Introduction
Obesity, which is defined as a body mass index
>30 kg/m2, has become one of the major public health
problems worldwide and its prevalence has increased in recent
years. The World Health Organization reported that the total number
of obese individuals is four times the number 30 years ago
(1). Obesity and overweight are
known to adversely affect human health and the risks and prognosis
for a number of serious medical conditions, such as type 2 diabetes
and cardiovascular disease, liver diseases, cardiovascular diseases
and even cancer (2,3).
Obesity involves pathological changes in a number of
tissues, such as adipose tissue and liver. Expanded visceral fat
deposits are also linked to dyslipidemia, which is characterized by
high triacylglycerol (TG), low-density lipoprotein cholesterol
(LDL-C), high-density lipoprotein cholesterol (HDL-C) and insulin
resistance, which are often accompanied by impaired metabolic
regulation in adipose tissue, leading to an over-production of free
fatty acids (FFAs) (4). The flux of
FFAs toward the liver results in increased TG deposition and
secretion of TG-rich lipoproteins, which in turn affect lipolipase
lipase (LPL) activity and the distribution of lipoprotein subtypes
(5).
Currently, several drugs, such as sibutramine and
orlistat, have been approved by the Food and Drug Administration of
the United States, and have been applied for long-term use to
prevent obesity (6). Sibutramine
increases energy consumption and reduces fat accumulation by
inhibiting the re-uptake of monoamine signaling elements (7). Orlistat is a gastrointestinal lipase
inhibitor that inhibits intestinal triglyceride absorption
(8). However, these drugs
inevitably cause side effects such as uneven cardiac rhythm and
chest pain, which restrict their clinical application (9).
Polysaccharides are widely distributed in nature:
Plants, animals and fungi are valuable sources of polysaccharides.
Several types of polysaccharides have been determined to exert
multiple functions, such as anti-inflammatory, anti-hyperlipidemic
and anti-hypertension effects (10). Among these functions, the
lipid-lowering effect of polysaccharides has become a focus of
research in recent years. Polysaccharides from soybean (11), flaxseed (12) and Momordica charantia
(13) have been demonstrated to be
effective in treating or preventing obesity.
Polygonatum sibiricum F. Delaroche (P.
sibiricum) is a perennial herb of the Asparagaceae family. It
is mainly distributed in the Northern Hemisphere, including China,
Mongolia, North Korea and the USA (14). P. sibiricum is a well-known
traditional medicinal herb in China (15), as well as a health-improving
substance (16). P.
sibiricum has been widely used in a number of Chinese herbal
medicine compounds treating different diseases as recorded in
Ben Cao Gang Mu (Compendium of Materia Medica) and
its effects in treating diabetes (17), cardiovascular and cerebrovascular
diseasess18), somnipathy (19),
dyslipidemia and pathological fatigue (20) have also been determined by modern
medical methods.
A number of the functional components of P.
sibiricum have been documented, including polysaccharides,
phytosterols, triterpenoids, alkaloids, lignans, flavonoids and
phytosterols (21). Among them,
polysaccharides and phytosterols are the most abundant medicinal
components. P. sibiricum polysaccharide (PSP) is mainly
composed of galactose, mannose, glucose and galacturonic acid, in
molar ratios of 29.63:36.10:15.09:10.20 (22). PSP has been verified to exert a
variety of medicinal functions, including anti-inflammatory
(23) and anti-tumor effects
(24), lowering blood glucose
(25), enhancing immunity and
promoting the differentiation of bone cells (20).
To investigate if PSP ameliorates high-fat diet
(HFD)-induced obesity mice were fed an HFD to establish a high-fat
model and PSP was administered intragastrically. The molecular
mechanisms were investigated by reverse transcription-quantitative
PCR (RT-qPCR) and western blotting.
Materials and methods
Reagents
PSP (purity 90%) were obtained from Shaanxi
Yikanglong Biotechnology Co., Ltd. Triacylglycerol (TG), total
cholesterol (TC), high-density lipoprotein cholesterol (HDL-C) and
low-density lipoprotein cholesterol (LDL-C) kits were obtained from
Nanjing Jiancheng Bioengineering Institute. The glucometer was from
Sannuo Bio-Sensing Technology Co., Ltd., TRIzol® was
obtained from Invitrogen; Thermo Fisher Scientific, Inc. and
high-capacity cDNA reverse transcription kits were obtained from
Applied Biosystems (Thermo Fisher Scientific, Inc.). Antibodies
against fatty acid synthase (FAS; cat. no. 4233), phosphorylated
(p-) 5′ adenosine monophosphate-activated protein kinase (AMPK)α
(Thr172; cat. no. 50081), AMPKα (cat. no. 5832), p-acetyl-coenzyme
A carboxylase (ACC; Ser79; cat. no. 11818), TNF-α (cat. no. 11948),
IL-6 (cat. no. 12912), IL-1β (cat. no. 31202), inducible nitric
oxide synthase (iNOS; cat. no. 13120), β-actin (cat. no. 4970),
carnitine palmitoyl transferase I (CPT1; cat. no. 12252) were
purchased from Cell Signaling Technology, Inc. The peroxisome
proliferator-activated receptor (PPARα; cat. no. ab245119) antibody
was purchased from Abcam. The sterol regulatory-element binding
protein (SREBP-1; cat. no. PA1-337) antibody and the HRP-conjugated
goat anti-rabbit IgG (cat. no. 31460) antibody were obtained from
Thermo Fisher Scientific, Inc. RIPA Lysis and Extraction Buffer was
obtained from Beyotime Institute of Biotechnology and protease and
phosphatase inhibitors were obtained from Roche Diagnostics.
Polyvinylidene fluoride (PVDF) membranes were obtained from Cytiva.
An enhanced chemiluminescence (ECL) detection kit was purchased
from Pierce (Thermo Fisher Scientific, Inc.).
Experimental animals
A total of 50 eight-week-old C57BL male mice were
purchased from Hunan SJA Laboratory Animal Co., Ltd. The mice were
housed in a constant temperature and humidity animal room at 23±2°C
and a relative humidity of 40–45%, with a 12-h light/dark cycle,
and were acclimated for 7 days. Prior to the animal experiments,
the mice were fed a normal diet (Hunan SJA Laboratory Animal Co.,
Ltd.) and provided water ad libitum. The 50 mice were
randomly divided into 5 groups: A normal diet (control) group; a
high fat diet (HFD) group and the three PSP groups (mice separately
received 200, 400 and 800 mg/kg of PSP daily by intragastric
administration for 3 days before HFD feeding and for the whole
experiment). The animal body weights and food intake were recorded
weekly. The care and use of the animals and experimental protocols
complied with the Guidelines for the Care and Use of Experimental
Animals of Hunan University of Chinese Medicine. The project was
approved by the Office of Animal Experiment Ethics of Hunan
University of Chinese Medicine (approval no. 20190165). At the end
of the experiments, the mice were fasted for 12 h and sacrificed
with CO2 for 5 min as recommended by Pritchett et
al (26). The CO2
flow rate was 40% chamber volume/min. Blood samples were gathered
from the eye for glucose test and biochemical analysis. The liver
and epididymal fat pad were immediately removed, weighed, imaged,
flash frozen in liquid nitrogen and stored at −80°C for further
experiments.
According to the results of body weight changes, the
body weights of the 400 and 800 mg/kg PSP groups were significantly
lower compared with the HFD group. However, no significant
difference was observed between these two groups. Therefore, the
400 mg/kg PSP group was chosen for further research.
