Introduction
Acute myeloid leukemia (AML) is characterized by
blocked myeloid differentiation and abnormal proliferation of
immature myeloid cells in the bone marrow and peripheral blood
(1,2). The current clinical treatments for AML
include induced differentiation therapy, combined chemotherapy,
hematopoietic stem cell transplantation and immunotherapy (3–6).
Although progress has been made in medicine and chemotherapy, the
treatment outcomes of AML are not ideal. Only 35–40% of young (age
<60 years) and 5–15% of elderly patients with AML (age ≥60
years) survive for >5 years post-treatment (7). The primary reason for therapeutic
failure and poor prognosis is the chemotherapy resistance of
leukemia cells (8,9). Therefore, there is an urgent need to
find more effective drugs and therapies for treating AML.
Corilagin is the primary active ingredient of the
Euphorbia phyllanthus plant and has antiviral,
anti-inflammatory, antioxidant and other biological activity
(10–12). Corilagin can effectively diminish
inflammation in ulcerative colitis and cystic fibrosis bronchial
cells. By blocking the activation of nuclear factor κB (NF-κB),
corilagin decreases expression of interleukin (IL)-1β, IL-6, TNF-α
and other inflammatory factors (13,14).
Previous studies have shown that corilagin has a significant
inhibitory effect on glioblastoma, as well as breast and ovarian
cancer (15–17). Corilagin inhibits the proliferation
of breast cancer cells by inducing apoptosis and autophagy and that
of ovarian cancer cells by blocking the typical Smad and atypical
ERK/AKT pathways (16,17). Although corilagin possesses
anti-inflammatory and antitumor effects, to the best of our
knowledge, its function in inhibiting leukemia has rarely been
reported.
MicroRNA (miRNAs/miRs) are a class of small,
non-coding RNAs with 19–25 nucleotides (nts) that negatively
regulate target gene expression at the post-transcriptional level.
Abnormal expression of miRNAs is associated with a variety of human
diseases, including AML (18–20).
Modulation of dysregulated miRNAs is used in AML therapy and
sensitizes AML cells to chemotherapy (21,22).
Overexpression of miR-34a, which is expressed at low levels in AML,
promotes apoptosis and inhibits autophagy in AML cells (23). Upregulation of miR-142-3p improves
the drug sensitivity of AML cells by inhibiting cell viability and
promoting apoptosis (24).
Additionally, miR-451 is abnormally downregulated in patients with
AML, and miR-451 acts as a tumor suppressor by repressing malignant
proliferation and inducing apoptosis (25,26).
Upregulation of miR-451 can prevent cardiomyocyte
anoxia/reoxygenation (A/R) injury by suppressing high mobility
group protein B1 (HMGB1) (27).
However, the association between miR-451 and HMGB1 in AML remains
unclear.
HMGB1 is a highly conserved and widely expressed
non-histone, chromatin-binding protein, which can bind with
specific chromatin DNA and participate in DNA recombination, gene
transcription regulation, cell division, and differentiation
(28,29). Under metabolic stress, HMGB1
transfers from the nucleus to the cytoplasm and binds Beclin-1 to
prevent its aggregation, thus promoting autophagy and inhibiting
apoptosis (30). HMGB1 can induce
chemotherapy resistance by promoting autophagy in lung and
colorectal cancer, as well as other types of tumor (31,32).
HMGB1 levels in the serum of patients with AML increase
significantly following chemotherapy, and inhibition of HMGB1 can
improve the sensitivity of AML cells to chemotherapy drugs
(24,33). Therefore, HMGB1 may be an important
clinical target for the treatment of leukemia.
In the present study, the effects of
corilagin-induced proliferation inhibition were investigated in the
AML cell line HL-60. The effect of corilagin on apoptosis and
autophagy in AML cells was investigated. The expression levels of
miR-451 and HMGB1 were evaluated following corilagin treatment.
Finally, the role of miR-451 in corilagin-induced proliferation
inhibition on HL-60 cells was assessed. The present study aimed to
elucidate the effect and potential mechanism of corilagin on AML
cells, which may provide a promising treatment for AML.
Materials and methods
Cell culture and treatment
The human promyelocytic leukemia cell line HL-60,
obtained from the American Type Culture Collection, was maintained
in DMEM supplemented with 10% FBS and 100 U/ml
penicillin/streptomycin at 37°C under 5% CO2. HL-60
cells were treated with PBS or increasing concentrations of
corilagin (25 and 50 µg/ml) in a 37°C incubator for different
durations (0, 1, 2, 3, 4 and 5 days).
Cell transfection
HL-60 cells were seeded in 6-well plates
(1×106/well) before transfection. A total of 10 µl
diluted transfection reagent DharmaFECT (Tube 1) and 100 nM
negative control (NC) inhibitor or miR-451 inhibitor (Tube 2) were
mixed with 190 µl serum-free DMEM for 5 min at room temperature.
The contents of Tubes 1 and 2 were mixed and incubated for 20 min
at room temperature, then 1.6 ml antibiotic-free DMEM was added for
a total volume of 2 ml transfection medium. Culture medium was
replaced with transfection medium in each well. The NC inhibitor
and miR-451 inhibitor were synthesized by Shanghai GenePharma Co.,
Ltd. Sequence of NC inhibitor was 5′-CAGUACUUUUGUGUAGUACAA-3′;
sequence of miR-451 inhibitor was 5′-AACUCAGUAAUGGUAACGGUUU-3′.
Following transfection at 37°C for 72 h, cells were harvested for
further experiments.
Reagents and antibodies
Corilagin (purity ≥98%) was purchased from Shanghai
Yuanye Biotechnology Co., Ltd. DMEM, FBS and
penicillin/streptomycin were purchased from Gibco (Thermo Fisher
Scientific, Inc.). Cell Counting Kit-8 (CCK-8) and
Carboxyfluorescein Diacetate Succinimidyl Ester (CFSE) cell
proliferation kits were obtained from Dongren Chemical Technology
(Shanghai) Co., Ltd.. Annexin V-APC/7-AAD apoptosis detection kit
was from BioLegend, Inc., BSA, RIPA lysis buffer and BCA protein
quantification kit were purchased from Beyotime Institute of
Biotechnology. Polyvinylidene difluoride (PVDF) membrane and ECL
reagent were obtained from MilliporeSigma. Primary antibodies (all
1:1,000) against light chain (LC)3 (cat. no. 12741),
autophagy-related 5 (Atg5; cat. no. 12994), pro- and cleaved
caspase-3 (cat. no. 14220), Bcl-xl (cat. no. 2764) and Bax (cat.
no. 5023) were obtained from Cell Signalling Technology, Inc.; p62
(cat. no. 18420-1-AP, 1:1,000), Beclin-1 (cat. no. 11306-1-AP,
1:1000), β-actin (cat. no. 20536-1-AP, 1:5,000), HMGB1 (cat. no.
