Introduction
Sepsis is a systemic inflammatory disease that
occurs after severe trauma, burns, infection and major surgery, and
is associated with multiple organ failure and mortality (1,2). The
excessive inflammation, immunosuppression and tissue damage caused
by sepsis can increase susceptibility to secondary infection and
greatly aggravate the state of illness of a patient, and may
ultimately lead to death (3).
Therefore, the early diagnosis of sepsis and septic shock is
essential for reducing the mortality rate. Although sepsis has
become the leading cause of death in critically ill patients
worldwide, the underlying molecular mechanism of sepsis remains
poorly understood. Therefore, it is of crucial importance to
explore the pathogenesis of the inflammatory response induced by
sepsis.
The genome-wide analysis of gene expression in
critically ill patients has shown that more >80% of basic
genetic factors undergo changes, suggesting that the molecular
basis of sepsis may be associated with regulation at the genetic
level (4). Long non-coding RNAs
(lncRNAs) are non-protein-coding RNAs that are >200 nucleotides
in length (5). The mechanisms of
action of lncRNAs appear diverse, as they can regulate gene
expression at multiple levels and play important roles in tumor
formation, virus replication, inflammatory injury and other
pathological processes. lncRNAs have also gradually been considered
as biomarkers for various diseases, including sepsis (6,7).
Studies have reported on several lncRNAs that participate in
cellular immune regulation, the release of inflammatory factors and
the inflammatory response of sepsis. For example, Chen et al
(8) demonstrated that lncRNA NEAT1
is significantly upregulated in patients with sepsis-induced acute
kidney injury, and that NEAT1 exacerbates lipopolysaccharide
(LPS)-induced cell injury by directly targeting microRNA
(miRNA/miR)-204 and activating the NF-κB pathway. Jia et al
(9) reported that lncRNA CCL2
upregulates the expression of inflammatory factors in macrophages
from septic mice. Growth arrest-specific 5 (GAS5) is a lncRNA that
is closely associated with the regulation of malignant tumors. The
expression of GAS5 is downregulated in renal cell carcinoma, and
its downregulation has been suggested to serve an oncogenic role
(10). Also, GAS5 expression is
negatively correlated with the malignancy of cervical cancer, and
its overexpression has been shown to reduce cell viability and
increase apoptosis in cervical cancer cells (11). Furthermore, GAS5 has been reported
to be involved in cell proliferation, invasion, metastasis,
apoptosis, epithelial-mesenchymal transition and drug resistance
through various molecular mechanisms (12). However, studies concerning the
functional role and mechanisms of GAS5 in sepsis are lacking.
miRNAs are naturally occurring, non-coding,
single-stranded RNA molecules that are 21–25 nucleotides in length,
and are able to bind to mRNA and reduce protein translation
(13). miRNAs are widely involved
in numerous biological processes, including the growth and
development of the organism, substance metabolism, hematopoietic
cell differentiation, tumor formation and immune regulation, and
are expected to become new targets for the treatment of various
diseases. A previous study detected a significant reduction in the
expression of miR-23a-3p in patients with acute kidney injury
caused by sepsis (14). In another
study using myeloid leukemia cells, the downregulation of
miR-23a-3p inhibited cell proliferation, promoted apoptosis and
influenced the cell cycle (15). A
number of studies have shown that lncRNA can cause a series of
changes in biological functions through mutual regulation with
miRNA (16–18). Bioinformatic analysis predicts that
miR-23a-3p is a potential target gene of GAS5; however, the
interaction between GAS5 and miR-23a-3p in sepsis has not yet been
studied.
Accordingly, in the present study, the functional
mechanism of GAS5 was explored to evaluate the effects of GAS5 on
LPS-induced inflammation and cell apoptosis and provide a
theoretical basis for molecular-targeted therapy and the prognostic
diagnosis of sepsis.
Materials and methods
Cell culture and treatment
The THP-1 human monocytic leukemia cell line was
purchased from the American Type Culture Collection. The cells were
cultured in RPMI-1640 medium (HyClone; Cytiva) containing 10% fetal
bovine serum (FBS; HyClone; Cytiva) and 1% penicillin/streptomycin
(Invitrogen; Thermo Fisher Scientific, Inc.) in a humidified
atmosphere of 5% CO2 at 37°C. After subculture, cells in
the logarithmic phase were taken for the following experiments.
To establish the cell sepsis model, cells were
treated with 1 µg/ml LPS for 24 h at 37°C to simulate the sepsis
environment. Reverse transcription-quantitative PCR (RT-qPCR) was
used to detect the expression levels of IL-6, TNF-α and IL-1β to
verify that the sepsis model was constructed successfully.
Cell transfection
Short hairpin RNAs (shRNAs) targeting GAS5
(sh-GAS5-1 and sh-GAS5-2), shRNA control (sh-NC), miR-23a-3p
overexpression vector (miR-23a-3p mimic; cat. no. B01001) and miRNA
negative control (miR-NC; cat. no. B01001) were obtained from
Shanghai GenePharma Co., Ltd. The Toll-like receptor 4 (TLR4)
overexpression vector pcDNA-TLR4 and empty control vector pcDNA-NC
were constructed by Guangzhou RiboBio Co., Ltd. In strict
accordance with the manufacturer's instructions, following LPS
treatment, 1 µg plasmids were transfected into THP-1 cells using
Lipofectamine® 3000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Following incubation for 6 h, the medium was
replaced with DMEM with 10% FBS (HyClone; Cytiva). After 48 h of
incubation, the cells were collectedand the transfection efficiency
was analyzed by RT-qPCR.
Bioinformatics
StarBase v3.0 (http://starbase.sysu.edu.cn/) was used to predict the
target miRNA of GAS5 and the downstream target genes of
miR-23a-3p.
RT-qPCR analysis
The assay was performed in accordance with the
instructions of the respective kits. Total RNAs were isolated from
cells using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Then, the RNAs were reverse transcribed into
cDNA using the PrimeScript RT Reagent kit (Takara Biotechnology
Co., Ltd.). qPCR was then performed using Light Cycler 480 SYBR
Green I Master mix (Roche Applied Science). The reaction was
carried out under the following conditions: Initial denaturation at
95°C for 1 min 40 sec, followed by 42 cycles of denaturation at
94°C for 30 sec, annealing at 55°C for 1 min and elongation at 72°C
for 60 sec, followed by a final extension at 72°C for 5 min.