Biochemical analysis
Blood samples were placed in refrigerator at 4°C
overnight and then centrifuged at 1,000 × g for 15 min at 4°C. The
supernatants were collected as plasma samples. TG, TC, LDL-c and
HDL-c levels were measured by commercial assay kits from Nanjing
Jiancheng Bioengineering Institute following the manufacturer's
instructions. Serum adiponectin (cat. no. MRP300), leptin (cat. no.
MOB00B) and resistin (cat. no. MRSN00) levels were measured by
ELISAs (R&D Systems, Inc.), and insulin levels were measured
using a mouse insulin-1 ELISA kit (Sigma-Aldrich; Merck KGaA; cat.
no. RAB0327).
Histopathological examinations
Samples (epididymal fat pads and liver tissues) were
fixed with 10% neutrally buffered formalin for 2 days at room
temperature, dehydrated in an ascending series of alcohol, and then
embedded in paraffin by the paraffin embedding machine for 16 h.
Paraffin-embedded samples were then cut into 5 µm sections. The
sections were then deparaffinized with xylene, rehydrated in a
descending series of alcohol, rinsed with distilled water for 1 min
and stained with hematoxylin (20 min) and eosin (1 min) (H&E)
at room temperature. Images were captured under a light microscope
(magnification, ×200) in five randomly selected fields of view.
RNA isolation and RT-qPCR
Total RNA was isolated from the liver tissues by
using RNA extract reagent (TRIzol®). A total of 2 µg of
RNA was reverse transcribed into cDNA with cDNA reverse
transcription kits (Thermo Fisher Scientific, Inc.) at 42°C for 30
min and then 80°C for 10 min for enzymatic inactivation. qPCR was
performed on a CFX96 Real Time PCR system (Applied Biosystems;
Thermo Fisher Scientific, Inc.) using the TransStart Green qPCR
SuperMix UDG kit (Beijing Transgen Biotech Co., Ltd.; cat. no.
AQ111-01). The following thermocycling conditions were used for the
qPCR: Initial denaturation at 95°C for 1 min; followed by 40 cycles
of amplification (94°C for 30 sec, 58°C for 25 sec and 72°C for 1
min), followed by an extension at 72°C for 3 min. The primer
sequences were described in our previous publication and are shown
in Table I (27). The 2−∆∆Cq method was used
for quantification of the expression levels (28). All these methods were according to
the manufacturer's protocols.
 | Table I.Primers sequences used in reverse
transcription-quantitative PCR. |
Table I.
Primers sequences used in reverse
transcription-quantitative PCR.
| Gene | Forward | Reverse |
|---|
| β-actin |
5′-ATCATGTTTGAGACCTTCACACC-3′ |
5′-TAGAGCAACATAGCACAGCTTCTCTT-3′ |
| Peroxisome
proliferator-activated receptor α |
5′-TAAAGTACGGTGTGTATGAAGCCAT-3′ |
5′-ATGTAGCCTATGTTTAGAAGGCCAG-3′ |
| Sterol
regulatory-element binding protein-1C |
5′-GCCATCGACTACATCCGCTTCTT-3′ |
5′-TGGGCTTTGACCTGGCTATCCTC-3′ |
| Fatty acid
synthase |
5′-AGCACTGCCTTCGGTTCAGTC-3′ |
5′-AAGAGCTGTGGAGGCCACTTG-3′ |
| Carnitine
palmitoyltransferase I |
5′-CAGCTCGCACATTACAAGGA-3′ |
5′-TGCACAAAGTTGCAGGACTC-3′ |
| IL-1β |
5′-GCAACTGTTCCTGAACTCAACT-3′ |
5′-ATCTTTTGGGGTCCGTCAACT-3′ |
| TNF-α |
5′-CCCTCACACTCAGATCATCTTCT-3′ |
5′-GCTACGACGTGGGCTACAG-3′ |
| IL-6 |
5′-TAGTCCTTCCTACCCCAATTTCC-3′ |
5′-TTGGTCCTTAGCCACTCCTTC-3′ |
| Inducible nitric
oxide synthase |
5′-CCTTACGAGGCGAAGAAGGACAG-3′ |
5′-CAGTTTGAGAGAGGAGGCTCCG-3′ |
Western blot analysis
The liver tissues were homogenized in RIPA Lysis and
Extraction Buffer (Beyotime Institute of Biotechnology) on ice for
30 min and then the suspension was centrifuged at 12,000 × g for 5
min at 4°C. The protein concentration was determined using a BCA
Protein assay kit (Abcam). The protein-containing supernatants were
equally (15 µg) subjected to 10% SDS-PAGE and then transferred onto
a PVDF membrane. The membrane was first blocked with 0.1% TBS-Tween
20 (TBST) containing 5% skimmed milk at room temperature for 1 h,
followed by incubations with the following primary antibodies:
Anti-PPARα (1:2,000), anti-SREBP-1C (1:2,000), anti-FAS (1:2,000),
anti-CPT-1 (1:2,000), anti-IL-1β (1:2,000), anti-TNF-α (1:2,000),
anti-IL-6 (1:2,000), anti-iNOS (1:2,000) and β-actin (1:5,000)
overnight at 4°C. Subsequently the membranes were washed in TBST
(3×15 min) and incubated with secondary antibodies (1:10,000) at
room temperature for 1 h. Finally, the membrane was developed with
an ECL Plus™ western blotting detection system. The images were
captured using an imaging system (ChemiDoc™ XRS+, Bio-Rad
Laboratories, Inc.). The densitometric analysis was performed using
Adobe Photoshop 7.01 (Adobe Systems, Inc.).
Statistical analysis
SPSS v13.0 statistical software (SPSS, Inc.) was
used to analyze the data which were expressed as the mean ±
standard deviation. A one-way ANOVA was used for statistical
analyses among three groups with the Tukey HSD post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
PSP prevents HFD-induced obesity
To evaluate whether PSP prevents HFD-induced
obesity, the mice were administered PSPs as described in the
methods section. The body weights of mice in the control group
increased steadily up to the end of the experiment. The body
weights of the HFD group mice increased quickly from 25.33±0.44 g
at the beginning to 34.46±0.89 g on week 7 and then plateaued until
the end of the experiment. The body weights of the different PSP
dosages increased more slowly than those of the HFD group, as the
200 mg/kg PSP group increased from 25.42±0.46 to 34.97±0.88 g,
which was lower compared with the HFD group (P<0.05), while the
400 mg/kg PSP group was from 25.70±0.46 to 33.19±0.73 g and the 800
mg/kg PSP group was from 25.55±0.48 to 32.81±0.71 g, all lower
compared with the HFD group (P<0.01), However, no significant
difference was observed between these two groups. Thus, the 400
mg/kg PSP group was chosen for further experiments and this group
was defined as the PSP group hereafter (Fig. 1A).
