Introduction
Vascular smooth muscle cells (VSMCs) are the main
cellular components of blood vessels and serve a key role in
intimal hyperplasia, a pathophysiological process that causes
numerous vascular disorders, such as atherosclerosis, hypertension
and post-angioplasty restenosis (1). VSMCs are not terminally differentiated
and maintain considerable diversity and plasticity. For instance,
these cells undergo phenotypic switching under the action of
locally produced growth factors, such as platelet-derived growth
factor-BB (PDGF-BB), and migrate to the intima and proliferate to
induce intimal disease (1). During
the process of neointima formation, VSMCs are characterized by
decreased expression levels of genes associated with cell
differentiation and contraction, as well as increased
proliferation, migration and synthesis of extracellular matrix
components, which lead to the conversion to a synthetic phenotype
(2). Therefore, investigating the
mechanisms underlying VSMC phenotypic modulation is essential for
developing effective treatments in the management of vascular
intimal hyperplasia.
MicroRNAs (miRNAs/miRs) are a cluster of non-coding
RNAs containing 19–25 nucleotides that can regulate >60% of
human genes involved in almost all the cellular biological
behaviors, including proliferation, migration and differentiation
(3). miRNAs exert their biological
functions mainly by binding to the 3′-untranslated region (3′-UTR)
of target genes, leading to the degradation or/and preventing the
translation of mRNA, thereby negatively regulating gene expression
in a post-transcriptional manner (4). Accumulating evidence has shown the
significance of miRNAs in vascular diseases and their potentials as
therapeutic targets (5). For
example, some miRNAs, such as miR-21, miR-26a and miR-29b, have
been reported to participate in arterial intimal hyperplasia by
regulating the phenotypic switching of VSMCs (6).
Homeobox B13 (HOXB13) belongs to the homeobox family
and is a master regulator in cell identity, differentiation,
proliferation and migration during embryonic development,
tumorigenesis and inflammatory reactions (7,8). In
particular, homeobox genes are involved in vasculogenesis and
vascular remodeling, which are common events during neointimal
lesion formation in atherosclerosis and post-angioplasty restenosis
(9). Moreover, HOXB13 is a
well-known regulator of the terminal differentiation of prostate
epithelial cells and represents an important biomarker for prostate
cancer (10). However, it remains
unknown whether HOXB13 participates in regulating the phenotypic
switching, proliferation and migration of VSMCs, and the upstream
regulatory miRNAs in VSMCs are yet to be determined. Therefore,
present study aimed to investigate the mechanisms underlying VSMC
phenotypic modulation.
Materials and methods
Regents and antibodies (Abs)
Recombinant PDGF-BB protein was purchased from
Sigma-Aldrich (Merck KGaA). Abs against HOXB13 (cat. no. sc-28333),
proliferating cell nuclear antigen (PCNA; cat. no. sc-56), α-smooth
muscle actin (α-SMA; cat. no. sc-53142), transgelin (cat. no.
sc-53932), smoothelin (cat. no. sc-376902), MMP-2 (cat. no.
sc-13594), MMP-9 (cat. no. sc-21733) and β-actin (cat. no.
sc-47778) were purchased from Santa Cruz Biotechnology, Inc.
Isolation and culture of VSMCs
In total, six male specific pathogen-free
Sprague-Dawley rats (age, 8 weeks; body weight, 250±20 g) were
obtained from the Animal Laboratory of Nanjing Jiangning Hospital
(Nanjing, China). Rats were housed at 23±2°C under a controlled
humidity (45–65%) with free access to food and water under a 12-h
light/dark cycle. Rats were intraperitoneally injected with 3%
sodium pentobarbital (100 mg/kg). Immediately after the loss of
consciousness, rats underwent thoracotomy and the thoracic aorta
was quickly dissected and separated. At this stage, the rats were
dead. After being rinsed with PBS, a section of the artery was
soaked in sterile Hanks balanced salt solution. The outer membrane
was gently peeled, and the artery was cut into small pieces of
0.5–1 mm3 and placed in a 25-ml culture flask with a
distance of ~0.5 cm. The flask was placed in a 37°C incubator with
a humidified atmosphere of 5% CO2, and 3 h later DMEM
(Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% FBS
(Thermo Fisher Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml
streptomycin was added. The media were refreshed every 3–5 days.
When cells grew to ~70% confluence, they were sub-cultured with
trypsinization. Moreover, 5–6 generations of cells were used for
the experiments.
Cell proliferation assay
VSMCs were seeded at a density of 3×103
cells/well in 96-well plates and incubated overnight with DMEM
supplemented with 10% FBS. The medium was removed and the plates
were washed with PBS. The same volume of serum-free medium was
added and cells were cultured for 12 h. After being washed with
PBS, cells were incubated at 37°C for 24 h with recombinant PDGF-BB
protein at 1, 2, 4, 8, 16 or 32 or 15 ng/ml PDGF-BB protein at 37°C
for 6, 12, 18, 24, 30 or 36 h.
A Cell Counting Kit-8 (CCK-8) assay (Dojindo
Molecular Technologies, Inc.) was used to determine cell
proliferation according to the manufacturer's instructions. The
culture medium was replaced with 100 µl fresh serum-free medium
containing 10 µl CCK-8 solution. The cells were further incubated
for 2 h at 37°C, and the optical density (OD) at 450 nm was
measured. The cell proliferation rate (%) was calculated using the
following formula: (Experimental OD-control OD)/control OD
×100%.
Migration assay
Transwell chambers (8-µm pore size) were purchased
from BD Bioscience. Cells (2×104) suspended in 200 µl
serum-free medium were seeded on the polycarbonate membrane in the
upper Transwell chamber, which was incubated for 60 min at 37°C
with 5% CO2. The lower chamber was then filled with 750
µl medium with 5% FBS. After being incubated for 12, 24 or 36 h at
37°C in a humidified atmosphere of 5% CO2, the Transwell
chamber was washed with PBS and cells on the top surface of the
polycarbonate membrane were removed. At room temperature, cells
that migrated to the bottom surface of the insert were fixed with
100% methanol for 20 min and stained with 0.25% crystal violet
solution for 10 min. Using light microscopy, the cells were counted
from digital images, with 10 visual fields captured randomly at a
magnification of ×200.
Prediction of potential miRNAs
Multiple miRNA prediction tools [miRWalk (version
2.0; http://mirwalk.umm.uni-heidelberg.de/), TargetScan
(version 7.1; targetscan.org/), miRanda (version
1.9; omictools.com/miranda-tool), miRTarBase (version 2.0;
mirtarbase.mbc.nctu.edu.tw/) and
mirdb (mirdb.org/)] were used to predict the potential
upstream miRNAs that can regulate HOXB13 gene by targeting its
3′-UTR.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from cell lysates using
TRIzol® reagent (cat. no. 15596026; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions. RNA
quantity and quality were measured using a NanoDrop ND-1000 system.
For amplifying miR-17-5p, RNA was reversely transcribed into cDNA
using a TaqMan® MicroRNA Reverse Transcription kit (cat
no. 4366597; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol, and a stem-loop RT primer
(5′-GTCGTATCCAGTGCAGGTATTCGCACTGGATACGACCTACC-3′), a sense primer
(5′-TGCAAAGTGCTTACAGTGCAG−3′) and an antisense primer
(5′-GTGCAGGGTCCGAGGTATTC-3′) were used. U6 RNA expression served as
a positive control, using a pair of primers: Sense,
5′-CTCGCTTCGGCAGCACA-3′ and antisense, 5′-AACGCTTCACGAATTTGCGT-3′.
