Introduction
Diabetic retinopathy (DR) is one of the most common
complications of diabetes and the leading cause of vision loss
globally (1,2). It is estimated that diabetes will
affect ~642 million people worldwide by 2035 (3) and nearly all patients with type 1, and
more than half with type 2 diabetes, will develop DR (1). DR pathophysiology is complex and
involves several mechanisms, such as oxidative stress,
mitochondrial dysfunction, an increase in inflammatory mediators
and metabolic abnormalities initiated by hyperglycaemia (4–6).
Apoptosis of retinal neurons serves a critical role in the
pathogenesis of DR observed in patients with diabetes and
streptozotocin (STZ)-induced diabetic rats (5,7–9).
Retinal neuron apoptosis occurs one month after STZ injection,
which is considered the early stage of DR (6,10). The
molecular mechanisms involved in the suppression of retinal neuron
apoptosis during the early stage of DR remain unclear.
RNA-dependent protein kinase (PKR) is a
serine/threonine-protein kinase implicated in the modulation of the
stress response and pro-inflammatory pathways, serving a role in
the regulation of critical cell processes (11), including the apoptosis of retinal
ganglion cells (RGCs) and neurons in patients with Parkinson's,
Alzheimer's and Huntington's disease (12–16).
PKR is ubiquitously expressed in mammalian cells and was initially
identified by its activation by double-stranded RNA (dsRNA)
intermediates during viral infection via a mechanism involving
autophosphorylation (11,17). PKR serves an essential role in cell
antiviral defence; studies on its activation have been performed
primarily with viral and synthetic dsRNA (18–20).
Single-stranded RNAs containing secondary structure motifs activate
PKR (14,19). Mitochondrial RNA (mtRNA) is present
as dsRNA in the cytosol where it interacts with PKR, resulting in
PKR autophosphorylation, especially under stress (21).
Notably, the activation of PKR is stress responsive
and its induction results in inhibition of cell proliferation,
suppression of translation and induction of apoptosis (15,22,23).
PKR serves a role in endoplasmic reticulum (ER) stress-dependent
apoptosis via the eukaryotic translation initiation factor 2α
(eIF2-α)/activating transcription factor 4/CHOP signalling pathway
(15) and ER stress has a role in
the early stage of DR (24).
PKR-associated protein X (RAX), a direct
activator of PKR, induces apoptosis by activating the PKR
signalling pathway under stress (25). Our previous study (26) showed that RAX expression is
indirectly modulated by microRNA (miR-29b) at the early stage of
STZ-induced diabetic rats. This finding suggests that miR-29b
serves a protective effect against apoptosis of retinal neurons via
the PKR signalling pathway. The present study investigated the
potential mechanisms underlying protection of retinal neurons
against apoptosis by PKR in the early stage of DR. The present
study investigated the subcellular localization, expression levels
and activity of PKR in the retina of normal and STZ-induced
diabetic rats.
Materials and methods
Animals and STZ treatment
A total of 168 Wistar rats (male; weight, 130–150 g;
age, 5–6 weeks) were obtained from the Animal Center of School of
Medicine, University of São Paulo, São Paulo Brazil. The animals
were housed in suspended wire-bottom cages under environmentally
controlled temperature (25±2°C) and humidity (55±5%) in a 12/12 h
dark/light cycle. The animals were provided food and water ad
libitum. Animals were maintained under these conditions for ≥4
days prior to the experiments. The animals were randomly divided
into seven groups (n=8 rats/group): Control group and six
experimental groups, in which animals were killed 3, 6, 15, 22, 28
and 35 days after STZ injection. Experimental diabetes mellitus was
induced by single intravenous injection of STZ (Sigma-Aldrich;
Merck KGaA) dissolved in 0.01 M citrate buffer (pH, 4.5) within 5
min of preparation. The rats were fasted overnight, anesthetized
with 5% isoflurane until loss of righting reflex and were
immediately injected with STZ in the jugular vein at a dose of 45
mg/kg body weight. Control rats received only citrate buffer.
At 24 h after STZ administration, blood glucose
levels were measured via the colorimetric oxidase glucose method
(Labtest) and animals with a blood glucose value ≥400 mg/dl were
used in subsequent experiments (27). All experiments were performed
between 8:00 and 10:00 a.m. STZ-injected and control rats were
sacrificed by rapid cervical dislocation. The death of animals was
confirmed by observing cardiac and respiratory arrest for 3–5 min
before retinas were dissected and used for analysis of PKR and
p-eIF2-α expression levels or immunofluorescence. The care and
treatment of the animals received prior institutional approval from
the Ethical Commission on Animal Research of the School of Medicine
at the University of São Paulo.
TUNEL apoptosis assay
Apoptotic cells in the retina were investigated
using a DeadEnd™ Colorimetric TUNEL system (Promega Corporation)
according to the manufacturer's protocol. Briefly, slides were
fixed in 4% formaldehyde for 15 min at room temperature and then
washed with PBS before permeabilization with protein k (10 ng/ml
for 15 min). The slides were rewashed in PBS, fixed in 4%
paraformaldehyde for 15 min at room temperature and immersed in
PBS. Both test and negative control slides were covered with buffer
solution (Promega Corporation) while the positive control reaction
was performed. For the positive control, DNase I and its respective
buffer (Promega Corporation) were added for 10 min at 37°C in a
humid chamber. The negative control was performed using RNase-free
water instead of DNase. The slides were then washed in 2X
saline-sodium citrate buffer diluted in deionized water (1:10),
incubated for 5 min at room temperature in 0.3% hydrogen peroxide,
diluted in PBS and washed twice with PBS (5 min each). Horseradish
peroxidase (HRP)-labelled streptavidin solution was prepared at a
1:500 dilution in PBS. Each slide received 100 µl streptavidin-HRP
and was incubated for 5 min at room temperature followed by washing
in PBS. Finally, the slides were incubated in 1X DAB solution (DAB
10X chromogen in DAB substrate 1X Buffer) for 10 min in a humid
dark chamber at room temperature. The slides were dehydrated,
mounted in Permount (Thermo Fisher Scientific, Inc.) and examined
under a light microscope (Leica DFC 340 FX; Leica Microsystems
GmbH) at an original magnification of ×100.
