Introduction
Liver cancer (LC) is one of the most frequently
occurring gastrointestinal malignancies and leading causes of
tumor-related death worldwide (1,2).
Certain primary risk factors, including alcohol abuse and hepatitis
virus infection, are correlated with LC pathogenesis (3). Despite advances in early diagnosis and
therapeutic strategies, including surgical resection, radiotherapy
and chemotherapy (4,5), the 5-year overall survival rate for LC
(34%) has not significantly improved, primarily due to the high
rate of recurrence and metastasis (6). Therefore, improving the current
understanding of the molecular mechanisms associated with
hepatocarcinogenesis is important for improving the prognosis of
LC.
Long non-coding RNAs (lncRNAs) are a class of
non-protein coding RNAs that are >200 nucleotides in length.
lncRNAs have become a hot research topic due to their important
regulatory functions in various biological behaviors, including
proliferation, differentiation and metastasis (7,8).
Accumulating evidence has indicated that lncRNAs are frequently
aberrantly expressed, and function as positive or negative
regulators of coding genes in the progression of different types of
cancer, including LC, colorectal cancer and lung cancer (9–11). For
instance, Jiang et al (12)
reported that upregulated double homeobox A pseudogene 8 expression
predicted poor outcomes in patients with LC and promoted cell
proliferation in vitro. Zheng et al (13) highlighted the oncogenic role of long
intergenic non-protein coding RNA 467 in LC progression via
regulating the microRNA-18a/neural precursor cell expressed,
developmentally downregulated 9 axis. Moreover, lncRNA F11
antisense RNA 1 negatively regulated LC cell proliferation,
migration and invasion (14). Novel
lncRNA tumor protein translationally controlled 1 antisense RNA 1
(TPT1-AS1) has been reported to be associated with the prognosis of
patients with glioma (15,16). Moreover, the oncogenic function of
TPT1-AS1 has been further validated in other tumors. For example,
Jiang et al (17) not only
demonstrated that high TPT1-AS1 was correlated with adverse
prognostic characteristics, but also reported that TPT1-AS1
promoted cell proliferation and metastasis in cervical cancer.
Jiang et al (17)
demonstrated the oncogenic effects of TPT1-AS1 on epithelial
ovarian cancer cell proliferation, migration and invasion. In
addition, Jia et al (18)
demonstrated that TPT1-AS1 inhibited glioma cell autophagy and
promoted cell proliferation. In gastric cancer, TPT1-AS1 knockdown
significantly inhibited cell proliferation, G1/S
transition and epithelial-mesenchymal transition (EMT) (19). However, the clinical significance
and functional roles of TPT1-AS1 in LC are not completely
understood.
The present study aimed to explore the role of
TPT1-AS1 expression in LC tissues and cells, as well as the
association between TPT1-AS1 and LC clinicopathological features or
survival prognosis were investigated. Subsequently,
loss-of-function experiments, including Cell Counting Kit-8
(CCK-8), colony formation, flow cytometry, wound healing and
Transwell assays, were performed to explore the potential effects
of TPT1-AS1 on LC cell functions. The results of the present study
may improve the current understanding of LC progression to aid with
the identification of novel therapeutic targets for LC.
Materials and methods
Clinical sample collection
Tumor tissues and matched adjacent paracancerous
tissues (2 cm away from the tumor margin) were collected from 50
patients with LC at Taizhou People's Hospital (Jiangsu, China)
between March 2019 and March 2021. Tissues were pathologically
confirmed. The patients had not received any antitumor treatments.
Collected tissues were immediately frozen in liquid nitrogen and
stored at −80°C until further analysis. The basic
clinicopathological characteristics, including gender, age and TNM
stage (20), of the patients are
presented in Table I. Written
informed consent was obtained from all patients. The present study
was performed in accordance with the Declaration of Helsinki and
approved by the Ethical Research Committee of Taizhou People's
Hospital.
 | Table I.Association between TPT1-AS1
expression and clinicopathological characteristics of patients with
liver cancer. |
Table I.
Association between TPT1-AS1
expression and clinicopathological characteristics of patients with
liver cancer.
|
|
| TPT1-AS1
expression |
|
|---|
|
|
|
|
|
|---|
| Characteristic | n (n=50) | High (n=25) | Low (n=25) | P-value |
|---|
| Age, years |
|
|
| 0.248 |
|
<60 | 20 | 12 | 8 |
|
|
≥60 | 30 | 13 | 17 |
|
| Sex |
|
|
| 0.777 |
|
Male | 27 | 13 | 14 |
|
|
Female | 23 | 12 | 11 |
|
|
Differentiation |
|
|
| 0.371 |
|
Well/moderate | 33 | 18 | 15 |
|
|
Poor | 17 | 7 | 10 |
|
| TNM stage |
|
|
| 0.024a |
|
I–II | 26 | 9 | 17 |
|
|
III–IV | 24 | 16 | 8 |
|
| Lymph node
metastasis |
|
|
| 0.048a |
|
Negative | 25 | 9 | 16 |
|
|
Positive | 25 | 16 | 9 |
|
Cell transfection
Human LC cell lines (HepG2 and SNU-182) and a
transformed human liver epithelial-3 cell line (THLE-3) were
purchased from American Type Culture Collection. All cell lines
were authenticated via STR profiling. Cells were cultured in DMEM
(Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.) at 37°C with 5%
CO2. Specific small interfering (si)RNAs targeted
against TPT1-AS1 were used, including si-TPT1-AS1#1
(5′-AAGGTACCGAAAGCACAGTAA-3′), si-TPT1-AS1#2
(5′-AACCATCACCTGCAGGAAACA-3′) and a scrambled siRNA control (si-NC)
(5′-AACCATCACTTACAAGAAACC-3′), which were purchased from Shanghai
GeneChem Co., Ltd. HepG2 and SNU-182 cells (2×104
cells/well) were transfected with 50 nM si-TPT1-AS1#1,
si-TPT1-AS1#2 or si-NC using Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.) for 48 h at 37°C. At
48 h post-transfection, cells were used for subsequent
experiments.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from tissue samples or cell
lines using TRIzol® reagent (Takara Bio, Inc.). Total
RNA was reverse transcribed into cDNA using the PrimeScript TM RT
Master Mix kit (Takara Bio, Inc.) according to the manufacturer's
protocol. Subsequently, qPCR was performed using an ABI 7500
Real-time PCR instrument (Applied Biosystems; Thermo Fisher
Scientific, Inc.) with SYBR-Green Master mix (Toyobo Life Science).