For amplifying HOXB13 mRNA, RNA was reversely transcribed into cDNA
using TaqMan™ Reverse Transcription Reagents (cat. no. N8080234;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol, and a pair of primers (sense, 5′-CAGATGTGTTGCCAGGGAGA-3′
and antisense, 5′-AGGCGTCAGGAGGGTGCT−3′) were used. The expression
level of GAPDH served as an internal control, using a pair of
primers: Sense, 5′-GAAATCCCATCACCATCTTCCAGG−3′ and antisense:
5′-GAGCCCCAGCCTTCTCCATG-3′. Following RT, qPCR was performed using
an Applied Biosystems 7500 Real-Time PCR system and SYBR Green
(cat. no. S7563; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instruction. The thermocycling conditions were as
follows: 10 min initial denaturation at 94°C, 15 sec denaturation
at 94°C and 30 sec of annealing at 55°C (40 cycles) and final
extension for 1 min at 72°C. Experiments were performed in
triplicate, and the relative expression levels were calculated
using the 2−ΔΔCq method (11).
Transfection of oligonucleotides
The miR-17-5p mimics
(5′-CAAAGUGCUUACAGUGCAGGUAG-3′), antagomiR-17-5p
(5′-CUACCUGCACUGUAAGCACUUUG-3′), mimics negative control
(5′-UUCUCCGAACGUGUCACGUTT−3′), antagomiR negative control
(5′-UCACAACCUCCUAGAAAGAGUAGA−3′), small interfering RNA (siRNA)
targeting HOXB13 (sense, 5′-GCGGCCGCAAGAAACGCAUTT−3′ and antisense,
5′-AUGCGUUUUGUGCGGCCGCTT−3′) and siRNA negative control (sense,
5′-UUCUCCGAACGUGUCACGU-3′ and antisense, 5′-ACGUGACACGUUCGGAGAA-3′)
oligonucleotides were purchased from Shanghai GenePharma Co., Ltd.
VSMCs (1×105/well) were seeded in 6-well plates and
cultured with DMEM supplemented with 10% FBS. When cells were grown
to 60–70% confluence, they were incubated with oligonucleotides at
100 nM using Lipofectamine® RNAiMAX transfection reagent
(Thermo Fisher Scientific, Inc.) in serum-free media at 37°C for 48
h, and were then immediately subjected to subsequent assays.
Luciferase reporter assay
The 3′-UTR fragment of HOXB13 mRNA containing
miR-17-5p-targeting sequence was cloned into a pMIR-REPORT
luciferase reporter vector (Ambion; Thermo Fisher Scientific, Inc.)
to create wild-type (WT) and mutated type (MT)
pMIR-luc-HOXB13-3′-UTR vectors. VSMCs (1×104/well) were
cultured in 24-well plates. When at 60–70% confluence, cells were
co-transfected with the vectors (100 nM), together with 50 nM
miR-175p mimics, 50 nM antagomiR-17-5p or 50 nM negative control
oligonucleotides using Lipofectamine® 3000 (Thermo
Fisher Scientific, Inc.) at 37°C. A luciferase reporter vector
without the miR-17-5p targeting sequence was transfected in
parallel. After transfection for 48 h, luciferase activities were
measured using the Dual-luciferase Reporter Assay system (Promega
Corporation) and normalized to Renilla according to the
manufacturer's instructions. The relative luciferase activity in
cells was expressed as a percentage of the luciferase activity over
that of cells transfected with the vector without miR-17-5p
targeting sequence.
Western blot analysis
Cells were homogenized in protein lysate buffer (50
mM Tris, pH 7.4, 100 mM EDTA, 0.25 M sucrose, 1% SDS, 1% NP40, 1
mg/ml leupeptin, 1 mg/ml pepstatin A and 100 mM phenyl methyl
sulfonyl fluoride), and debris was removed via centrifugation at
10,000 × g for 10 min at 4°C. Protein concentrations were
determined using the Bio-Rad protein assay (Bio-Rad Laboratories,
Inc.). Protein (30 µg/lane) were resolved on 10% SDS-polyacrylamide
gels and then electrophoretically transferred to PVDF membranes,
which were blocked in TBS-Tween 20 [137 mM NaCl, 20 mM Tris HCl (pH
7.6) and 0.1% (v/v) Tween 20] containing 5% (w/v) non-fat dry milk
at 37°C for 2 h. The membranes were incubated at 4°C overnight with
primary Abs (Anti-HOXB13, 1:500; anti-PCNA, 1:400; anti-α-SMA,
1:1,000; anti-transgelin, 1:1,000; anti-smoothelin, 1:1,000;
anti-MMP-2, 1:800; anti-MMP-9, 1:1,000; anti-β-actin, 1:1,000),
followed by an incubation with alkaline phosphatase-conjugated
secondary Abs (1:2,000) for 2 h at room temperature in the dark.
Membranes were then developed with 5-bromo-4-chloro-3-indolyl
phosphate/nitro blue tetrazolium (Tiangen Biotech Co., Ltd.). The
density of each band was measured using the ChemiDoc XRS system
(Version 3.0; Bio-Rad Laboratories, Inc.) and the semi-quantitative
measurement of band density was repeated thrice.
Statistical analysis
GraphPad Prism 8.02 (GraphPad Software, Inc.) was
used for statistical analyses. Data are presented as the mean ± SD.
Comparisons were conducted using one-way ANOVA followed by a
Tukey's post hoc test. The correlation between HOXB13 and miR-17-5p
expression levels was assessed using a Pearson's test. P<0.05
was considered to indicate a statistically significant difference.
The experiments were repeated at least three times.
Results
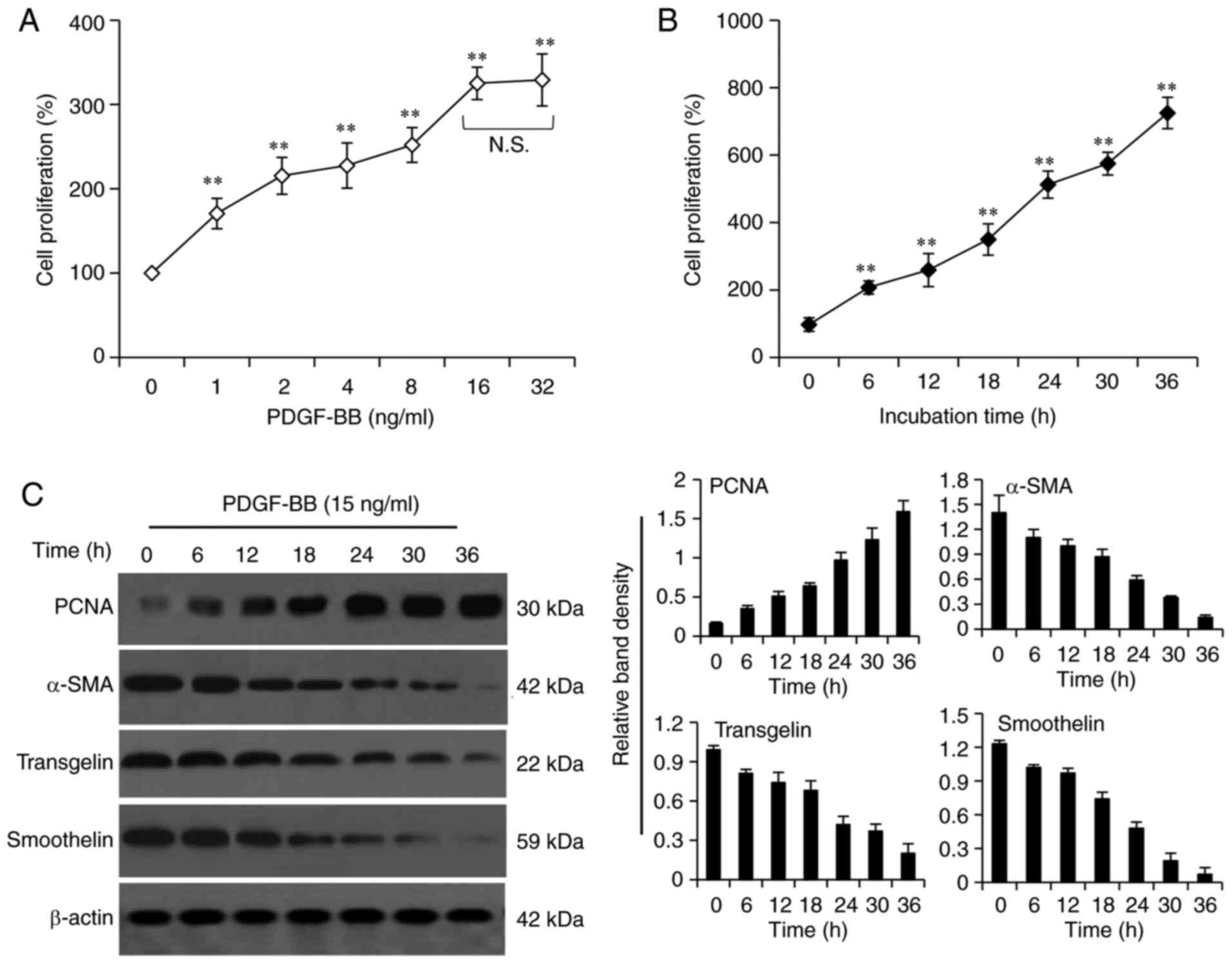
PDGF-BB stimulates the proliferation
and induces phenotypic switching of VSMCs
The stimulation of VSMCs with PDGF-BB protein is
widely used as an in vitro functional model for studying
vascular intimal hyperplasia (12).