10829-1-AP, 1:1,000) primary and horseradish peroxidase
(HRP)-conjugated secondary antibodies (both 1:5,000; goat
anti-mouse IgG, cat. no. SA00001-1; goat anti-rabbit IgG, cat. no.
SA00001-2) were from ProteinTech Group, Inc. TRIzol®,
Alexa Fluor488-conjugated goat anti-rabbit secondary antibody (cat.
no. A-11008; 1:500) and anti-fade mounting medium with DAPI were
purchased from Invitrogen (Thermo Fisher Scientific, Inc.).
DharmaFECT transfection reagent and RevertAid Reverse Transcription
(RT) kit were obtained from Thermo Fisher Scientific, Inc.
UltraSYBR Mixture was from CWBio.
Cell viability assay
HL-60 cells were seeded in 96-well plates at a
density of 2×104 cells/well and cultured overnight in a
37°C incubator. The cells were treated with PBS or increasing
concentrations of corilagin (25 and 50 µg/ml) for different
durations (0, 1, 2, 3, 4 and 5 days). CCK-8 was added to each well
and the cells were incubated for 4–6 h. The absorbance was measured
at 450 nm using a microplate reader. Cell viability was calculated
as follows: Mean optical density (OD) of treated wells-mean OD of
blank wells/mean OD of control wells-mean OD of blank wells.
Flow cytometry
HL-60 cells were treated with PBS or increasing
concentrations of corilagin (25 and 50 µg/ml) at 37°C for 48 h,
collected and washed with PBS three times. For the cell
proliferation assay, cells were incubated with 10 µM CFSE for 10
min at room temperature, then heat-inactivated FBS was added to
terminate the labelling reaction. The labelled cells were washed
twice, resuspended in PBS and analyzed by flow cytometry
(FACSCalibur; BD Biosciences). For the cell apoptosis assay, cells
were stained with 5 µl Annexin V-APC and 7-AAD for 15 min at room
temperature, then assessed by flow cytometry (CytoFlex; Beckman
Coulter, Inc.). The percentage of early- and late-stage apoptotic
cells was calculated using FlowJo v10 software (BD
Biosciences).
Small RNA single-end sequencing
Total RNA was extracted from cells using TRIzol
according to the manufacturers' instructions. The integrity, purity
and concentration of RNA were detected by Agilent 2100, Nanodrop
and Qubit, respectively. After the samples passed quality control
tests, the Small RNA Sample Prep kit (Illumina, Inc.) was used to
produce cDNA libraries. Briefly, adapters were added to the 5′ and
3′ ends of small RNA, followed by RT-quantitative (q)PCR
amplification and PAGE to separate the target DNA fragments. After
the library was built, the insert size was detected using Agilent
2100, and the effective concentration (>2 nM) was quantified by
qPCR. The cDNA libraries were sequenced using an Illumina HiSeq
2000 platform and 50 bp single-end reads were generated.
Subsequently, the raw reads were processed and filtered for the
length of 21–22 nt to obtain miRNA reads. The miRNAs were aligned
with miRBase (https://www.mirbase.org) and
annotated using Bowtie (http://bowtie-bio.sourceforge.net). To identify the
differentially expressed miRNAs, the expression levels of miRNAs
were normalized using Transcripts Per Million. Normalized
expression was determined as follows: Read count of miRNA
×1,000,000)/read count of total miRNAs.
The criteria for identifying differentially
expressed miRNAs were P<0.01 and log2(fold change)>1.
Western blotting
HL-60 cells were lysed with RIPA buffer for 30 min
on ice and supernatant was collected by centrifugation at 12,000 ×
g for 15 min at 4°C. Protein concentrations were determined using a
BCA protein quantification kit. Protein (30 µg/lane) was loaded,
separated via 12% SDS-PAGE (Bio-Rad Laboratories, Inc.) and
transferred onto PVDF membranes. After blocking with 3% BSA for 1 h
at room temperature, the membranes were incubated with primary
antibodies overnight at 4°C and with HRP-conjugated secondary
antibodies for 1 h at room temperature. Finally, protein bands were
exposed to ECL reagent and visualized using a chemiluminescent
detector (Tanon Science and Technology Co., Ltd.). The relative
protein expression was semi-quantified using ImageJ v.1.46r
software (National Institutes of Health) using β-actin as the
control.
Immunofluorescence
Cells were fixed with 4% paraformaldehyde for 10 min
at room temperature and washed three times with PBS. After blocking
with 10% FBS containing 0.1% Triton for 20 min at room temperature,
cells were incubated with LC3 primary antibody at 4°C overnight and
subsequently stained with 488-conjugated anti-rabbit secondary
antibody at room temperature for 1 h. Finally, cells were washed
three times with PBS and mounted on glass slides using anti-fade
mounting medium with DAPI. The stained cells were imaged using a
confocal microscope (Zeiss LSM800; Zeiss AG; magnification, ×630).
The number of LC3-positive puncta/cell was quantified using ImageJ
v.1.46r software (National Institutes of Health).
RT-qPCR
Total RNA was extracted from HL-60 cells using
TRIzol and reverse transcribed to cDNA using an RT kit according to
the manufacturer's protocol. RT-qPCR was performed using an
UltraSYBR Mixture in a CFX-96 Touch Real-Time PCR Detection system
(Bio-Rad Laboratories, Inc.). The thermocycling conditions were as
follows: 95°C for 5 min, followed by 40 cycles of 95°C for 10 sec
and 60°C for 30 sec. The expression level of HMGB1 was normalized
to that of the reference gene β-actin, and miR-451 to that of U6.
The relative expression levels of genes were calculated using the
2−ΔΔCq method (34). The
primers sequences are shown in Table
I.
 | Table I.Primer sequences for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences for reverse
transcription-quantitative PCR.
| Gene | Forward, 5′→3′ | Reverse, 5′→3′ |
|---|
| miR-451 |
AAACCGTTACCATTACTGAGTT |
AACTCAGTAATGGTAACGGTTT |
| U6 |
CTCGCTTCGGCAGCACATATACT |
ACGCTTCACGAATTTGCGTGTC |
| HMGB1 |
TGGTATTTTGGACTGCGGGG |
TGACATTTTGCCTCTCGGCT |
| GAPDH |
TGAAGGTCGGAGTCAACGG |
TCCTGGAAGATGGTGATGGGA |
Survival analysis of patients with AML
with differentially expressed HMGB1 by Gene Expression Profiling
Interactive Analysis (GEPIA)
The GEPIA database (http://gepia.cancer-pku.cn) was used to analyze the
survival percentage of patients with AML with high or low levels of
HMGB1 using 106 samples from the Genotype Tissue Expression
(https://gtexportal.org) and The Cancer Genome
Atlas (http://tcga-data.nci.nih.gov/tcga).