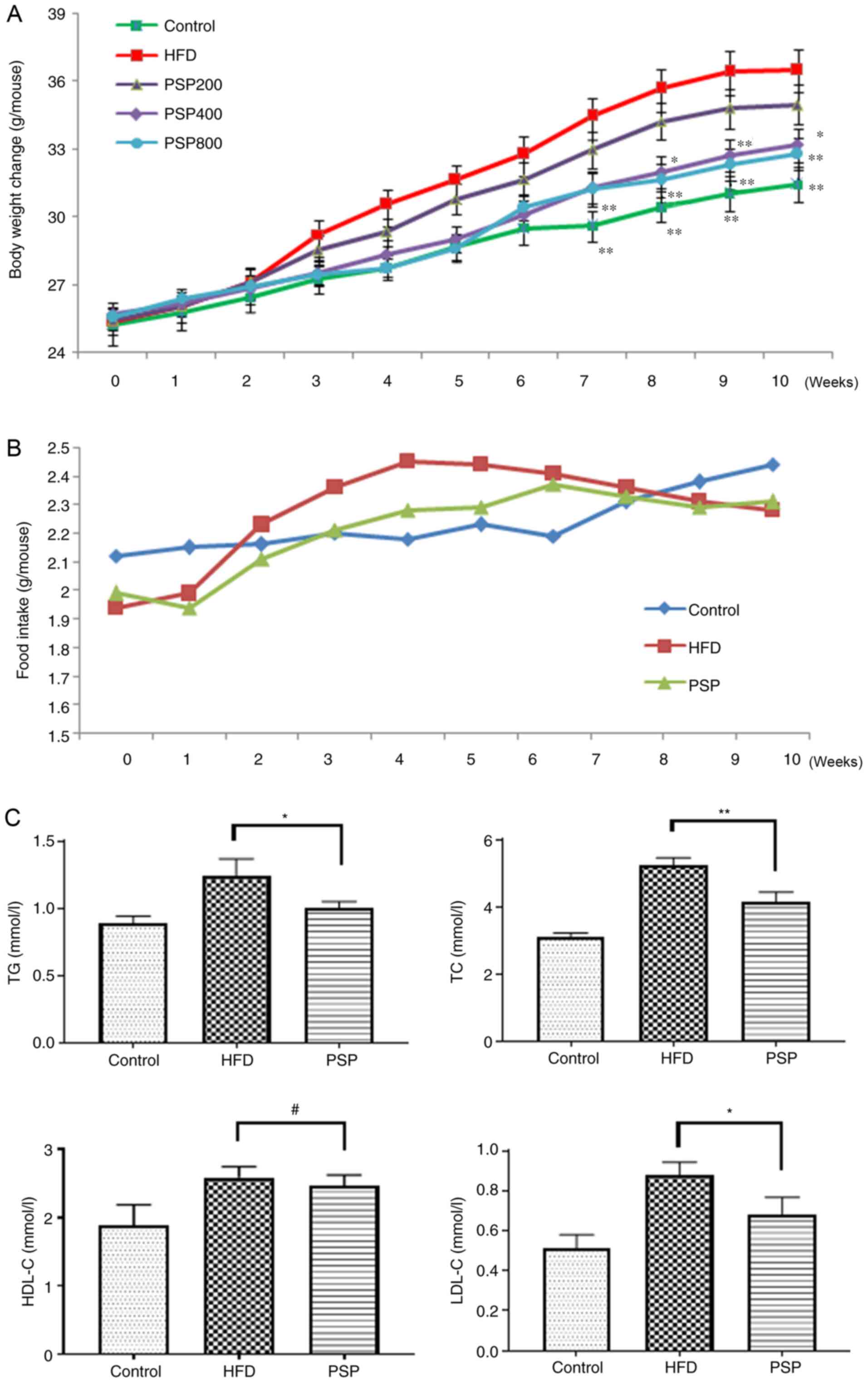 | Figure 1.Inhibitory effects of PSP on
HFD-induced obesity in mice. C57BL mice were divided into 5 groups.
Control group mice received a normal diet, HFD group mice received
HFD and three PSP groups mice received HFD and 200, 400 and 800
mg/kg of PSP by intragastric administration. Body weight and food
intake were recorded weekly, Serum lipid index were measured after
the mice were sacrificed. (A) Body weight changes and (B) food
intake. (C) Serum TG, TC, HDL-C and LDL-C. #P>0.05,
*P<0.05, **P<0.01. PSP, Polygonatum sibiricum F.
Delaroche polysaccharide; HFD, high-fat diet; TG,
triacylglycerol; TC, total cholesterol; HDL-C, high-density
lipoprotein cholesterol; LDL-C, low-density lipoprotein
cholesterol. |
The average food intake of control group increased
steadily and slowly, while the HFD group grew quickly in the first
4 weeks and plateaued with a slight decrease toward the end of the
experiment at a level that was higher compared with the control
group until week 8. The PSP group mice received less food than the
HFD group during weeks 2 and 6 (Fig.
1B). Serum lipid profiles were analyzed using a detection kit.
Compared with the levels in the control group, TC, TG, HDL-C and
LDL-C were increased following HFD feeding. PSP significantly
decreased the levels of TG, TC and LDL-C, while HDL-C was not
significantly affected by PSP (Fig.
1C). The serum glucose of the HFD group was 9.62±0.98 mmol/l,
much higher compared with the control group (5.63±0.53 mmol/l)
while the PSP group was 7.23±0.55 mmol/l. The insulin concentration
was also increased from 0.48±0.06 to 0.97±0.07 ng/ml by HFD, while
the PSP group was only 0.68±0.09 ng/ml, significantly lower than
the HFD group (P<0.01; Table
II).
| Table II.Effects of PSP on serum
parameters. |
Table II.
Effects of PSP on serum
parameters.
| Parameters | Control | HFD | PSP | P-value; Control
vs. HFD | P-value; HFD vs.
