Introduction
Alzheimer's disease (AD) is a degenerative disease
of the central nervous system (CNS) and is characterized by
progressive memory loss and cognitive dysfunction (1). The neurological dysfunction observed
in AD is accompanied by various cognitive symptoms and behavioral
abnormalities that seriously affect the daily life and work of
patients (1). The main pathological
features of AD are amyloid plaques or senile plaques that are
composed of extracellular β-amyloid protein (Aβ) and
neurofibrillary tangles (NFT), formed by abnormal accumulation in
cells of hyperphosphorylated tau protein (2). As the final manifestation of AD,
neuron synaptic loss is primarily associated with the abnormal
signal transmission of excitatory glutamate (Glu) and
depolymerization of the cytoskeleton (3).
The N-methyl-D-aspartate receptors (NMDARs) are
widely distributed on postsynaptic membranes of neurons in the CNS,
particularly in the hippocampus (HC). NMDARs are essential signal
mediators and play critical roles in synaptic transmission and
synaptic plasticity (4,5). Over-activation of NMDARs leads to the
opening of Ca2+ channels coupled with Glu receptors,
resulting in increases in the intracellular concentration of
Ca2+, which activates Ca2+/CaMKII (6). Phosphorylated (p) CaMKII directly
activate cyclic adenosine monophosphate (cAMP) response element
binding protein (CREB), a key regulator of long-term memory
(7,8). p-CREB further regulates the synthesis
of activity-regulated cytoskeleton-associated protein and synapsin,
which participate in the formation of long-term-potentiation (LTP)
and maintain the formation of long-term memory (9). Brain-derived neurotrophic factor is an
essential target gene of CREB, which affects the formation of
synapses and neuronal plasticity and plays a role in promoting the
growth and development of neurons (10,11).
Therefore, the Ca2+/CaMKII/CREB signaling pathway is
closely interconnected with synaptic plasticity.
It is widely accepted that the inflammatory response
induced by the activation of Aβ deposition may be the most
important pathological mechanism of AD (12). Accumulating evidence indicates that,
as the central element of the inflammatory response, the
inflammasome is closely related to numerous immune inflammatory and
metabolic diseases. Furthermore, it plays a vital role in the
occurrence and development of nervous system diseases (13,14).
Evidence suggests that Aβ and other abnormal aggregation proteins
activate the inflammasome and promote the maturation and secretion
of critical inflammatory factors, such as IL-1β that participate in
the internal immune inflammatory response and cause pyroptosis,
which is an essential natural immune response (15,16).
Additionally, Aβ causes K+ to flow out of neurons and a
low K+ concentration is an effective activator of the
NOD-, LRR- and pyrin domain-containing protein 3 (NLRP3)
inflammasome (17). The activated
NLRP3 inflammasome participates in the neurotoxicity caused by
pyroptosis, which aggravates neurodegenerative diseases and causes
progressive cognitive impairment (18,19).
Microglia are important recruiters and executors of
the inflammatory response in the brain (20,21).
Once activated, microglia produce pro-inflammatory cytokines and
other cytotoxic mediators, ultimately affecting the normal function
of the brain (22). Previous
studies have revealed that microglia may participate in the
formation of synaptic connections between neurons and play a key
role in the regulation of synaptic plasticity in the brain
(23,24).
κ-opioid receptor (KOR) agonists are widely used in
perioperative analgesia due to their strong analgesic effects; they
also regulate emotional and cognitive functions (25). The KOR agonist, U50488H was found to
reduce cognitive impairment significantly (26). Therefore, the present study
investigated the effects of U50488H on spatial memory, synaptic
plasticity and inflammatory cells in an amyloid precursor protein
(APP)/presenilin-1 (PS1) mouse model. Synaptic function was
assessed by examining the expression level of postsynaptic density
protein 95 (PSD95). Furthermore, the underlying mechanism of
microglia pyroptosis in AD and the regulatory mechanism of the
Ca2+/CaMKII/CREB signaling pathway in synaptic
plasticity were investigated. It was identified that the KOR
agonist, U50488H regulated NLRP3 via the
Ca2+/CaMKII/CREB signaling pathway, inhibiting
microglial cell pyroptosis and improving synaptic plasticity in
APP/PS1 mice.
Materials and methods
Animals and experimental groups
In total, 45 male (weight, 25–30 g; age, 6 months)
AD mice expressing human APP and PS1 genes and 15 C57BL/6 mice
(age, 6 months) were obtained from the Experimental Animal Center
of the China Medical University. The present study was approved by
the Animal Welfare and Ethics Committee of the China Medical
University Laboratory (Institutional Animal Care and Use Committees
approval no. 2018236). All mice were offered food and water ad
libitum and housed in pathogen-free facilities under a 12-h
light/dark cycle in a controlled room temperature of 12–24°C and
60% relative humidity. The mice used in the present study exhibited
amyloid plaques and NFT from age 9–12 months. Upon completion of
the experiments, mice were euthanized by intraperitoneal injection
of 50 mg/kg body weight sodium pentobarbital, followed by heart
perfusion or dislocation of the cervical spine. Brain tissue
samples were collected from wild-type or APP/PS1 transgenic mice.
The pathology was primarily localized to the HC, amygdala and
cerebral cortex. To reduce potential variations among the
experimental mice, the mice used in the present study were first
grouped by similar weight and at 12-months old. Then mice from each
litter were equally distributed to the different study groups.
Specifically, the mice were randomly divided into four groups as
follows: i) Wild-type mice (control group; n=15); ii) APP/PS1
transgenic mice (AD group; n=15); iii) APP/PS1 transgenic mice were
treated with the κ-opioid receptor agonist, U50488H for 28 days
(1.25 mg/kg) using an osmotic pump (U50488H group; n=15); iv)
APP/PS1 transgenic mice were treated with U50488H for 28 days (1.25
mg/kg) using an osmotic pump and were injected through an
intracerebral pump injection with CaMKII antagonist, KN93 (5
µM/day) for 28 days (KN93 group; n=15). KN93 (Sigma-Aldrich; Merck
KGaA) was dissolved in 0.9% saline containing 1% DMSO and diluted
to a concentration of 1 mM.
Morris water maze
Spatial memory was assessed using the Morris water
maze test, which included the concealed platform test and the space
exploration test (27,28). Before each trial period, the mice
were brought to the room with the water maze to allow for
acclimation. For spatial learning, the mice were trained for five
consecutive days to find a hidden platform in the Morris water
maze. During each trial, the mouse started from the middle of one
of the four quadrants, facing the wall of the pool. The trial ended
when the animal climbed onto the platform (diameter, 10 cm). The
mice were not allowed to search for the platform for more than 60
sec, after which they were guided to the platform. For the space
exploration test, the platform was removed after the end of the
hidden platform test. Then, 24 h after the hidden platform test,
the mice were placed in the same starting position as the hidden
platform test and their swimming paths were recorded for 60 sec.
The time each mouse spent in the original quadrant and the number
of times each mouse crossed the original platform location were
recorded. The Morris water maze video analysis system was used for
data processing.