Statistical analysis
The data are presented as the mean ± SD from three
independent experiments. Data were compared using an unpaired
Student's t-test or one-way ANOVA followed by Bonferroni's post hoc
test using GraphPad Prism 5 (GraphPad Software, Inc.). P<0.05
was considered to indicate a statistically significant
difference.
Results
Corilagin inhibits proliferation of
HL-60 AML cells
The inhibitory effect of corilagin on AML cells was
analyzed using CCK-8 and CFSE assay kits. The AML cell line HL-60
was treated with PBS or increasing concentrations of corilagin (25
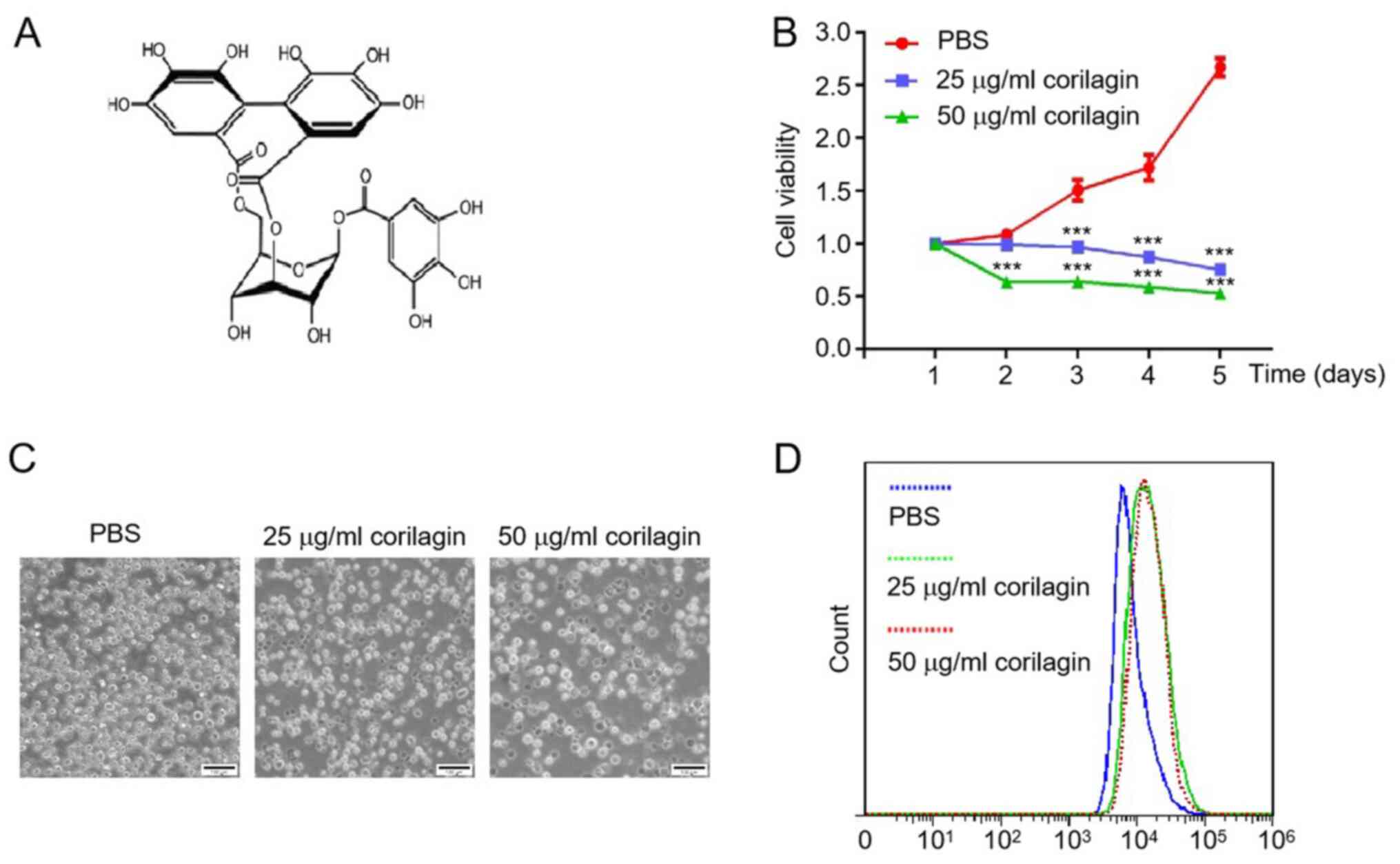
and 50 µg/ml) for different durations (Fig. 1A). Corilagin significantly inhibited
proliferation of HL-60 cells in a dose- and time-dependent manner
(Fig. 1B). The cytotoxicity of
corilagin on HL-60 cells was also evaluated by microscopy. The cell
number was notably decreased and the cell shape became irregular
following corilagin treatment for 48 h (Fig. 1C). Moreover, the fluorescence
intensity of the corilagin-treated group was higher than that of
the PBS group (Fig. 1D). These data
indicated that corilagin inhibited the proliferation of HL-60 AML
cells.