PSP |
|---|
| Glucose
(mmol/l) | 5.63±0.53 | 9.62±0.98 | 7.23±0.55 | <0.01 | <0.01 |
| Insulin
(ng/ml) | 0.48±0.06 | 0.97±0.07 | 0.68±0.09 | <0.01 | <0.01 |
| Resistin
(ng/ml) | 0.71±0.08 | 2.03±0.13 | 1.47±0.16 | <0.01 | <0.01 |
| Leptin (ng/ml) | 4.07±0.58 | 17.05±2.03 | 10.34±0.64 | <0.01 | <0.01 |
| Adiponectin
(µg/ml) | 6.39±0.72 | 4.50±0.39 | 5.83±0.80 | <0.01 | <0.05 |
PSP prevents HFD-induced hepatic
lesions and adipose hypertrophy
To evaluate the hepatoprotective effects of PSP,
mouse livers and abdominal adipose tissue were collected
immediately when the mice were sacrificed. The appearance of the
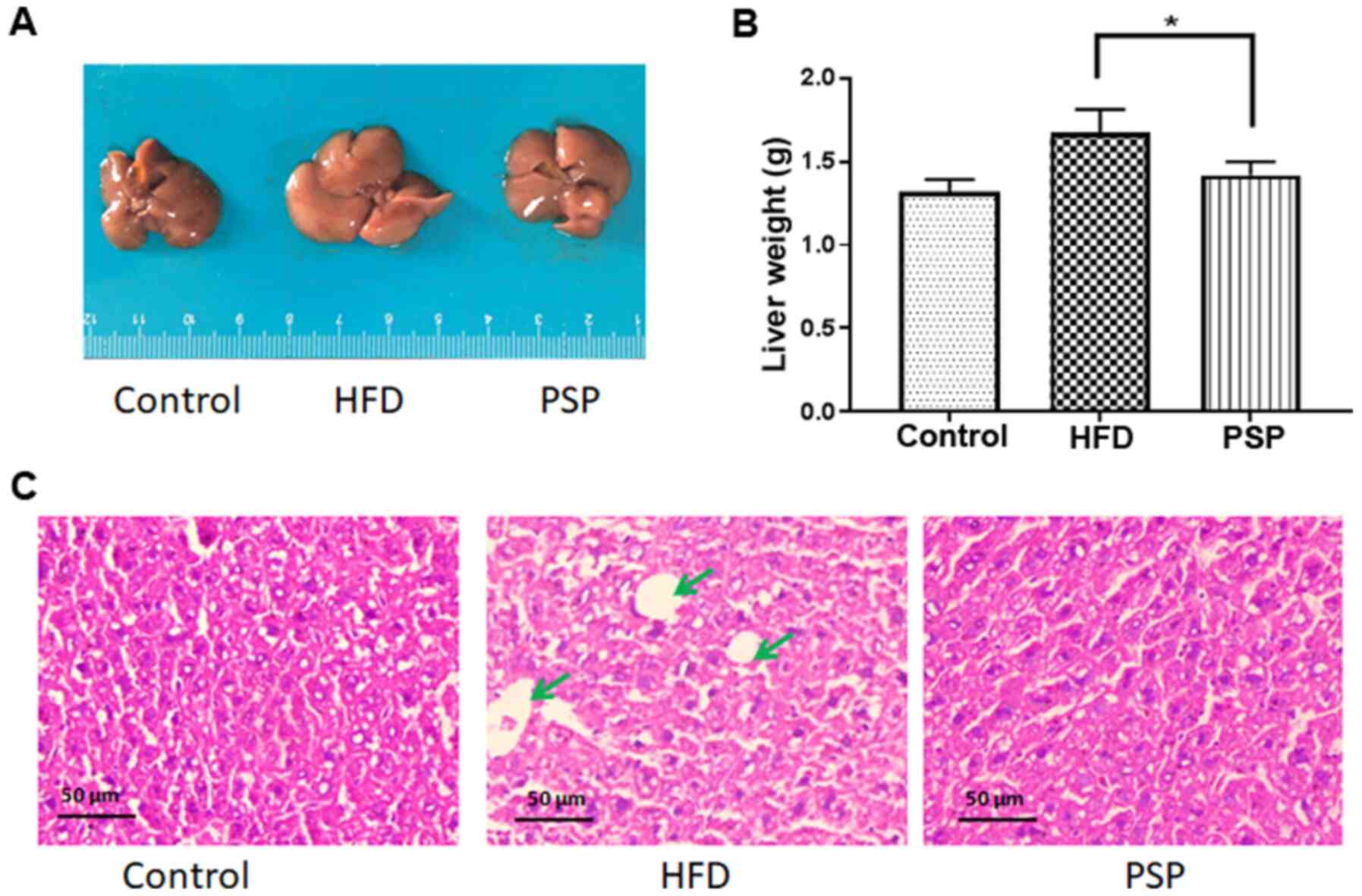
livers differed between the groups. As shown in Fig. 2A, the control group livers were in
dark red and were the smallest in size. The HFD group livers were
large in size and light in color; in addition, fat particles were
visible to the naked eye on those livers. The PSP group livers were
significantly smaller and the color was darker compared with those
of the HFD group. The differences in liver weights were as
expected; the liver weight of the control group was 1.32±0.12 g,
while that of the HFD group was 1.67±0.13 g and that of the PSP
group was 1.41±0.14 g, which was significantly lower compared with
the HFD group (Fig. 2B). The
H&E results revealed why the livers of the HFD group were large
and light in color. There was a large amount of fat in the
intercellular space of the livers. The PSP group livers contained
much less fat (Fig. 2C).
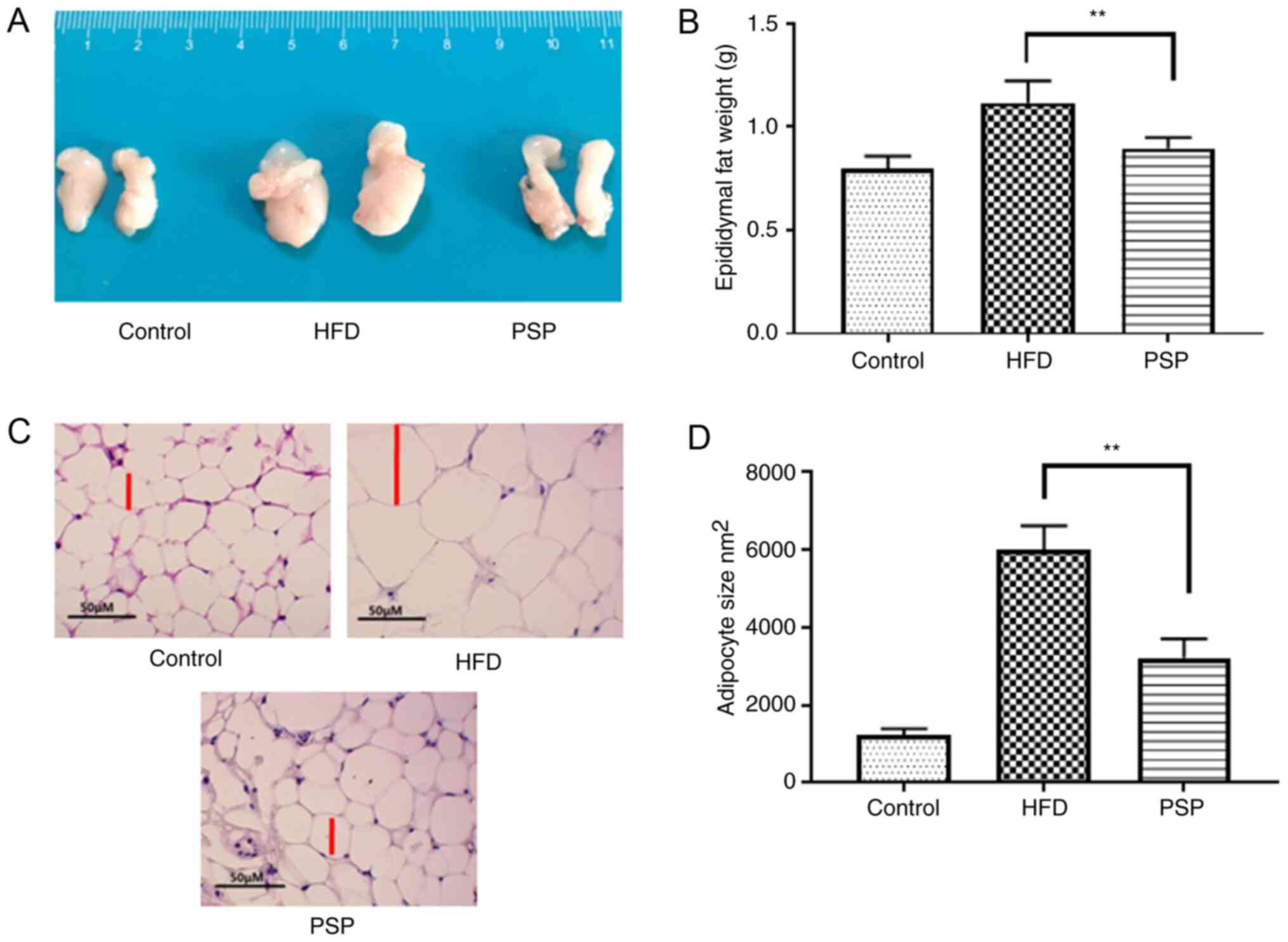
The trend in abdominal adipose tissue was similar to
that among the livers; mice in the control group had the smallest
adipose tissues weighing 0.80±0.06 g, while the HFD group was
1.12±0.13 g and PSP significantly decreased the weight of the
adipose tissues to 0.93±0.11 g (Fig. 3A
and B). H&E results indicated that the adipose cells of the
HFD group were larger compared with those of the control group. The
size of the PSP group adipose cells was significantly decreased
(Fig. 3C and D). Adipose secreted
factors are an important index of adipose health. The adiponectin
in control group was 6.39±0.72 µg/ml, while the HFD decreased it to
4.50±0.39 µg/ml and PSP effectively reversed this trend to
5.83±0.80 µg/ml (P<0.05). In contrast, leptin was increased from
4.07±0.58 to 17.05±2.03 ng/ml by HFD. Compared with the HFD group,
the PSP group exhibited a lower leptin content, only 10.34±0.64
ng/ml (P<0.01). Resistin in the HFD group increased from
0.71±0.08 to 2.03±0.13 ng/ml and PSP effectively decreased this to
1.47±0.16 ng/ml (Table II).