Immunohistochemistry
Upon completion of the water maze experiments, five
mice from each group were anesthetized by intraperitoneal injection
with 50 mg/kg sodium pentobarbital. Following anesthesia, mice were
perfused through the heart with PBS and then with pre-cooled 4%
(w/v) paraformaldehyde, then the mouse's brain was rapidly removed.
The brain tissue was placed in an embedding box, 4%
paraformaldehyde was added and the box was placed in a freezer at
−80°C for 24–48 h for rapid freezing. A cold microtome was used to
slice the tissue into 5-µm sections. The sections were
deparaffinized and rehydrated through a graded series of alcohol.
Hydrogen peroxide (3%) solution was used to inactivate endogenous
peroxidases and the sections were washed once with PBS (pH 7.4).
The sections were exposed to a 0.1 M sodium citrate solution for
antigen retrieval. Next, the sections were incubated overnight at
4°C in Aβ antibody (cat. no. sc-28365; 1:1,000; Santa Cruz
Biotechnology, Inc.). The next day the sections were washed three
times in PBS, incubated with a biotin-labeled secondary antibody
(cat. no. sc-525409; 1:1,000; Santa Cruz Biotechnology, Inc.) at
37°C for 30 min, then washed again in PBS. The
3,3′-diaminobenzidine, as a chromogen, was used to stain sections
for 3 min at room temperature to visualize the Aβ-positive cell
staining. The cell nuclei were counterstained with hematoxylin, a
neutral resin was used to seal coverslips onto the microscope
slides and the stained sections were observed with a light
microscope (Olympus Corporation) with a magnification of ×40.
Hematoxylin and eosin (H&E)
staining
The sections that were mounted on glass microscope
slides were heated to melt the paraffin. Then the sections were
deparaffinized and rehydrated through a series of graded alcohol.
The tissue sections were rinsed with tap water for 10 min.
Hematoxylin staining was performed for 2 min at room temperature
and the sections were rinsed with tap water for 10 min and immersed
in 1% hydrochloric alcohol for 3 sec. Eosin staining was performed
for 1 min at room temperature, then 95% ethanol followed by two
changes of anhydrous ethanol were used to dehydrate the sections.
The sections then were immersed in two changes of xylene for 5 min
each. Finally, a neutral mounting medium was used to seal
coverslips over the sections. Pathological changes in the tissue
sections were observed using a light microscope (Olympus
Corporation) with a magnification of ×40.
ELISA assay
At the end of the water maze experiment, five mice
in each group were sacrificed by dislocating the cervical spine.
The brain of the mice was collected. The brain tissue was lysed in
Laemmli buffer with homogenization and sonication three time for 10
sec/time on ice (QSonica LLC). Following tissue homogenization, the
Glu (cat. no. CES122Ge; Wuhan USCN Business Co., Ltd.), collagen II
(cat. no. CB85527920; Boswio; http://www.boswio.com), IL-18 (cat. no. SEA064Mu;
Wuhan USCN Business Co., Ltd.) and IL-1β (cat. no. SEA563Mu; Wuhan
USCN Business Co., Ltd.) content was detected using ELISA kits
according to the manufacturer's instructions. The kit was
equilibrated to room temperature and the required reaction plate
was removed. Then the standards and diluted samples were added into
the wells of the corresponding reaction plate and incubated at room
temperature for 20 min. Subsequently, the reaction plate was
washed, HRP-labeled secondary antibody provided in the kit was
added to each well and incubated at 37°C for 30 min. The plate was
rinsed, and the color developing solution was added in the dark.
The plate was incubated for 15 min at room temperature, and then
the stop solution was added to each well. The optical density (OD)
value at 450 nm was read using a microplate reader. The OD value
was used as the vertical coordinate and the standard concentrations
were used as the horizontal coordinate. A standard curve was
constructed and the curve equation and R-value were calculated to
determine the corresponding concentration values for each
sample.
Golgi-Cox staining
The brain tissue samples were immersed in a 30%
sucrose solution overnight followed by immersion in optimal cutting
temperature embedding reagent at 4°C for 6 h, then stored at −80°C
for 24 h and sectioned (thickness, 100 µm), according to a previous
publication (27). The Golgi-Cox
staining procedure was performed according to the instructions from
the FD Rapid GolgiStain™ kit (FD Neurotechnologies, Inc.). The
dendritic spine density of neurons was analyzed using ImageJ
software (version 1.52a; National Institutes of Health).
Western blotting
The brain tissue samples were homogenized in
iced-cold RIPA buffer containing protease inhibitor (Santa Cruz
Biotechnology, Inc.) and protein concentration was estimated using
BCA reagent. Then, 30 mg protein lysates per lane were loaded,
separated by 6–12% SDS-PAGE gradient gels and transferred onto
nitrocellulose membranes, then the membranes were blocked in a 5%
skimmed milk solution at room temperature for 1 h. Tris-buffered
saline with 0.1% Tween (TBST) was used to wash the membranes and
then the membranes were incubated with the primary antibody
including anti-NMDAR (1:1,000; cat. no. ab274377; Abcam),
anti-p-CaMKII [1:1,000; cat. no. 12716S; Cell Signaling Technology,
Inc. (CST)], anti-CaMKII (1:1,000; cat. no. 3362S; CST),
anti-p-CREB (1:1,000; cat. no. 9198S; CST), anti-CREB (1:1,000;
cat. no. 9197S; CST), anti-NCAM (1:1,000; cat. no. 99746S; CST),
anti-NR2B (1:1,000; cat. no. ab254356; Abcam), anti-GluR1 (1:1,000;
ab183797; Abcam), anti-SYN (1:1,000; cat. no. ab212184; Abcam),
anti-PSD95 (1:1,000; cat. no. 3409S; CST), anti-GAPDH (1:1,000;
cat. no. 2118S; CST), anti-pro-caspase-1 (1:1,000; cat. no. 24232S;
CST), anti-pro-IL-1β (1:1,000; cat. no. 31202S; CST), anti-ASC
(1:1,000; cat. no. 67824S; CST) and NLRP3 (1:1,000; cat. no.
15101S; CST) overnight at 4°C. The PVDF membranes were washed three
times in PBST, incubated with anti-rabbit IgG, HRP-linked antibody
(1:1,000; cat. no. 7074S; CST) at room temperature for 1 h and the
PVDF membranes were washed in TBST. The labeled protein bands were
visualized using an ECL kit (Amersham Biosciences) according to the
manufacturer's instructions.
Immunofluorescence
Paraffin-embedded tissues were sectioned (thickness,
4 µm) using a microtome and placed on glass microscope slides. The
sections were deparaffinized at room temperature by immersion in
two changes of xylene and two changes of absolute ethanol for 10
min each. Subsequently, the sections were rehydrated in a graded
series of ethanol and washed in 10 mM PBS three times for 5 min
each. Antigen retrieval was performed at 95°C for 3 min. After the
sections were cooled to room temperature, they were washed in PBS,
immersed in 5% normal goat serum (Invitrogen; Thermo Fisher
Scientific, Inc.) in PBS and maintained at 37°C for 1 h. Sections
were then incubated with anti-ionized calcium binding adaptor
molecule 1 (IBA1; 1:100; cat. no. sc-32725; Santa Cruz
Biotechnology) and NLRP3 (1:50; cat. no. NBP2-12446; Novus
Biologicals, LLC) or anti-ASC (1:800; cat. no. 67824S; CST) and
anti-pro-caspase-1 (1:100; cat. no. NBP2-15713; Novus Biologicals,
LLC) antibodies at 4°C overnight. The next day, the sections were
washed with PBS and incubated with goat anti-rabbit IgG antibody
(1:1,000; cat. no. 8889S; CST) or goat anti-mouse IgG antibody
(1:1,000; cat. no. 4408S; CST) at 37°C for 1 h, then washed with
PBS. DAPI was added at room temperature for 7 min. Subsequently,
the sections were sealed with neutral mounting medium and observed
with a fluorescence microscope.