Relative gene expression was analyzed using the 2−ΔΔCq
method (19). U6 was used as an
endogenous control for miR-23a-3p and GAPDH was used as a control
for other genes. The primer sequences used are as follows: IL-6
forward, 5′-GCAAGGGTCTGGTTTCAGCC-3′ and reverse,
5′-TGAGGTAAGCCTACACTTTCCAA-3′; TNF-a forward,
5′-CCCTCTCTCCCCTGGAAAGG-3′ and reverse, 5′-GCCACTGAATAGGGCGAT-3′;
IL-1b forward, 5′-ATTGCTCAAGTGTCTGAAGCAG-3′ and reverse,
5′-AGAGAGCACACCAGTCCAA-3′; GAS5 forward 5′-CTTCTGGGCTCAAGTGATCCT-3′
and reverse, 5′-TTGTGCCATGAGACTCCATCAG-3′; TLR4 forward,
5′-GGAGACTTGGCCCTAAACCA-3′ and reverse, 5′-GACATGGAAACACACCCAGG-3′;
miR-23a-3p forward 5′-GCGATCACATTGCCAGGG-3′ and reverse,
5′-CAGTGCGTGTCGTGGAGT-3′; GAPDH forward,
5′-CGGAGTCAACGGATTTGGTCGTAT-3′ and reverse,
5′-AGCCTTCTCCATGGTGGTGAAGAC-3′; U6 forward, 5′-CTCGCTTCGGCAGCACA-3′
and reverse, 5′-AACGCTTCACGAATTTGCGT-3′.
ELISA
Experiments were performed according to the protocol
outlined in the IL-6 (cat. no. 130-094-065; Miltenyi Biotec), TNF-α
(cat. no. 130-101-688; Miltenyi Biotec) and IL-1β ELISA kits (cat.
no. 130-094-053; Miltenyi Biotec). Cells were incubated at 37°C for
30 min. According to the manufacturer's protocol: A total of 100 µl
supernatant was added onto the IL-6/TNF-α/IL-1β antibody-coated
plate, and incubated at 25°C for 2 h. After adding the
biotin-conjugated detecting IL-6/TNF-α/IL-1β antibody and
incubating at 25°C for 2 h, streptavidin-HRP was added and
3,3′-5,5′tetramethylbenzidin was used for development, which was
incubated for 20 min at room temperature and protected from light.
The optical density value was measured at 450 nm using a Multiskan
spectrum spectrophotometer (Thermo Fisher Scientific, Inc.).
Experiments were performed in triplicate.
Cell apoptosis analysis
Transfected cells were seeded into 24-well plates at
a density of 1×105 cells/well and routinely incubated
for 48 h. After this, the cells were collected, resuspended with
100 µl 1X binding buffer and 5 µl Annexin V-FITC and 5 µl PI
staining solution (Beyotime Institute of Biotechnology) were added.
The suspension was incubated for 10 min at room temperature in the
dark. Finally, 400 µl of 1X binding buffer was added. The samples
were analyzed by a BD FACSCalibur flow cytometry (BD Biosciences)
and FlowJo software (version 7.6.1; Tree Star, Inc.) within 1 h.
Each measurement was repeated three times and the mean value was
taken.
Luciferase reporter assay
The binding in the GAS5/miR-23a-3p/TLR4 cascade was
detected using a Luciferase Reporter Assay System kit (Promega
Corporation). miR-23a-3p mimic or miR-NC together with the
wild-type (WT) or mutant (MUT) GAS5-3′untranslated region (UTR) or
TLR4-3′UTR were co-transfected into 293T cells (Beyotime Institute
of Biotechnology) using Lipofectamine 3000 (Invitrogen; Thermo
Fisher Scientific, Inc.). Luciferase activity was detected 48 h
after transfection using the dual-luciferase reporter system
(Promega Corporation). Firefly luciferase activity was normalized
to Renilla luciferase activity.
Western blot analysis
Total intracellular protein was extracted from the
cells by lysis with RIPA buffer (Beyotime Institute of
Biotechnology) and the protein concentration was detected using a
BCA kit (Beyotime Institute of Biotechnology). Proteins (30
µg/lane) were separated by 10% SDS-PAGE and transferred to a PVDF
membrane (EMD Millipore). Membranes were then blocked with 5% milk
for 2 h at room temperature and incubated with anti-TLR4 (1:1,000;
cat. no. 14358; Cell Signaling Technology Inc.), anti-Bcl-2 (cat.
no. 4223; 1:1,000; Cell Signaling Technology, Inc.), anti-Bax (cat.
no. 14796; 1:1,000; Cell Signaling Technology, Inc.), anti-cleaved
caspase-3 (cat. no. 9661; 1:1,000; Cell Signaling Technology,
Inc.), anti-caspase-3 (cat. no. 9662; 1:1,000; Cell Signaling
Technology, Inc.), anti-cleaved poly (ADP ribose) polymerase (PARP;
cat. no. 5625; 1:1,000; Cell Signaling Technology, Inc.), anti-PARP
antibodies (cat. no. 9532; 1:1,000; Cell Signaling Technology,
Inc.) and anti-GAPDH (1:1,000; cat. no. 5174; Cell Signaling
Technology Inc.) primary antibodies overnight at 4°C. Membranes
were washed with PBS twice and then incubated with HRP-conjugated
second antibody (cat. no. bs-0295G; 1:2,000; BIOSS) at room
temperature for 2 h. Enhanced chemiluminescence (Chemilucent Plus
western blot enhancing kit; cat. no. 2650; EMD Millipore) was used
for film exposure. The gray value of each band was analyzed using
ImageJ software v1.8.0 (National Institutes of Health). GADPH
served as an internal reference.
Statistical analysis
Data were analyzed using SPSS version 17.0
statistical software (SPSS, Inc.). Measurement data are expressed
as the mean ± standard deviation. An unpaired Student's t-test or
one-way ANOVA followed by Tukey's post hoc test was employed to
examine the difference between two or multiple groups,
respectively. P<0.05 was considered to indicate a statistically
significant difference.