Immunofluorescence
Enucleated eyes were fixed using 4% buffered
paraformaldehyde dissolved in 0.2 M phosphate buffer (pH 7.3) for
24 h at 4°C, then treated with 70% ethanol (2 h), 99% ethanol (1
h), absolute ethanol (30 min) and xylene (1 h) at room temperature,
followed by treatment with xylene for 10 min at 37°C before being
placed in a paraffin bath (1 h; 56°C). The fixed samples were
subjected to vacuum pressure for 30 min and then embedded in
paraffin (2 baths at 60°C, 2 h each). The paraffin blocks were cut
into 5-µm serial sections and placed on Superfrost™ (Cole-Parmer
Ltd.; Thermo Fisher Scientific, Inc.) slides. In order to confirm
cellular integrity, the samples were stained using Harris
hematoxylin and eosin-phloxine. Sections were adhered to the slides
in an oven at 80°C for 15 min, deparaffinized in xylene and
rehydrated via graded ethanol washes (100, 95, 80, and 70%) for 10
min each, followed by washing in distilled water for 5 min.
Sections were stained at room temperature with 5% w/v hematoxylin
in water for 1 min, washed in distilled water and stained with 0.5%
w/v eosin in water for 2 min. Samples were blocked at room
temperature in 1% goat serum, 2% BSA and 0.05% Triton X-100 in PBS
for 1 h, then incubated with anti-PKR mouse monoclonal (1:50; cat.
no. sc-6282; Santa Cruz Biotechnology, Inc.) or anti-p-eIF2-α
rabbit polyclonal antibodies (1:50; cat. no. 44-728G; Thermo Fisher
Scientific, Inc.) for 3 h at 4°C. The samples were washed for 15
min in PBS and incubated with species-specific fluorescent Alexa
Fluor 594 rabbit anti-mouse or goat anti-rabbit (H+L) (both
1:2,000; cat. nos. A-11062 and A-11037, respectively; both Thermo
Fisher Scientific, Inc.) in a humidified chamber at room
temperature for 1 h in the dark. Then, slides were mounted in
Vectashield Mounting Medium with DAPI (Sigma-Aldrich; Merck KGaA)
and examined under a microscope (magnification, ×100; Leica DFC 340
FX). Images were analyzed using Adobe PhotoShop CS6 version 13.0×
64 software (Adobe Systems, Inc.). The specificity of antibody
staining was confirmed by incubating adjacent sections in the
absence of the primary antibody.
Western blot analysis
Total protein was extracted from rat retina using
lysis buffer [20 mM Tris-HCl (pH 7.6), 400 mM NaCl, 50 mM KCl, 0.2
mM phenylmethylsulfonyl fluoride, 2 µg/ml leupeptin, 2 µg/ml
aprotinin, 1 mM dithiothreitol, 1 mM EDTA, 1% Triton X-100 and 20%
glycerol]. All reagents were purchased from Sigma-Aldrich (Merck
KGaA). The homogenates were centrifuged at 15,500 × g at 4°C for 40
min and the supernatant was used for analysis. The protein
concentrations were determined using Pierce BCA Protein Assay
(Thermo Fisher Scientific, Inc.). Equal amounts of protein (30 µg)
were separated by electrophoresis using a 10% SDS-PAGE resolving
gel with 4% SDS-PAGE stacking gel and transferred onto a
Hybond-C-supported nitrocellulose membrane (Amersham Biosciences;
Cytiva). The membranes were subsequently blocked with 3% BSA in 100
mM Tris-HCl (pH 7.5), 100 mM NaCl, 0.1% Tween-20 (TBST) for 1 h at
4°C with gentle agitation. After washing in TBST for 20 min, the
membranes were incubated overnight with anti-PKR (1:300; cat. no.
sc-6282; Santa Cruz Biotechnology, Inc.) and anti-β-actin mouse
monoclonal antibodies at 4°C (1:1,000; cat. no. sc-517582; Santa
Cruz Biotechnology, Inc.). The membranes were subsequently washed
with TBST for 20 min and incubated with HRP-conjugated anti-mouse
IgG κ binding protein (1:5,000; cat. no. sc-516102; Santa Cruz
Biotechnology, Inc.) antibodies diluted in TBST at room temperature
for 60 min. Membranes were washed for 20 min with TBST.
Antibody-labeled protein bands were visualized with enhanced
chemiluminescence detection reagents (Amersham Biosciences; Cytiva)
according to the manufacturer's protocol. The films were observed
and the intensity of protein bands was determined using ImageJ
v1.4.1 software (National Institutes of Health).
Statistical analysis
Data are presented as the mean ± SD (n=3) and were
analyzed by one-way ANOVA followed by Dunnett's post hoc test.
Statistical analysis was performed using GraphPad Prism software
(version 4.0; GraphPad Software, Inc.). P<0.05 was considered to
indicate a statistically significant difference.