The molecular mechanisms by which PSP
prevents HFD-induced obesity
The liver is one of the key organs involved in lipid
regulation (12). In this present
study, mouse livers were collected to explore the potential
signaling pathways of PSP in HFD-induced hepatic steatosis. Lipid
metabolic and inflammatory mRNA levels were evaluated by RT-qPCR
and western blotting. The fatty acid and cholesterol
synthesis-associated genes FAS and SREBP-1 and the inflammatory
cytokines TNF-α, IL-6, IL-1β and iNOS were highly expressed, while
PPARα and CPT-1 were inhibited in the HFD group. PSP reversed this
trend, indicating that PSP inhibits lipid synthesis and
inflammation (Figs. 4 and 5A).
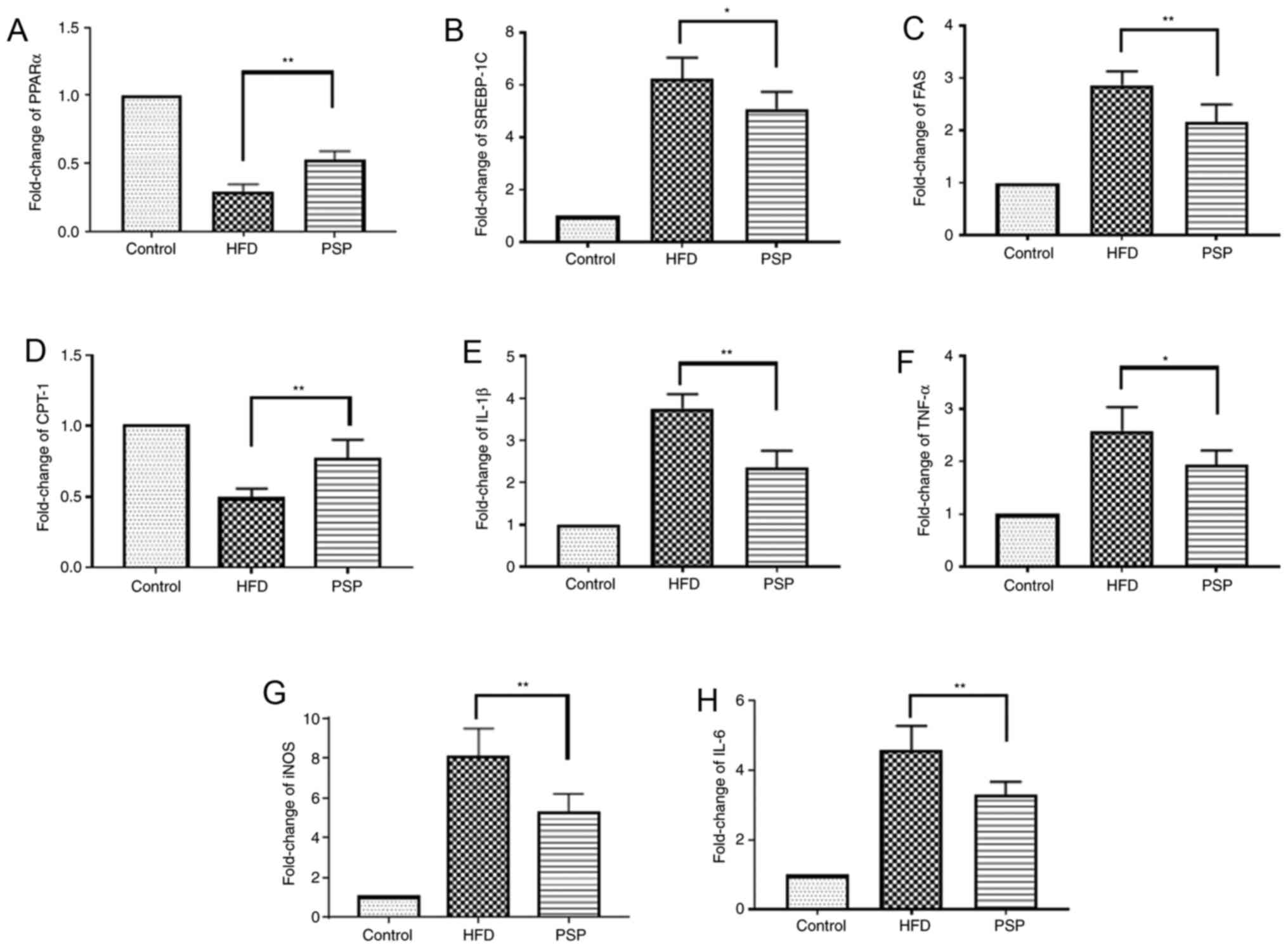 | Figure 4.Lipid-associated and inflammatory
cytokine mRNA changes. Results of reverse
transcription-quantitative PCR. Fold change of lipid-associated (A)
PPARα, (B) SREBP-1, (C) FAS and(D) CPT-1 and inflammatory (E)
IL-1β, (F) TNF-α, (G) iNOS and (H) IL-6 mRNA. *P<0.05,
**P<0.01.PSP, Polygonatum sibiricum F. Delaroche
polysaccharide; HFD, high-fat diet; PPAR, peroxisome
proliferator-activated receptor; SREBP, sterol regulatory-element
binding protein; FAS, fatty acid synthase; CPT-1, carnitine
palmitoyltransferase I; iNOS, inducible nitric oxide synthase. |
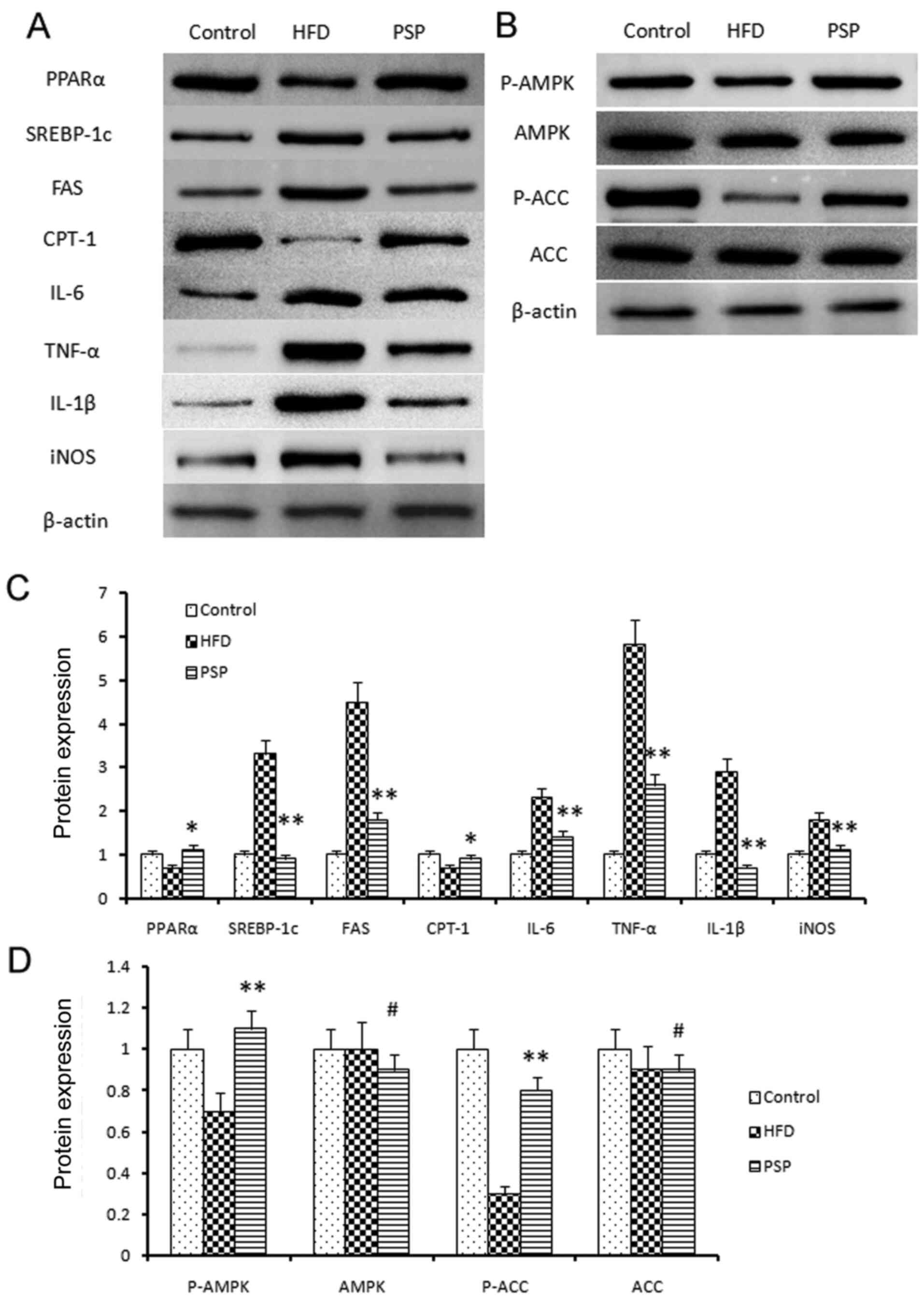 | Figure 5.Signal pathway changes and
lipid-associated genes and inflammatory cytokine protein changes.
(A) Western blotting was performed to reveal the protein
expressions of the lipid and inflammatory genes as well as the key
genes of the AMPK signal pathway. Lipid synthesis-associated genes
and inflammatory genes were inhibited by PSP. (B) P-AMPK and the
downstream gene p-ACC was clearly activated by PSP, which
demonstrated that the PSP could ameliorate HFD-induced mouse
obesity through activating the AMPK signal pathway. (C)
Semi-quantification of protein expression levels from part (A) (D)
Semi-quantification of protein expression levels from part (B)
#P>0.05, *P<0.05, **P<0.01. AMPK, 5′ adenosine
monophosphate-activated protein kinase; p-, phosphorylated; ACC,
acetyl-coenzyme A carboxylase; PSP, Polygonatum sibiricum F.
Delaroche polysaccharide; HFD, high-fat diet; PPAR, peroxisome
proliferator-activated receptor; SREBP, sterol regulatory-element
binding protein; FAS, fatty acid synthase; CPT-1, carnitine
palmitoyltransferase I; iNOS, inducible nitric oxide synthase. |
The AMPK signaling pathway was assessed and western
blotting results indicated that p-AMPK was suppressed in the HFD
group, while PSP significantly activated p-AMPK. P-ACC results were
similar to p-AMPK; clearly blocked in the HFD group and activated
in the PSP group (Fig. 5B-D).
Discussion
Obesity is the commonest chronic metabolic disease
in the modern world. Indeed, >13% of adults are overweight
(29). Obesity is the main cause of
a number of serious diseases, including hepatic diseases, diabetes
and even cancer (30). The
pathogenesis of obesity is very complex. It is widely accepted that
the development of obesity is mainly determined by genetic and