VSMCs were first incubated for 24 h with recombinant PDGF-BB
protein at 1, 2, 4, 8, 16 or 32 ng/ml, which were selected based on
previous studies (12–15). The CCK-8 assay results demonstrated
that incubation with the PDGF-BB protein stimulated the
proliferation of VSMCs in a concentration-dependent manner
(Fig. 1A). Since PDGF-BB exerted
its maximal effect at a concentration of 15 ng/ml, this was used
for the subsequent experiments. PDGF-BB stimulation also promoted
the proliferation of VSMCs in a time-dependent manner when VSMCs
were incubated with 15 ng/ml PDGF-BB protein for 6, 12, 18, 24, 30
or 36 h (Fig. 1B). The western blot
analysis results identified the reduced expression levels of VSMC
differentiation-related markers, including α-SMA (16), transgelin (also known as smooth
muscle 22α) (17) and smoothelin
(18), and increased expression
levels of the proliferation marker PCNA, in VSMCs after PDGF-BB
stimulation in a time-dependent manner (Fig. 1C). These results indicated that
PDGF-BB can induce the phenotypic switch and stimulate the
proliferation of VSMCs.
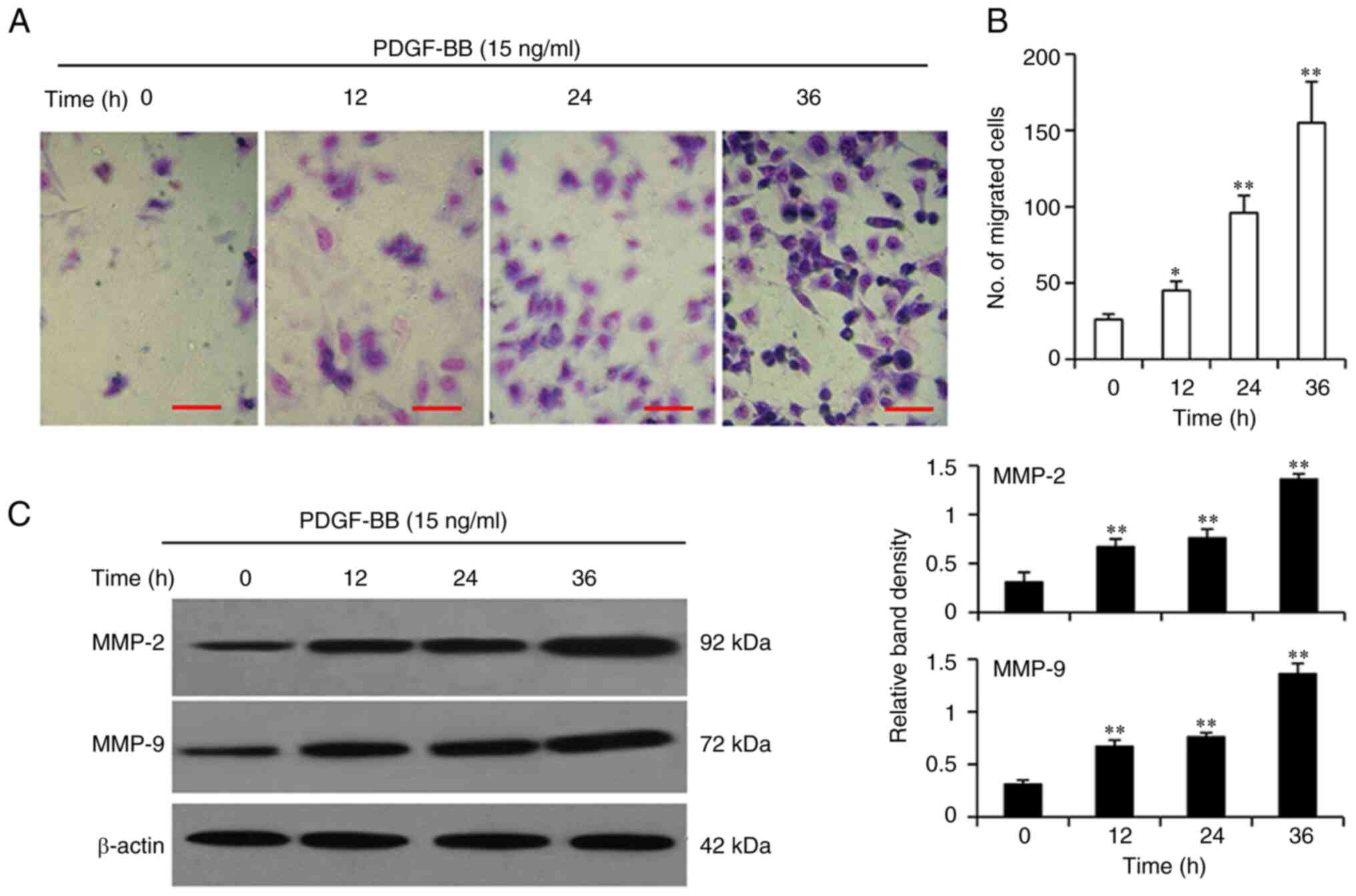
PDGF-BB promotes the migration of
VSMCs
Transwell migration assays revealed that the ability
of VSMCs to migrate was significantly enhanced by PDGF-BB protein
in a time-dependent manner (Fig. 2A and
B). Western blot analysis also confirmed that PDGF-BB
stimulation resulted in an upregulation of MMP-2 and MMP-9
expression in a time-dependent manner (Fig. 2C), which are two important mediators
of VSMC migration (19).