Results
TUNEL apoptosis assays
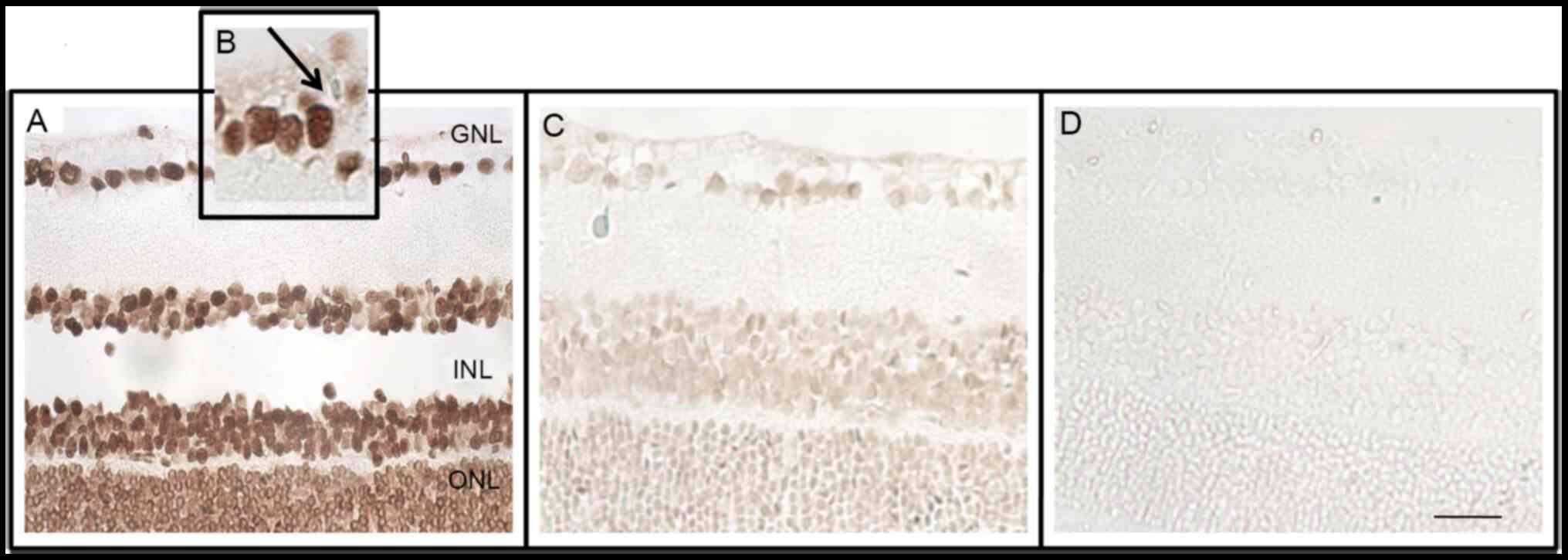
The in situ presence of apoptotic cells in
the sections of control and treated rat retina following STZ
injection on days 6, 15, 22, and 35 was investigated by the TUNEL
method. The morphological patterns obtained in the positive
control, such as condensed and fragmented nuclei and the presence
of apoptotic bodies, were compared with retinal cells of the
diabetic and negative groups (Fig. 1A
and B). At 35 days after treatment with STZ (Fig. 1C), there was no increase in the
number of apoptotic cells, indicating that the present study
evaluated molecular events that preceded apoptosis of neurons in
the retina of diabetic rats. The negative control is shown in
Fig. 1D.
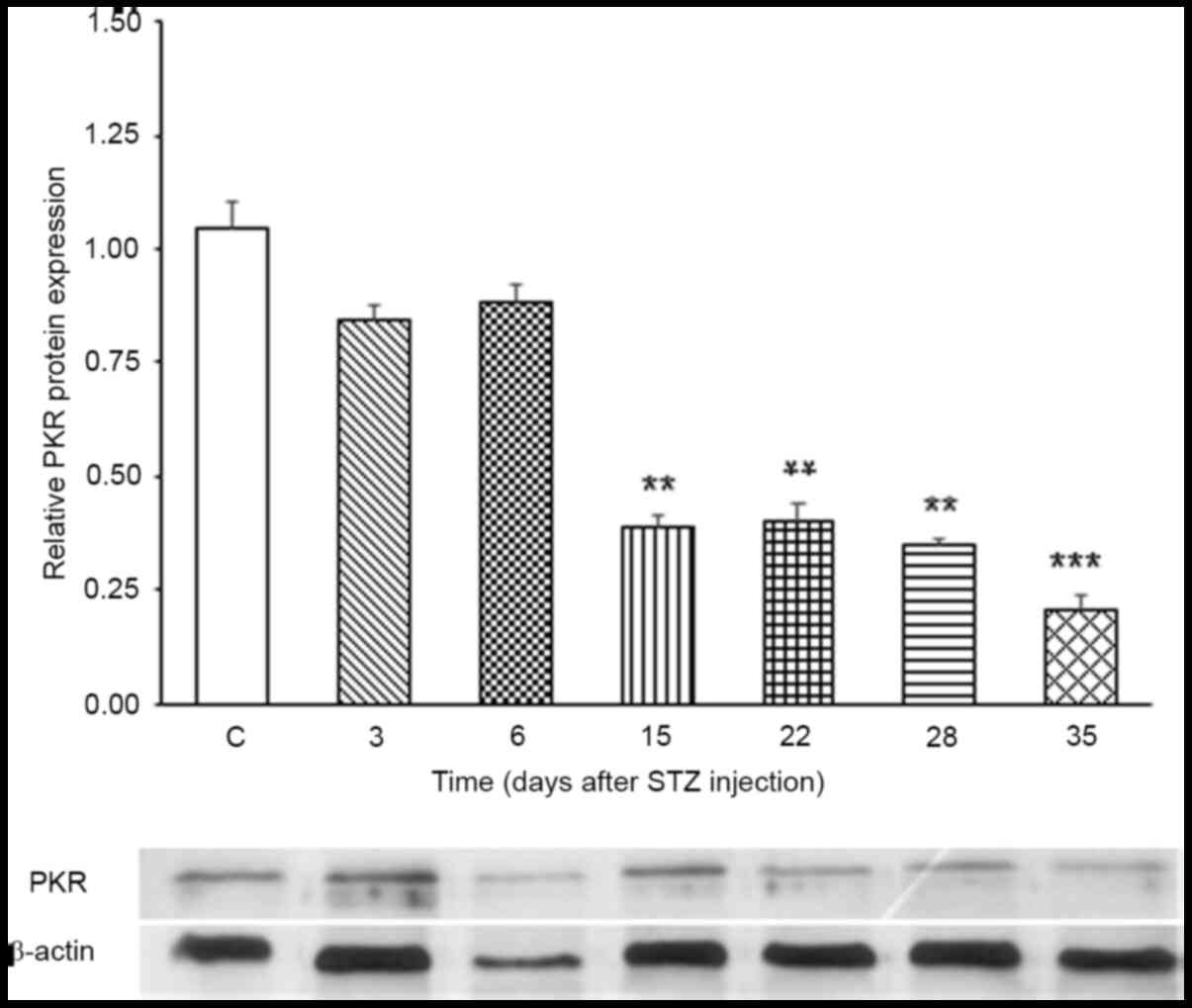
Expression levels of PKR protein
Western blot analysis showed that PKR protein
expression levels did not change on days 3 and 6 after STZ
treatment compared with the control (Fig. 2). However, on days 15, 22, 28 and
35, a significant decrease in PKR protein expression levels was
observed.