Corilagin activates apoptosis in HL-60
AML cells
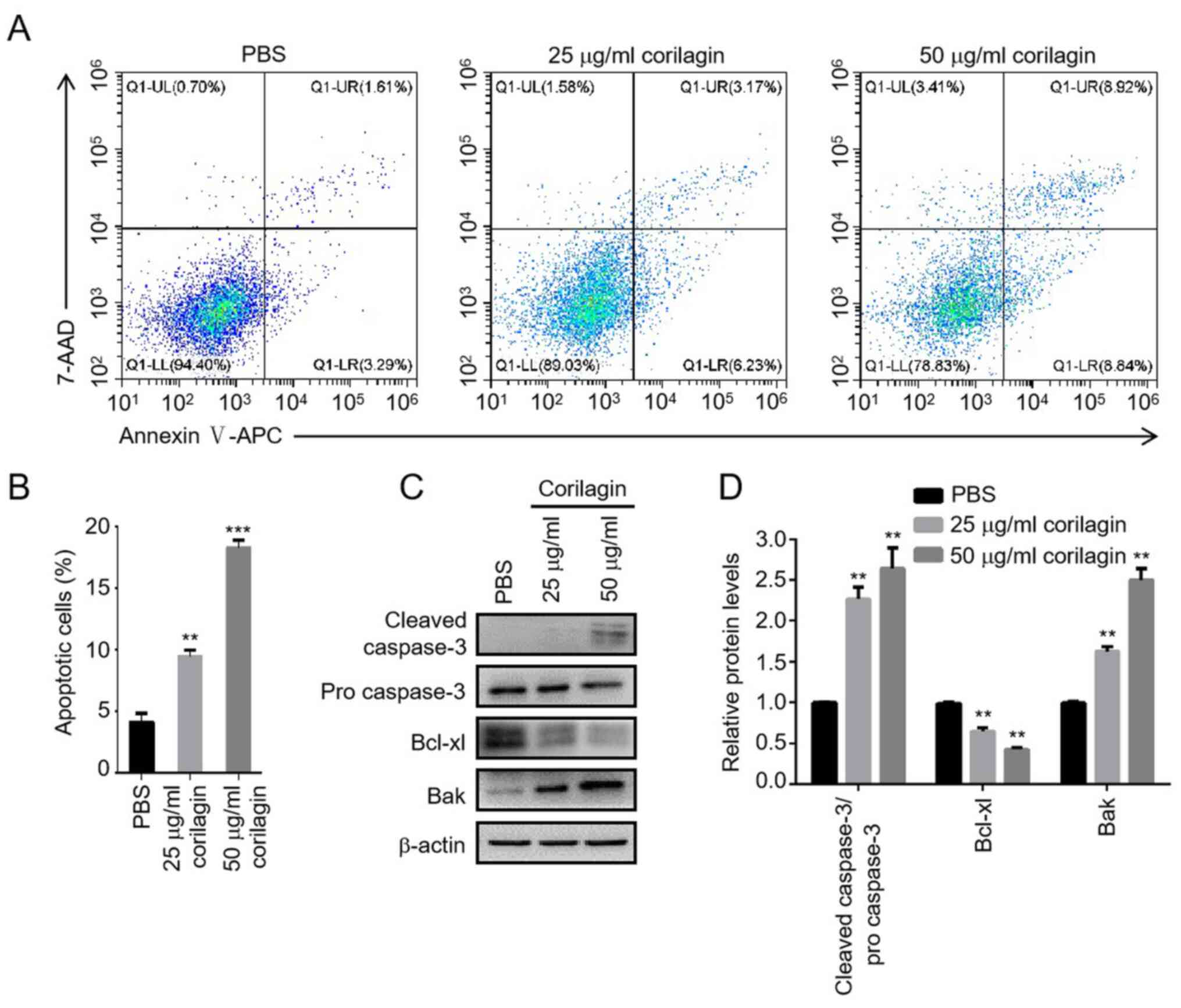
To investigate the apoptotic effect of corilagin on
AML cells, Annexin V/7-AAD staining assay was used to assess
apoptotic cell death by flow cytometry. HL-60 cells were treated
with PBS or different concentrations of corilagin (25 and 50 µg/ml)
for 48 h. The percentage of early- and late-stage apoptotic cells
increased following corilagin treatment in a dose-dependent manner
(Fig. 2A and B). To verify whether
corilagin-induced apoptosis was triggered by the intrinsic
signalling pathway, the expression levels of key proteins were
detected by western blotting. Corilagin increased the protein
levels of cleaved caspase-3 and Bak and decreased the level of
Bcl-xl, indicating that the intrinsic mitochondrial apoptosis
pathway was activated following exposure to corilagin (Fig. 2C and D).
Corilagin inhibits autophagy of HL-60
AML cells
Autophagy may function as a protective mechanism in
tumor cells, and inhibition of autophagy promotes cell apoptosis
(30). Microtubule-associated
protein LC3 is a key autophagy-associated protein involved in
autophagosome formation. Cytosolic LC3-I is converted to
membrane-bound LC3-II by conjugation to phosphatidylethanolamine
and localizes to the autophagosome (35). To determine the effect of corilagin
on AML cell autophagy, the ratio of LC3-II/I and number of
LC3-positive puncta were assessed by western blotting and
immunofluorescence, respectively. Corilagin significantly decreased
the conversion of LC3-I into LC3-II and formation of LC3-positive
puncta. In addition, the expression levels of Atg5 and Beclin-1
decreased, while p62 expression increased following corilagin
treatment. These results suggested that corilagin exhibited an