Results
GAS5 is upregulated in LPS-stimulated
THP-1 cells
To test the effects of GAS5 on the inflammation and
apoptosis induced by sepsis, an in vitro cell model of
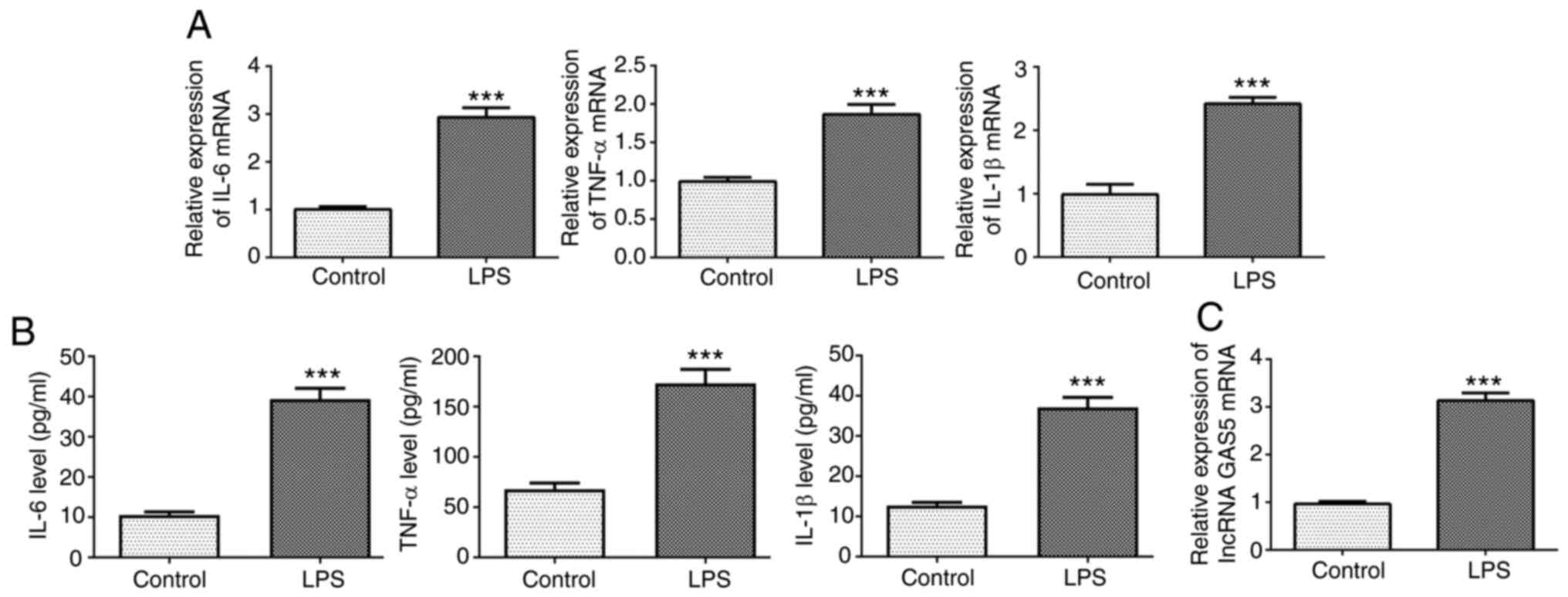
sepsis was established using LPS-induced THP-1 cells. The RT-qPCR
and ELISA results showed that the expression levels of IL-6, TNF-α
and IL-1β in THP-1 cells stimulated with LPS were significantly
increased compared with those in the untreated control group
(Fig. 1A and B). In addition, the
expression of GAS5 was significantly increased in the THP-1 cells
stimulated with LPS compared with the untreated control (Fig. 1C).
GAS5 knockdown alleviates LPS-induced
inflammation and apoptosis in THP-1 cells
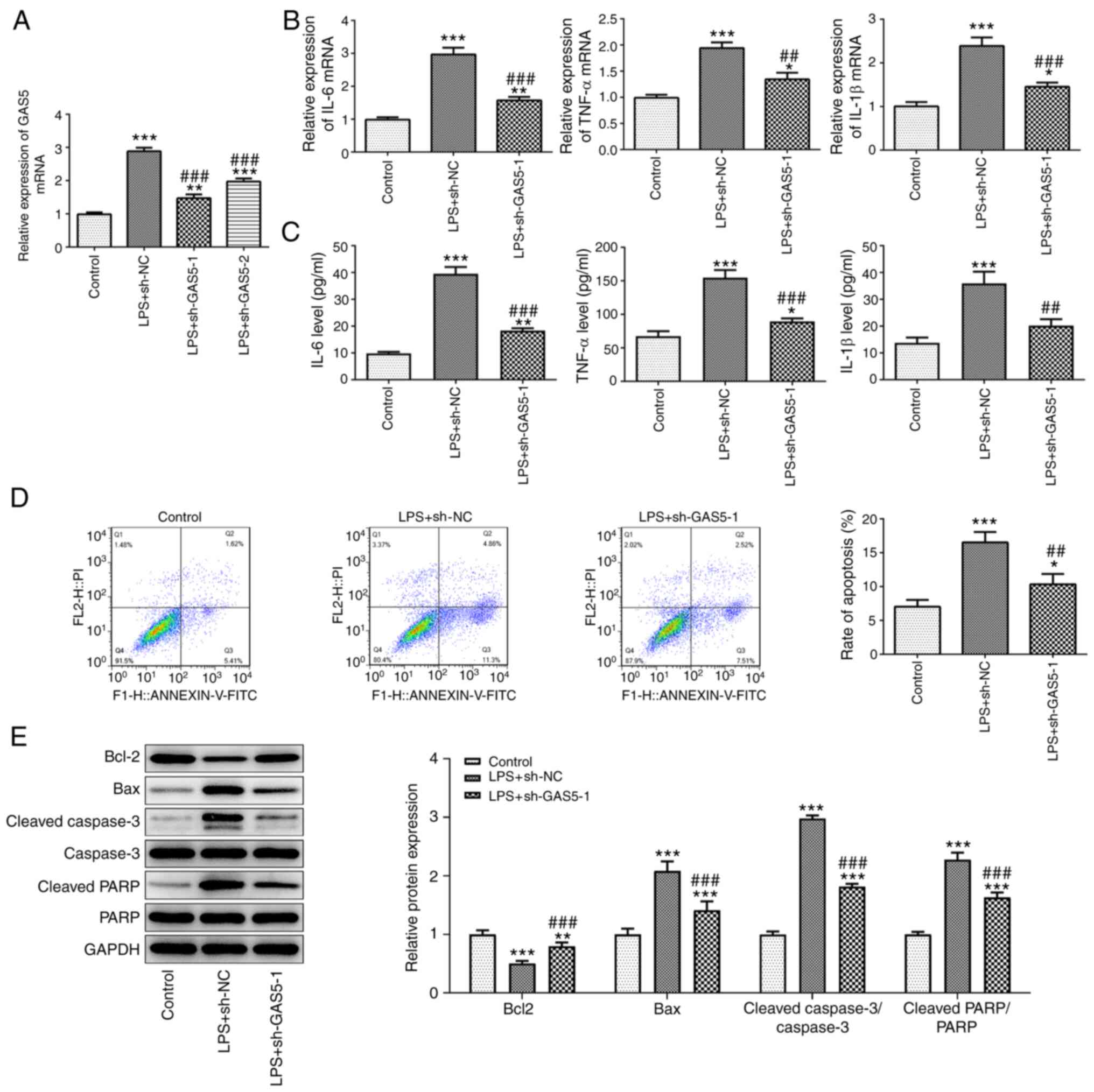
Next, two shRNAs targeting GAS5 were transfected
into THP-1 cells to knockdown the expression of GAS5. As shown in
Fig. 2A, the expression of GAS5 in
the cells transfected with sh-GAS5-1 was the lowest, which
indicated that sh-GAS5-1 had the better knockdown efficiency; thus
sh-GAS5-1 was selected for use in the following experiments. The
effect of GAS5 knockdown on the biological functions of THP-1 cells
treated with LPS was then detected. As shown in Fig. 2B and C, GAS5 knockdown significantly
reduced the release of the inflammatory cytokines IL-6, TNF-α and
IL-1β from THP-1 cells stimulated with LPS. The results of a flow
cytometry assay showed that the increase in cell apoptosis induced
by LPS treatment was attenuated when GAS5 was knocked down
(Fig. 2D). The western blotting
results confirmed the increase in apoptosis in the
sh-GAS5-1-transfected cells (Fig.