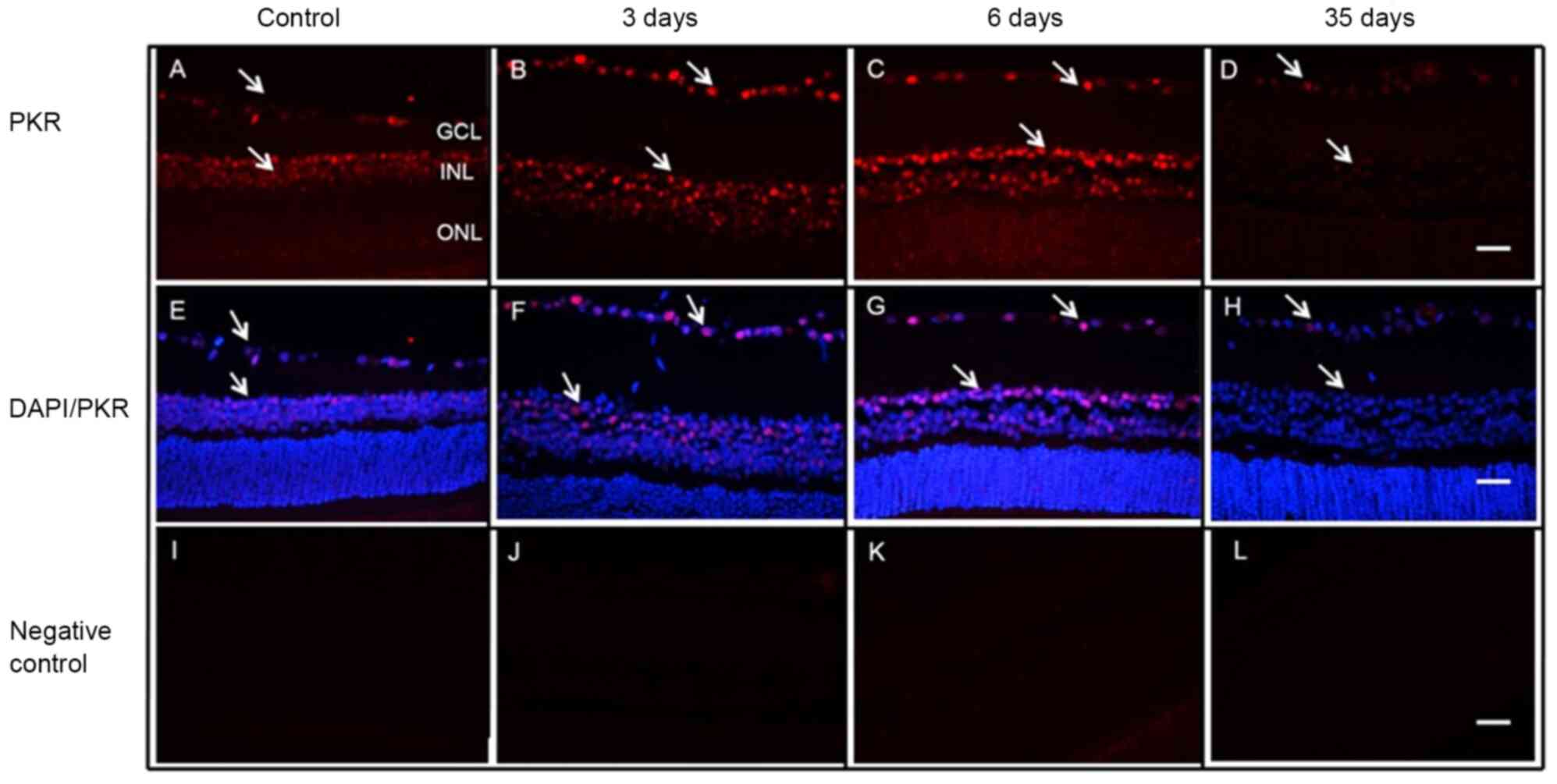
Cellular localization of PKR
Cellular localization of PKR was investigated in the
retina of normal and diabetic rats on days 3, 6 and 35 after STZ
injection (Fig. 3). PKR staining
was observed in the nuclei of RGCs and INL cells. No change was
observed in the cellular localization of PKR in the retinas of
normal or diabetic rats. However, expression levels of nuclear PKR
in the retinas of diabetic rats were more evident on days 3 and 6
(Fig. 3B and C) compared with
control animals (Fig. 3A). Minimal
expression was detected at 35 days after STZ injection (Fig. 3D). The DAPI-stained nuclei are shown
in Fig. 3E-H and the negative
controls in Fig. 3I-L. PKR protein