Statistical analysis
The data were analyzed using SPSS version 21.0 (IBM
Corp.) and are presented as means ± standard deviation. Differences
between two groups were analyzed using unpaired Student's t-test.
Differences among multiple groups were analyzed using one-way ANOVA
followed by Tukey's post hoc test for pairwise comparisons.
P<0.05 was considered to indicate a statistically significant
difference.
Results
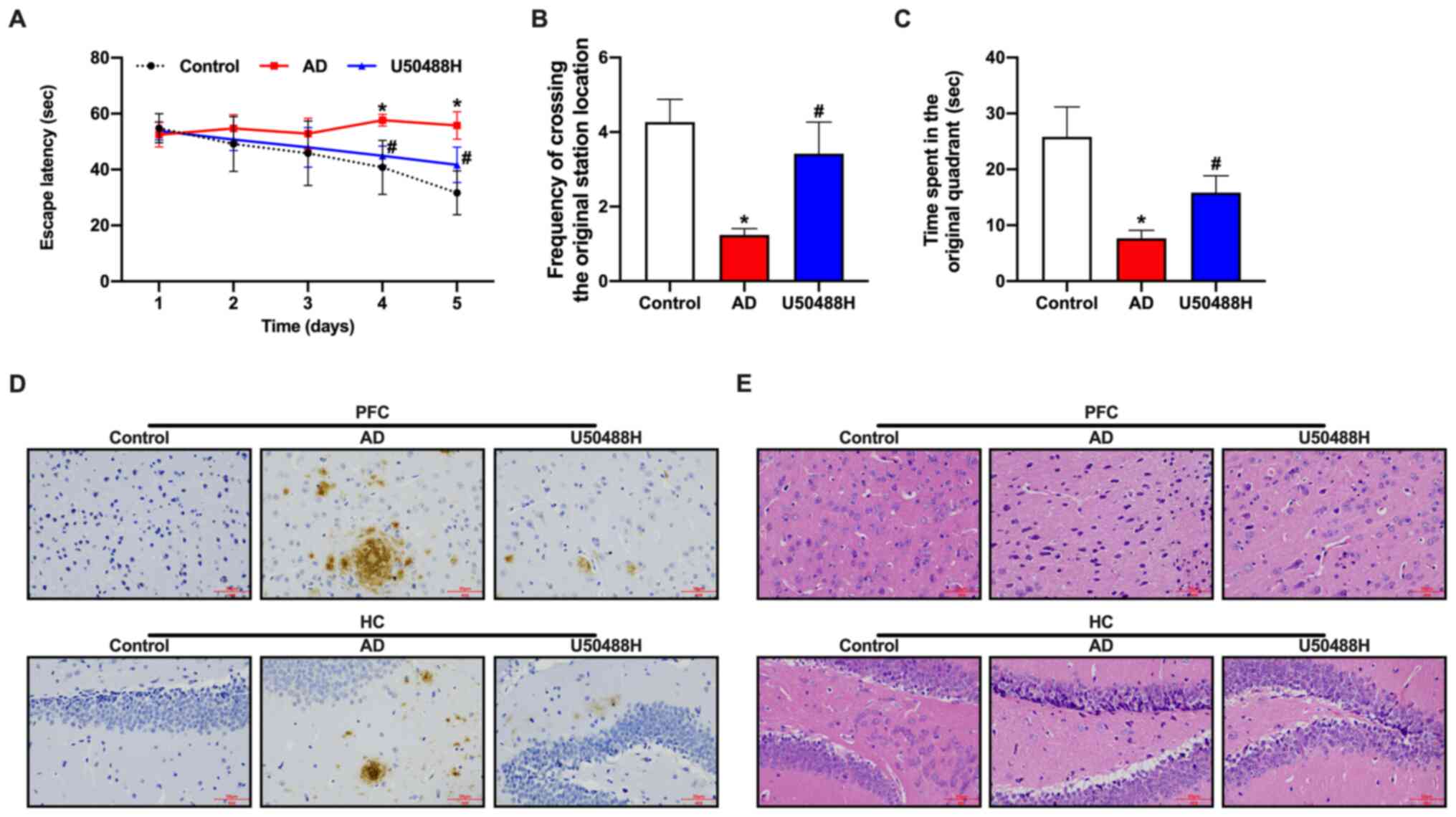
KOR agonist, U50488H, diminishes brain
injury in APP/PS1 mice
To investigate the effect of the KOR agonist,
U50488H, on the spatial memory of APP/PS1 mice, U50488H was
administered subcutaneously for 28 days, using an osmotic pump. The
Morris water maze was used to evaluate the cognitive function of
the mice. Although impaired learning and spatial memory in the
Morris water maze was observed in APP/PS1 mice compared with
control mice, the mice in the U50488H group showed a significant
improvement in their cognitive abilities (Fig. 1A-C). Aβ plaque deposition was
decreased in the prefrontal cortex and HC in the APP/PS1 mice
treated with U50488H (Fig. 1D). The
pathological changes in brain tissue samples were observed using
H&E staining. The neurons in 12 month-old APP/PS1 mice were
sparse and disorganized, with the loss of numerous neurons.
However, the morphology of neurons in the group treated with
U50488H was improved (Fig. 1E).
Furthermore, the expression of PSD95 was increased in the HC of
U50488H-treated mice. Taken together, these results demonstrated
that U50488H reduced brain injury in APP/PS1 mice.
KOR agonist, U50488H inhibits
Ca2+ overload and improves synaptic plasticity in
APP/PS1 mice
Glu is the primary neurotransmitter in the CNS. When
the Glu concentration increases significantly, extensive
pathological damage occurs in brain tissues (29,30).
ELISA analysis revealed that Glu concentrations in the APP/PS1 mice
were significantly higher than in the U50488H treatment group
(Fig. 2A). As the
Ca2+/CaMKII/CREB signaling pathway is involved in
synaptic plasticity, the central components of the pathway,
including NMDAR, CaMKII, p-CaMKII, CREB and p-CREB were analyzed.