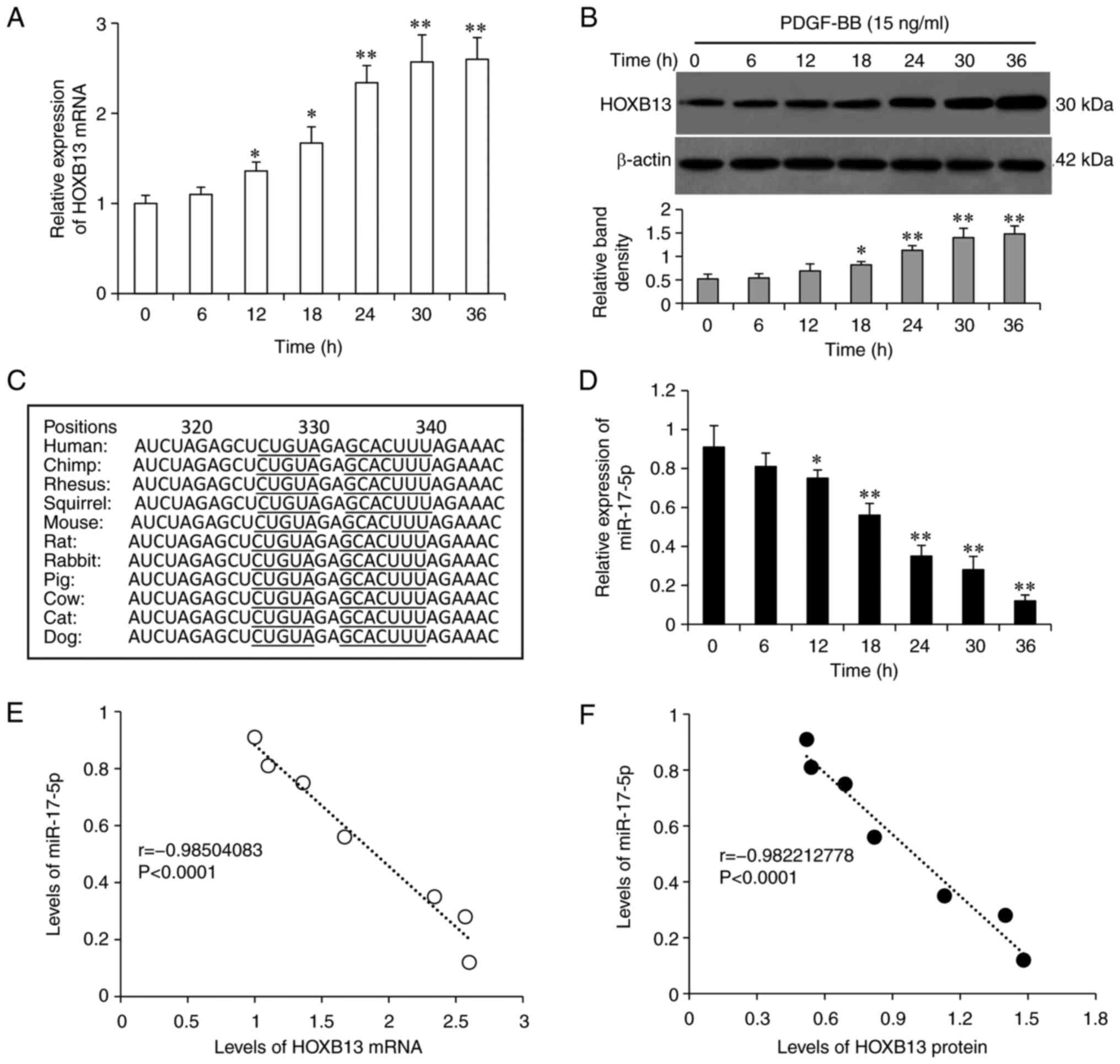
PDGF-BB stimulation leads to the
upregulation of HOXB13 and the downregulation of miR-17-5
It was identified that incubation with 15 ng/ml
PDGF-BB protein increased the mRNA and protein expression levels of
HOXB13 in VSMCs in a time-dependent manner, as determined via
RT-qPCR and western blot analyses, respectively (Fig. 3A and B). Using multiple miRNA
prediction tools, the potential upstream miRNAs that target HOXB13
were examined. miR-17-5p was found to have a 7-base perfect binding
site and a 5-base imperfect binding site with the 3′-UTR of HOXB13
mRNA, and the binding sites were highly conserved across species
(Fig. 3C), indicating that
miR-17-5p may be a potential miRNA to regulate HOXB13. Moreover, a
literature search supported the involvement of miR-17-5p in the
phenotypic modulation of VSMCs (12). It was demonstrated that the
expression levels of miR-17-5p were significantly decreased in
VSMCs upon PDGF-BB stimulation in a time-dependent manner (Fig. 3D). Furthermore, HOXB13 mRNA
(Fig. 3E) and protein (Fig. 3F) expression levels were very
strongly negatively correlated with those of miR-17-5p in VSMCs
given PDGF-BB stimulation for different durations, as determined
using a Pearson's test.
miR-17-5p negatively regulates the
expression level of HOXB13 in VSMCs
Based on the identified binding sites, the WT and MT
pMIR-REPORT luciferase report vectors were constructed (Fig. 4A). VSMCs were co-transfected with
the WT vector and miR-17-5p mimics oligonucleotides, and the
luciferase assays revealed that miR-17-5p inhibited luciferase
activities in a concentration-dependent manner (Fig. 4B). In addition, the miR-17-5p mimics
significantly decreased, while antagomiR-17-5p significantly
increased, the luciferase activity in cells co-transfected with the
WT vector but not the MT vector (Fig.
4C). Moreover, the miR-17-5p mimics significantly reduced,
while the antagomiR-17-5p significantly increased, the mRNA
(Fig. 4D) and protein (Fig. 4E) expression levels of HOXB13. These
results indicated that miR-17-5p served a negative regulatory role
on HOXB13 expression by binding to its 3′-UTR in VSMCs.
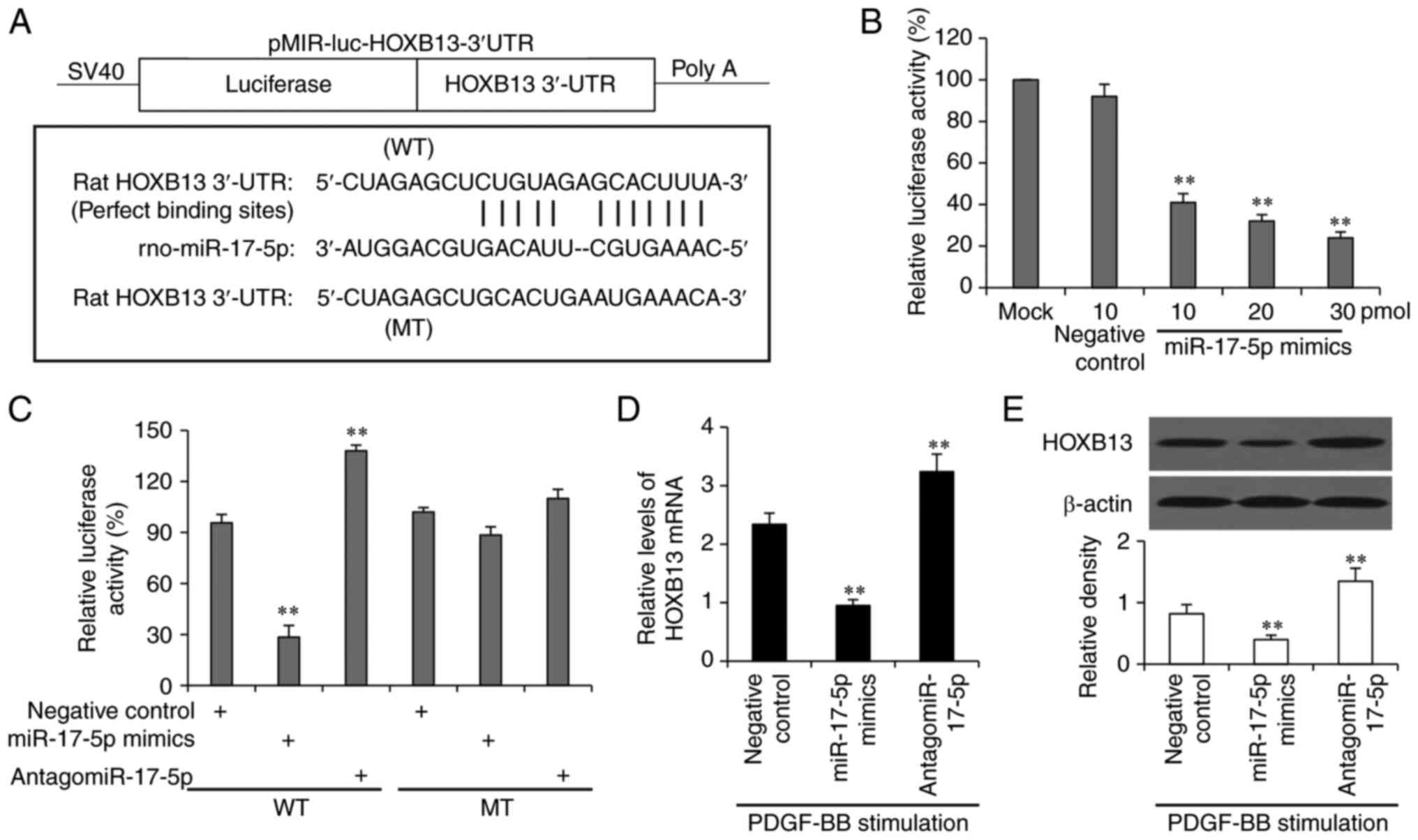 | Figure 4.miR-17-5p negatively regulates the
expression of HOXB13 by specifically binding to its 3′-UTR in
VSMCs. (A) The WT and MT pMIR-REPORT luciferase report vectors were
constructed. (B) VSMCs were either mock-treated or co-transfected
with the WT luciferase vector and 10 pmol negative control or
miR-17-5p mimics oligonucleotides at different concentrations. (C)
VSMCs were co-transfected with WT or MT luciferase reporter
vectors, and negative control, miR-17-5p mimics or antagomiR-17-5p
oligonucleotides. Luciferase activities are shown in (B) and (C)
VSMCs were transfected with negative control, miR-17-5p mimics or
antagomiR-17-5p oligonucleotides, and then subjected to reverse
transcription-quantitative PCR and western blot analyses for
detecting the (D) mRNA and (E) protein expression levels of HOXB13,
respectively. Band density was normalized to β-actin in (E)
**P<0.001 vs. negative control-transfected cells. WT, wild-type;
MT, mutated type; PDGF-BB, platelet-derived growth factor-BB;
VSMCs, vascular smooth muscle cells; miR, microRNA; HOXB13,
homeobox B13; UTR, untranslated region. |
miR-17-5p inhibits the proliferation
and phenotypic switching of VSMCs
Transfection of mimic negative control had no effect
on, while miR-17-5p transfection significantly increased, the
expression level of miR-17-5p in VSMCs (Fig. 5A). Transfection of antagomiR
negative control had no effect on, while antagomiR-17-5p
transfection significantly decreased, the expression level of
miR-17-5p in VSMCs (Fig. 5B).