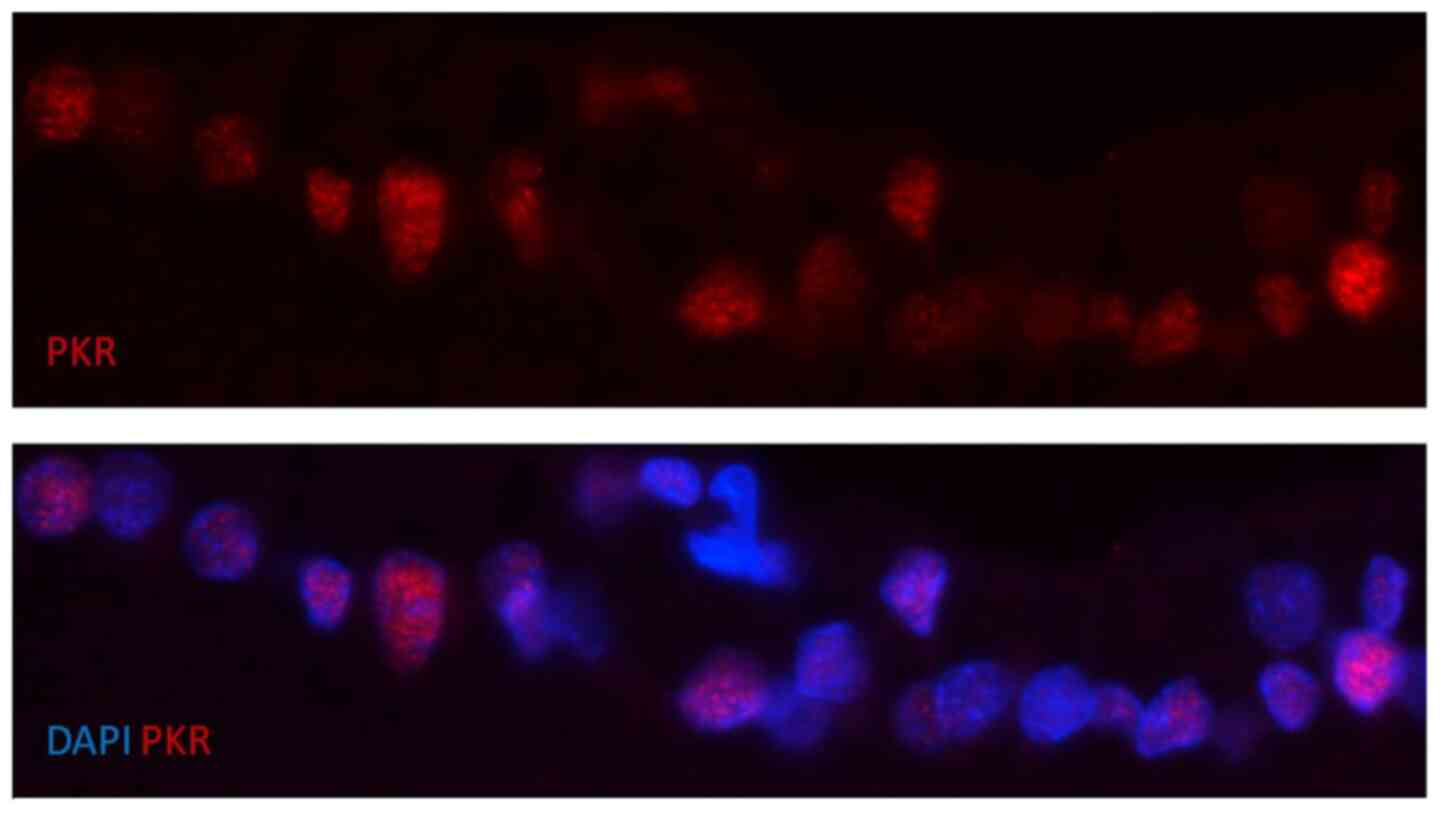
nuclear staining on day 6 after STZ injection is presented in
Fig. 4.
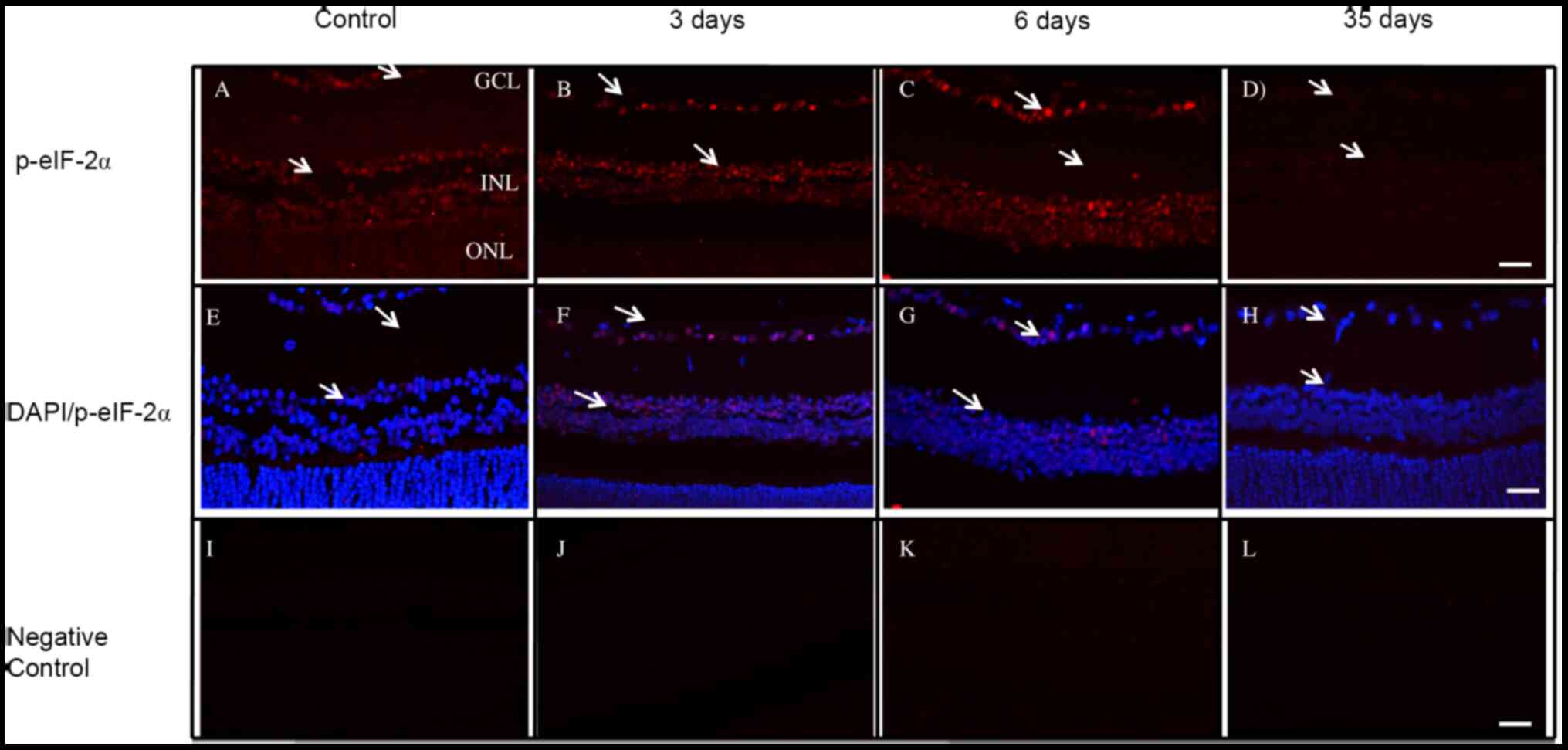
Cellular localization of p-eIF2-α
Activated eIF2-α protein (p-eIF2-α) was expressed in
the same retinal neurons where PKR was detected from the retina
observed for PKR (Fig. 5). In
addition, the number of p-eIF2-α-positive retinal cells was the
same in the normal and diabetic groups (Fig. 5A-C). Nuclear expression of p-eIF2-α
in RGCs and the inner nuclear layer (INL) was observed at 3 and 6
days. Moreover, at 35 days after STZ injection, p-eIF2-α expression
levels were decreased in the retina of treated rats compared with
controls (Fig. 5A and D). The
DAPI-stained nuclei are shown in Fig.