Based on the western blot results, the expression levels for NMDARs
and the phosphorylation levels of CaMKII and CREB in the
U50488H-treated group decreased significantly when compared to the
APP/PS1 mice (Fig. 2B). Golgi-Cox
staining was used to analyze the number of dendritic spines in the
HC. The results demonstrated that the number of spines increased in
the treated mice when compared to the APP/PS1 mice, which indicated
that U50488H helped improve the growth of dendritic spines and
improved synaptic plasticity in the treated mice (Fig. 2C). To investigate the underlying
mechanism, western blot analysis was performed to detect the
synaptic plasticity-related proteins, neural cell adhesion
molecule, N-methyl D-aspartate receptor subtype 2B, Glu receptor 1,
PSD95 and α-synuclein. The expression levels for these proteins
were observed to be higher in the U50488H-treated group than in the
APP/PS1 mice (Fig. 2D). These
results demonstrated that treatment with U50488H improved synaptic
plasticity in APP/PS1 mice.
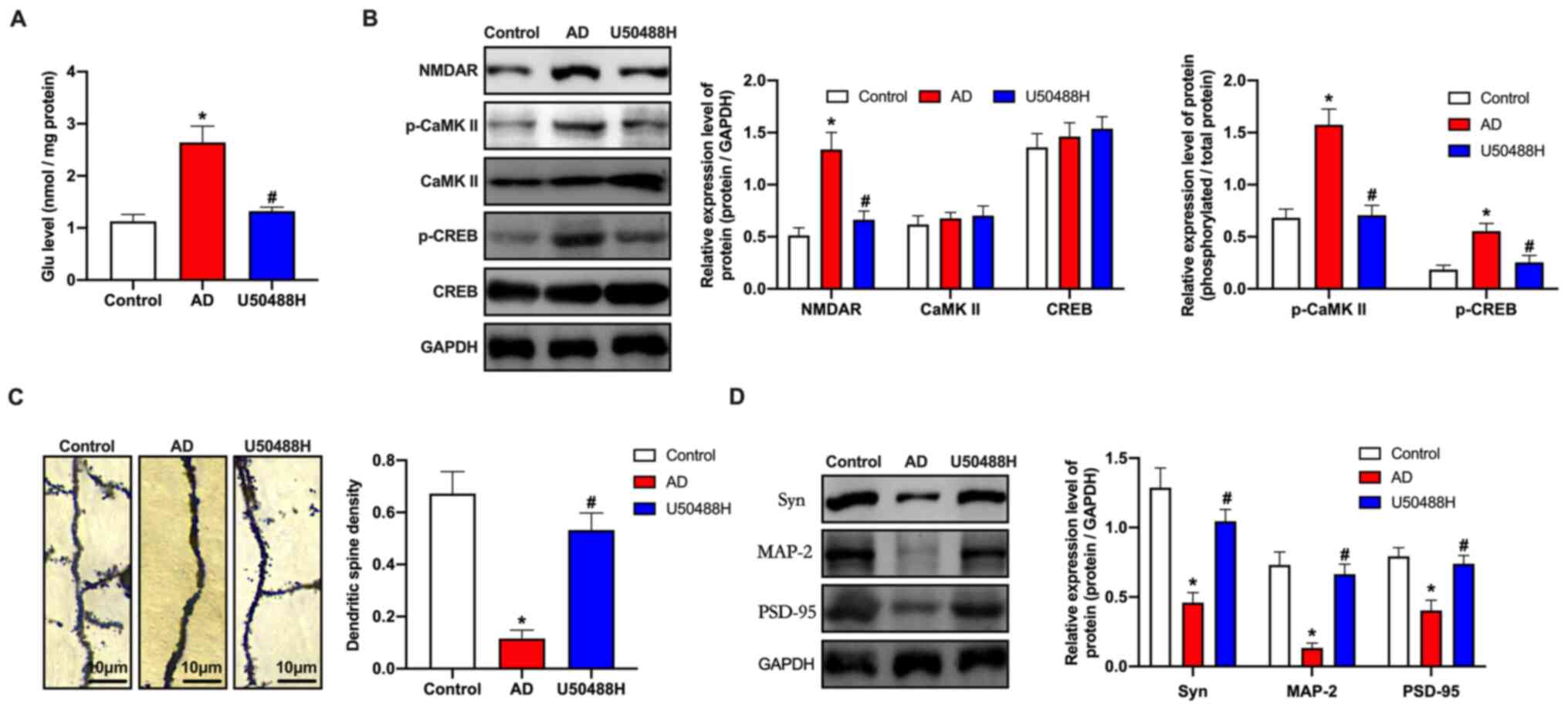 | Figure 2.KOR agonist inhibits Ca2+
overload and improves synaptic plasticity in amyloid precursor
protein/presenilin-1 mice. (A) ELISA analysis demonstrated the Glu
concentration of mice in each group. (B) Western blot analysis of
NMDAR, CaMKII, p-CaMKII, CREB and p-CREB expression levels of mice
in each group. (C) Golgi-Cox staining of brain tissue samples.
Scale bar, 10 µm. (D) Western blot analysis demonstrating NCAM,
NR2B, GluR1, PSD95 and SYN expression levels of mice in each group.
*P<0.05 vs. Control group; #P<0.05 vs. AD group.
Experiments were performed in triplicate and repeated three times.
KOR, κ-opioid receptor; Glu, glutamate; NMDAR, N-methyl-D-aspartate
receptor; CaMKII, calcium/calmodulin-dependent protein kinase II;
p, phosphorylated; CREB, cyclic adenosine monophosphate response
element binding protein; NCAM, neural cell adhesion molecule; NR2B,
N-methyl D-aspartate receptor subtype 2B; GluR1, glutamate receptor
1; PSD95, postsynaptic density protein 95; SYN, α-synuclein; AD,
Alzheimer's disease. |
KOR agonist, U50488H inhibits
microglial pyroptosis in APP/PS1 mice
It has previously been reported that cell pyroptosis
depends on cysteine aspartic acid-specific protease to promote the
release of the inflammatory mediators, IL-1β and IL-18 (31). Notably, the levels of collagen II
antibody, IL-18 and IL-1β in the serum of mice treated with U50488H
decreased significantly (Fig. 3A).
This indicated that U50488H played a critical role in inhibiting
microglial pyroptosis in APP/PS1 mice. Therefore,
immunofluorescence was performed to detect cell pyroptosis-related
proteins. The results showed that the expression levels of NLRP3
and pro-caspase-1 in the U50488H-treated mice decreased
significantly (Fig. 3B and C). To
confirm the fluorescence results, pro-caspase-1, pro-IL-1β,
apoptosis-associated speck-like protein containing a C-terminal
caspase recruitment domain (ASC) and NLRP3 were analyzed using
western blot analysis. The expression levels of ASC, pro-IL-1β,
pro-caspase-1 and NLRP3 in the U50488H-treated group were
significantly lower when compared with those of the APP/PS1 mice
(Fig. 3D). These results indicated
that the KOR agonist inhibited microglial pyroptosis in APP/PS1
mice.