2E). Compared with the LPS plus sh-NC group, Bcl-2 was
downregulated and Bax, caspase3 and PARP were upregulated in the
LPS plus sh-GAS5-1 group.
miR-23a-3p is a direct target gene of
GAS5
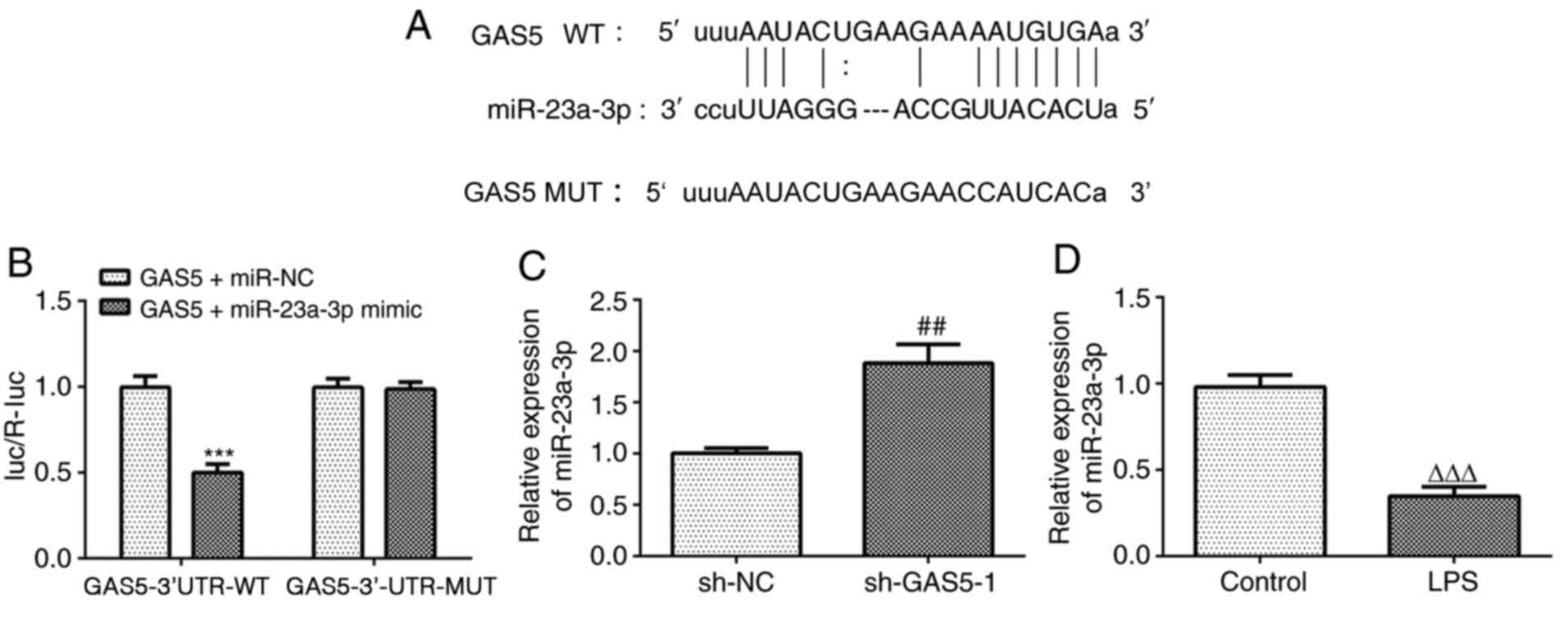
miR-23a-3p was predicted as the target miRNA of GAS5
using the StarBase database. The predicted binding sequence is
shown in Fig. 3A. The results of
the luciferase reporter assay demonstrated that the miR-23a-3p
mimic significantly reduced the luciferase activity of the reporter
vector containing GAS5-3′UTR-WT, but had no effect on the
luciferase activity of the reporter vector containing
GAS5-3′UTR-MUT (Fig. 3B). In
addition, GAS5 knockdown significantly increased the expression of
miR-23a-3p (Fig. 3C). Furthermore,
the expression of miR-23a-3p was downregulated in THP-1 cells
treated with LPS (Fig. 3D). These
results indicate that GAS5 can specifically regulate the expression
of miR-23a-3p and that there is a negative association between GAS5
and miR-23a-3p.
TLR4 is a direct target gene of
miR-23a-3p
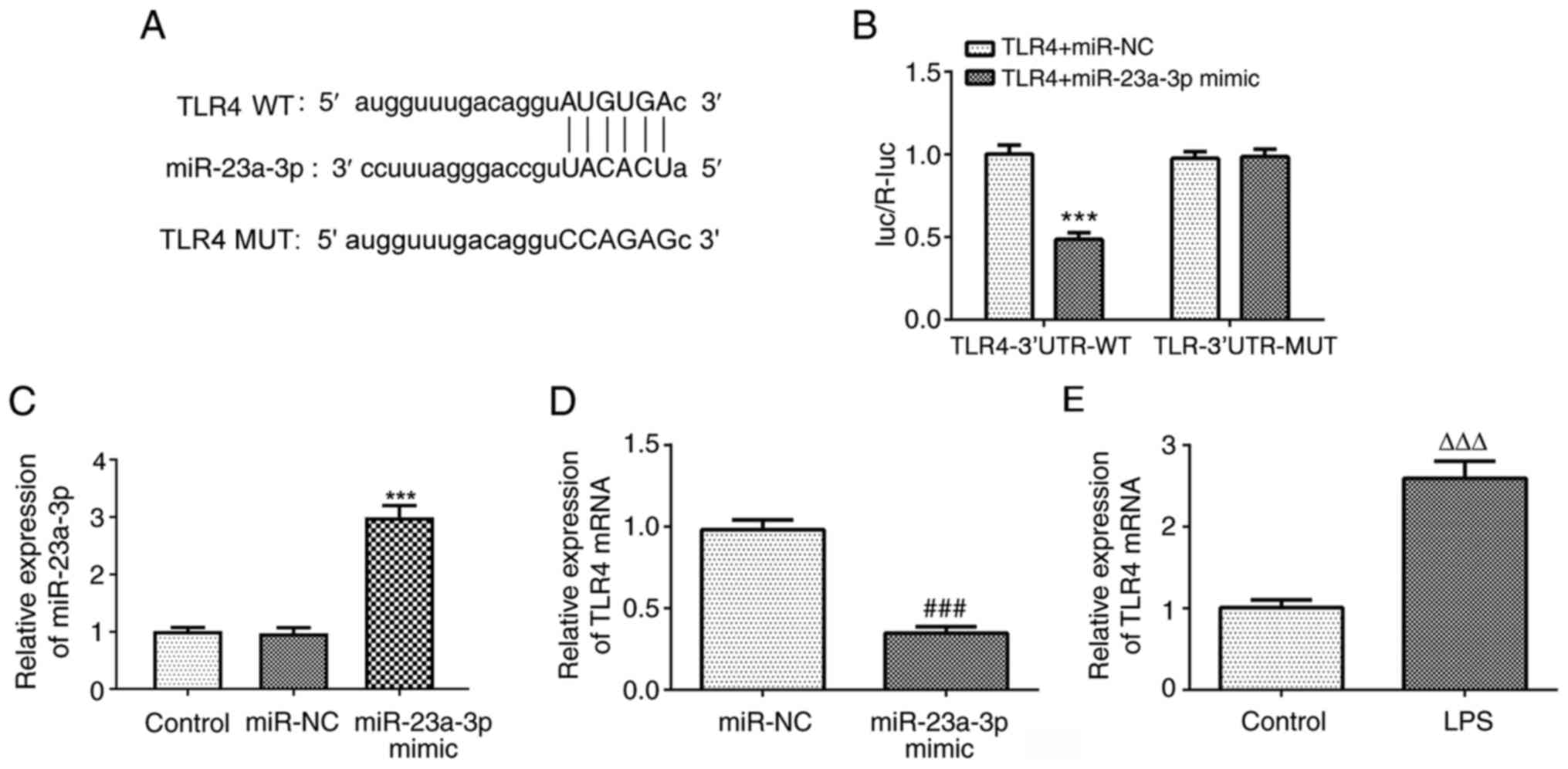
To investigate the downstream target genes of
miR-23a-3p, the StarBase database was used, which predicted that
TLR4 is a potential target gene of miR-23a-3p. The predicted
binding sequence is shown in Fig.
