. 2012. View Article : Google Scholar
|
|
7
|
Hellström I, Raycraft J, Hayden-Ledbetter
M, Ledbetter JA, Schummer M, McIntosh M, Drescher C, Urban N and
Hellström KE: The HE4 (WFDC2) protein is a biomarker for ovarian
carcinoma. Cancer Res. 63:3695–3700. 2003.
|
|
8
|
Aarenstrup Karlsen M, Høgdall C,
Nedergaard L, Philipsen Prahm K, Schou Karlsen NM, Weng Ekmann-Gade
A, Henrichsen Schnack T, Svenstrup Poulsen T, Jarle Christensen I
and Høgdall E: HE4 as a predictor of adjuvant chemotherapy
resistance and survival in patients with epithelial ovarian cancer.
APMIS. 124:1038–1045. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nassir M, Guan J, Luketina H, Siepmann T,
Rohr I, Richter R, Castillo-Tong DC, Zeillinger R, Vergote I, Van
Nieuwenhuysen E, et al: The role of HE4 for prediction of
recurrence in epithelial ovarian cancer patients-results from the
OVCAD study. Tumour Biol. 37:3009–3016. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ribeiro JR, Schorl C, Yano N, Romano N,
Kim KK, Singh RK and Moore RG: HE4 promotes collateral resistance
to cisplatin and paclitaxel in ovarian cancer cells. J Ovarian Res.
9:282016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tanaka T, Akatsuka S, Ozeki M, Shirase T,
Hiai H and Toyokuni S: Redox regulation of Annexin II and its
implications for oxidative stress-induced renal carcinogenesis and
metastasis. Oncogene. 23:3980–3989. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bjørnland K, Winberg JO, Odegaard OT,
Hovig E, Loennechen T, Aasen AO, Fodstad O and Maelandsmo GM:
S100A4 involvement in metastasis: Deregulation of matrix
metalloproteinases and tissue inhibitors of matrix
metalloproteinases in osteosarcoma cells transfected with an
anti-S100A4 ribozyme. Cancer Res. 59:4702–4708. 1999.
|
|
13
|
Mathisen B, Lindstad RI, Hansen J,
El-Gewely SA, Maelandsmo GM, Hovig E, Fodstad O, Loennechen T and
Winberg JO: S100A4 regulates membrane induced activation of matrix
metalloproteinase-2 in osteosarcoma cells. Clin Exp Metastasis.
20:701–711. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhuang H, Tan M, Liu J, Hu Z, Liu D, Gao
J, Zhu L and Lin B: Human epididymis protein 4 in association with
Annexin II promotes invasion and metastasis of ovarian cancer
cells. Mol Cancer. 13:2432014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang F, Zhang H, Wang Z, Yu M, Tian R, Ji
W, Yang Y and Niu R: P-glycoprotein associates with Anxa2 and
promotes invasion in multidrug resistant breast cancer cells.
Biochem Pharmacol. 87:292–302. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang ZD, Li Y, Fan Q, Zhao B, Tan B and
Zhao XF: Annexin II is implicated in multi-drug-resistance in
gastric cancer through p38MAPK and AKT pathway. Neoplasma.
61:627–637. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li N, Yang L, Wang H, Yi T, Jia X, Chen C
and Xu P: MiR-130a and MiR-374a function as novel regulators of
cisplatin resistance in human ovarian cancer A2780 Cells. PLoS One.
10:e01288862015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jung H, Kim JS, Kim WK, Oh KJ, Kim JM, Lee
HJ, Han BS, Kim DS, Seo YS, Lee SC, et al: Intracellular Annexin II
regulates NF-κB signaling by binding to the p50 subunit:
Implications for gemcitabine resistance in pancreatic cancer. Cell
Death Dis. 6:e16062015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang R, Lu M, Zhang Z, Tian X, Wang S and
Lv D: Resveratrol reverses P-glycoprotein-mediated multidrug
resistance of U2OS/ADR cells by suppressing the activation of the
NF-κB and p38 MAPK signaling pathways. Oncol Lett. 12:4147–4154.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
FIGO Committee on Gynecologic Oncology, .
Current FIGO staging for cancer of the vagina, fallopian tube,
ovary, and gestational trophoblastic neoplasia. Int J Gynaecol
Obstet. 105:3–4. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Morgan RJ Jr, Copeland L, Gershenson D,
Locker G, McIntosh D, Ozols R and Teng N; The National
Comprehensive Cancer Network, : NCCN ovarian cancer practice
guidelines. Oncology (Williston Park). 10 (Suppl 11):293–310.
1996.PubMed/NCBI
|
|
22
|
Zhuang H, Gao J, Hu Z, Liu J, Liu D and
Lin B: Co-expression of Lewis y antigen with human epididymis
protein 4 in ovarian epithelial carcinoma. PLoS One. 8:e689942013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang H, Tan M, Zhang S, Li X, Gao J, Zhang
D, Hao Y, Gao S, Liu J and Lin B: Expression and significance of
CD44, CD47 and c-met in ovarian clear cell carcinoma. Int J Mol
Sci. 16:3391–3404. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Moore RG, Hill EK, Horan T, Yano N, Kim K,
MacLaughlan S, Lambert-Messerlian G, Tseng YD, Padbury JF, Miller
MC, et al: HE4 (WFDC2) gene overexpression promotes ovarian tumor
growth. Sci Rep. 4:35742014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Armstrong DK, Alvarez RD, Bakkum-Gamez JN,
Barroilhet L, Behbakht K, Berchuck A, Berek JS, Chen LM, Cristea M,
DeRosa M, et al: NCCN guidelines insights: Ovarian cancer, Version
1. J Natl Compr Canc Netw. 17:896–909. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kim VN and Nam JW: Genomics of microRNA.
Trends Genet. 22:165–173. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wu Q, Yang Z, Xia L, Nie Y, Wu K, Shi Y
and Fan D: Methylation of miR-129-5p CpG island modulates
multi-drug resistance in gastric cancer by targeting ABC
transporters. Oncotarget. 5:11552–11563. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Islam SU, Shehzad A, Sonn JK and Lee YS:
PRPF overexpression induces drug resistance through actin
cytoskeleton rearrangement and epithelial-mesenchymal transition.
Oncotarget. 8:56659–56671. 2017. View Article : Google Scholar : PubMed/NCBI
|