The following thermocycling conditions were used for qPCR: Initial
denaturation at 95°C for 1 min; followed by 40 cycles at 95°C for
20 sec and 60°C for 40 sec. The following primers were used for
qPCR: TPT1-AS1 forward, 5′-AGCCTTGAGGCTATGCCCATC-3′ and reverse,
5′-AACAGTGTTGGAGGCCTCTGAA-3′; and GAPDH forward,
5′-GCCCTCCGACACCCACTACCTTT-3′ and reverse,
5′-TGAATTCTGTAGCCACGTTGTCATA-3′. mRNA expression levels were
quantified using the 2−ΔΔCq method (21) and normalized to the internal
reference gene GAPDH.
CCK-8 assay
At 48 h post-transfection, cells were seeded
(3×103 cells/well) into 96-well plates and incubated
with 10 µl CCK-8 solution (Sigma-Aldrich; Merck KGaA) for 24, 48 or
72 h. Then, the culture medium was changed and following incubation
for a further 2 h, cells were harvested and absorbance was measured
at a wavelength of 450 nm using a microplate reader.
Colony formation assay
Transfected cells were seeded (5×102
cells/well) into 6-well plates and cultured for 2 weeks under
standard culture conditions. Subsequently, colonies were washed
twice with PBS, fixed with 70% methanol for 30 min and stained with
0.1% crystal violet for 1 h at room temperature. Colonies (>50
cells) were observed and counted using a light microscope
(magnification, ×40).
Cell cycle analysis
At 48 h post-transfection, cells were washed twice
with precooled PBS and fixed with precooled 70% ethanol overnight
at 4°C. Cells were centrifuged for 5 min at 112 × g at 4°C, and the
supernatant was discarded. Subsequently, cells were incubated with
20 µl RNase A solution and 0.2% Triton X-100 at 37°C for 10 min,
followed by incubation with 400 µl PI staining solution (Beyotime
Institute of Biotechnology) at 37°C for 20 min in the dark. Cell
cycle distribution was analyzed using a flow cytometer (BD
FACSLyric™ Flow Cytometer; BD Biosciences). FCSalyzer 0.9.22 alpha
software (https://sourceforge.net/projects/fcsalyzer/) was used
for data analysis.
Wound healing assay
Cell migration was assessed by performing a wound
healing assay. Briefly, transfected cells were seeded
[5×105 cells/well containing 5% (vol/vol) FBS] into
6-well plates. Subsequently, a 200 µl pipette tip was used to make
a scratch in the (95–100%) confluent cell monolayer. The wound was
observed at 0 and 24 h, which were recorded as W0 and 24,
respectively, using a light inverted microscope (magnification,
×40). Cell migration (%) was calculated according to the formula:
(W0-24)/W0 ×100.
Cell invasion assay
Cell invasion was assessed using 24-well
Matrigel-coated Transwell invasion inserts (BD Biosciences).
Briefly, cells (5×104) were resuspended in serum-free
DMEM and plated into the upper chamber, whereas DMEM supplemented
with 20% FBS was added to the lower chamber. Following incubation
for 24 h at 37°C, invading cells were fixed with methanol for 15
min and stained with 0.2% crystal violet for 30 min at room
temperature. Invasive cells were visualized using an inverted
microscope (magnification, ×40) and photographed. Cells were
counted in five randomly selected fields.
Western blotting
Total protein was extracted from tissues and
cultured cells using RIPA lysis buffer (Beyotime Institute of
Biotechnology) and quantified using a BCA Protein Assay kit
(Beyotime Institute of Biotechnology). Proteins (10 µg/well) were
separated via 10% SDS-PAGE and transferred onto PVDF membranes (EMD
Millipore), which were blocked with 5% skimmed milk for 1 h at room
temperature. Subsequently, the membranes were incubated overnight
at 4°C with primary antibodies (all purchased from Abcam) targeted
against: CDK4 (cat. no. ab137675; 1:1,000), Cyclin D1 (cat. no.