4A. The results of the luciferase reporter assay showed that
the miR-23a-3p mimic significantly reduced the luciferase activity
of the reporter vector containing TLR4-3′UTR-WT, but had no effect
on the luciferase activity of the reporter vector containing
TLR4-3′UTR-MUT (Fig. 4B). The
expression of miR-23a-3p was successfully increased following
transfection with miR-23a-3p mimic (Fig. 4C). Furthermore, miR-23a-3p
overexpression significantly reduced the expression of TLR4
(Fig. 4D). In addition, the
expression of TLR4 was upregulated in THP-1 cells treated with LPS
(Fig. 4E). These results indicate
that miR-23a-3p can specifically regulate the expression of TLR4
and that there is a negative association between miR-23a-3p and
TLR4.
miR-23a-3p affects LPS-induced
inflammation and apoptosis by targeting TLR4
The effects of miR-23a-3p and TLR4 on the biological
functions of THP-1 cells treated with LPS were studied. Firstly, a
TLR4 overexpression plasmid was successfully transfected into THP-1
cells, and the TLR4 overexpression efficiency was verified by
western blotting. The results also showed that miR-23a-3p
overexpression significantly attenuated the increase in expression
of TLR4 induced by the TLR4 plasmid (Fig. 5A). Furthermore, RT-qPCR and ELISA
results revealed that the overexpression of miR-23a-3p
significantly attenuated the LPS-induced expression of the
inflammatory cytokines IL-6, TNF-α and IL-1β in THP-1 cells, and
the overexpression of TLR4 reduced the inhibitory effect of
miR-23a-3p overexpression on the release of inflammatory factors
(Fig. 5B and C). In addition,
miR-23a-3p overexpression significantly suppressed the LPS-induced
apoptosis of THP-1 cells, and the overexpression of TLR4 alleviated
the inhibitory effect of miR-23a-3p overexpression on apoptosis
(Fig. 5D and E).
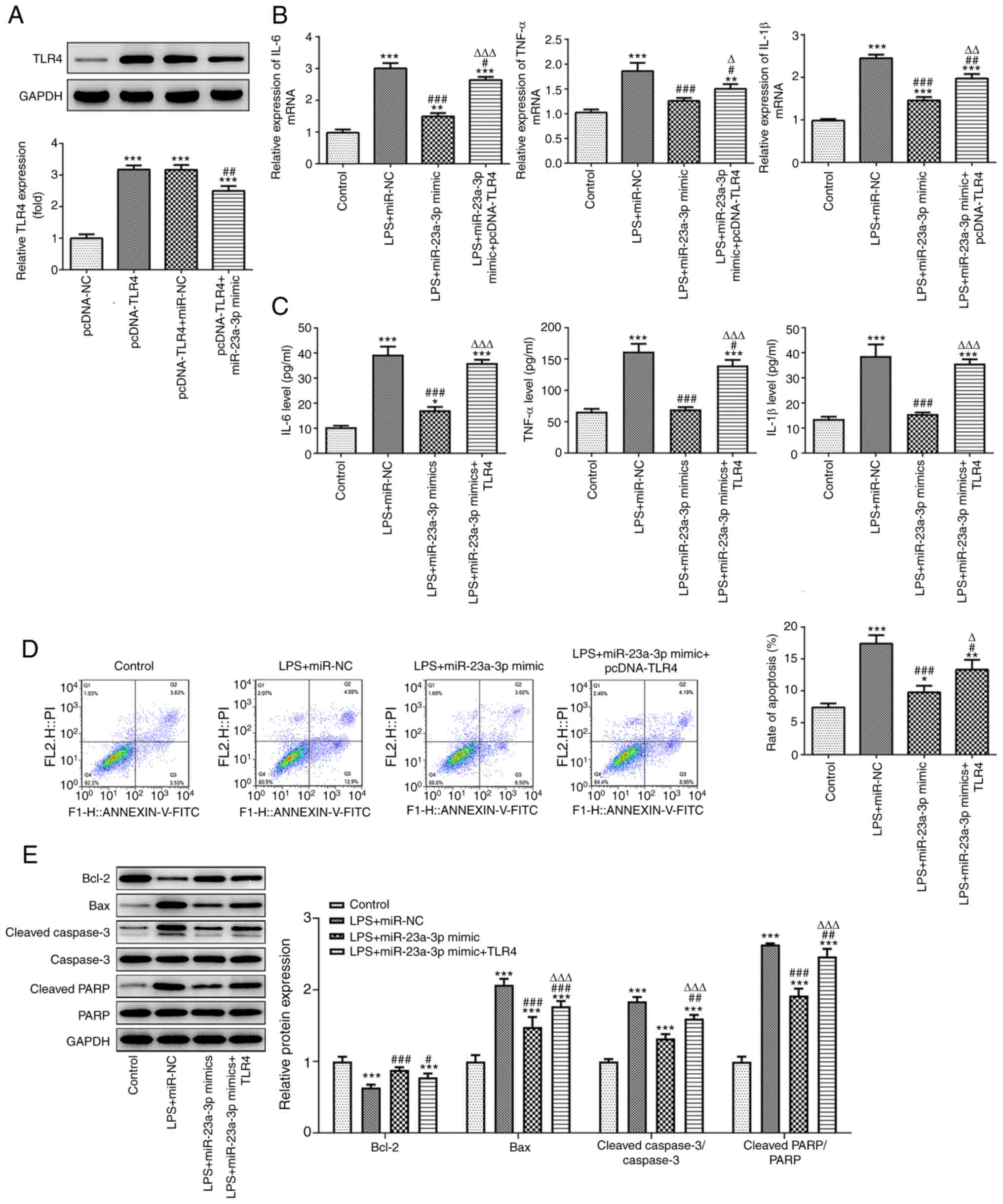 | Figure 5.miR-23a-3p affects LPS-induced
inflammatory and apoptosis by targeting TLR4. (A) The efficiency of
the TLR4 expression plasmid and the effect of miR-23a-3p
overexpression on TLR4 expression were detected by western
blotting. ***P<0.001 vs. pcDNA-NC; ##P<0.01 vs.