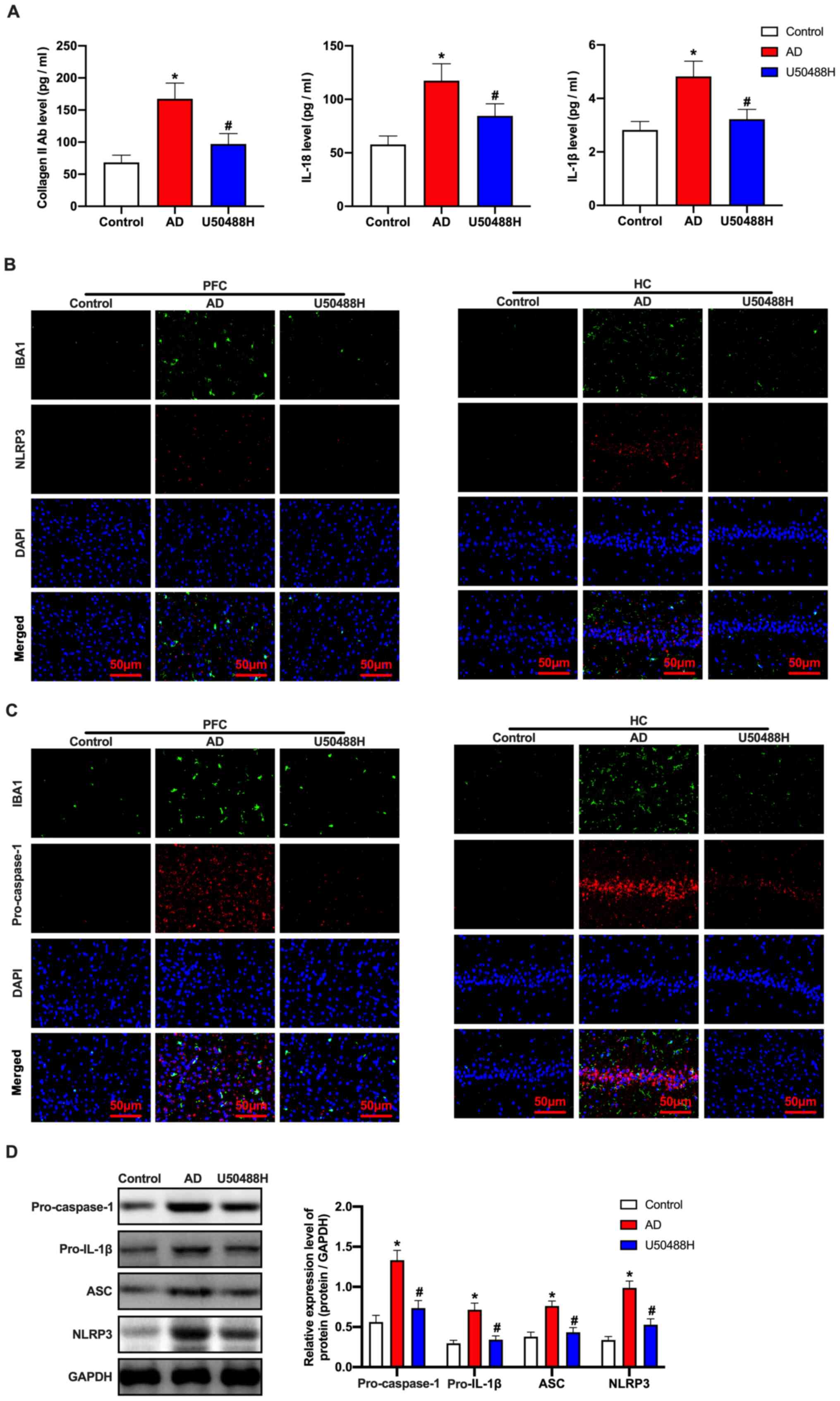 | Figure 3.KOR agonist inhibits microglial
pyrolysis in amyloid precursor protein/presenilin-1 mice. (A) The
level of collagen II antibody, IL-18 and IL-1β were detected by
ELISA. (B) The NLRP3 and IBA1 expression levels and (C)
pro-caspase-1 and IBA1 expression levels of mice in each group were
detected by immunofluorescence. Scale bars, 50 µm. (D) Western blot
analysis of ASC, pro-IL-1β, pro-caspase-1 and NLRP3 expression
levels of mice in each group. *P<0.05 vs. Control group;
#P<0.05 vs. AD group. Experiments were performed in
triplicate and repeated three times. KOR, κ-opioid receptor; Glu,
glutamate; NLRP3, NOD-, LRR- and pyrin domain-containing protein 3;
IBA1, ionized calcium binding adaptor molecule 1; ASC,
apoptosis-associated speck-like protein containing a C-terminal
caspase recruitment domain; AD, Alzheimer's disease. |
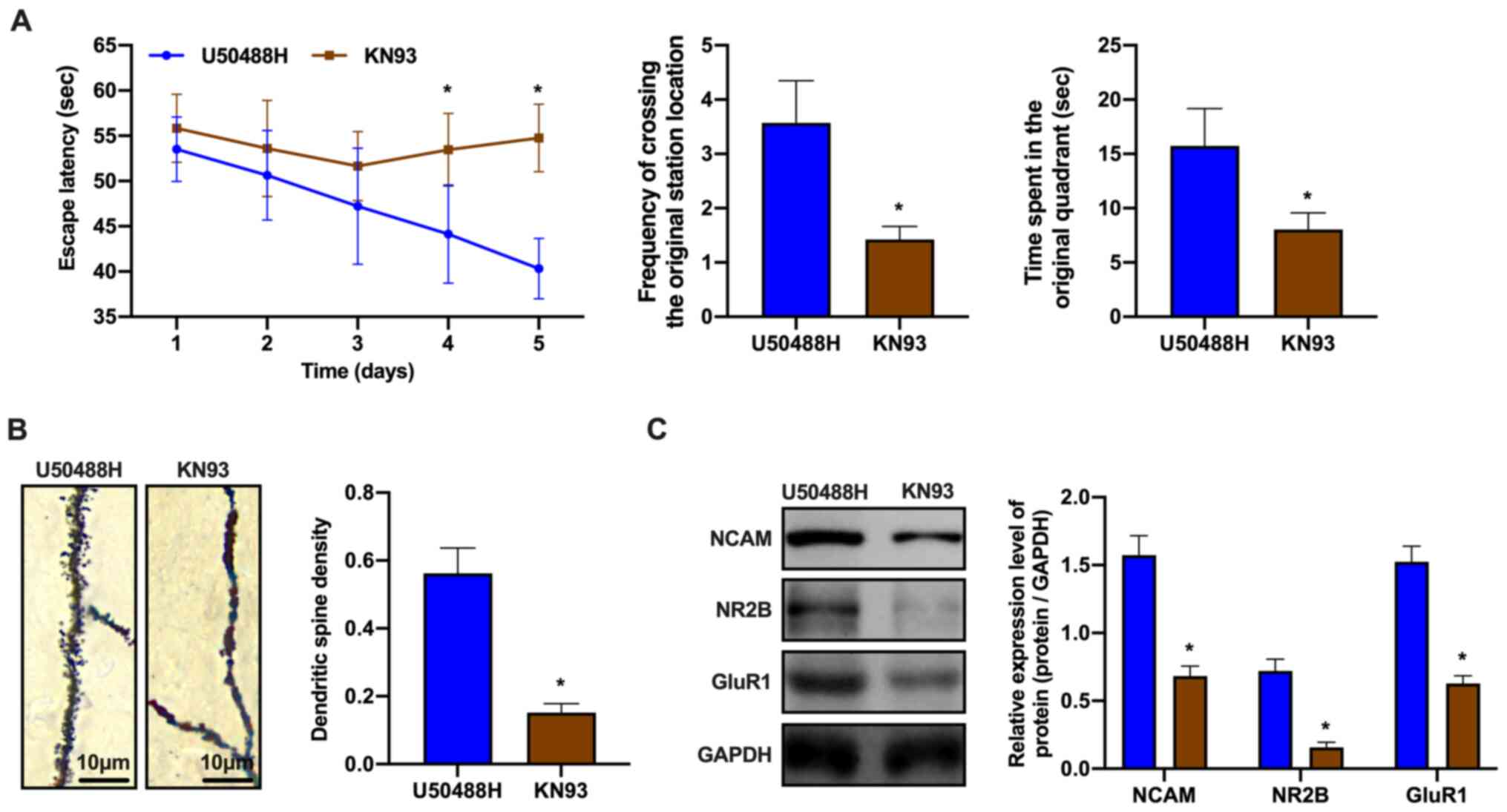
CaMKII inhibitor, KN93 blocks changes
in synaptic plasticity in APP/PS1 mice induced by the KOR
agonist
CaMKII is an important member of the calmodulin
regulatory protein family and plays a role in the
pathophysiological processes of numerous diseases, such as
neuropsychological disorders (32).