inhibitory effect on AML cell autophagy (Fig. 3A-D).
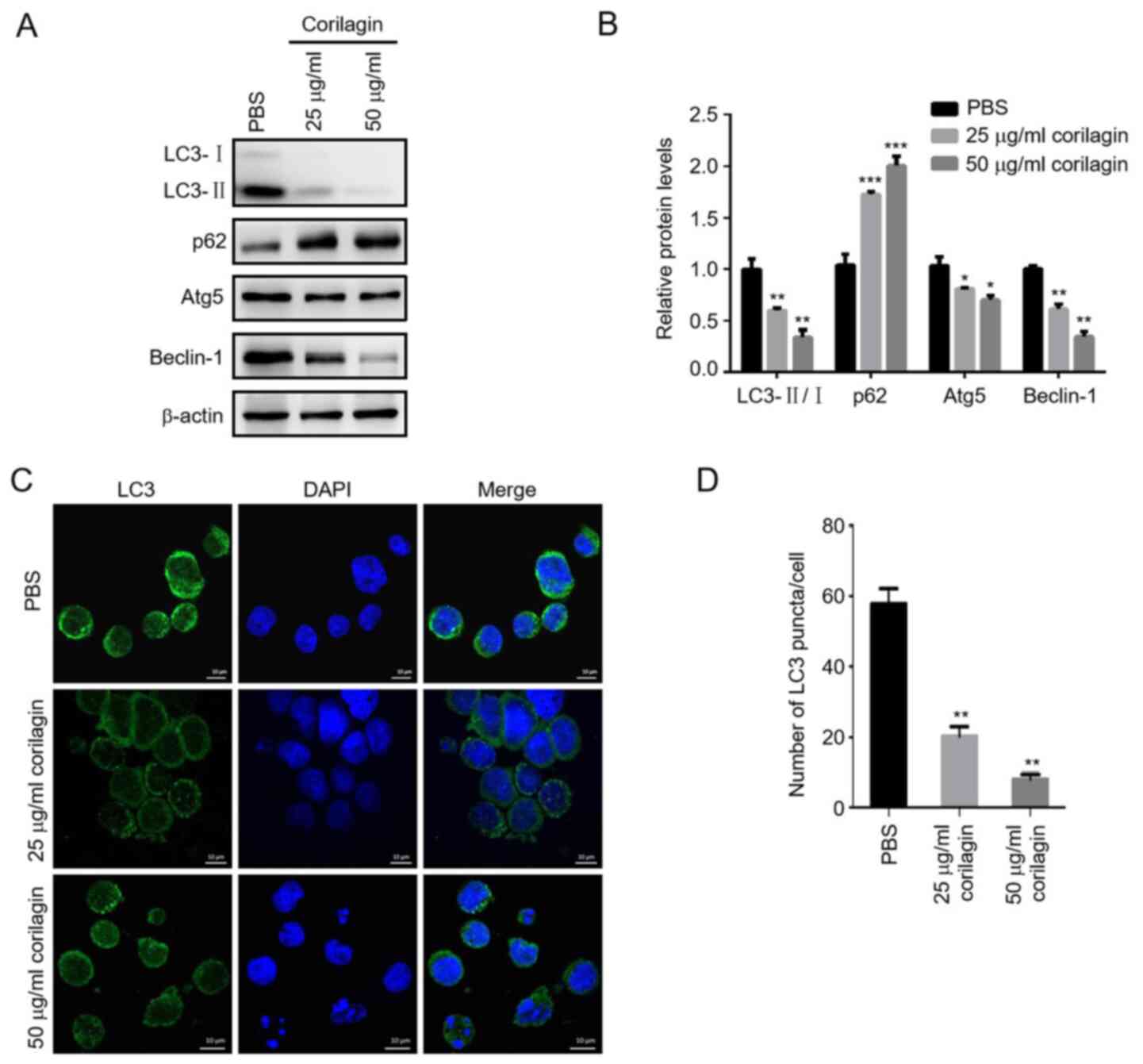 | Figure 3.Corilagin inhibits autophagy of HL-60
cells. (A) HL-60 cells were incubated with PBS or 25 and 50 µg/ml
corilagin for 48 h and expression of autophagy-associated proteins
LC3, p62, Atg5 and Beclin-1 was determined by western blotting. (B)
Relative protein expression levels of LC3-II/LC3-I, p62, Atg5 and
Beclin-1 were quantified using Image J software. (C) Confocal
images of LC3-positive puncta, representing autophagosomes. Scale
bar, 10 µm. (D) Quantification of the average number of
LC3-positive puncta per cell using Image J software. *P<0.05,
**P<0.01, ***P<0.001 vs. PBS group. LC, light chain; Atg,
autophagy-related. |
Corilagin elevates expression of tumor
suppressor miR-451
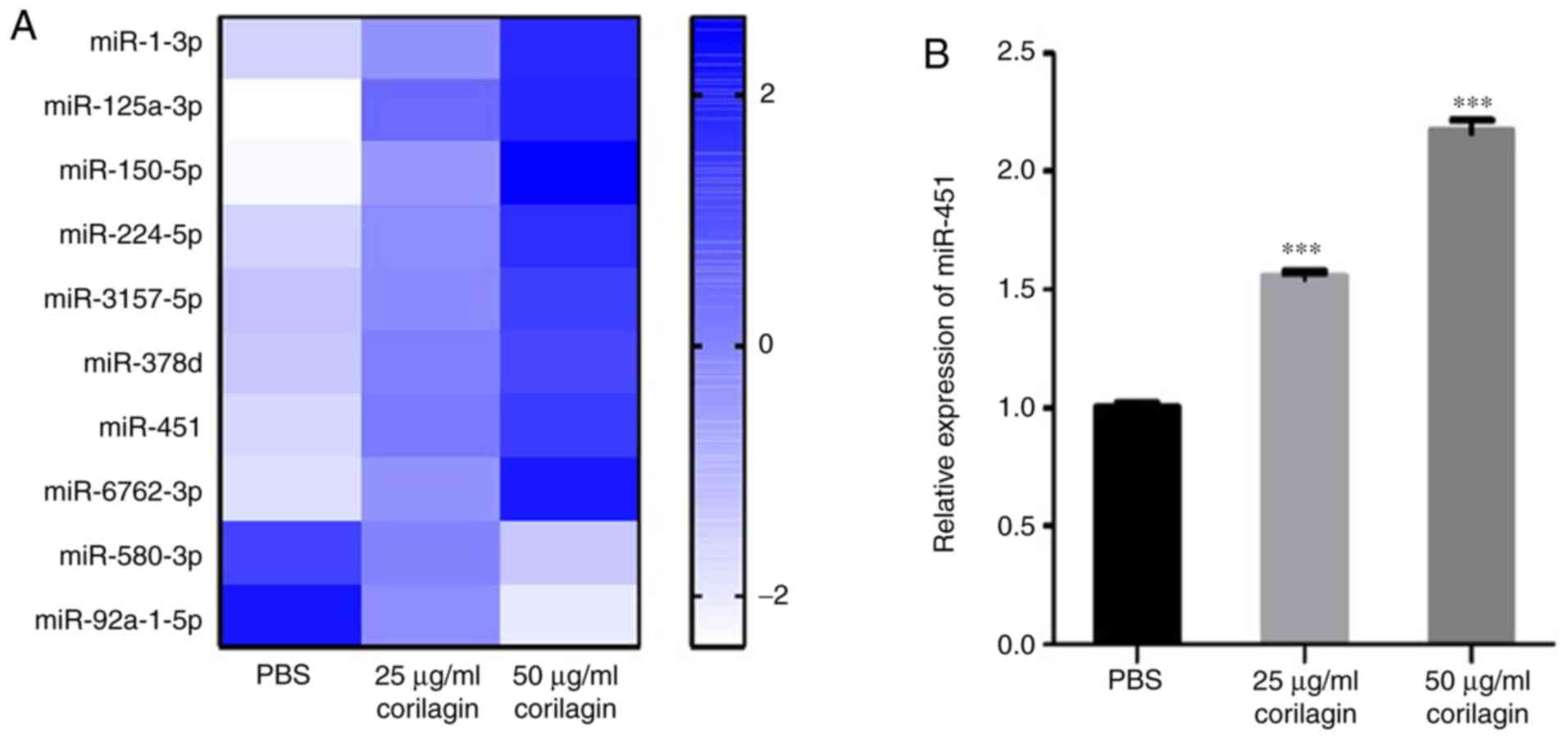
Previous studies have revealed that corilagin
prevents liver fibrosis by blocking the miR-21-regulated Smad
signalling pathway (36,37). To assess changes in miRNA expression
levels caused by corilagin, small RNA sequencing was performed.
Compared with the PBS group, eight miRNAs were notably upregulated
and two were downregulated in the corilagin-treated groups. In
addition, the expression levels of the differentially expressed
miRNAs showed a dose-dependent change when treated with corilagin
(Fig. 4A). miR-451 has been
reported to be abnormally downregulated in patients with AML and
serves as a tumor suppressor by inducing apoptosis (25,26).
The present microRNA sequencing data showed that miR-451 was
notably upregulated following exposure to corilagin. RT-qPCR
results verified the elevated expression of miR-451 following
corilagin treatment in a dose-dependent manner (Fig. 4B).