ab134175; 1:1,000), p21 (cat. no. ab188224; 1:1,000), E-cadherin
(cat. no. ab1416; 1:1,000), N-cadherin (cat. no. ab76057; 1:1,000),
Vimentin (cat. no. ab24525; 1:1,000) and GAPDH (cat. no. ab8245;
1:1,000). Following primary incubation, the membranes were washed
twice with PBS and incubated with a HRP-conjugated anti-Rabbit
secondary antibody (cat. no. ab6721; 1:10,000; Abcam) for 2 h at
room temperature. Protein bands were visualized using an ECL
Detection kit (Thermo Fisher Scientific, Inc.). GAPDH was used as
the loading control.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism software (version 6.0; GraphPad Software, Inc.). The
χ2 test was used to analyze the association between
TPT1-AS1 and clinicopathological characteristics. Survival curves
were plotted using the Kaplan-Meier method and statistically
compared using the log-rank test. Quantitative data are presented
as the mean ± SD of three independent experiments. Comparisons
between two groups were analyzed using the paired or unpaired
Student's t-test. Comparisons among multiple groups were analyzed
using one-way ANOVA followed by Bonferroni's post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
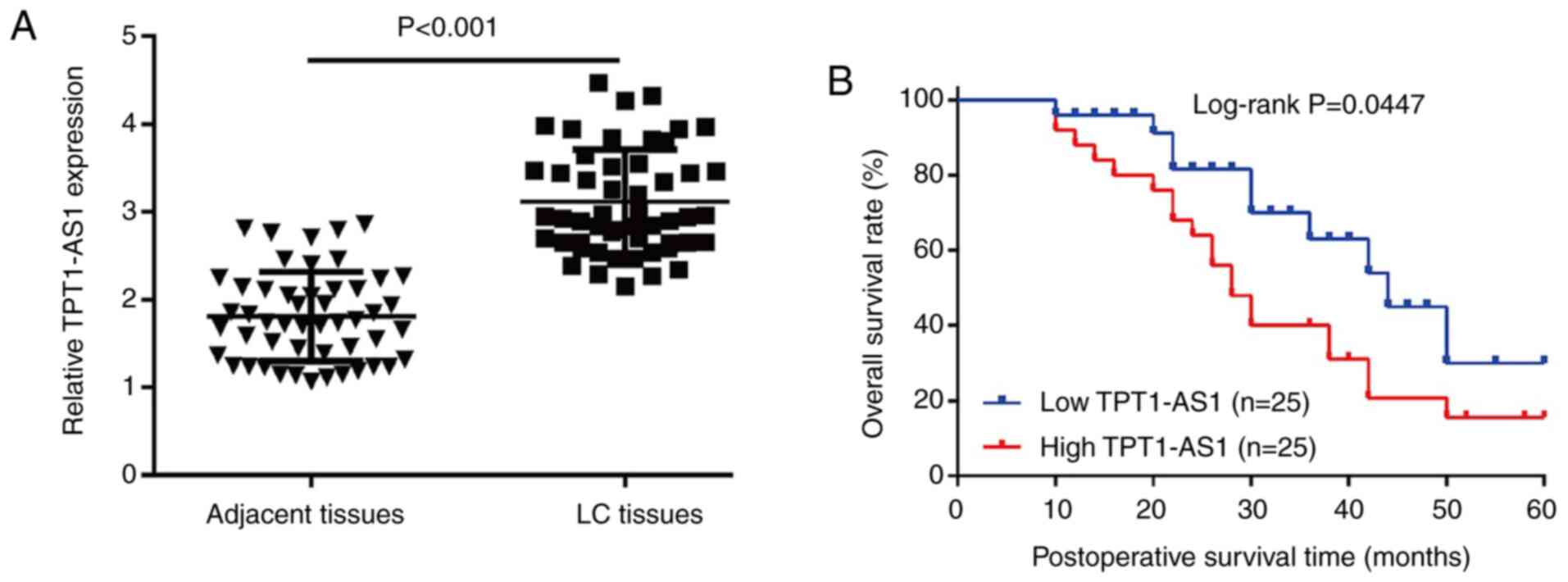
Significantly upregulated TPT1-AS1
expression is associated with poor survival prognosis in patients
with LC
To evaluate the clinical significance of TPT1-AS1 in
LC, TPT1-AS1 expression levels were determined in 50 paired LC and
adjacent paracancerous tissues via RT-qPCR. TPT1-AS1 expression
levels were significantly increased in LC tissues compared with
adjacent paracancerous tissues (Fig.
1A). Subsequently, the 50 patients with LC were divided into
TPT1-AS1 high expression group (n=25) and low expression (n=25)
groups using the median TPT1-AS1 expression level (cut-off, 0.62)
in LC tissues as a cut-off value. Moreover, high TPT1-AS1
expression was more frequent in patients with late TNM stage and
lymph node metastasis compared with patients with early TNM stage