environmental factors (27). One of
the most important environmental factors is the modern lifestyle,
which involves the excessive intake of carbohydrates and fat, with
a low intake of fiber (31), as
well as a lack of physical activity. Although this reason is widely
accepted, unhealthy lifestyles remain hard to change. Compared with
the effects of lipid-lowering medicines, healthy foods provide
long-term protection against obesity and fewer side effects
(32). Consequently, research on
preventing the pathogenesis of obesity by eating healthy foods has
become increasingly popular.
Non-starch polysaccharides are considered effective
healthy foods for preventing obesity and one of the main reasons is
their unique chemical structures (33). They consist of or contain a number
of monosaccharides, such as fructose, galactose, mannose, ribose,
rhamnose, xylose and arabinose (34). In addition, the bonds differ from
those of starch; for example, oat β-glucan is β-1,3/1,4-D-glucan
and mushroom and yeast glucans are β-1,3/1,6-D-glucan (27). These characteristics indicate that
polysaccharides are difficult for mammals, which lack the relevant
enzymes, to digest and are broken down into monosaccharides, which
can be absorbed and used as an energy source (34).
P. sibiricum is a traditional Chinese herbal
medicine that is effective in treating metabolic diseases such as
atherosclerosis and hypertension (31). PSP is one of the main functional
components of P. sibiricum and has been determined to be
effective in enhancing immunity (35) and treating diabetes and diabetic
retinal injury (17), acute heart
failure (36) and cancer (24). An ethanol extract of P.
sibiricum is effective in high-fat diet-induced obesity
(37). Additionally, PSP regulates
bone marrow-derived macrophages to osteoclast differentiation
through the ERK/GSK-3β/β-Catenin signaling pathway (38). However, to the best of the authors'
knowledge, there has been no study on the anti-obesity effect of
PSP. Thus, the present study evaluated the anti-obesity effect of
PSP and the latent molecular mechanisms. PSP groups mice had slower
bodyweight increases and the livers and abdominal fat pads were
smaller compared with those of the HFD group. The glucose and
insulin contents of the PSP group were lower compared with those of
the HFD group. PSP also increased the adiponectin and decreased the
leptin and resistin levels. Adiponectin represents one potential
contributor to improved insulin sensitivity, which exerts
anti-inflammatory and insulin sensitizing effects in the liver and
adipocytes (39). Resistin levels
are positively associated with increased levels of insulin and
glucose and an elevation of resistin levels induces insulin
resistance in mice (40). Leptin is
another important factor in obesity and inflammation. A reduced
leptin level is considered a beneficial effect because leptin has
pro-inflammatory effects and contributes to the development of
comorbidities of obesity (41).
Lipid metabolic and inflammatory cytokines, as well as the AMPK
signaling pathway, were also altered by PSP.