5E-H and the negative controls in Fig. 5I-L.
Discussion
The present study evaluated the subcellular
localization, expression levels and activity of the pro-apoptotic
kinase PKR in retinal neurons during the first 35 days after STZ
injection, which is considered the early stage of DR in this
experimental model of type 1 diabetes (28). The first step was to investigate PKR
expression levels and subcellular localization in the retina of
control and STZ-treated rats. PKR protein expression levels were
downregulated at 15, 22, and 28 days and PKR was localized in the
nuclei of RGCs and INL cells in the rat retina. To the best of our
knowledge, this is the first demonstration of the nuclear
localization of PKR in retinal neurons. The activation of PKR was
indirectly assessed by eIF2-α phosphorylation, as previously
reported (29). A strong signal of
p-eIF2-α-positive cells was observed up to 22 days after STZ
treatment compared with normal retinas. In addition, analysis of
cellular localization of p-eIF2-α showed that this protein was
strongly expressed in the nucleus of neurons of the GCL in the
retina of diabetic animals, suggesting that nuclear PKR was active
in retinal neurons of diabetic rats.
PKR is stress-sensitive and considered to be one of
the most important pro-apoptotic kinases (15,22,23).
Thus, cellular stress due to glucotoxicity in the early stage of DR
induced by STZ may activate PKR and consequently induce apoptosis
of retinal neurons. However, the present study did not detect
apoptosis in retinal neurons in spite of the presence of active PKR
in these cells.
The accumulation of active PKR in the nucleus has
been proposed as a cellular stress response (30,31).
It has been linked to several types of pathology, such as leukaemia
development (30), radiation
resistance in lung cancer (32),
sporadic Alzheimer's disease (33),
Creutzfeldt-Jakob disease (34) and
myelodysplastic syndrome (31). The
accumulation of active PKR has also been reported in the nuclei of
cells treated with tunicamycin, an ER stress inductor, and in brain
neurons of patients with Parkinson's and Huntington's disease
(12,35). The biological significance of PKR
translocation to the nucleus is not entirely understood. Blalock
et al (30) reported that
PKR is phosphorylated in patients with myelodysplastic syndrome and
its subcellular localization depends on disease severity. Their
findings suggested that nuclear translocation of PKR may be a
mechanism to sequester active PKR, thus preventing this stress
kinase from activating signalling pathways in the cytosol.
Previous work has revealed that cellular stress due
to hyperglycaemia is responsible for mitochondrial dysfunction
(36). It was also demonstrated
that mtRNA is released into the cytosol and forms intermolecular
dsRNAs that activate cytoplasmic PKR (21,37).
Moreover, it has been shown that different types of cellular
stress, including that induced by hyperglycaemia, can induce
transcription of Alu elements that belong to the short
interspersed nuclear elements (SINE) family and are present at more
than one million copies in the human genome (38,39).
Alu RNA inhibits cytoplasmic PKR, suggesting a functional
role for mammalian SINEs, which have previously been considered
junk DNA (40). Accumulating
evidence supports the concept that Alu RNAs serve a
functional role during the stress response. Alu RNA inhibits
the transcription of genes involved in the response to heat shock
(39) and can inhibit activation of
the cytoplasmic PKR (40). The
induction of Alu element transcription in hyperglycaemic
endothelial cells has been observed, which is responsible for
inhibition of the synthesis of nitric oxide synthase and superoxide
dismutase 2 (41).
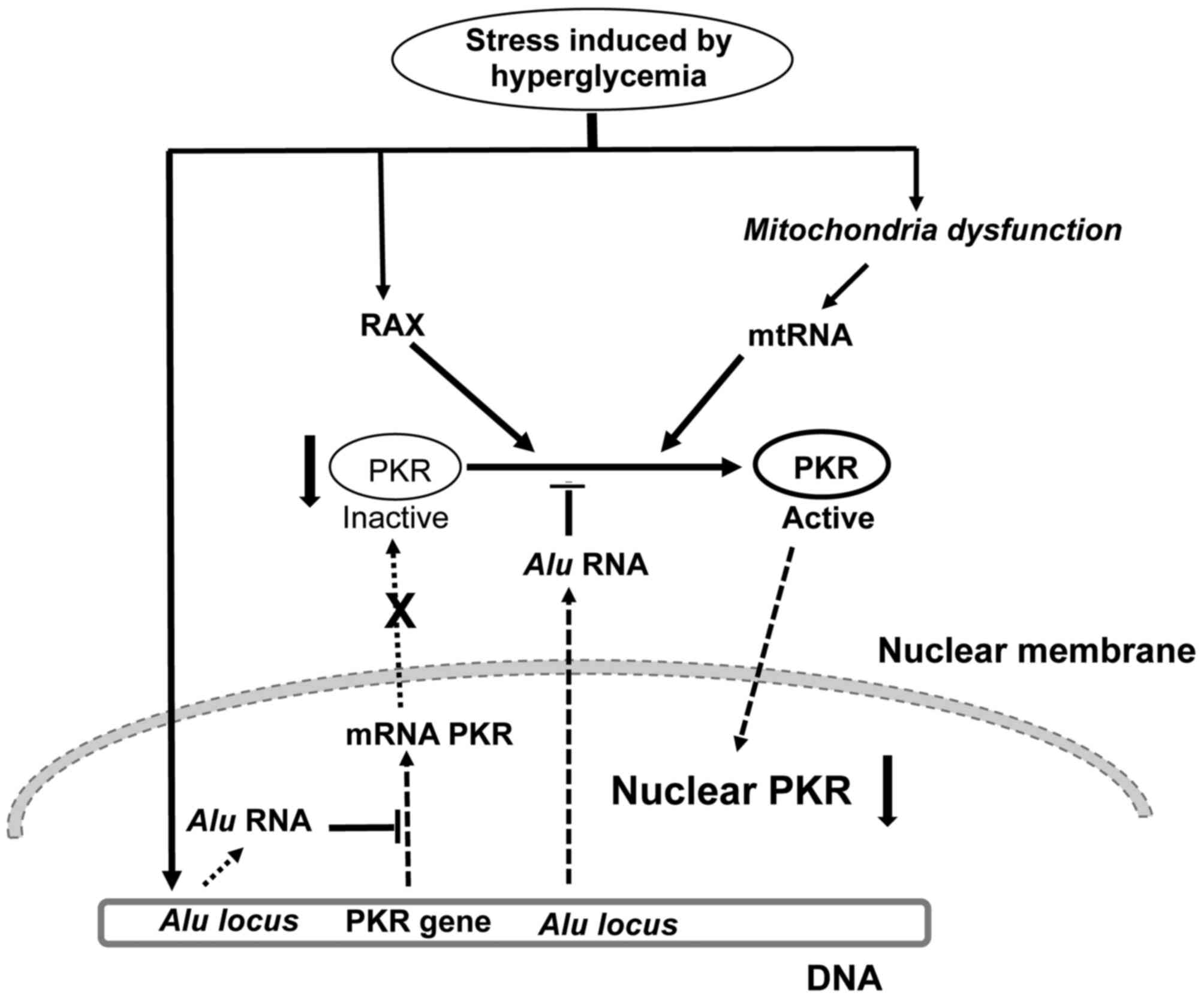
Based on the present results and the literature, we
proposed a mechanism to explain the decrease in nuclear PKR
expression levels in retinal neurons at 35 days after STZ treatment
(Fig. 6). We hypothesize that
stress caused by hyperglycaemia induces RAX expression (26) and mitochondrial dysfunction, with
subsequent release of mtRNA into the cytosol of neurons (21,37).