pcDNA-TLR4+miR-NC. The inhibitory effect of miR-23a-3p
overexpression on the inflammatory factors IL-6, TNF-α and IL-1β
was mediated via the targeting of TLR4, as revealed by (B) RT-qPCR
and (C) ELISA in LPS-induced THP-1 cells. The inhibitory effect of
miR-23a-3p overexpression on apoptosis was mediated via the
targeting of TLR4, as demonstrated by (D) flow cytometry and (E)
the western blotting of apoptosis-associated proteins in
LPS-induced THP-1 cells. *P<0.05, **P<0.01 and ***P<0.001
vs. control; #P<0.05, ##P<0.01 and
#P<0.05 vs. LPS+miR-NC; ΔP<0.05,
ΔΔP<0.01 and ΔΔΔP<0.001 vs.
LPS+miR-23a-3p mimic. miR, microRNA; LPS, lipopolysaccharide; TLR4,
Toll-like receptor 4; RT-qPCR, reverse transcription-quantitative
PCR; NC, negative control; PARP, poly (ADP ribose) polymerase. |
GAS5 affects LPS-induced inflammation
and apoptosis by regulating the miR-23a-3p/TLR4 pathway
To investigate the effect of GAS5 and TLR4 on the
biological function of THP-1 cells treated with LPS, cells
transfected with sh-GAS5-1 and TLR4 overexpression plasmid were
used. Western blotting analysis showed that GAS5 knockdown
significantly suppressed the expression of TLR4 in LPS-induced
THP-1 cells (Fig. 6A). In addition,
GAS5 knockdown significantly inhibited the expression of the
inflammatory cytokines IL-6, TNF-α and IL-1β in LPS-treated THP-1
cells, and the overexpression of TLR4 attenuated the inhibitory
effect of GAS5 knockdown on the release of inflammatory factors
(Fig. 6B and C). Furthermore, flow
cytometry and western blotting results demonstrated that TLR4
overexpression reduced the inhibitory effect of GAS5 knockdown on
apoptosis (Fig. 6D and E). These
results demonstrate that the overexpression of TLR4 can reverse the
inhibitory effects of GAS5 knockdown and miR-23a-3p overexpression
on the inflammatory response and apoptosis of THP-1 cells. In
particular, they suggest that GAS5 may affect LPS-induced
inflammation and apoptosis by regulating the miR-23a-3p/TLR4
pathway.
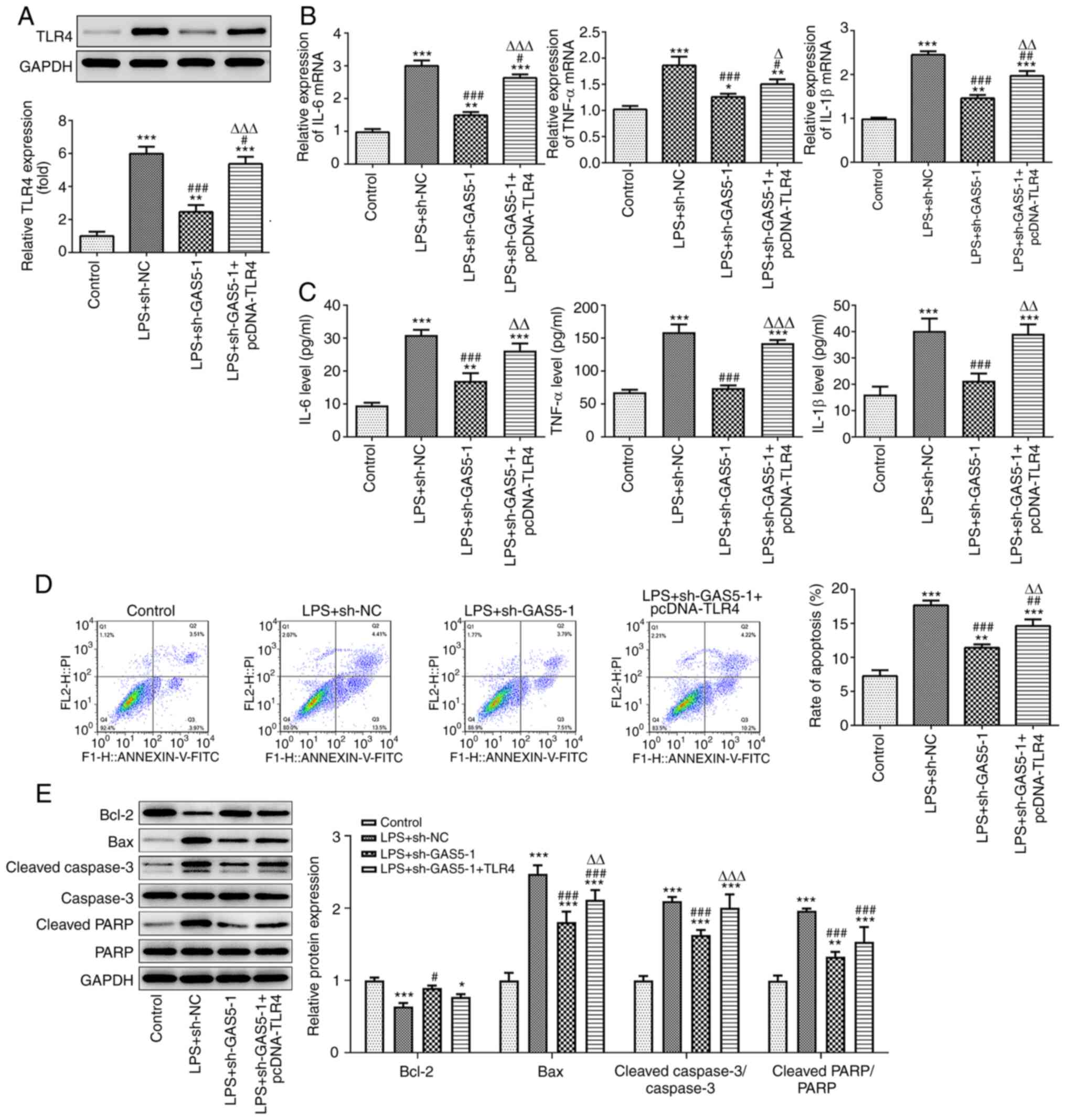 | Figure 6.GAS5 affects LPS-induced inflammation
and apoptosis by regulating the miR-23a-3p/TLR4 pathway. (A) The
effect of GAS5 knockdown on TLR4 expression was detected by western
blotting. The inhibitory effect of GAS5 knockdown on the
inflammatory factors IL-6, TNF-α and IL-1β was mediated via the
regulation of TLR4, as revealed by (B) RT-qPCR and (C) ELISA in
LPS-induced THP-1 cells. The inhibitory effect of GAS5 knockdown on
apoptosis was mediated via the regulation of TLR4, as demonstrated
by (D) flow cytometry and (E) the western blotting of
apoptosis-associated proteins in LPS-induced THP-1 cells.