Therefore, the regulatory mechanisms of the
Ca2+/CaMKII/CREB signaling pathway in synaptic
plasticity were investigated. KN93 was injected intraperitoneally
into APP/PS1 mice for 28 days. The mice also received U50488H,
which was administered subcutaneously for 28 days using an osmotic
pump. KN93 is a CaMKII specific inhibitor, which inhibits its
phosphorylation activity. Learning and memory was assessed in the
treated and control mice using the Morris water maze. The learning
and memory abilities of the KN93-treated group was observed to
decrease (Fig. 4A). In addition,
the density of the dendritic spines in the CA1 area of the HC was
evaluated using Golgi-Cox staining. The dendritic spine density in
the KN93-treated group was decreased when compared to that of the
untreated group (Fig. 4B).
Furthermore, the mechanisms by which the CaMKII antagonist blocked
changes in synaptic plasticity induced by the KOR agonist were
evaluated. Western blot analysis showed that the expression of
synaptic plasticity-related proteins decreased in the KN93-treated
group (Fig. 4C). These observations
demonstrated that the CaMKII inhibitor blocked the effects of
U50488H, which had improved synaptic plasticity in the APP/PS1
mice.
CaMKII inhibitor eliminates inhibition
by the KOR agonist on microglial pyroptosis in APP/PS1 mice
Based on the abovementioned results, it was
hypothesized that the CaMKII inhibitor blocked inhibition of the
KOR agonist on pyroptosis. The ELISA assay revealed a significant
increase in the expression levels of pyroptosis-related proteins
(Fig. 5A). Treatment with KN93
prevented the decrease in NLRP3 and pro-caspase-1 protein
expression when compared to mice in the U50488H-treated group
(Fig. 5B and C). In addition,
changes in the expression of pyroptosis-related proteins were
examined. The results consistently demonstrated that the expression
levels of NLRP3, ASC, pro-caspase-1 and pro-IL-1β proteins
increased in the KN93-treated mice (Fig. 5D). These observations demonstrated
that the inhibition of pyroptosis in APP/PS1 mice microglia by the
KOR agonist could be eliminated by KN93 treatment.
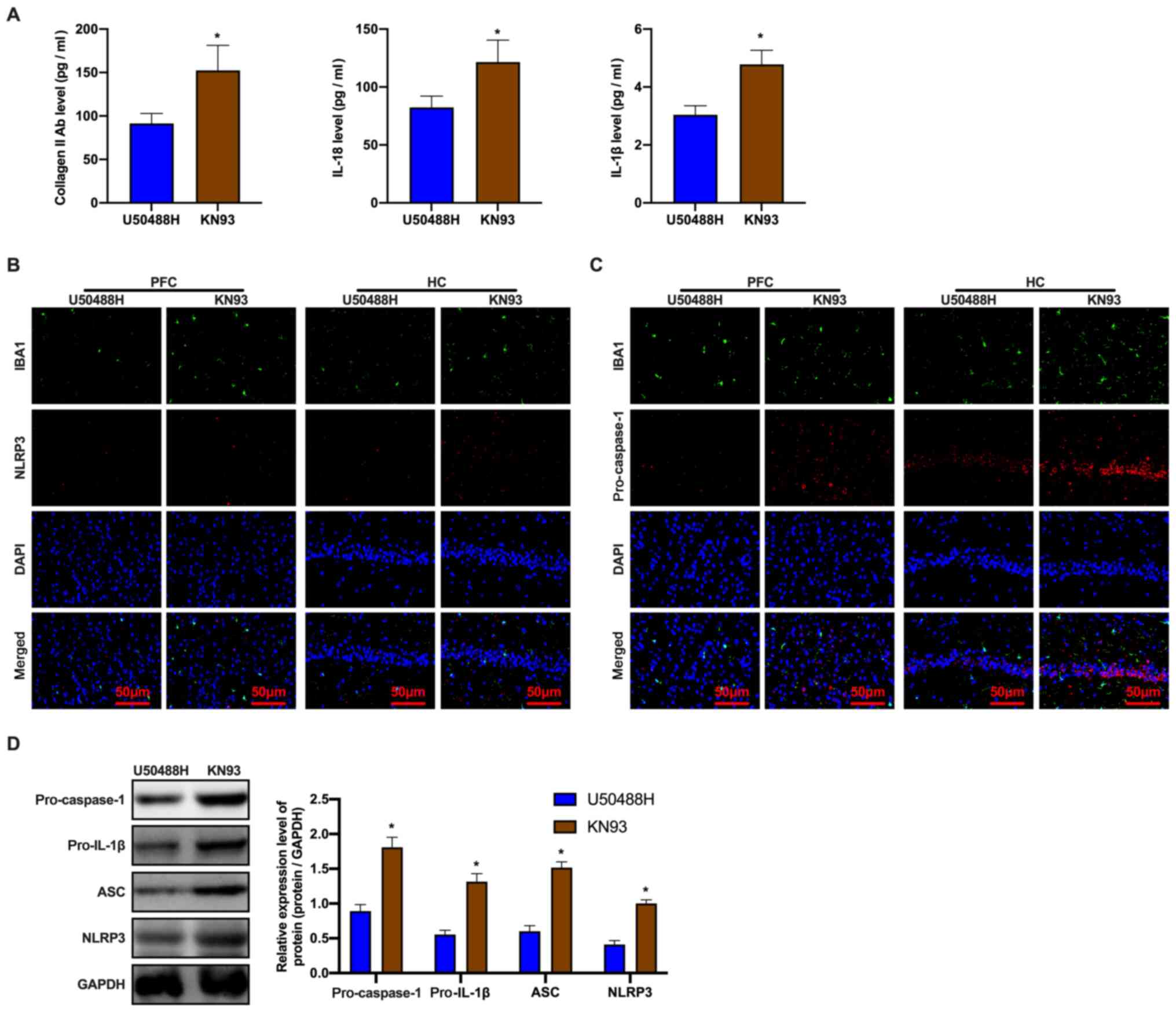 | Figure 5.CaMKII inhibitor, KN93 eliminates the
inhibition of KOR-agonist, U50488H on pyroptosis of amyloid
precursor protein/presenilin-1 mice microglia. (A) The level of
collagen II antibody, IL-18 and IL-1β were detected by ELISA. (B)
The NLRP3 and IBA1 expression levels and (C) The pro-caspase-1 and
IBA1 expression levels of mice in each group detected by
immunofluorescence. Scale bar, 50 µm. (D) Western blot analysis of
ASC, pro-IL-1β, pro-caspase-1 and NLRP3 expression levels of mice
in each group. *P<0.05 vs. U50488H group. Experiments were
performed in triplicate and repeated three times. CaMKII,
calcium/calmodulin-dependent protein kinase II; KOR, κ-opioid
receptor; NLRP3, NOD-, LRR- and pyrin domain-containing protein 3;
IBA1, ionized calcium binding adaptor molecule 1; ASC,
apoptosis-associated speck-like protein containing a C-terminal
caspase recruitment domain; PFC, prefrontal cortex; HC,
hippocampus. |
Discussion
Opioid receptors are widely but unevenly distributed
in the nervous system (33). There
are at least four opioid receptor subtypes in the CNS, µ, κ, δ and
σ. The present study focused on KORs. In addition to the dentate
gyrus, KORs are expressed in the hypothalamus, cerebral cortex and
spinal cord (34). Opioids are
common analgesic treatments in the clinical setting and KOR
agonists exert similar effects. Moreover, compared with traditional
opioids, KOR agonists antagonize the effects mediated by µ-opioid
receptors in the brain, including memory processes. Prior studies
have demonstrated that KOR-specific agonists exhibit
antinociceptive effects and, unlike morphine and other opioid
analgesics, KOR agonists do not result in respiratory depression or
addictive effects (35). However,
KORs agonists in the CNS produce irritable and sedative effects
(36). U50488H is a KOR agonist.