Because neither mimic nor antagomiR negative controls affected
miR-17-5p expression level, mimic negative control was used as a
negative control in the subsequent experiments. Transfection of
miR-17-5p mimics significantly inhibited the proliferation of VSMCs
under PDGF-BB stimulation, while the antagomiR-17-5p significantly
promoted the proliferation of VSMCs without PDGF-BB stimulation
(Fig. 5C). Furthermore, miR-17-5p
mimics transfection downregulated the expression levels of HOXB13
and PCNA, and increased the expression levels of α-SMA, transgelin
and smoothelin in VSMCs under PDGF-BB stimulation. However,
antagomiR-17-5p transfection showed the opposite effects in VSMCs
without PDGF-BB stimulation (Fig.
5D).
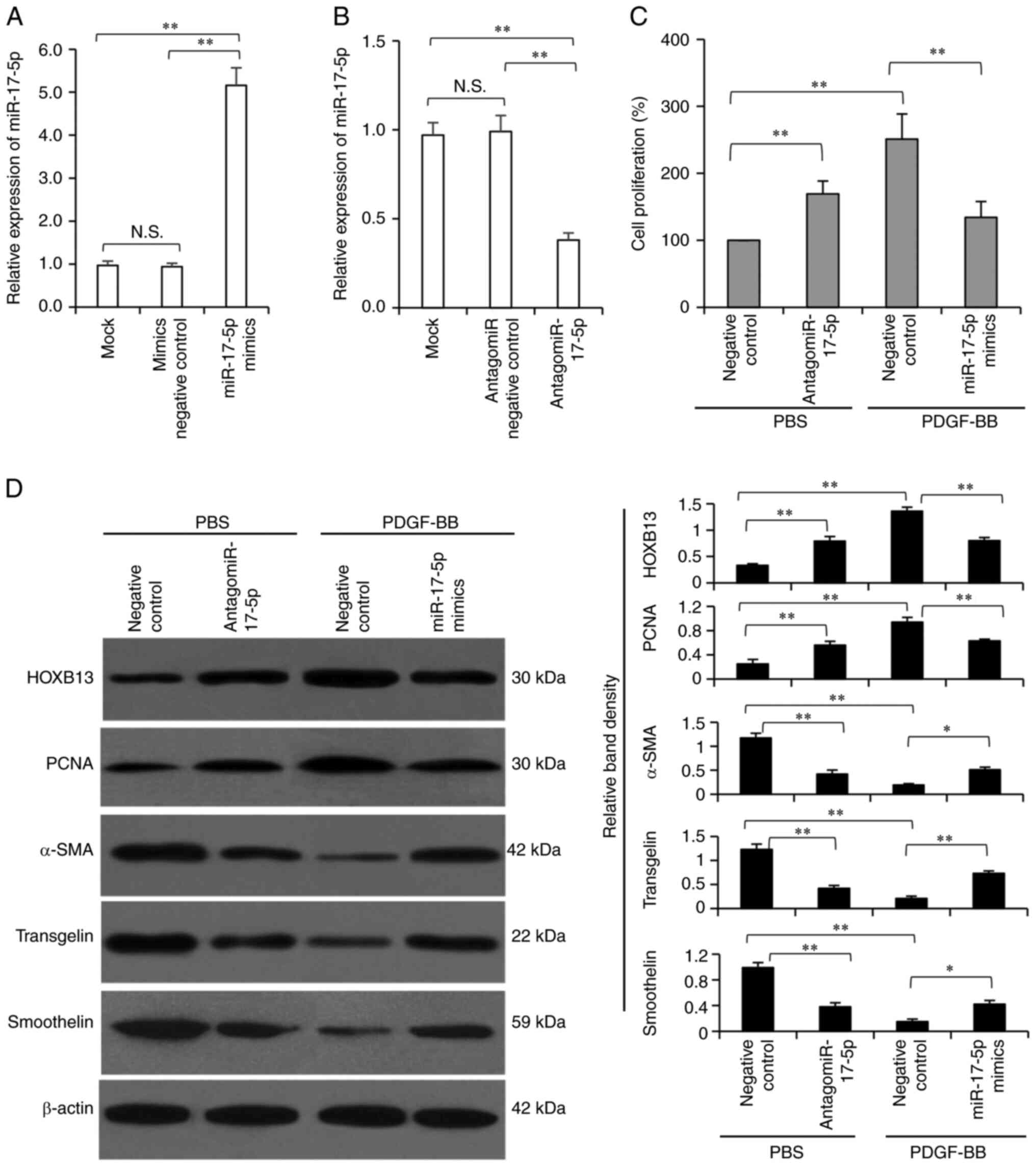 | Figure 5.miR-17-5p regulates the proliferation
and phenotypic switching of VSMCs. (A) VSMCs were mock-treated, or
transfected with mimics negative control or miR-17-5p mimics
oligonucleotides, and the expression level of miR-17-5p was
detected via RT-qPCR. (B) VSMCs were mock-treated, or transfected
with antagomiR negative control or antagomiR-17-5p
oligonucleotides, and the expression level of miR-17-5p was
detected RT-qPCR. VSMCs transfected with negative control,
miR-17-5p mimics or antagomiR-17-5p oligonucleotides were incubated
in the absence or presence of PDGF-BB protein for 24 h. (C) Cell
proliferation was measured. (D) Cells were subjected to western
blot analysis and band density was normalized to β-actin.