*P<0.05, **P<0.01 and ***P<0.001 vs. control;
#P<0.05, ##P<0.01 and
###P<0.001 vs. LPS+sh-NC; ΔP<0.05,
ΔΔP<0.01 and ΔΔΔP<0.001 vs. LPS +
sh-GAS5-1. GAS5, growth arrest-specific 5; LPS, lipopolysaccharide;
miR, microRNA; TLR4, Toll-like receptor 4; RT-qPCR, reverse
transcription-quantitative PCR; NC, negative control; PARP, poly
(ADP ribose) polymerase. |
Discussion
An LPS-induced THP-1 cell sepsis model was used in
the present study to investigate the expression and mechanism of
lncRNA GAS5. The results revealed that the expression of GAS5 was
significantly increased in THP-1 cells induced by LPS and was
accompanied by the downregulation of miR-23a-3p and upregulation of
TLR4, as well as increases in the release of pro-inflammatory
factors and the number of apoptotic cells. Further experiments
showed that both GAS5 and TLR4 target miR-23a-3p. However, the
overexpression of TLR4 was demonstrated to attenuate the protective
effects of GAS5 knockdown and miR-23a-3p overexpression on
inflammatory injury. These results suggest that the damaging effect
of GAS5 on LPS-induced THP-1 cells is mediated via the negative
regulation of miR-23a-3p, which subsequently increases the
expression of TLR4.
The lncRNA GAS5 was first discovered by Schneider
et al (20) in NIH3T3 mouse
fibroblasts, who found that serum-starved cells showed growth
arrest accompanied by an increase in the expression of GAS5.
Further research has shown that GAS5 is aberrantly expressed in a
variety of tumors and acts as a tumor suppressor (21,22).
However, GAS5 is seldom studied in fields outside oncology.
Notably, a previous study showed that the knockdown GAS5 reduced
the inflammatory injury of cardiomyocytes induced by high glucose
(23). This anti-inflammatory
property suggests that an investigation into the potential
inhibitory effects of GAS5 knockdown on the inflammatory injury in
sepsis is merited. In the present study, the role of GAS5 in sepsis
was evaluated for the first time, to the best of our knowledge. The
results showed that the expression levels of GAS5 and inflammatory
factors were significantly increased in THP-1 cells following
treatment with LPS, and that GAS5 knockdown suppressed the
inflammatory reaction and apoptosis in the LPS-induced THP-1 cells.
This suggests that GAS5 may be involved in the occurrence and
development of the inflammatory response caused by sepsis.
TLR4 is expressed on the surfaces of numerous kinds
of cells, including neutrophils and macrophages, and participates
in the regulation of various physiological functions of cells.
Previous studies have shown that the upregulation of TLR4 promotes
an inflammatory response and inhibits cell growth (24,25).
In another study, miR-23a-3p was reported to inhibit monocyte
function and phagocytosis by targeting TLR4/TNF-α/TGF-β1/IL-10
signaling in patients with active tuberculosis with a high
bacterial load via interferon regulatory factor 1/transcription
factor SP1 (26). In addition, the
downregulation of miR-21-5p in patients with obstructive sleep
apnea has been shown to regulate intermittent hypoxia and
reoxygenation-induced apoptosis and cytotoxicity by targeting
pro-inflammatory TNF-α/TLR4 signaling (27). In the present study, TLR4 was
identified as a gene that is directly targeted by miR-23a-3p, and a
clear negative association between miR-23a-3p and TLR4 expression
was observed. The overexpression of miR-23a-3p significantly
reduced the expression of the inflammatory factors IL-6, TNF-α and
IL-1β, as well as apoptosis, and the overexpression TLR4 was able
to reverse this change. This suggests that miR-23a-3p and TLR4 act
antagonistically to regulate the inflammatory response as follows:
miR-23a-3p inhibits the inflammatory response induced by sepsis,
while TLR4 promotes the inflammatory response.
A number of studies have shown that lncRNA and miRNA
can regulate each other and participate in the occurrence and
development of a variety of diseases by forming a complex molecular
regulatory network (28,29). The results of the present study
demonstrate that GAS5 targets miR-23a-3p and negatively regulate
its expression, and that miR-23a-3p and TLR4 antagonistically
regulate the inflammatory response induced by sepsis. Experiments
in which the GAS5 knockdown plasmid and TLR4 overexpression plasmid
were cotransfected into THP-1 cells demonstrated that TLR4
overexpression attenuated the reductions in inflammatory cytokine
levels and apoptosis caused by GAS5 knockdown. Together, these
results indicate that GAS5 promotes sepsis-induced inflammation and
apoptosis by negatively regulating miR-23a-3p and thereby
increasing the expression of TLR4.
In summary, the present study demonstrated that GAS5
is upregulated in LPS-treated THP-1 cells and appears to promote
the inflammation and apoptosis of LPS-induced THP-1 cells by
modulating the miR-23a-3p/TLR4 axis. Moreover, the
GAS5/miR-23a-3p/TLR4 axis may become a new potential therapeutic
target in the treatment of sepsis. However, there are some
limitations to the present study. Firstly, only in vitro
experiments were conducted and no in vivo experiments were
performed for validation. In future research, animal models and
clinical samples may be used to confirm the findings of the present
study. Secondly, the mechanisms underlying the roles of lncRNA GAS5
and miR-23a-3p in the progression of sepsis have not been fully
investigated. Therefore, these issues require further in-depth
study in the future.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
DH designed the study and was mainly involved in the
bioinformatics and data analysis. ZG mainly conducted most of the
experiments. Both authors read and approved the final manuscript.