AMPK is a serine/threonine kinase and plays critical
roles in metabolism (42) and
inflammation (43). AMPK activation
promotes glucose metabolism and adiposis oxidation which is
associated with significant upregulation of autophagy, mitophagy
and mitochondrial metabolism. By contrast, it inhibits
gluconeogenesis and glycogenesis, fatty acid synthesis and
cholesterol synthesis (44).
Obesity is associated with chronic energy imbalance and with
reduced AMPK activation, accompanied with alterations in
glycolysis, hepatic lipid metabolism and inflammation (45).
AMPK downstream genes such as FAS, SREBP-1c, CPT-1
and PPAR-α are associated with lipid biosynthesis and metabolism.
An activation of FAS results in lipid synthesis and deposition
(44). Conversely, SREBP-1c is
negatively correlated with AMPK, meaning that AMPK activity
controls SREBP-1c inhibition (46).
CPT-1 and PPAR-α are the main directors regulating β-oxidation
which is associated with the hepatic diseases (47). In the present study, AMPK signaling
was suppressed in the HFD group and the liver fatty acid and
cholesterol synthesis-associated genes FAS and SREBP-1 were highly
expressed, while CPT-1 and PPAR-α were inhibited by HFD. PSP
significantly activated AMPK signaling, suppressed the expression
of FAS and SREBP-1c and activated CPT-1 and PPAR-α. HFD-induced
obesity is also associated with chronic inflammation (48). In the present study, proinflammatory
cytokines, including TNF-α, IL-6, IL-1β and iNOS, were highly
expressed in the HFD group but significantly inhibited in the PSP
group.
In conclusion, the present study is the first, to
the best of the authors' knowledge, to investigate the effect of
PSP treatment in the prevention of HFD-induced obesity. It is
possible that PSP will have the beneficial property of decreasing
inflammation in the liver, along with protecting against
dyslipidemia, hepatic steatosis and blood glucose increase. It is
hypothesized that the mechanism by which PSP supplementation
protects against lipid accumulation and inflammatory stress might
be due to modulation of the energy balance in the signaling pathway
of AMPK.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81774174), the
Provincial Natural Science Foundation of Hunan (grant no.
2020JJ4345), the China Postdoctoral Foundation (grant no.
2019M652784), the Key Project of the Education Department of Hunan
Province (grant no. 18C0400) and the Open Foundation Research
Center for Standardization and Functional Engineering Technology of
Traditional Chinese Medicine Decoction Pieces of Hunan Province
(grant no. 201806).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BL and JG designed the experiments and wrote the
manuscript; YT performed the animal experiments, histological
examination and statistical analysis; BL and ZS performed the
RT-qPCR and western blotting experiments. BL and JG confirm the
authenticity of all the raw data. JG revised the manuscript and
given final approval of the version to be published. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The care and use of the animals and experimental
protocols complied with the Guidelines for the Care and Use of
Experimental Animals of Hunan University of Chinese Medicine. The
project was approved by the Office of Animal Experiment Ethics of
Hunan University of Chinese Medicine (approval no. 20190165).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
PSP
|
Polygonatum sibiricum F.
Delaroche polysaccharide
|
|
HFD
|
high-fat diet
|
|
TG
|
triacylglycerol
|
|
LDL-C
|
low-density lipoprotein
cholesterol
|
|
HDL-C
|
high-density lipoprotein
cholesterol
|
|
FFAs
|
free fatty acids
|
|
PVDF
|
Polyvinylidene fluoride
|
|
TBST
|
Tris-buffered saline containing 0.1%
Tween-20
|
References
|
1
|
Mishra AK, Dubey V and Ghosh AR: Obesity:
An overview of possible role(s) of gut hormones, lipid sensing and
gut microbiota. Metabolism. 65:48–65. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mokdad AH, Ford ES, Bowman BA, Dietz WH,
Vinicor F, Bales VS and Marks JS: Prevalence of obesity, diabetes,
and obesity-related health risk factors, 2001. Jama. 289:76–79.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Van Gaal LF, Mertens IL and De Block CE:
Mechanisms linking obesity with cardiovascular disease. Nature.
444:875–880. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Peng CH, Yang MY, Yang YS, Yu CC and Wang
CJ: Antrodia cinnamomea prevents obesity, dyslipidemia, and the
derived fatty liver via Regulating AMPK and SREBP Signaling. Am J
Chin Med. 45:67–83. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Despres JP: Is visceral obesity the cause
of the metabolic syndrome? Ann Med. 38:52–63. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Joyal SV: A perspective on the current
strategies for the treatment of obesity. Curr Drug Targets CNS
Neurol Disord. 3:341–356. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Connoley IP, Liu YL, Frost I, Reckless IP,
Heal DJ and Stock MJ: Thermogenic effects of sibutramine and its
metabolites. Br J Pharmacol. 126:1487–1495. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hsieh CJ, Wang PW, Liu RT, Tung SC, Chien
WY, Chen JF, Chen CH, Kuo MC and Hu YH: Orlistat for obesity:
Benefits beyond weight loss. Diabetes Res Clin Pract. 67:78–83.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mead E, Atkinson G, Richter B, Metzendorf
MI, Baur L, Finer N, Corpeleijn E, O'Malley C and Ells LJ: Drug
interventions for the treatment of obesity in children and
adolescents. Cochrane Database Syst Rev. 11:CD0124362016.PubMed/NCBI
|
|
10
|
Nie Y, Lin Q and Luo F: Effects of
non-starch polysaccharides on inflammatory bowel disease. Int J Mol
Sci. 18:13722017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lu Y, Zhao A, Wu Y, Zhao Y and Yang X:
Soybean soluble polysaccharides enhance bioavailability of
genistein and its prevention against obesity and metabolic syndrome
of mice with chronic high fat consumption. Food Funct.
10:4153–4165. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Luo J, Qi J, Wang W, Luo Z, Liu L, Zhang
G, Zhou Q, Liu J and Peng X: Antiobesity effect of flaxseed
polysaccharide via inducing satiety due to leptin resistance
removal and promoting lipid metabolism through the AMP-Activated
Protein Kinase (AMPK) Signaling Pathway. J Agric Food Chem.
67:7040–7049. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wen JJ, Gao H, Hu JL, Nie QX, Chen HH,
Xiong T, Nie SP and Xie MY: Polysaccharides from fermented
Momordica charantia ameliorate obesity in high-fat induced obese
rats. Food Funct. 10:448–457. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao P, Zhao C, Li X, Gao Q, Huang L, Xiao
P and Gao W: The genus Polygonatum: A review of ethnopharmacology,
phytochemistry and pharmacology. J Ethnopharmacol. 214:274–291.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang H, Cao Y, Chen L, Wang J, Tian Q,
Wang N, Liu Z, Li J, Wang N, Wang X, et al: A polysaccharide from
Polygonatum sibiricum attenuates amyloid-β-induced
neurotoxicity in PC12 cells. Carbohydr Polym. 117:879–886. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zong S, Zeng G, Zou B, Li K, Fang Y, Lu L,
Xiao D and Zhang Z: Effects of Polygonatum sibiricum
polysaccharide on the osteogenic differentiation of bone
mesenchymal stem cells in mice. Int J Clin Exp Pathol. 8:6169–6180.
2015.PubMed/NCBI
|
|
17
|
Wang Y, Lan C, Liao X, Chen D, Song W and
Zhang Q: Polygonatum sibiricum polysaccharide potentially
attenuates diabetic retinal injury in a diabetic rat model. J
Diabetes Investig. 10:915–924. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kong X, Liu JJ, Li H and Chen ZB: Effect
of polysaccharides from Polygonatum sibiricum on
lipid-metabolism related mRNA and protein expression in
hyperlipidemic mice. Zhongguo Zhong Yao Za Zhi. 43:3740–3747.