Corilagin downregulates HMGB1
expression by increasing miR-451
To determine the mechanisms by which miR-451
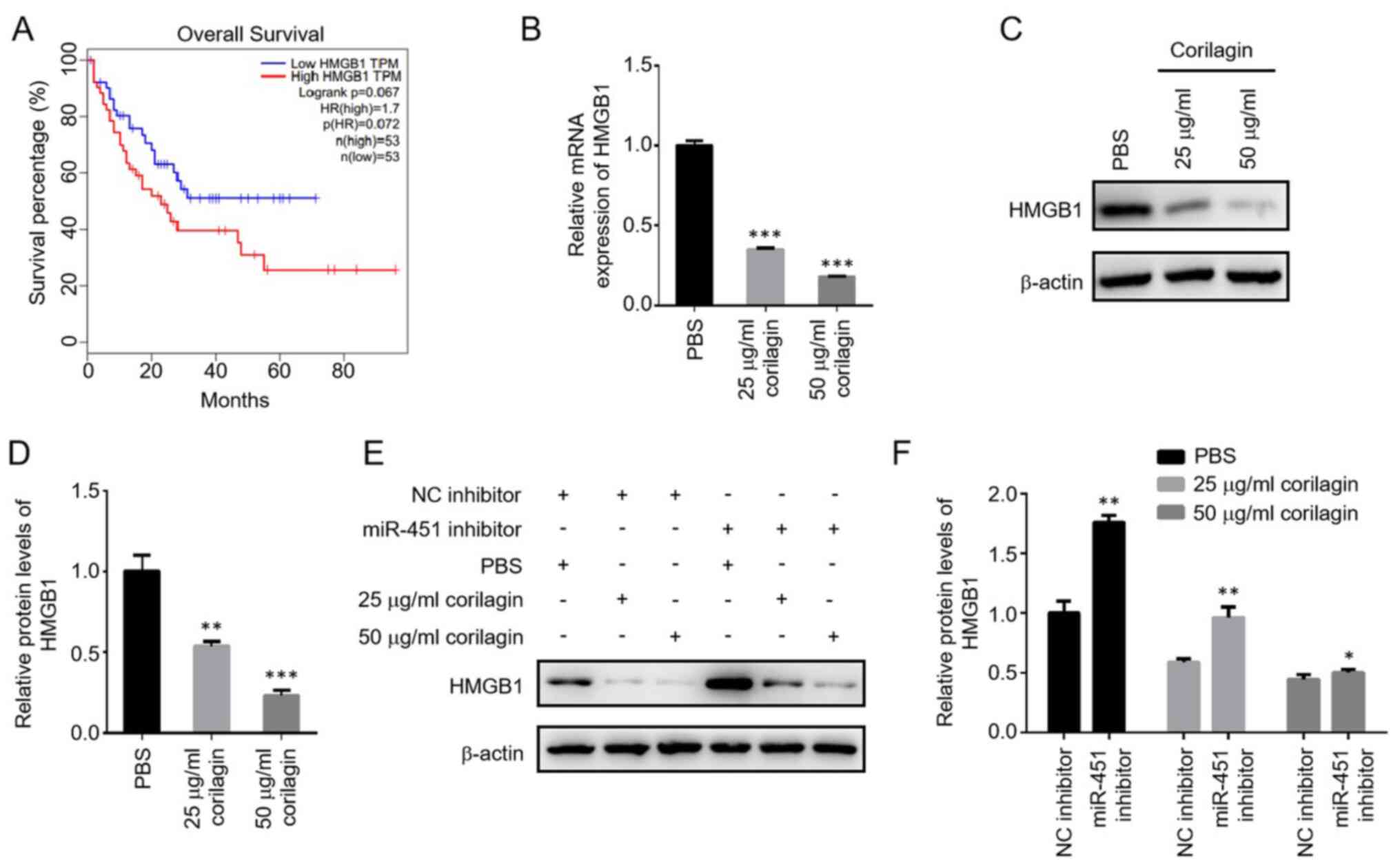
regulates AML, target genes of miR-451 were screened. A previous
study reported that the expression of HMGB1 was negatively
regulated by miR-451 during cardiomyocyte A/R injury (27). The GEPIA database showed that HMGB1
levels were negatively associated with survival of patients with
AML. The low HMGB1 expression group had a higher survival rate than
the high HMGB1 expression group (Fig.
5A). To test whether miR-451 functions by regulating HMGB1
during corilagin treatment, the expression of HMGB1 was measured
using RT-qPCR and western blotting. The mRNA and protein levels of
HMGB1 were significantly decreased in response to corilagin in a
dose-dependent manner (Fig. 5B-D).
Knockdown of miR-451 decreased the corilagin-induced downregulation
of HMGB1, indicating a negative regulation of HMGB1 by miR-451
during corilagin treatment (Fig. 5E and
F).
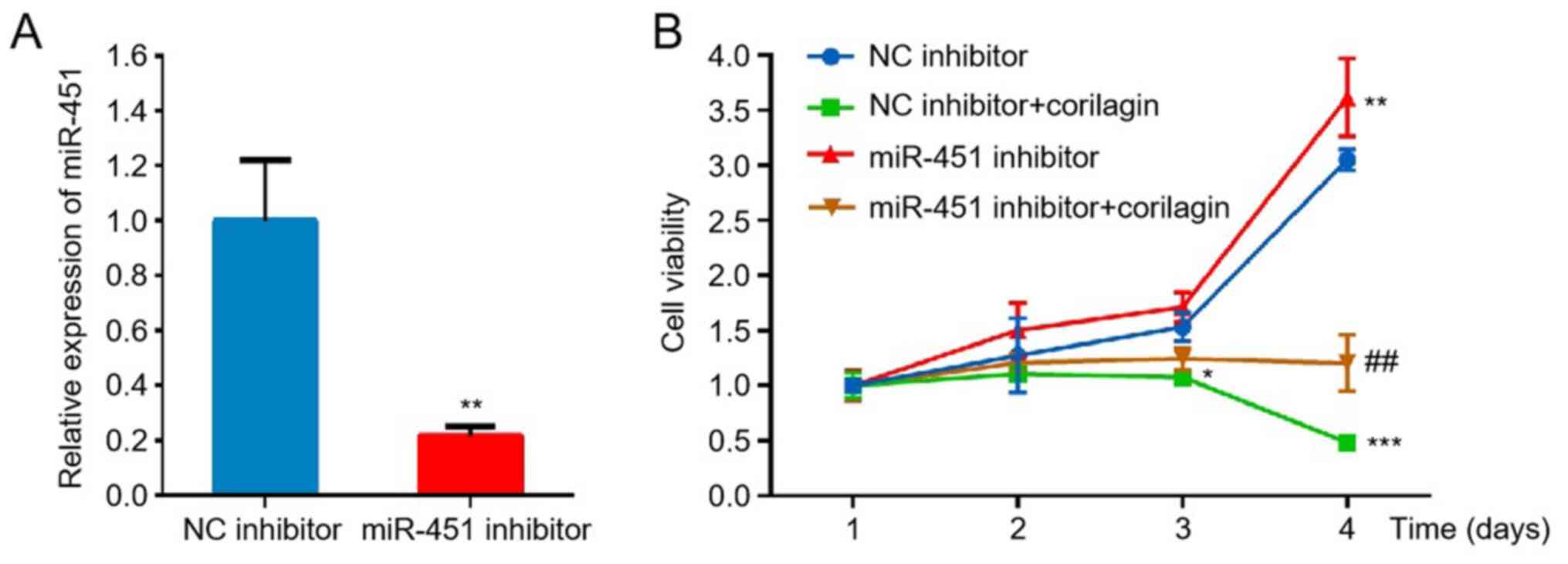
Corilagin inhibits proliferation of
HL-60 cells via the miR-451/HMGB1 axis
To investigate whether regulation of miR-451/HMGB1
axis by corilagin is involved in inhibition of HL-60 cell
proliferation, miR-451 was knocked down and cell proliferation was
analyzed. The CCK-8 assay showed that knockdown of miR-451 promoted
proliferation of HL-60 cells, indicating a tumor suppressive effect
of miR-451 on AML cells. Compared with corilagin treatment-alone,
knockdown of miR-451 attenuated corilagin-induced inhibition of
HL-60 cell proliferation, which implied that miR-451 was involved
in corilagin-induced inhibition of proliferation (Fig. 6A and B). A recent study showed that
miR-451 enhances death of AML cells by targeting HMGB1 (38). As aforementioned, the present
results also suggested negative regulation of HMGB1 by miR-451
during corilagin treatment (Fig. 5E and
F). Therefore, it was concluded that corilagin inhibited the
proliferation of HL-60 cells via the miR-451/HMGB1 axis.