and no lymph node metastasis, respectively (Table I). The Kaplan-Meier survival
analysis demonstrated that patients with LC with high TPT1-AS1
expression displayed significantly worse overall survival compared
with patients with LC with low TPT1-AS1 expression (Fig. 1B).
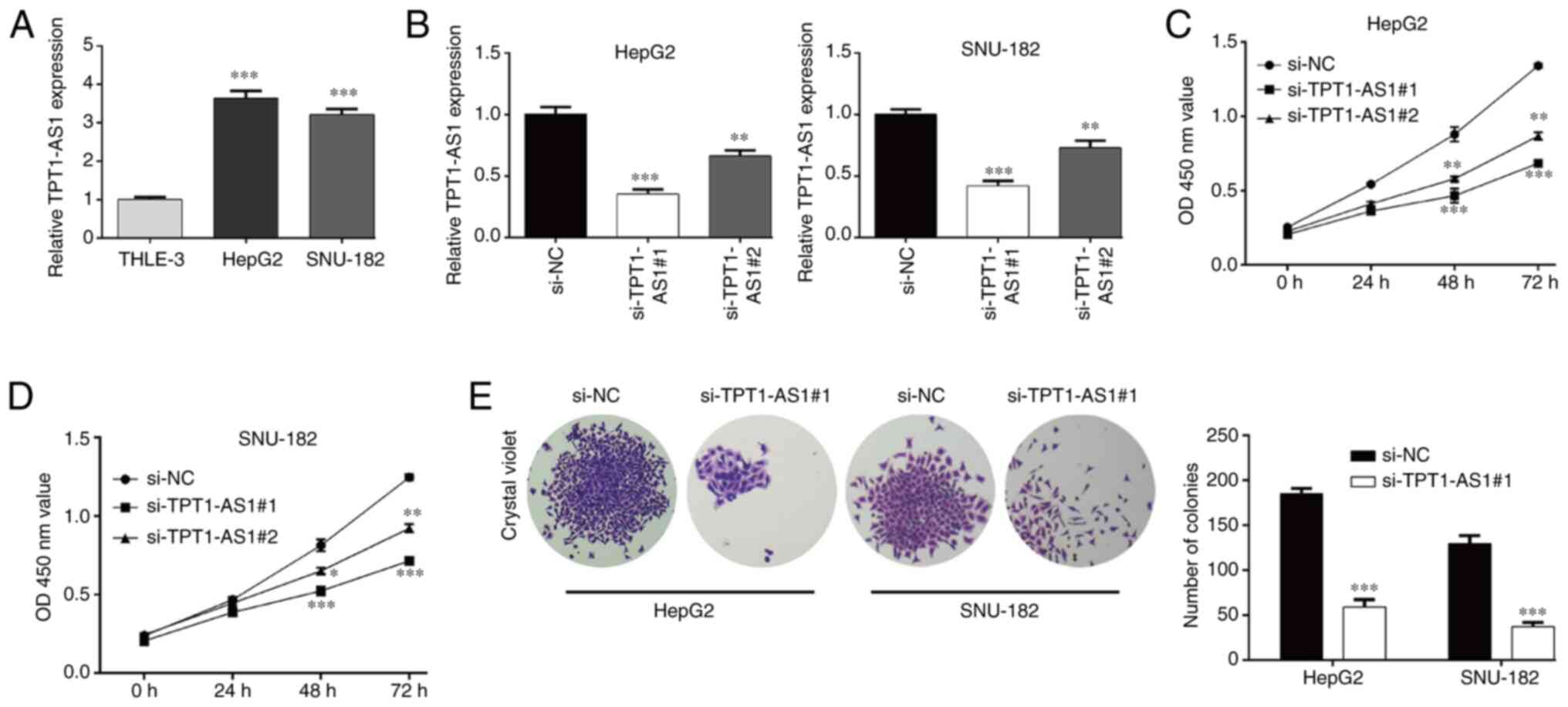
TPT1-AS1 knockdown inhibits LC cell
proliferation and induces G0/G1 phase
arrest
Consistent with TPT1-AS1 expression in LC tissues,
TPT1-AS1 expression levels were significantly upregulated in the LC
cell lines (HepG2 and SNU-182) compared with the THLE-3 cell line
(Fig. 2A). Subsequently, TPT1-AS1
expression was knocked down to investigate the function of TPT1-AS1
in HepG2 and SNU-182 cells. The RT-qPCR results indicated that
si-TPT1-AS1#1 and si-TPT1-AS1#2 significantly decreased TPT1-AS1
expression levels in HepG2 and SNU-182 cells compared with the
si-NC group (Fig. 2B). The CCK-8
assay results demonstrated that compared with the si-NC group,
TPT1-AS1 knockdown significantly suppressed HepG2 (Fig. 2C) and SNU-182 (Fig. 2D) cell viability at 48 and 72 h.
Moreover, si-TPT1-AS1#1 displayed enhanced suppressive effects on
TPT1-AS1 expression and cell viability compared with si-TPT1-AS1#2,
thus si-TPT1-AS1#1 was selected for subsequent experiments.
Similarly, HepG2 and SNU-182 cell proliferation was significantly
inhibited by TPT1-AS1 knockdown, as evidenced by a significantly
decreased number of colonies in the si-TPT1-AS1#1 group compared
with the si-NC group (Fig. 2E).
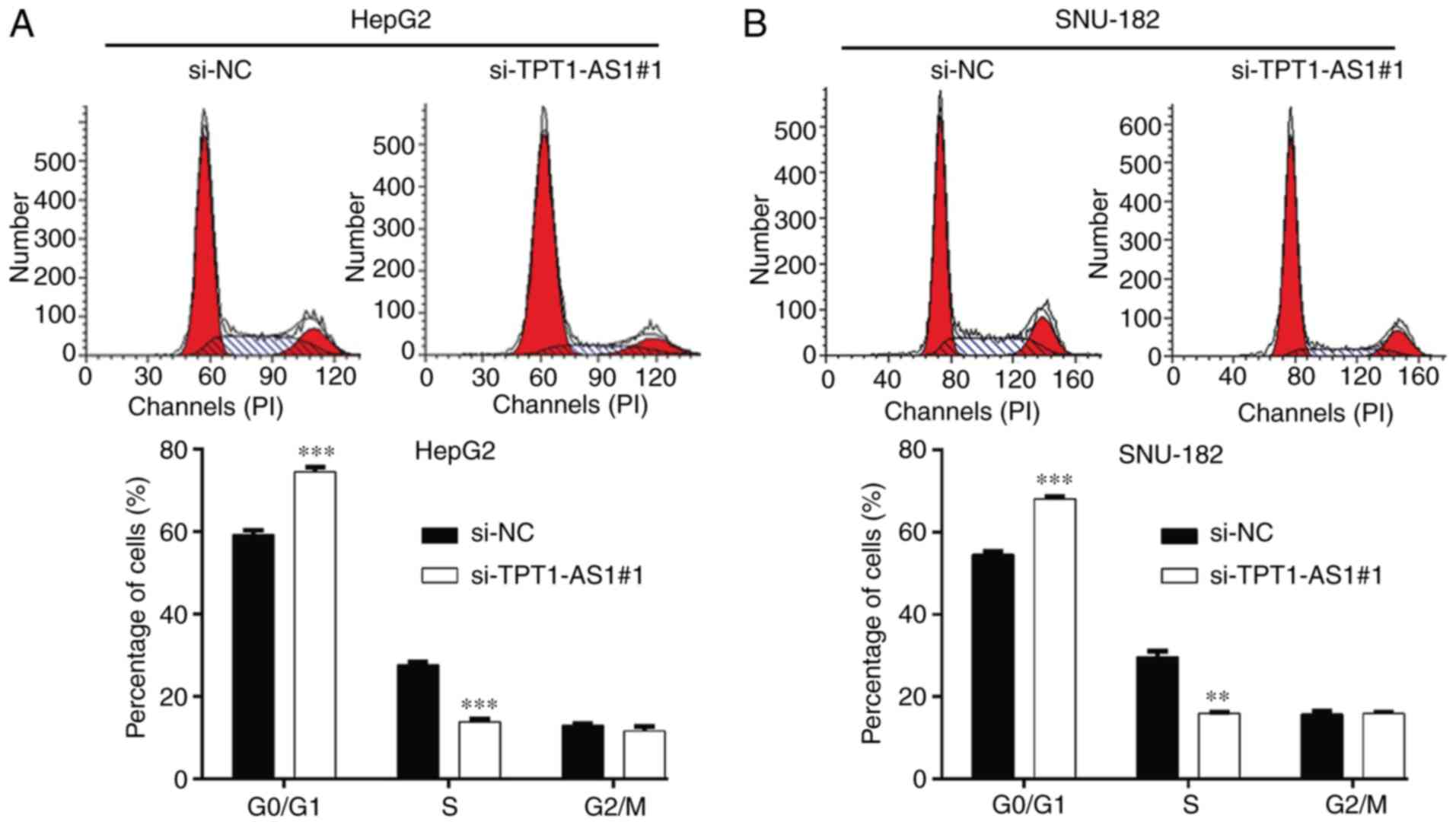
Furthermore, the effects of TPT1-AS1 knockdown on LC cell cycle
distribution were assessed. The flow cytometry results demonstrated
that compared with the si-NC group, si-TPT1-AS1#1 transfection
significantly increased the percentage of cells in the
G0/G1 phase and significantly decreased the
percentage of cells in the S phase in HepG2 (Fig. 3A) and SNU-182 (Fig. 3B) cells, which indicated that
TPT1-AS1 knockdown induced G0/G1 phase arrest
in LC cells.