*P<0.05 and **P<0.001. N.S., not significant; PDGF-BB,
platelet-derived growth factor-BB; VSMCs, vascular smooth muscle
cells; miR, microRNA; RT-qPCR, reverse transcription-quantitative
PCR; PCNA, proliferating cell nuclear antigen; α-SMA, α-smooth
muscle actin. |
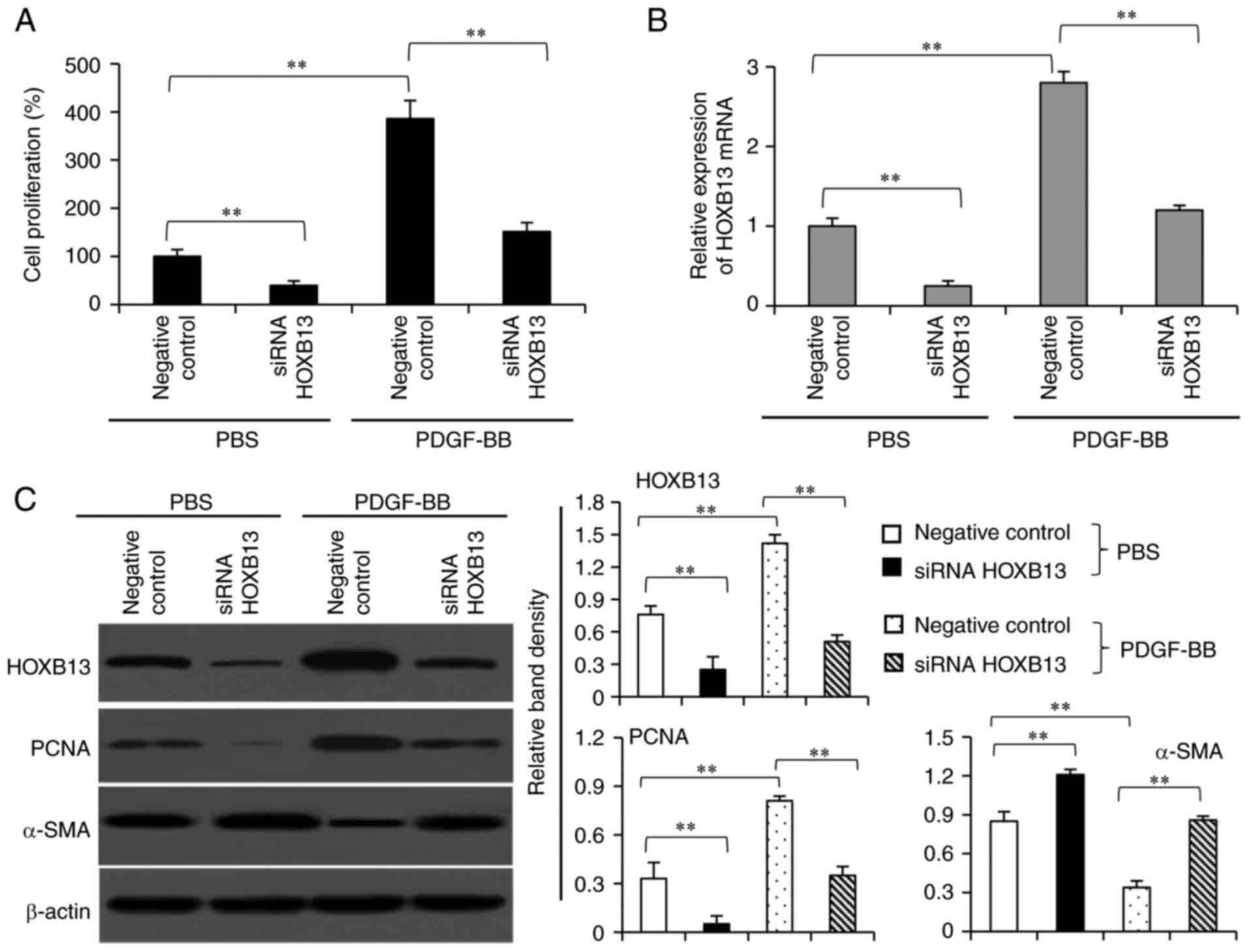
Knockdown of HOXB13 using siRNA
inhibits the proliferation and phenotype switching of VSMCs
Knockdown of HOXB13 using siRNA significantly
inhibited the proliferation of VSMCs in the presence or absence of
PDGF-BB stimulation, compared with the negative controls (Fig. 6A). siRNA HOXB13 also significantly
decreased the mRNA (Fig. 6B) and
protein (Fig. 6C) expression levels
of HOXB13 in VSMCs in the presence or absence of PDGF-BB
stimulation. In addition, transfection of siRNA HOXB13
significantly decreased the expression level of PCNA and increased
that of α-SMA, compared with the negative control (Fig. 6C).
VSMC proliferation, migration and
phenotype switching are regulated via the miR-17-5p/HOXB13
axis
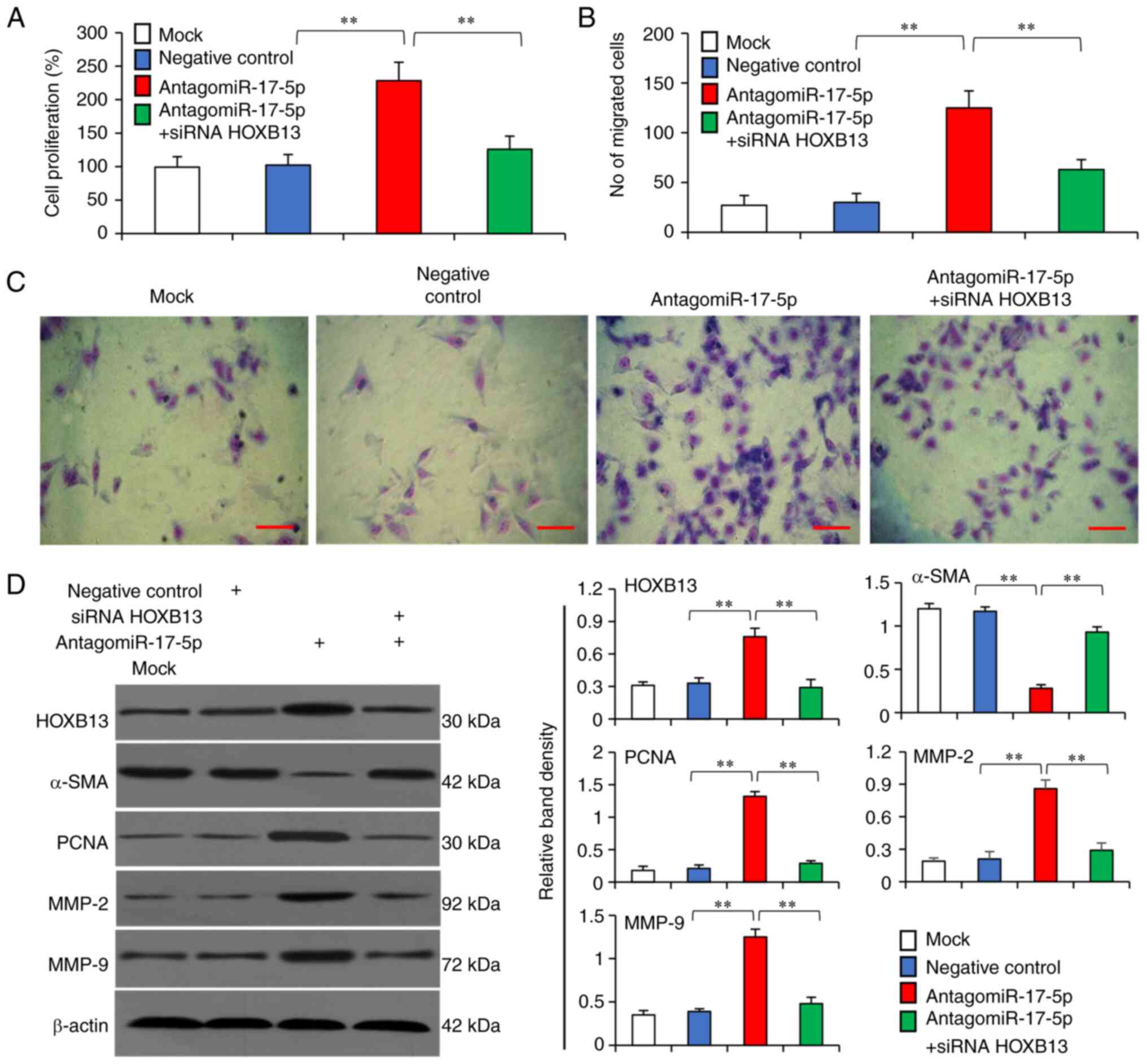
Compared with the negative control, the
antagomiR-17-5p significantly promoted the proliferation of VSMCs,
while knockdown of HOXB13 using siRNA abolished this effect
(Fig. 7A). The antagomiR-17-5p also
increased the migratory capability of VSMCs but knockdown of HOXB13
abrogated this effect (Fig. 7B and
C). Moreover, the antagomiR-17-5p significantly increased the
expression levels of HOXB13, PCNA, MMP-2 and MMP-9, and
downregulated α-SMA expression, while HOXB13 knockdown could
reverse these effects (Fig.