Discussion
Corilagin, a water-soluble tannin, is the primary
bioactive ingredient of the E. phyllanthus plant. Corilagin
possesses anti-viral, anti-inflammatory, antioxidant and antitumor
properties (10–12). Corilagin inhibits proliferation of a
variety of cancer cells, including hepatic carcinoma, as well as
breast, gastric and ovarian cancer (15–17).
To the best of our knowledge, however, the effect of corilagin on
leukemia cells has rarely been reported. In the present study,
corilagin significantly inhibited proliferation of HL-60 AML cells
in a concentration- and time-dependent manner.
Apoptosis, a type of programmed cell death, is a key
mechanism underlying the activity of numerous antitumor drugs, such
as oblimersen sodium and nutlins (39–41).
The present results indicated that the suppressive effect of
corilagin on HL-60 cells was associated with apoptotic cell death.
Flow cytometry analysis of Annexin V/7-AAD staining showed that the
percentage of early- and late-stage apoptotic cells increased
following corilagin treatment in a dose-dependent manner. Apoptosis
is primarily regulated by the mitochondria-mediated intrinsic,
death receptor-mediated extrinsic and endoplasmic reticulum
stress-induced apoptotic pathways (42). The intrinsic pathway is accompanied
by activation of pro-apoptotic proteins (Bax and Bak), inhibition
of anti-apoptotic proteins (Bcl-2 and Bcl-xl) and cleavage of
procaspases-3/6/7, leading to cell death. In the present study,
corilagin increased protein levels of cleaved caspase-3 and Bak and
decreased levels of Bcl-xl, indicating that the intrinsic
mitochondrial apoptosis pathway was activated following exposure to
corilagin.
Autophagy is a highly conserved process that
maintains homeostasis by degrading and recycling protein
aggregates, long-lived proteins and damaged organelles (43). Under stress, such as nutrient
deprivation, hypoxia, chemotherapy and radiotherapy, autophagy is a
survival mechanism that provides nutrients for vital cellular
functions and protects cells from apoptosis or necrosis (43). There is crosstalk between autophagy
and apoptosis and inhibition of autophagy may lead to apoptotic
cell death (44,45). The present results showed that
corilagin markedly decreased the conversion of LC3-I into LC3-II
and the formation of LC3 puncta, leading to the accumulation of
substrate p62. These data suggested that corilagin exhibited an
inhibitory effect on AML cell autophagy; further research should
investigate the association between autophagy and corilagin-induced
apoptosis.
Numerous types of miRNA are aberrantly expressed in
AML, including miR-181, miR-21, miR-34 and miR-451 (20). miR-451, which serves as a tumor
suppressor by suppressing proliferation of malignant cells and
inducing apoptosis, is abnormally downregulated in patients with
AML. Previous studies have showed that miR-451 inhibits AML
development by targeting tyrosine 3-monooxygenase/tryptophan
5-monooxygenase activation protein ζ and heterogeneous nuclear
ribonucleoprotein A1 (25,26). However, a recent study found that
miR-451 enhances the death of AML cells by targeting HMGB1
(38). HMGB1 levels in the serum of
patients with AML increase significantly following chemotherapy,
and inhibition of HMGB1 improves the sensitivity of AML cells to
chemotherapy drugs (24,33).
The present study investigated whether corilagin
exerts its anti-proliferative effects by regulating the
miR-451/HMGB1 axis. microRNA sequencing and RT-qPCR both
demonstrated that corilagin notably elevated the expression of
miR-451 in a concentration-dependent manner, while mRNA and protein
levels of HMGB1 were markedly decreased in response to corilagin in
a dose-dependent manner. Knockdown of miR-451 decreased
corilagin-induced downregulation of HMGB1, indicating negative
regulation of HMGB1 by miR-451 following corilagin treatment.
Furthermore, knockdown of miR-451 also attenuated corilagin-induced
inhibition of HL-60 cell proliferation, implying that miR-451 was
involved in the inhibitory effect of corilagin on cell
proliferation. Therefore, it was hypothesized that corilagin may
function in AML treatment by regulating the miR-451/HMGB1 axis.
In conclusion, the present study demonstrated the
inhibitory effects of corilagin on HL-60 AML cell proliferation and
viability in a dose- and time-dependent manner. Corilagin induced
apoptosis in HL-60 cells by activating the intrinsic mitochondrial
apoptosis pathway. Moreover, inhibition of autophagy in HL-60 cells
induced by corilagin may be associated with apoptotic cell death.
Corilagin modulated miR-451 and HMGB1 expression, and knockdown of
miR-451 alleviated corilagin-induced downregulation of HMGB1 and
inhibition of HL-60 cell proliferation, indicating that corilagin
may exert its anticancer effects by regulating the miR-451/HMGB1
axis. These results suggested that corilagin may be a potential
novel drug for treating AML.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and Technology
Development Program of Medical and Health from Shandong Province
(grant no. 2018WSB34003), PhD Research Foundation of Affiliated
Hospital of Jining Medical University (grant no. 2018-BS-011) and
Incubation Project of National Natural Science Foundation of China
(grant no. 700103005).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the authors on reasonable request.