RAX and mtRNA activate PKR in the cytosol of retinal neurons, which
is followed by its nuclear translocation. Nuclear translocation of
active PKR has previously been suggested as a cell protection
mechanism against the pro-apoptotic activity of PKR (31). Stress caused by hyperglycaemia
induces the expression of Alu RNAs (41), which inhibit transcription of the
PKR gene and activation of PKR (39), leading to a decrease in nuclear PKR
protein expression levels in retinal neurons of STZ-induced
diabetic animals. Thus, the present results suggest a novel
mechanism that may underly the protection of retinal neurons from
apoptosis by PKR at the early stage of DR, in addition to the
inhibition of RAX expression by miR-29b, as previously described
(26). Bone marrow mononuclear
cells of high-risk patients with myelodysplastic syndrome display
nuclear localization of active PKR and a lower number of apoptotic
cells compared with low-risk patients (31). In conclusion, the present study may
contribute to understanding of the biological significance of PKR
nuclear translocation observed in pathologies, such as
myelodysplastic syndrome (31) and
Alzheimer's (33) and
Creutzfeldt-Jakob disease (34).
Acknowledgements
The authors would like to thank Cacilda Pereira and
Zuleica Moraes from the Department of Biochemistry and Immunology
and Neusa Maria Zanon from the Department of Physiology, Ribeirão
Preto School of Medicine, São Paulo, Brazil, for technical
assistance.
Funding
The present study was supported by Research Support
Foundation of the State of São Paulo (grant no. 08/58325-4) and
Coordination for higher Education Staff Development.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
VAOS and FLDL designed the experiments and wrote the
manuscript. VAOS, NDA, TAES, IDCK and VMA performed the
experiments. VAOS and NDA analyzed the data. All authors discussed
the data and commented on the manuscript. VAOS, NDA and FLDL
confirm the authenticity of all the raw data. All authors read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethical
Commission on Animal Research of the School of Medicine at the
University of São Paulo (approval no. 012/2008).
Patient consent for publication
Not applicable.
Competing interests
The authors confirm that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
PKR
|
RNA-dependent protein kinase
|
|
RAX
|
PKR-associated protein X
|
|
mtRNA
|
mitochondrial RNA
|
|
SINE
|
short interspersed nuclear
elements
|
References
|
1
|
Fong DS, Aiello L, Gardner TW, King GL,
Blankenship G, Cavallerano JD, Ferris FL III and Klein R; American
Diabetes Association, : Diabetic retinopathy. Diabetes Care.
26:226–229. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Antonetti DA, Klein R and Gardner TW:
Diabetic retinopathy. N Engl J Med. 366:1227–1239. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Guariguata L, Whiting DR, Hambleton I,
Beagley J, Linnenkamp U and Shaw JE: Global estimates of diabetes
prevalence for 2013 and projections for 2035. Diabetes Res Clin
Pract. 103:137–149. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kowluru RA and Mishra M: Oxidative stress,
mitochondrial damage and diabetic retinopathy. Biochim Biophys
Acta. 1852:2474–2483. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Barber AJ: Diabetic retinopathy: Recent
advances towards understanding neurodegeneration and vision loss.
Sci China Life Sci. 58:541–549. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ung L, Pattamatta U, Carnt N,
Wilkinson-Berka JL, Liew G and White AJR: Oxidative stress and
reactive oxygen species: A review of their role in ocular disease.
Clin Sci. 131:2865–2883. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Barber AJ, Lieth E, Khin SA, Antonetti DA,
Buchanan AG and Gardner TW: Neural apoptosis in the retina during
experimental and human diabetes: Early onset and effect of insulin.
J Clin Invest. 102:783–791. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Oshitari T and Roy S: Diabetes: A
potential enhancer of retinal injury in rat retinas. Neurosc Lett.
390:25–30. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Luo D, Fan Y and Xu X: The effects of
aminoguanidine on retinopathy in STZ-induced diabetic rats. Bioorg
Med Chem Lett. 22:4386–4390. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Naderi A, Zahed R, Aghajanpour L, Amoli FA
and Lashay A: Long term features of diabetic retinopathy in
streptozotocin-induced diabetic Wistar rats. Exp Eye Res.
184:213–220. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gal-Ben-Ari S, Barrera I, Ehrlich M and
Rosenblum K: PKR: A Kinase to Remember. Front Mol Neurosci.
11:4802019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bando Y, Onuki R, Katayama T, Manabe T,
Kudo T, Taira K and Tohyama M: Double-strand RNA dependent protein
kinase (PKR) is involved in the extrastriatal degeneration in
Parkinson's disease and Huntington's disease. Neurochem Int.
46:11–18. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Page G, Rioux Bilan A, Ingrand S,
Lafay-Chebassier C, Pain S, Perault Pochat MC, Bouras C, Bayer T
and Hugon J: Activated double-stranded RNA-dependent protein kinase
and neuronal death in models of Alzheimer's disease. Neuroscience.