7D).
Discussion
Previous studies have reported that the
transformation of VSMCs from a highly differentiated phenotype to a
poorly differentiated phenotype leads to egregious cell
proliferation, migration and excessive synthesis of extracellular
matrix, resulting in a series of pathological changes, such as
vascular intimal hyperplasia or even luminal stenosis (1,2,12,20–22).
Unfortunately, there are a lack of effective treatment strategies
for this disorder. Therefore, it is crucial to elucidate the
molecular mechanisms underlying the phenotypic switch,
proliferation and migration of VSMCs. A culture of primary VSMCs
under PDGF-BB stimulation is the most commonly used in vitro
model to study this specific pathophysiological response (23). By adopting this model, the current
study identified a novel miR-17-5p/HOXB13 axis that participates in
the regulation of the phenotypic transformation of VSMCs. However,
this axis has not been validated in in vivo models, which is
a limitation of the present study.
miRNAs serve a key role in the regulation of cell
differentiation, proliferation and migration by inhibiting the
translation of target genes or promoting the degradation of mRNAs
(3). Previous in vivo and
in vitro studies have revealed that miRNAs were key
molecular regulators involved in the process of vascular intimal
hyperplasia. For instance, miR-663 regulated the phenotypic
switching of VSMCs by targeting JunB and myosin light chain 9
(24), while miR-15b and miR-16
promoted the conversion of VSMCs to a contractile phenotype and
inhibited vascular neointima formation by targeting Yes-associated
protein (21). Moreover, miR-22
suppressed the formation of vascular neointima by downregulating
methyl-CpG binding protein 2, histone deacetylase 4 and ecotropic
virus integration site 1 protein homolog, and upregulating the
expression levels of VSMC marker genes including α-SMA and smooth
muscle myosin heavy chain (22). It
has also been shown that miR-145 overexpression inhibited intimal
hyperplasia by increasing the expression level of myosin heavy
chain smooth muscle-2 and reducing the expression level of the
proliferation marker Ki-67 in a rabbit model of vein graft disease
(25). Furthermore, miR-143 and
miR-145 cooperatively promote the differentiation and repress the
proliferation of VSMCs by targeting multiple genes (26), and the reduced expression of miR-143
and miR-145 by cholesterol loading decreases the differentiation of
VSMCs (27). These studies indicate
that numerous miRNAs contribute to intimal hyperplasia by
modulating the phenotypic transformation, proliferation and
migration of VSMCs.
In a recent study, Li et al (12) reported that miR-17 knockdown
promoted the phenotypic switching of VSMCs via the upregulation of
interferon regulatory factor 9, which was in agreement with the
present results, although miR-17 was shown to target different
genes between the two studies. Interestingly, a study by Yang et
al (28) revealed that miR-17
promoted the proliferation of VSMCs when upregulated by the
activation of the NF-κB p65 pathway. These inconsistent results
indicate that the role of miR-17 may be cell-specific, as the study
by Yang et al (28) used
human VSMCs, while primary mouse and rat VSMCs were utilized in the
study by Li et al (12) and
the present study. Moreover, miR-26a has been shown to promote the
proliferation and inhibit the differentiation of human VSMCs by
altering the TGF-β pathway signaling (29), but exerted an opposite effect on rat
jugular vein VSMCs by targeting mitogen-activated protein kinase 6
(30). The miR-17-92 cluster
(consisting of six mature miRNAs including miR-17, miR-18a,
miR-19a, miR-19b, miR-20a and miR-92a) increased the proliferation
of mouse VSMCs when its maturation was facilitated by DiGeorge
critical region 8 (31), and
promoted the proliferation of human pulmonary artery VSMCs by
inhibiting prolyl hydroxylase 2 and inducing hypoxia-inducible
factors (32). Based on the
presence of the regulatory mechanisms between miRNAs (3), the results of the two previous studies
(31,32) do not suggest that miR-17 alone has
the same function as the miR-17-92 cluster in enhancing the
proliferation of VSMCs. The miRNA regulatory network is extremely
complex; one miRNA can regulate multiple target genes, while one
target gene may be regulated by multiple miRNAs, and the regulatory
function of miRNAs is affected by several factors (3). Therefore, the function of miR-17-5p in
the phenotypic switching, proliferation and migration of VSMCs
requires further investigation.
As a homeobox transcription factor, HOXB13 serves an
important role in regulating the proliferation, differentiation and
invasion of cells from various cancer types (33). HOXB13 has been considered to be a
prognostic indicator for prostate cancer as it is found to be
mutated in families with an increased risk of early-onset
hereditary prostate cancer (34).
The function of HOXB13 in a variety of diseases has been reported
in recent years (8). It has been
shown that HOXB13 contributes to the maturation and proliferation
of infant cardiomyocytes (35).
Moreover, HOXB13 is highly expressed in liver fibrotic tissues and
its expression level is positively correlated with the degree of
liver inflammation and fibrosis (36). To the best of our knowledge, the
present study may be the first to reveal the role of HOXB13 in
regulating the proliferation, migration and phenotypic switch of
VSMCs.
In conclusion, the present study demonstrated that
HOXB13 was upregulated, while miR-17-5p was downregulated in VSMCs
upon PDGF-BB stimulation. Furthermore, the current study revealed
the regulatory link between miR-17-5p and HOXB13, and elucidated
some of the mechanisms accounting for the phenotypic modulation and
the changes in the biological behaviors of VSMCs. It was found that
miR-17-5p specifically binds to the 3′-UTR of HOXB13 to
downregulate its expression. The miR-17-5p/HOXB13 axis was shown to
be involved in the phenotypic switching, proliferation and
migration of VSMCs by regulating the expression of the VSMC markers
α-SMA, transgelin and smoothelin, the cell proliferating marker
PCNA and the cell migration proteins MMP-2 and MMP-9. Collectively,
the present results indicated that these factors may represent
potential therapeutic targets for the management of intimal
hyperplasia.
Acknowledgements
Not applicable.
Funding
This study was supported by General grants of
Medical Science and Technology Development in Nanjing City, China
(grant no. YKK18211).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HZ conceived and designed the study, and finalized
the manuscript. TY and TW performed the experiments. SK and GZ
contributed to data collection and analysis. KZ contributed to the
result interpretation and manuscript revision. TY and TW drafted
the manuscript. All authors read and approved the final manuscript.
TY and HZ confirming the authenticity of all the raw data. TY and
TW contributed equally to this work.
Ethics approval and consent to
participate
The study had been approved by the Animal Ethics
Committee of Nanjing Jiangning Hospital, (Nanjing, China; approval
no. IACUC-2004055). It is confirmed that animals were anesthetized
and sacrificed using acceptable methods.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang G, Jacquet L, Karamariti E and Xu Q:
Origin and differentiation of vascular smooth muscle cells. J
Physiol. 593:3013–3030. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Alexander MR and Owens GK: Epigenetic
control of smooth muscle cell differentiation and phenotypic
switching in vascular development and disease. Annu Rev Physiol.