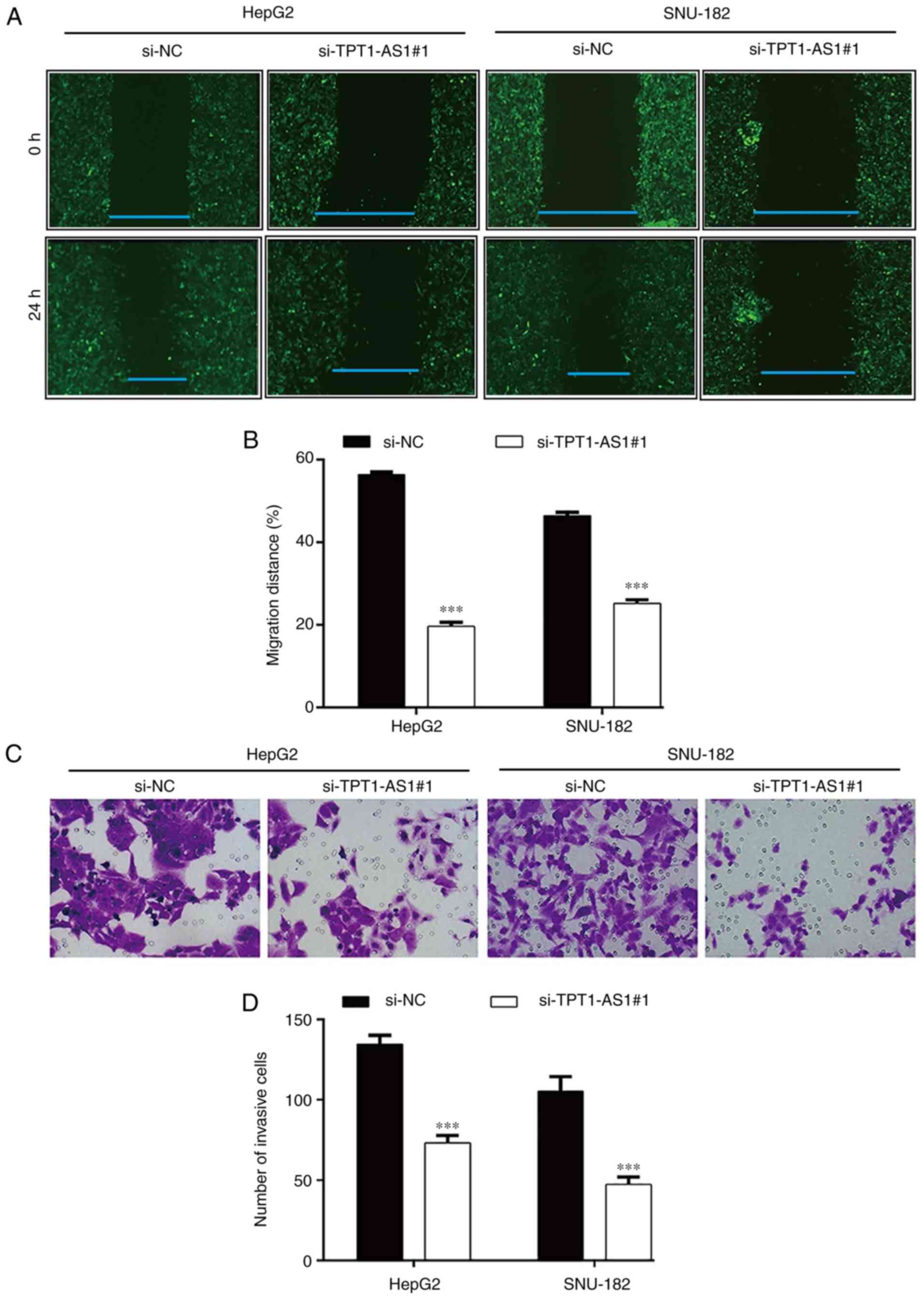
TPT1-AS1 knockdown suppresses LC cell
migration and invasion
In addition, the effects of TPT1-AS1 knockdown on LC
cell motility were assessed. The wound healing assay results
demonstrated that the relative migration rate was significantly
decreased in the si-TPT1-AS1#1 group compared with the si-NC group
in HepG2 (19.6±1.0 vs. 56.4±0.6) and SNU-182 (25.2±0.8 vs.
46.5±0.8) cells (Fig. 4A and B).
Similarly, the Transwell invasion assay demonstrated that TPT1-AS1
knockdown significantly reduced the number of invasive cells
compared with the si-NC group in HepG2 (73.3±4.5 vs. 134.7±5.5) and
SNU-182 (47.3±4.7 vs. 105.3±9.1) cells (Fig. 4C and D).