2018.(In Chinese). PubMed/NCBI
|
|
19
|
Jo K, Kim H, Choi HS, Lee SS, Bang MH and
Suh HJ: Isolation of a sleep-promoting compound from Polygonatum
sibiricum rhizome. Food Sci Biotechnol. 27:1833–1842. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cui X, Wang S, Cao H, Guo H, Li Y, Xu F,
Zheng M, Xi X and Han C: A review: The bioactivities and
pharmacological applications of polygonatum sibiricum
polysaccharides. Molecules. 23:11702018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhao X and Li J: Chemical constituents of
the genus Polygonatum and their role in medicinal treatment. Nat
Prod Commun. 10:683–688. 2015.PubMed/NCBI
|
|
22
|
Sun T, Zhang H, Li Y, Liu Y, Dai W, Fang
J, Cao C, Die Y, Liu Q, Wang C, et al: Physicochemical properties
and immunological activities of polysaccharides from both crude and
wine-processed Polygonatum sibiricum. Int J Biol Macromol.
143:255–264. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cai J, Zhu Y, Zuo Y, Tong Q, Zhang Z, Yang
L, Li X and Yi G: Polygonatum sibiricum polysaccharide
alleviates inflammatory cytokines and promotes glucose uptake in
high-glucose and high-insulin-induced 3T3L1 adipocytes by promoting
Nrf2 expression. Mol Med Rep. 20:3951–3958. 2019.PubMed/NCBI
|
|
24
|
Long T, Liu Z, Shang J, Zhou X, Yu S, Tian
H and Bao Y: Polygonatum sibiricum polysaccharides play
anti-cancer effect through TLR4-MAPK/NF-kappaB signaling pathways.
Int J Biol Macromol. 111:813–821. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang Y, Qin S, Pen G, Chen D, Han C, Miao
C, Lu B, Su C, Feng S, Li W, et al: Original Research: Potential
ocular protection and dynamic observation of Polygonatum
sibiricum polysaccharide against streptozocin-induced diabetic
rats' model. Exp Biol Med (Maywood). 242:92–101. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pritchett K, Corrow D, Stockwell J and
Smith A: Euthanasia of neonatal mice with carbon dioxide. Comp Med.
55:275–281. 2005.PubMed/NCBI
|
|
27
|
Liu B, Lin Q, Yang T, Zeng L, Shi L, Chen
Y and Luo F: Oat β-glucan ameliorates dextran sulfate sodium
(DSS)-induced ulcerative colitis in mice. Food Funct. 6:3454–3463.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
GBD 2015 Obesity Collaborators, ; Afshin
A, Forouzanfar MH, Reitsma MB, Sur P, Estep K, Lee A, Marczak L,
Mokdad AH, Moradi-Lakeh M, et al: Health effects of overweight and
obesity in 195 Countries over 25 Years. N Engl J Med. 377:13–27.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Santoliquido A, Di Campli C, Miele L,
Gabrieli ML, Forgione A, Zocco MA, Lupascu A, Di Giorgio A, Flore
R, Pola P, et al: Hepatic steatosis and vascular disease. Eur Rev
Med Pharmacol Sci. 9:269–271. 2005.PubMed/NCBI
|
|
31
|
Burgess E, Hassmen P and Pumpa KL:
Determinants of adherence to lifestyle intervention in adults with
obesity: A systematic review. Clin Obes. 7:123–135. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu B, Yang T, Luo Y, Zeng L, Shi L, Wei
C, Nie Y, Cheng Y, Lin Q and Luo F: Oat β-glucan inhibits
adipogenesis and hepatic steatosis in high fat diet-induced
hyperlipidemic mice via AMPK signaling. J Func Foods. 41:72–82.
2018. View Article : Google Scholar
|
|
33
|
Zhang T, Yang Y, Liang Y, Jiao X and Zhao
C: Beneficial effect of intestinal fermentation of natural
polysaccharides. Nutrients. 10:10552018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hervik AK and Svihus B: The role of fiber
in energy balance. J Nutr Metab. 2019:49836572019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yelithao K, Surayot U, Park W, Lee S, Lee
DH and You S: Effect of sulfation and partial hydrolysis of
polysaccharides from Polygonatum sibiricum on
immune-enhancement. Int J Biol Macromol. 122:10–18. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhu X, Wu W, Chen X, Yang F, Zhang J and
Hou J: Protective effects of Polygonatum sibiricum
polysaccharide on acute heart failure in rats 1. Acta Cir Bras.
33:868–878. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ko JH, Kwon HS, Yoon JM, Yoo JS, Jang HS,
Kim JY, Yeon SW and Kang JH: Effects of Polygonatum
sibiricum rhizome ethanol extract in high-fat diet-fed mice.
Pharm Biol. 53:563–570. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Peng X, He J, Zhao J, Wu Y, Shi X, Du L,
Nong M, Zong S and Zeng G: Polygonatum sibiricum
polysaccharide promotes osteoblastic differentiation through the
ERK/GSK-3β/β-Catenin Signaling Pathway In Vitro. Rejuvenation Res.
21:44–52. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Monk JM, Liddle DM, Hutchinson AL, Wu W,
Lepp D, Ma DW, Robinson LE and Power KA: Fish oil supplementation
to a high-fat diet improves both intestinal health and the systemic
obese phenotype. J Nutr Biochem. 72:1082162019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Steppan CM, Bailey ST, Bhat S, Brown EJ,
Banerjee RR, Wright CM, Patel HR, Ahima RS and Lazar MA: The
hormone resistin links obesity to diabetes. Nature. 409:307–312.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Paz-Filho G, Mastronardi C, Franco CB,
Wang KB, Wong ML and Licinio J: Leptin: Molecular mechanisms,
systemic pro-inflammatory effects, and clinical implications. Arq
Bras Endocrinol Metabol. 56:597–607. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hardie DG: The AMP-activated protein
kinase pathway-new players upstream and downstream. J Cell Sci.
117((Pt 23)): 5479–5487. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Pan MH, Li MY, Tsai ML, Pan CY, Badmaev V,
Ho CT and Lai CS: A mixture of citrus polymethoxyflavones, green
tea polyphenols and lychee extracts attenuates adipogenesis in
3T3-L1 adipocytes and obesity-induced adipose inflammation in mice.
Food Funct. 10:7667–7677. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lyons CL and Roche HM: Nutritional
modulation of AMPK-impact upon metabolic-Inflammation. Int J Mol
Sci. 19:30922018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Garcia D and Shaw RJ: AMPK: Mechanisms of
cellular energy sensing and restoration of metabolic balance. Mol
Cell. 66:789–800. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Song CY, Shi J, Zeng X, Zhang Y, Xie WF
and Chen YX: Sophocarpine alleviates hepatocyte steatosis through
activating AMPK signaling pathway. Toxicol In Vitro. 27:1065–1071.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tailleux A, Wouters K and Staels B: Roles
of PPARs in NAFLD: Potential therapeutic targets. Biochim Biophys
Acta. 1821:809–818. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Cox AJ, West NP and Cripps AW: Obesity,
inflammation, and the gut microbiota. Lancet Diabetes Endocrinol.
3:207–215. 2015. View Article : Google Scholar : PubMed/NCBI
|