ZG and DH confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wu H, Liu J, Li W, Liu G and Li Z:
LncRNA-HOTAIR promotes TNF-α production in cardiomyocytes of
LPS-induced sepsis mice by activating NF-κB pathway. Biochem
Biophys Res Commun. 471:240–246. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Singer M, Deutschman CS, Seymour CW,
Shankar-Hari M, Annane D, Bauer M, Bellomo R, Bernard GR, Chiche
JD, Coopersmith CM, et al: The third international consensus
definitions for sepsis and septic shock (Sepsis-3). JAMA.
315:801–810. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cawcutt KA and Peters SG: Severe sepsis
and septic shock: Clinical overview and update on management. Mayo
Clin Proc. 89:1572–1578. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xiao W, Mindrinos MN, Seok J, Cuschieri J,
Cuenca AG, Gao H, Hayden DL, Hennessy L, Moore EE, Minei JP, et al:
A genomic storm in critically injured humans. J Exp Med.
208:2581–2590. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rinn JL and Chang HY: Genome regulation by
long noncoding RNAs. Annu Rev Biochem. 81:145–166. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dey BK, Mueller AC and Dutta A: Long
non-coding RNAs as emerging regulators of differentiation,
development, and disease. Transcription. 5:e9440142014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wapinski O and Chang HY: Long noncoding
RNAs and human disease. Trends Cell Biol. 21:354–361. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen Y, Qiu J, Chen B, Lin Y, Chen Y, Xie
G, Qiu J, Tong H and Jiang D: Long non-coding RNA NEAT1 plays an
important role in sepsis-induced acute kidney injury by targeting
miR-204 and modulating the NF-κB pathway. Int Immunopharmacol.
59:252–260. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jia Y, Li Z, Cai W, Xiao D, Han S, Han F,
Bai X, Wang K, Liu Y, Li X, et al: SIRT1 regulates inflammation
response of macrophages in sepsis mediated by long noncoding RNA.
Biochim Biophys Acta Mol Basis Dis. 1864:784–792. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Qiao HP, Gao WS, Huo JX and Yang ZS: Long
non-coding RNA GAS5 functions as a tumor suppressor in renal cell
carcinoma. Asian Pac J Cancer Prev. 14:1077–1082. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yao T, Lu R, Zhang J, Fang X, Fan L, Huang
C, Lin R and Lin Z: Growth arrest-specific 5 attenuates
cisplatin-induced apoptosis in cervical cancer by regulating STAT3
signaling via miR-21. J Cell Physiol. 234:9605–9615. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ghaforui-Fard S and Taheri M: Growth
arrest specific transcript 5 in tumorigenesis process: An update on
the expression pattern and genomic variants. Biomed Pharmacother.
112:1087232019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ge QM, Huang CM, Zhu XY, Bian F and Pan
SM: Differentially expressed miRNAs in sepsis-induced acute kidney
injury target oxidative stress and mitochondrial dysfunction
pathways. PLoS One. 12:e01732922017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao C, Wang S, Zhao Y, Du F, Wang W, Lv P
and Qi L: Long noncoding RNA NEAT1 modulates cell proliferation and
apoptosis by regulating miR-23a-3p/SMC1A in acute myeloid leukemia.
J Cell Physiol. 234:6161–6172. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Su Z, Zhi X, Zhang Q, Yang L, Xu H and Xu
Z: LncRNA H19 functions as a competing endogenous RNA to regulate
AQP3 expression by sponging miR-874 in the intestinal barrier. FEBS
Lett. 590:1354–1364. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang JY, Yang Y, Ma Y, Wang F, Xue A, Zhu
J, Yang H, Chen Q, Chen M, Ye L, et al: Potential regulatory role
of lncRNA-miRNA-mRNA axis in osteosarcoma. Biomed Pharmacother.
121:1096272020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang Y, Yujiao W, Fang W, Linhui Y, Ziqi
G, Zhichen W, Zirui W and Shengwang W: The roles of miRNA, lncRNA
and circRNA in the development of osteoporosis. Biol Res.
53:402020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schneider C, King RM and Philipson L:
Genes specifically expressed at growth arrest of mammalian cells.
Cell. 54:787–793. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pickard MR, Mourtada-Maarabouni M and
Williams GT: Long non-coding RNA GAS5 regulates apoptosis in
prostate cancer cell lines. Biochim Biophys Acta. 1832:1613–1623.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mourtada-Maarabouni M, Pickard MR, Hedge
VL, Farzaneh F and Williams GT: GAS5, a non-protein-coding RNA,
controls apoptosis and is downregulated in breast cancer. Oncogene.
28:195–208. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao J, Liu B and Li C: Knockdown of long
noncoding RNA GAS5 protects human cardiomyocyte-like AC16 cells
against high glucose-induced inflammation by inhibiting
miR-21-5p-mediated TLR4/NF-κB signaling. Naunyn Schmiedebergs Arch
Pharmacol. 393:1541–1547. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang J, Guo X, Yang J, Ding JW, Li S, Yang
R, Fan ZX and Yang CJ: RP105 protects against apoptosis in
ischemia/reperfusion-induced myocardial damage in rats by
suppressing TLR4-mediated signaling pathways. Cell Physiol Biochem.
36:2137–2148. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hsu RY, Chan CH, Spicer JD, Rousseau MC,
Giannias B, Rousseau S and Ferri LE: LPS-induced TLR4 signaling in
human colorectal cancer cells increases beta1 integrin-mediated
cell adhesion and liver metastasis. Cancer Res. 71:1989–1998. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen YC, Lee CP, Hsiao CC, Hsu PY, Wang
TY, Wu CC, Chao TY, Leung SY, Chang YP and Lin MC: MicroRNA-23a-3p
down-regulation in active pulmonary tuberculosis patients with high
bacterial burden inhibits mononuclear cell function and
phagocytosis through TLR4/TNF-α/TGF-β1/IL-10 signaling via
targeting IRF1/SP1. Int J Mol Sci. 21:85872020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen YC, Hsu PY, Su MC, Chin CH, Liou CW,
Wang TY, Lin YY, Lee CP, Lin MC and Hsiao CC: miR-21-5p
under-expression in patients with obstructive sleep apnea modulates
intermittent hypoxia with Re-oxygenation-induced-cell apoptosis and
cytotoxicity by targeting pro-inflammatory TNF-α-TLR4 signaling.
Int J Mol Sci. 21:9992020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Karreth FA and Pandolfi PP: ceRNA
cross-talk in cancer: When ce-bling rivalries go awry. Cancer
Discov. 3:1113–1121. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu X, Sun M, Nie FQ, Ge YB, Zhang EB, Yin
DD, Kong R, Xia R, Lu KH, Li JH, et al: Lnc RNA HOTAIR functions as
a competing endogenous RNA to regulate HER2 expression by sponging
miR-331-3p in gastric cancer. Mol Cancer. 13:922014. View Article : Google Scholar : PubMed/NCBI
|