TPT1-AS1 knockdown alters the
expression of G1/S transition- and EMT-associated
markers
To further assess the suppressive role of TPT1-AS1
knockdown on LC cell cycle G1/S transition, migration
and invasion, the expression levels of G1/S transition-
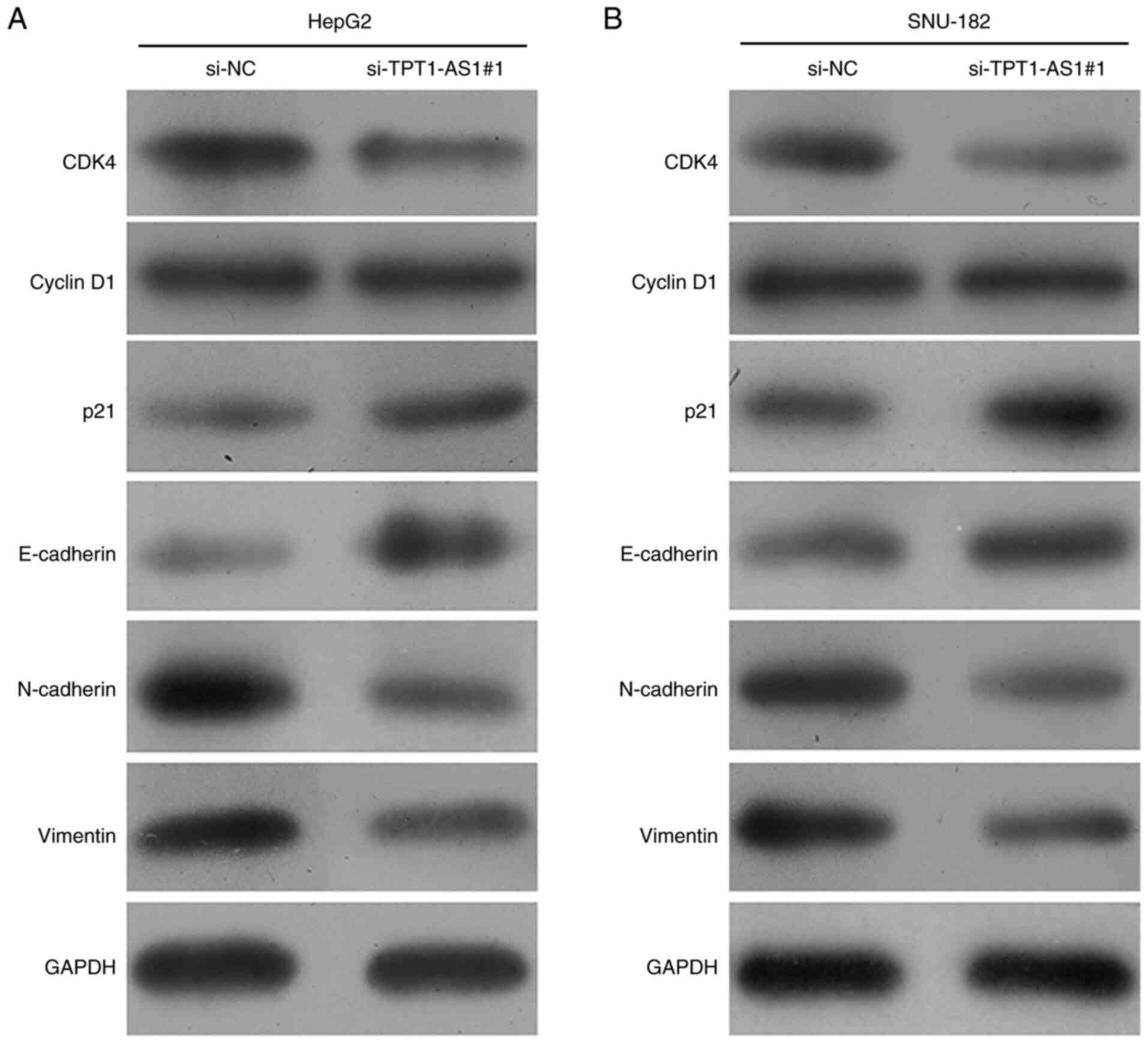
and EMT-associated markers were measured via western blotting.
Compared with the si-NC group, si-TPT1-AS1#1 transfection obviously
downregulated the expression levels of CDK4, N-cadherin and
Vimentin, and markedly upregulated the expression levels of p21 and
E-cadherin, but did not obviously alter the expression levels of
Cyclin D1 in HepG2 cells (Fig. 5A).
Similar effects of TPT1-AS1 knockdown on the expression levels of
the aforementioned protein markers were also observed in SNU-182
cells (Fig. 5B). Collectively, the
results demonstrated that TPT1-AS1 knockdown negatively regulated
LC cell proliferation, migration and invasion.
Discussion
The identification of specific and reliable
biomarkers for early-stage diagnostic modalities to improve
prognostic outcomes in LC is important, of which lncRNAs have been
highlighted as potential markers in cancer diagnosis and prognosis
(22). In the present study,
TPT1-A1 expression was significantly upregulated in LC tissues
compared with adjacent paracancerous tissues. Moreover, high
TPT1-AS1 expression was significantly associated with adverse
clinical features, including TNM stage, lymph node metastasis and
poorer prognosis in patients with LC. Consistent with the results
of the present study, ectopic TPT1-AS1 expression was strongly
associated with unfavorable clinicopathological features and poor
survival in epithelial ovarian (23), cervical (18) and gastric (19) cancer.
Subsequently, two LC cell lines (HepG2 and SNU-182)
with significantly higher TPT1-AS1 expression levels compared with
the THLE-3 cell line were selected to perform loss-of-function
experiments to assess the biological function of TPT1-AS1 in LC
in vitro. The results demonstrated that TPT1-AS1 knockdown
significantly inhibited LC cell proliferation, G1/S
transition, migration and invasion compared with the si-NC group.