139:1343–1354. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
García MA, Gil J, Ventoso I, Guerra S,
Domingo E, Rivas C and Esteban M: Impact of protein kinase PKR in
cell biology: From antiviral to antiproliferative action. Microbiol
Mol Biol Rev. 70:1032–1060. 2006. View Article : Google Scholar
|
|
15
|
Lee ES, Yoon C, Kim Y and Bae Y: The
double-strand RNA-dependent protein kinase PKR plays a significant
role in a sustained ER stress-induced apoptosis. FEBS Lett.
581:4325–4332. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hugon J, Mouton-Liger F, Dumurgier J and
Paquet C: PKR involvement in Alzheimer's disease. Alzheimer Res
Ther. 9:832017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ung TL, Cao C, Lu J, Ozato K and Dever TE:
Heterologous dimerization domains functionally substitute for the
double-stranded RNA binding domains of the kinase PKR. EMBO J.
20:3728–3737. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Williams BR: Signal integration via PKR.
Sci STKE. 2001:re22001.PubMed/NCBI
|
|
19
|
De Lucca FL, Serrano SV, Souza LR and
Watanabe MA: Activation of RNA-dependent protein kinase and nuclear
factor-kB by regulatory RNA from lipopolysaccharide-stimulated
macrophages: Implications for cytokine production. Eur J Pharmacol.
450:85–89. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Robertson HD and Mathews MB: The
regulation of the protein kinase PKR by RNA. Biochimie. 78:909–914.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim Y, Lee JH, Park JE, Cho J, Yi H and
Kim VN: PKR is activated by cellular dsRNAs during mitosis and acts
as a mitotic regulator. Genes Dev. 28:1310–1322. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee SB and Esteban M: The
interferon-induced double-stranded RNA-activated protein kinase
induces apoptosis. Virology. 199:491–496. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gil J and Esteban M: Induction of
apoptosis by the dsRNA-dependent protein kinase (PKR): Mechanism of
action. Apoptosis. 5:107–114. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li B, Wang HS, Li GG, Zhao MJ and Zhao MH:
The role of endoplasmic reticulum stress in the early stage of
diabetic retinopathy. Acta Diabetol. 48:103–111. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ito T, Yang M and May WS: RAX, a cellular
activator for double-stranded RNA-dependent protein kinase during
stress signaling. J Biol Chem. 274:15427–15432. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Silva VA, Polesskaya A, Sousa TA, Corrêa
VM, André ND, Reis RI, Kettelhut IC, Harel-Bellan A and De Lucca
FL: Expression and cellular localization of microRNA-29b and RAX,
an activator of the RNA-dependent protein kinase (PKR), in the
retina of streptozotocin-induced diabetic rats. Mol Vis.
17:2228–2240. 2011.PubMed/NCBI
|
|
27
|
Trinder P: Determination of blood glucose
using 4-amino phenazone as oxygen acceptor. J Clin Pathol.
22:2461969. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Furman BL: Streptozotocin-induced diabetic
models in mice and rats. Curr Protoc Pharmacol. 70:5.47.1–5.47.20.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Muaddi H, Majumder M, Peidis P, Papadakis
AI, Holcik M, Scheuner D, Kaufman RJ, Hatzoglou M and Koromilas AE:
Phosphorylation of eIF2α at serine 51 is an important determinant
of cell survival and adaptation to glucose deficiency. Mol Biol
Cell. 21:3220–3231. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Blalock WL, Bavelloni A, Piazzi M,
Tagliavini F, Faenza I, Martelli AM, Follo MY and Cocco L: Multiple
forms of PKR present in the nuclei of acute leukemia cells
represent an active kinase that is responsive to stress. Leukemia.
25:236–245. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Follo MY, Finelli C, Mongiorgi S, Clissa
C, Bosi C, Martinelli G, Blalock WL, Cocco L and Martelli AM: PKR
is activated in MDS patients and its subcellular localization
depends on disease severity. Leukemia. 22:2267–2269. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hao C, Shao R, Raju U, Fang B, Swisher SG
and Pataer A: Accumulation of RNA-dependent protein kinase (PKR) in
the nuclei of lung cancer cells mediates radiation resistance.
Oncotarget. 7:38235–38242. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bose A, Mouton-Liger F, Paquet C, Mazot P,
Vigny M, Gray F and Hugon J: Modulation of tau phosphorylation by
the kinase PKR: Implications in Alzheimer's disease. Brain Pathol.
21:189–200. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Paquet C, Bose A, Polivka M, Peoch K,
Brouland JP, Keohane C, Hugon J and Gray F: Neuronal phosphorylated
RNA-dependent protein kinase in Creutzfeldt-Jakob disease. J
Neuropathol Exp Neurol. 68:190–198. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Onuki R, Bando Y, Suyama E, Katayama T,
Kawasaki H, Baba T, Tohyama M and Taira K: A RNA-dependent protein
kinase is involved in tunicamycin-induced apoptosis and Alzheimer's
disease. EMBO J. 23:959–968. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dey A and Swaminathan K:
Hyperglycemia-induced mitochondrial alterations in liver. Life Sci.
87:197–214. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim Y, Park J, Kim S, Kim M, Kang MG, Kwak
C, Kang M, Kim B, Rhee HW and Kim VN: PKR senses nuclear and
mitochondrial signals by interacting with endogenous
double-stranded RNAs. Mol Cell. 71:1051–1063.e6. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen LL and Yang L: ALUternative
regulation for gene expression. Trends Cell Biol. 27:480–490. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu WM, Chu WM, Choudary PV and Schmid CW:
Cell stress and translational inhibitors transiently increase the
abundance of mammalian SINE transcripts. Nucleic Acids Res.
23:1758–1765. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chu WM, Ballard R, Carpick BW, Williams BR
and Schmid CW: Potential alu function: Regulation of the activity
of double-stranded rna-activated kinase PKR. Mol Cell Biol.
18:58–68. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang W, Wang W, Azadzoi KM, Dai P, Wang Q,
Sun JB, Zhang WT, Shu Y, Yang JH and Yan Z: Alu RNA accumulation in
hyperglycemia augments oxidative stress and impairs eNOS and SOD2
expression in endothelial cells. Mol Cell Endocrinol. 426:91–100.
2016. View Article : Google Scholar : PubMed/NCBI
|