74:13–40. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ha M and Kim VN: Regulation of microRNA
biogenesis. Nat Rev Mol Cell Biol. 15:509–524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mellis D and Caporali A: MicroRNA
regulation of vascular function. Vasc Biol. 1:H41–H46. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang D and Atanasov AG: The microRNAs
regulating vascular smooth muscle cell proliferation: A minireview.
Int J Mol Sci. 20:3242019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Holland PW: Evolution of homeobox genes.
Wiley Interdiscip Rev Dev Biol. 2:31–45. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aspuria PJ, Cheon DJ, Gozo MC, Beach JA,
Recouvreux MS, Walts AE, Karlan BY and Orsulic S: HOXB13 controls
cell state through super-enhancers. Exp Cell Res. 393:1120392020.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cantile M, Schiavo G, Terracciano L and
Cillo C: Homeobox genes in normal and abnormal vasculogenesis. Nutr
Metab Cardiovasc Dis. 18:651–658. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Attard G, Parker C, Eeles RA, Schröder F,
Tomlins SA, Tannock I, Drake CG and de Bono JS: Prostate cancer.
Lancet. 387:70–82. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li W, Deng P, Wang J, Li Z and Zhang H:
MiR-17 knockdown promotes vascular smooth muscle cell phenotypic
modulation through upregulated interferon regulator Factor 9
expression. Am J Hypertens. 33:1119–1126. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lee MH, Kwon BJ, Koo MA, You KE and Park
JC: Mitogenesis of vascular smooth muscle cell stimulated by
platelet-derived growth factor-bb is inhibited by blocking of
intracellular signaling by epigallocatechin-3-O-gallate. Oxid Med
Cell Longev. 2013:8279052013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Risinger GM Jr, Hunt TS, Updike DL, Bullen
EC and Howard EW: Matrix metalloproteinase-2 expression by vascular
smooth muscle cells is mediated by both stimulatory and inhibitory
signals in response to growth factors. J Biol Chem.
281:25915–25925. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hsuan CF, Lu YC, Tsai IT, Houng JY, Wang
SW, Chang TH, Chen YL and Chang CC: Glossogyne tenuifolia
attenuates proliferation and migration of vascular smooth muscle
cells. Molecules. 25:58322020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xin H, Wang Z, Wu S, Wang P, Tao X, Xu C
and You L: Calcified decellularized arterial scaffolds impact
vascular smooth muscle cell transformation via downregulating α-SMA
expression and upregulating OPN expression. Exp Ther Med.
18:705–710. 2019.PubMed/NCBI
|
|
17
|
Je HD and Sohn UD: SM22alpha is required
for agonist-induced regulation of contractility: Evidence from
SM22alpha knockout mice. Mol Cells. 23:175–181. 2007.PubMed/NCBI
|
|
18
|
van Eys GJ, Niessen PM and Rensen SS:
Smoothelin in vascular smooth muscle cells. Trends Cardiovasc Med.
17:26–30. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Newby AC: Matrix metalloproteinases
regulate migration, proliferation, and death of vascular smooth
muscle cells by degrading matrix and non-matrix substrates.
Cardiovasc Res. 69:614–624. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dzau VJ, Braun-Dullaeus RC and Sedding DG:
Vascular proliferation and atherosclerosis: New perspectives and
therapeutic strategies. Nat Med. 8:1249–1256. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu F, Ahmed AS, Kang X, Hu G, Liu F, Zhang
W and Zhou J: MicroRNA-15b/16 attenuates vascular neointima
formation by promoting the contractile phenotype of vascular smooth
muscle through targeting YAP. Arterioscler Thromb Vasc Biol.
35:2145–2152. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang F, Chen Q, He S, Yang M, Maguire EM,
An W, Afzal TA, Luong LA, Zhang L and Xiao Q: miR-22 Is a novel
mediator of vascular smooth muscle cell phenotypic modulation and
neointima formation. Circulation. 137:1824–1841. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Montezano AC, Lopes RA, Neves KB, Rios F
and Touyz RM: Isolation and culture of vascular smooth muscle cells
from small and large vessels. Methods Mol Biol. 1527:349–354. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li P, Zhu N, Yi B, Wang N, Chen M, You X,
Zhao X, Solomides CC, Qin Y and Sun J: MicroRNA-663 regulates human
vascular smooth muscle cell phenotypic switch and vascular
neointimal formation. Circ Res. 113:1117–1127. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ohnaka M, Marui A, Yamahara K, Minakata K,
Yamazaki K, Kumagai M, Masumoto H, Tanaka S, Ikeda T and Sakata R:
Effect of microRNA-145 to prevent vein graft disease in rabbits by
regulation of smooth muscle cell phenotype. J Thorac Cardiovasc
Surg. 148:676–682.e2. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cordes KR, Sheehy NT, White MP, Berry EC,
Morton SU, Muth AN, Lee TH, Miano JM, Ivey KN and Srivastava D:
miR-145 and miR-143 regulate smooth muscle cell fate and
plasticity. Nature. 460:705–710. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vengrenyuk Y, Nishi H, Long X, Ouimet M,
Savji N, Martinez FO, Cassella CP, Moore KJ, Ramsey SA, Miano JM
and Fisher EA: Cholesterol loading reprograms the
microRNA-143/145-myocardin axis to convert aortic smooth muscle
cells to a dysfunctional macrophage-like phenotype. Arterioscler
Thromb Vasc Biol. 35:535–546. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang D, Sun C, Zhang J, Lin S, Zhao L,
Wang L, Lin R, Lv J and Xin S: Proliferation of vascular smooth
muscle cells under inflammation is regulated by NF-κB
p65/microRNA-17/RB pathway activation. Int J Mol Med. 41:43–50.
2018.PubMed/NCBI
|
|
29
|
Leeper NJ, Raiesdana A, Kojima Y, Chun HJ,
Azuma J, Maegdefessel L, Kundu RK, Quertermous T, Tsao PS and Spin
JM: MicroRNA-26a is a novel regulator of vascular smooth muscle
cell function. J Cell Physiol. 226:1035–1043. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tan J, Yang L, Liu C and Yan Z:
MicroRNA-26a targets MAPK6 to inhibit smooth muscle cell
proliferation and vein graft neointimal hyperplasia. Sci Rep.
7:466022017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen Z, Wu J, Yang C, Fan P, Balazs L,
Jiao Y, Lu M, Gu W, Li C, Pfeffer LM, et al: DiGeorge syndrome
critical region 8 (DGCR8) protein-mediated microRNA biogenesis is
essential for vascular smooth muscle cell development in mice. J
Biol Chem. 287:19018–19028. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen T, Zhou Q, Tang H, Bozkanat M, Yuan
JX, Raj JU and Zhou G: miR-17/20 Controls prolyl hydroxylase 2
(PHD2)/Hypoxia-Inducible Factor 1 (HIF1) to regulate pulmonary
artery smooth muscle cell proliferation. J Am Heart Assoc.
5:e0045102016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang X, Sun Y, Xu T, Qian K, Huang B,
Zhang K, Song Z, Qian T, Shi J and Li L: HOXB13 promotes
proliferation, migration, and invasion of glioblastoma through
transcriptional upregulation of lncRNA HOXC-AS3. J Cell Biochem.
120:15527–15537. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ewing CM, Ray AM, Lange EM, Zuhlke KA,
Robbins CM, Tembe WD, Wiley KE, Isaacs SD, Johng D, Wang Y, et al:
Germline mutations in HOXB13 and prostate-cancer risk. N Engl J
Med. 366:141–149. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nguyen NUN, Canseco DC, Xiao F, Nakada Y,
Li S, Lam NT, Muralidhar SA, Savla JJ, Hill JA, Le V, et al: A
calcineurin-Hoxb13 axis regulates growth mode of mammalian
cardiomyocytes. Nature. 582:271–276. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zuo L, Tan T, Wei C, Wang H, Tan L, Hao Y,
Qian J, Chen Y and Wu C: HOXB13 expression is correlated with
hepatic inflammatory activity of patients with hepatic fibrosis. J
Mol Histol. 51:183–189. 2020. View Article : Google Scholar : PubMed/NCBI
|