Moreover, antisense lncRNAs have been reported to serve pivotal
regulatory roles in sense mRNA stability, sense-encoded protein
translation and cis-antisense RNA, and are highly expressed in
various tumors, such as breast cancer and colorectal adenocarcinoma
(24–26). In line with the results of the
present study, a variety of antisense RNAs, including MCM3AP
antisense RNA 1 (27), LIM and SH3
protein 1 antisense RNA 1 (28) and
DSCAM antisense RNA 1 (29), have
been reported to display oncogenic roles in LC. Moreover, TPT1-AS1
promoted cell proliferation, migration and invasion in cervical
cancer in vitro and in vivo (17). Wu et al (23) reported that TPT1-A1 overexpression
remarkably induced cell proliferation, migration and invasion,
whereas TPT1-AS1 knockdown results in the opposite effects in
epithelial ovarian cancer. Additionally, TPT1-AS1 knockdown
significantly inhibited gastric cancer cell proliferation, cell
cycle G1/S transition, migration and invasion (19). The results indicated that TPT1-AS1
displayed oncogenic effects, thus served a critical role in LC
progression.
In the present study, the possible molecular
mechanisms underlying TPT1-AS1 knockdown-mediated effects on LC
cell malignant behaviors were explored by performing western
blotting. Compared with the si-NC group, TPT1-AS1 knockdown
decreased CDK4 expression, but increased p21 expression in LC
cells. CDK4 and CDK inhibitor p21 serve an important role in cell
cycle G1/S transition, and have been investigated in
tumor cell proliferation (30,31).
Moreover, Wang et al (32)
demonstrated that cell adhesion molecule 1 antisense RNA 1 induced
G0/G1 phase arrest by decreasing CDK4 and
enhancing p21 expression. Consistently, Tang et al (19) indicated that TPT1-AS1 knockdown
downregulated the expression of CDK4 and upregulated the expression
of p21, negatively regulating G1/S transition and
proliferation in gastric cancer cells. According to the close
association between uncontrolled proliferation and cell cycle
dysfunction, it was hypothesized that TPT1-AS1 knockdown suppressed
LC cell proliferation by inducing G0/G1 phase
arrest via regulating CDK4/p21 expression. EMT is a pivotal
mechanism contributing to cancer invasion and metastasis, whereby
epithelial cells lose their polarity and acquire the migratory
properties of mesenchymal cells (33). In the present study, the suppressive
effects of TPT1-AS1 knockdown on EMT were observed in LC cells, as
evidenced by increased E-cadherin expression, and decreased
N-cadherin and Vimentin expression in the si-TPT1-AS1#1 group
compared with the si-NC group. In recent years, it has been
reported that knockdown of antisense lncRNAs, including HOXA
cluster antisense RNA 2 (34) and
SBF2 antisense RNA 1 (35),
suppressed LC cell migration and invasion by modulating EMT
ability. Interestingly, the effects of TPT1-AS1 knockdown on
E-cadherin and Vimentin have also been reported in gastric cancer
cells, resulting in decreased cell migration and invasion in
vitro (19). The aforementioned
results strongly supported the suppressive role of TPT1-AS1 on LC
cell proliferation and metastasis. Additionally, the present study
had a number of limitations, including the lack of overexpression
experiments, and recurrence rate and disease-free survival
analyses, which require further investigation in future
studies.
In summary, the present study demonstrated that
TPT1-AS1 may serve as an important predictor in the clinical
outcomes of patients with LC. Functionally, TPT1-AS1 facilitated LC
cell proliferation, migration and invasion by affecting
G1/S transition and EMT-associated markers.
Collectively, the results of the present study suggested that
TPT1-AS1 may serve as a promising therapeutic target for LC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HL drafted the work and performed the experiments.
JJ was responsible for the conception and design of the present
study, and gave final approval of the work. JX researched the
literature and performed the experiments. WW performed data
analysis and interpretation. All authors have read and approved the
final manuscript. HL and WW confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
The present study was performed in accordance with
the Declaration of Helsinki and approved by the Ethical Research
Committee of Taizhou People's Hospital (approval no. XCV/20190842;
Taizhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Qin G, Dang M, Gao H, Wang H, Luo F and
Chen R: Deciphering the protein-protein interaction network
regulating hepatocellular carcinoma metastasis. Biochim Biophys
Acta Proteins Proteom. 1865:1114–1122. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
El-Serag HB and Rudolph KL: Hepatocellular
carcinoma: Epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Carr BI: Hepatocellular carcinoma: Current
management and future trends. Gastroenterology. 127 (Suppl
1):S218–S224. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Llovet JM: Updated treatment approach to
hepatocellular carcinoma. J Gastroenterol. 40:225–235. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lopez PM, Villanueva A and Llovet JM:
Systematic review: Evidence-based management of hepatocellular
carcinoma-an updated analysis of randomized controlled trials.
Aliment Pharmacol Ther. 23:1535–1547. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chu C, Spitale RC and Chang HY:
Technologies to probe functions and mechanisms of long noncoding
RNAs. Nat Struct Mol Biol. 22:29–35. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kopp F and Mendell JT: Functional
classification and experimental dissection of long noncoding RNAs.
Cell. 172:393–407. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen J, Wang R, Zhang K and Chen LB: Long
non-coding RNAs in non-small cell lung cancer as biomarkers and
therapeutic targets. J Cell Mol Med. 18:2425–2436. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang Y, Chen L, Gu J, Zhang H, Yuan J,
Lian Q, Lv G, Wang S, Wu Y, Yang YT, et al: Recurrently deregulated
lncRNAs in hepatocellular carcinoma. Nat Commun. 8:144212017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu S, Liu J, Wang X, Li M, Chen Z and Tang
Y: Aberrant expression of the long non-coding RNA GHRLOS and its
prognostic significance in patients with colorectal cancer. J
Cancer. 8:4040–4047. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jiang H, Shi X, Ye G, Xu Y, Xu J, Lu J and
Lu W: Up-regulated long non-coding RNA DUXAP8 promotes cell growth
through repressing Krüppel-like factor 2 expression in human
hepatocellular carcinoma. Onco Targets Ther. 12:7429–7436. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zheng Y, Nie P and Xu S: Long noncoding
RNA linc00467 plays an oncogenic role in hepatocellular carcinoma
by regulating the miR-18a-5p/NEDD9 axis. J Cell Biochem.