Acetylcholine is blocked through the KORs in the nervous system,
which blocks the decrease in acetylcholine release and ultimately
improves learning and memory (37).
However, the specific mechanism by which KOR agonists promote
recovery from brain injury in AD remains unclear. In the present
study, the Ca2+/CaMKII/CREB signaling pathway was found
to play an essential role in this process.
The Ca2+/CaMKII/CREB signaling pathway is
a critical signal transduction pathway and is involved in the
formation and maintenance of learning and memory in the CNS
(38,39). Ca2+ is a second messenger
that participates in a range of physiological and biochemical
processes in cells. In the process of cell signal transduction, CaM
is the receptor for Ca2+, forming the
Ca2+/CaM complex. As an important target enzyme for
Ca2+, CaMKII phosphorylates numerous substrates to
participate in neuronal plasticity, synthesis, neurotransmitter
release and LTP (40). cAMP
regulatory element modulator can be phosphorylated by CaMKII, which
regulates gene transcription and enhances LTP formation in the HC
(41). The
Ca2+/CaMKII/CREB signaling pathway also regulates
synaptic plasticity, which is consistent with the experimental
results of the present study. The KOR agonist inhibited
Ca2+ overload and improved synaptic plasticity in
APP/PS1 mice in the present study and the CaMKII inhibitor blocked
those changes.
In the present study, it was shown that the
Ca2+/CaMKII/CREB signaling pathway could regulate
microglial pyroptosis. The KOR agonist significantly inhibited the
inflammatory response according to the ELISA analysis. In addition,
the immunofluorescence results that detected cell
pyroptosis-related proteins revealed that the KOR agonist inhibited
microglial pyroptosis in APP/PS1 mice. It has been established that
KOR agonists significantly improved cognitive dysfunction in
cardiopulmonary bypass rats via the JAK2/STAT3 signaling pathway
(42). However, to the best of our
knowledge, it has not previously been reported that KOR agonists
could improve memory impairment in AD. In the present study,
U50488H regulated synaptic plasticity and microglia through the
Ca2+/CaMKII/CREB signaling pathway and the CaMKII
inhibitor reversed this outcome. These observations provide
theoretical evidence that might prove useful for future treatment
of patients with AD.
Thus, the present study demonstrated that KOR
agonists provided neuroprotective effects against AD brain damage
in APP/PS1 mice, which was at least partially mediated by
inhibition of the Ca2+/CaMKII/CREB signaling pathway.
Further investigation is required to assess the possible
associations among other signaling pathways involved in the
underlying mechanisms by which KOR agonists are able to repair the
damage that occurs in the AD brain.
Acknowledgements
Not applicable.
Funding
The present study was supported by Liaoning Province
Key R&D Program Guidance Project (grant no.
2020JH2/10300044).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XSo and XSu designed the project, wrote the
manuscript. JH performed the western blot analysis and PCR. JH and
TY performed the immunohistochemistry and immunofluorescence,
Golgi-Cox staining, ELISA assay and H&E staining. ZC and TY
performed the animal experiment. XSu was the project leader, was
responsible for the design of the project, the revision of the
manuscript and performed some of the experiments. ZC and XSo are
responsible for confirming the authenticity of the raw data. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The animal study was approved by the Institutional
Animal Care and Use Committees of the China Medical University
Laboratory.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Soria Lopez JA, Gonzalez HM and Leger GC:
Alzheimer's disease. Handb Clin Neurol. 167:231–255. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tiwari S, Atluri V, Kaushik A, Yndart A
and Nair M: Alzheimer's disease: Pathogenesis, diagnostics, and
therapeutics. Int J Nanomedicine. 14:5541–5554. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferreira-Vieira TH, Guimaraes IM, Silva FR
and Ribeiro FM: Alzheimer's disease: Targeting the Cholinergic
System. Curr Neuropharmacol. 14:101–115. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Parsons MP and Raymond LA: Extrasynaptic
NMDA receptor involvement in central nervous system disorders.
Neuron. 82:279–293. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang R and Reddy PH: Role of glutamate and
NMDA receptors in Alzheimer's disease. J Alzheimers Dis.
57:1041–1048. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sanhueza M and Lisman J: The CaMKII/NMDAR
complex as a molecular memory. Mol Brain. 6:102013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang D, Noda Y, Zhou Y, Nitta A, Nabeshima
T and Yu Q: Effects of sodium houttuyfonate on phosphorylation of
CaMK II, CREB and ERK 1/2 and expression of c-Fos in macrophages.
Int Immunopharmacol. 4:1083–1088. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baek A, Park EJ, Kim SY, Nam BG, Kim JH,
Jun SW, Kim SH and Cho SR: High-Frequency repetitive magnetic
stimulation enhances the expression of brain-derived neurotrophic
factor through activation of Ca2+-calmodulin-dependent
protein kinase II-cAMP-response element-binding protein pathway.
Front Neurol. 9:2852018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang Z, Cao X, Bao X, Zhang Y, Xu Y and
Sha D: Cocaine- and amphetamine-regulated transcript protects
synaptic structures in neurons after ischemic cerebral injury.
Neuropeptides. 81:1020232020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Islam R, Matsuzaki K, Sumiyoshi E, Hossain
ME, Hashimoto M, Katakura M, Sugimoto N and Shido O: Theobromine
improves working memory by activating the CaMKII/CREB/BDNF pathway
in rats. Nutrients. 11:8882019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ko HR, Ahn SY, Chang YS, Hwang I, Yun T,
Sung DK, Sung SI, Park WS and Ahn JY: Human UCB-MSCs treatment upon
intraventricular hemorrhage contributes to attenuate hippocampal
neuron loss and circuit damage through BDNF-CREB signaling. Stem
Cell Res Ther. 9:3262018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Moir RD, Lathe R and Tanzi RE: The
antimicrobial protection hypothesis of Alzheimer's disease.
Alzheimers Dement. 14:1602–1614. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rathinam VA and Fitzgerald KA:
Inflammasome complexes: Emerging mechanisms and effector functions.
Cell. 165:792–800. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Man SM and Kanneganti TD: Regulation of
inflammasome activation. Immunol Rev. 265:6–21. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Olsen I and Singhrao SK: Inflammasome
involvement in Alzheimer's disease. J Alzheimers Dis. 54:45–53.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao N, Sun C, Zheng M, Liu S and Shi R:
Amentoflavone suppresses amyloid β1–42 neurotoxicity in Alzheimer's
disease through the inhibition of pyroptosis. Life Sci.
239:1170432019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
La Rosa F, Saresella M, Marventano I,
Piancone F, Ripamonti E, Al-Daghri N, Bazzini C, Zoia CP, Conti E,
Ferrarese C and Clerici M: Stavudine reduces NLRP3 inflammasome
activation and modulates amyloid-β autophagy. J Alzheimers Dis.
72:401–412. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang S, Yuan YH, Chen NH and Wang HB: The
mechanisms of NLRP3 inflammasome/pyroptosis activation and their
role in Parkinson's disease. Int Immunopharmacol. 67:458–464. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Qiu Z, Lei S, Zhao B, Wu Y, Su W, Liu M,
Meng Q, Zhou B, Leng Y and Xia ZY: NLRP3 inflammasome
activation-mediated pyroptosis aggravates myocardial
ischemia/reperfusion injury in diabetic rats. Oxid Med Cell Longev.
2017:97432802017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hickman S, Izzy S, Sen P, Morsett L and El
Khoury J: Microglia in neurodegeneration. Nat Neurosci.
21:1359–1369. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hansen DV, Hanson JE and Sheng M:
Microglia in Alzheimer's disease. J Cell Biol. 217:459–472. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Orihuela R, McPherson CA and Harry GJ:
Microglial M1/M2 polarization and metabolic states. Br J Pharmacol.
173:649–665. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sominsky L, De Luca S and Spencer SJ:
Microglia: Key players in neurodevelopment and neuronal plasticity.
Int J Biochem Cell Biol. 94:56–60. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu Y, Dissing-Olesen L, MacVicar BA and
Stevens B: Microglia: Dynamic mediators of synapse development and
plasticity. Trends Immunol. 36:605–613. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Corder G, Castro DC, Bruchas MR and
Scherrer G: Endogenous and exogenous opioids in pain. Annu Rev
Neurosci. 41:453–473. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Takahashi K, Nakagawasai O, Sugawara M,
Sato A, Nemoto W, Tadano T and Tan-No K: Kappa opioid receptor
agonist administration in olfactory bulbectomized mice restores
cognitive impairment through cholinergic neuron activation. Biol
Pharm Bull. 41:957–960. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
D'Hooge R and De Deyn PP: Applications of
the Morris water maze in the study of learning and memory. Brain
Res Brain Res Rev. 36:60–90. 2001. View Article : Google Scholar
|
|
28
|
Garthe A and Kempermann G: An old test for
new neurons: Refining the Morris water maze to study the functional
relevance of adult hippocampal neurogenesis. Front Neurosci.
7:632013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Meldrum BS: Glutamate as a
neurotransmitter in the brain: Review of physiology and pathology.
J Nutr. 130 (Suppl 4S):1007S–1015S. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Colombo MN and Francolini M: Glutamate at
the vertebrate neuromuscular junction: From modulation to
neurotransmission. Cells. 8:9962019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Man SM, Karki R and Kanneganti TD:
Molecular mechanisms and functions of pyroptosis, inflammatory
caspases and inflammasomes in infectious diseases. Immunol Rev.
277:61–75. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Takemoto-Kimura S, Suzuki K, Horigane SI,
Kamijo S, Inoue M, Sakamoto M, Fujii H and Bito H: Calmodulin
kinases: Essential regulators in health and disease. J Neurochem.
141:808–818. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Waldhoer M, Bartlett SE and Whistler JL:
Opioid receptors. Annu Rev Biochem. 73:953–990. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ding YQ, Kaneko T, Nomura S and Mizuno N:
Immunohistochemical localization of mu-opioid receptors in the
central nervous system of the rat. J Comp Neurol. 367:375–402.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Baumann MH, Majumdar S, Le Rouzic V,
Hunkele A, Uprety R, Huang XP, Xu J, Roth BL, Pan YX and Pasternak
GW: Pharmacological characterization of novel synthetic opioids
(NSO) found in the recreational drug marketplace.
Neuropharmacology. 134((Pt A)): 101–107. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Beck TC, Hapstack MA, Beck KR and Dix TA:
Therapeutic potential of kappa opioid agonists. Pharmaceuticals
(Basel). 12:952019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang Q, Sun Y, Li J, Xing W, Zhang S, Gu
X, Feng N, Zhao L, Fan R, Wang Y, et al: Quaternary ammonium salt
of U50488H, a new κ-opioid receptor agonist, protects rat heart
against ischemia/reperfusion injury. Eur J Pharmacol. 737:177–184.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ding S, Xu Z, Yang J, Liu L, Huang X, Wang
X and Zhuge Q: The involvement of the decrease of astrocytic Wnt5a
in the cognitive decline in minimal hepatic encephalopathy. Mol
Neurobiol. 54:7949–7963. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang L, Chen ZW, Yang SF, Shaer M, Wang
Y, Dong JJ and Jiapaer B: MicroRNA-219 decreases hippocampal
long-term potentiation inhibition and hippocampal neuronal cell
apoptosis in type 2 diabetes mellitus mice by suppressing the NMDAR
signaling pathway. CNS Neurosci Ther. 25:69–77. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shonesy BC, Jalan-Sakrikar N, Cavener VS
and Colbran RJ: CaMKII: A molecular substrate for synaptic
plasticity and memory. Prog Mol Biol Transl Sci. 122:61–87. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ahmed T and Frey JU: Plasticity-specific
phosphorylation of CaMKII, MAP-kinases and CREB during late-LTP in
rat hippocampal slices in vitro. Neuropharmacology. 49:477–492.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li X, Sun Y, Jin Q, Song D and Diao Y:
Kappa opioid receptor agonists improve postoperative cognitive
dysfunction in rats via the JAK2/STAT3 signaling pathway. Int J Mol
Med. 44:1866–1876. 2019.PubMed/NCBI
|