121:3135–3144. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Deng Y, Wei Z, Huang M, Xu G, Wei W, Peng
B, Nong S and Qin H: Long non-coding RNA F11-AS1 inhibits
HBV-related hepatocellular carcinoma progression by regulating
NR1I3 via binding to microRNA-211-5p. J Cell Mol Med. 24:1848–1865.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang W, Yang F, Zhang L, Chen J, Zhao Z,
Wang H, Wu F, Liang T, Yan X, Li J, et al: LncRNA profile study
reveals four-lncRNA signature associated with the prognosis of
patients with anaplastic gliomas. Oncotarget. 7:77225–77236. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zan XY and Li L: Construction of
lncRNA-mediated ceRNA network to reveal clinically relevant lncRNA
biomarkers in glioblastomas. Oncol Lett. 17:4369–4374.
2019.PubMed/NCBI
|
|
17
|
Jiang H, Huang G, Zhao N, Zhang T, Jiang
M, He Y, Zhou X and Jiang X: Long non-coding RNA TPT1-AS1 promotes
cell growth and metastasis in cervical cancer via acting AS a
sponge for miR-324-5p. J Exp Clin Cancer Res. 37:1692018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jia L, Song Y, Mu L, Li Q, Tang J, Yang Z
and Meng W: Long noncoding RNA TPT1-AS1 downregulates the
microRNA-770-5p expression to inhibit glioma cell autophagy and
promote proliferation through STMN1 upregulation. J Cell Physiol.
235:3679–3689. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tang J, Huang F, Wang H, Cheng F, Pi Y,
Zhao J and Li Z: Knockdown of TPT1-AS1 inhibits cell proliferation,
cell cycle G1/S transition, and epithelial-mesenchymal transition
in gastric cancer. Bosn J Basic Med Sci. 21:39–46. 2021.PubMed/NCBI
|
|
20
|
Mirsadraee S, Oswal D, Alizadeh Y, Caulo A
and van Beek E Jr: The 7th lung cancer TNM classification and
staging system: Review of the changes and implications. World J
Radiol. 4:128–134. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang ZL, Li W, Chen QF, Wu PH and Shen
LJ: Eight key long non-coding RNAs predict hepatitis virus positive
hepatocellular carcinoma as prognostic targets. World J
Gastrointest Oncol. 11:983–997. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu W, Gao H, Li X, Zhu Y, Peng S, Yu J,
Zhan G, Wang J, Liu N and Guo X: LncRNA TPT1-AS1 promotes
tumorigenesis and metastasis in epithelial ovarian cancer by
inducing TPT1 expression. Cancer Sci. 110:1587–1598. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
He Y, Vogelstein B, Velculescu VE,
Papadopoulos N and Kinzler KW: The antisense transcriptomes of
human cells. Science. 322:1855–1857. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Munroe SH and Zhu J: Overlapping
transcripts, double-stranded RNA and antisense regulation: A
genomic perspective. Cell Mol Life Sci. 63:2102–1218. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Faghihi MA and Wahlestedt C: Regulatory
roles of natural antisense transcripts. Nat Rev Mol Cell Biol.
10:637–643. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Y, Yang L, Chen T, Liu X, Guo Y, Zhu
Q, Tong X, Yang W, Xu Q, Huang D and Tu K: A novel lncRNA
MCM3AP-AS1 promotes the growth of hepatocellular carcinoma by
targeting miR-194-5p/FOXA1 axis. Mol Cancer. 18:282019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yin L, Chen Y, Zhou Y, Deng G, Han Y, Guo
C, Li Y, Zeng S and Shen H: Increased long noncoding RNA LASP1-AS
is critical for hepatocellular carcinoma tumorigenesis via
upregulating LASP1. J Cell Physiol. 234:13493–13509. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ji D, Hu G, Zhang X, Yu T and Yang J: Long
non-coding RNA DSCAM-AS1 accelerates the progression of
hepatocellular carcinoma via sponging miR-338-3p. Am J Transl Res.
11:4290–4302. 2019.PubMed/NCBI
|
|
30
|
Ye D, Luo H, Lai Z, Zou L, Zhu L, Mao J,
Jacob T, Ye W, Wang L and Chen L: ClC-3 chloride channel proteins
regulate the cell cycle by up-regulating cyclin D1-CDK4/6 through
suppressing p21/p27 expression in nasopharyngeal carcinoma cells.
Sci Rep. 6:302762016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu J, Min L, Zhu S, Guo Q, Li H, Zhang Z,
Zhao Y, Xu C and Zhang S: Cyclin-dependent kinase inhibitor 3
promoted cell proliferation by driving cell cycle from G1 to S
phase in esophageal squamous cell carcinoma. J Cancer.
10:1915–1922. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang F, Qi X, Li Z, Jin S, Xie Y and Zhong
H: lncRNA CADM1-AS1 inhibits cell-cycle progression and invasion
via PTEN/AKT/GSK-3β axis in hepatocellular carcinoma. Cancer Manag
Res. 11:3813–3828. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ma C, Huang S, Xu L, Tian L, Yang Y and
Wang J: Transcription co-activator P300 activates Elk1-aPKC-ι
signaling mediated epithelial-to-mesenchymal transition and
malignancy in hepatocellular carcinoma. Oncogenesis. 9:322020.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang Y, Xu J, Zhang S, An J, Zhang J,
Huang J and Jin Y: HOXA-AS2 promotes proliferation and induces
epithelial-mesenchymal transition via the miR-520c-3p/GPC3 axis in
hepatocellular carcinoma. Cell Physiol Biochem. 50:2124–2138. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li Y, Liu G, Li X, Dong H, Xiao W and Lu
S: Long non-coding RNA SBF2-AS1 promotes hepatocellular carcinoma
progression through regulation of miR-140-5p-TGFBR1 pathway.
Biochem Biophys Res Commun. 503:2826–2832. 2018. View Article : Google Scholar : PubMed/NCBI
|