Authors' contributions
SJ and DH performed experiments and interpreted
data. XL, PC, JL, MC and CW performed the experiments. SJ conceived
the study, directed the experiments and wrote the manuscript. CM
and HZ designed and conceived the study. SJ and DH confirm the
authenticity of all the raw data. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dohner H, Weisdorf DJ and Bloomfield CD:
Acute myeloid leukemia. N Engl J Med. 373:1136–1152. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Khwaja A, Bjorkholm M, Gale RE, Levine RL,
Jordan CT, Ehninger G, Bloomfield CD, Estey E, Burnett A,
Cornelissen JJ, et al: Acute myeloid leukaemia. Nat Rev Dis
Primers. 2:160102016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sasine JP and Schiller GJ: Emerging
strategies for high-risk and relapsed/refractory acute myeloid
leukemia: Novel agents and approaches currently in clinical trials.
Blood Rev. 29:1–9. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shafer D and Grant S: Update on rational
targeted therapy in AML. Blood Rev. 30:275–283. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nair R, Salinas-Illarena A and Baldauf HM:
New strategies to treat AML: Novel insights into AML survival
pathways and combination therapies. Leukemia. 35:299–311. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Short NJ, Konopleva M, Kadia TM, Borthakur
G, Ravandi F, DiNardo CD and Daver N: Advances in the treatment of
acute myeloid leukemia: New drugs and new challenges. Cancer
Discov. 10:506–525. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dohner H, Estey EH, Amadori S, Appelbaum
FR, Buchner T, Burnett AK, Dombret H, Fenaux P, Grimwade D, Larson
RA, et al: Diagnosis and management of acute myeloid leukemia in
adults: Recommendations from an international expert panel, on
behalf of the European LeukemiaNet. Blood. 115:453–474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Karathedath S, Rajamani BM, Musheer Aalam
SM, Abraham A, Varatharajan S, Krishnamurthy P, Mathews V,
Velayudhan SR and Balasubramanian P: Role of NF-E2 related factor 2
(Nrf2) on chemotherapy resistance in acute myeloid leukemia (AML)
and the effect of pharmacological inhibition of Nrf2. PLoS One.
12:e01772272017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shafat MS, Gnaneswaran B, Bowles KM and
Rushworth SA: The bone marrow microenvironment-Home of the leukemic
blasts. Blood Rev. 31:277–286. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tong F, Zhang J, Liu L, Gao X, Cai Q, Wei
C, Dong J, Hu Y, Wu G and Dong X: Corilagin attenuates
radiation-induced brain injury in mice. Mol Neurobiol.
53:6982–6996. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li X, Deng Y, Zheng Z, Huang W, Chen L,
Tong Q and Ming Y: Corilagin, a promising medicinal herbal agent.
Biomed Pharmacother. 99:43–50. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lu J, Ye C, Huang Y, Huang D, Tang L, Hou
W, Kuang Z, Chen Y, Xiao S, Yishake M and He R: Corilagin
suppresses RANKL-induced osteoclastogenesis and inhibits oestrogen
deficiency-induced bone loss via the NF-κB and PI3K/AKT signalling
pathways. J Cell Mol Med. 24:10444–10457. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gambari R, Borgatti M, Lampronti I, Fabbri
E, Brognara E, Bianchi N, Piccagli L, Yuen MC, Kan CW, Hau DK, et
al: Corilagin is a potent inhibitor of NF-kappaB activity and
downregulates TNF-alpha induced expression of IL-8 gene in cystic
fibrosis IB3-1 cells. Int Immunopharmacol. 13:308–315. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xiao HT, Lin CY, Ho DH, Peng J, Chen Y,
Tsang SW, Wong M, Zhang XJ, Zhang M and Bian ZX: Inhibitory effect
of the gallotannin corilagin on dextran sulfate sodium-induced
murine ulcerative colitis. J Nat Prod. 76:2120–2125. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Milani R, Brognara E, Fabbri E, Finotti A,
Borgatti M, Lampronti I, Marzaro G, Chilin A, Lee KK, Kok SH, et
al: Corilagin induces high levels of apoptosis in the
temozolomide-resistant T98G glioma cell line. Oncol Res.
26:1307–1315. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tong Y, Zhang G, Li Y, Xu J, Yuan J, Zhang
B, Hu T and Song G: Corilagin inhibits breast cancer growth via
reactive oxygen species-dependent apoptosis and autophagy. J Cell
Mol Med. 22:3795–3807. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jia L, Jin H, Zhou J, Chen L, Lu Y, Ming Y
and Yu Y: A potential anti-tumor herbal medicine, Corilagin,
inhibits ovarian cancer cell growth through blocking the TGF-β
signaling pathways. BMC Complement Altern Med. 13:332013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Emmrich S, Katsman-Kuipers JE, Henke K,
Khatib ME, Jammal R, Engeland F, Dasci F, Zwaan CM, den Boer ML,
Verboon L, et al: MiR-9 is a tumor suppressor in pediatric AML with
t(8;21). Leukemia. 28:1022–1032. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
So AY, Sookram R, Chaudhuri AA,
Minisandram A, Cheng D, Xie C, Lim EL, Flores YG, Jiang S, Kim JT,
et al: Dual mechanisms by which miR-125b represses IRF4 to induce
myeloid and B-cell leukemias. Blood. 124:1502–1512. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu Y, Cheng Z, Pang Y, Cui L, Qian T,
Quan L, Zhao H, Shi J, Ke X and Fu L: Role of microRNAs, circRNAs
and long noncoding RNAs in acute myeloid leukemia. J Hematol Oncol.
12:512019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jiang X, Hu C, Arnovitz S, Bugno J, Yu M,
Zuo Z, Chen P, Huang H, Ulrich B, Gurbuxani S, et al: MiR-22 has a
potent anti-tumour role with therapeutic potential in acute myeloid
leukaemia. Nat Commun. 7:114522016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wallace JA and O'Connell RM: MicroRNAs and
acute myeloid leukemia: Therapeutic implications and emerging
concepts. Blood. 130:1290–1301. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu L, Ren W and Chen K: MiR-34a promotes
apoptosis and inhibits autophagy by targeting HMGB1 in acute
myeloid leukemia cells. Cell Physiol Biochem. 41:1981–1992. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Y, Liu Y and Xu X: Upregulation of
miR-142-3p improves drug sensitivity of acute myelogenous leukemia
through reducing P-glycoprotein and repressing autophagy by
targeting HMGB1. Transl Oncol. 10:410–418. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Su R, Gong JN, Chen MT, Song L, Shen C,
Zhang XH, Yin XL, Ning HM, Liu B, Wang F, et al: c-Myc suppresses
miR-451 dash, verticalYWTAZ/AKT axis via recruiting HDAC3 in acute
myeloid leukemia. Oncotarget. 7:77430–77443. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Song L, Lin HS, Gong JN, Han H, Wang XS,
Su R, Chen MT, Shen C, Ma YN, Yu J and Zhang JW:
MicroRNA-451-modulated hnRNP A1 takes a part in granulocytic
differentiation regulation and acute myeloid leukemia. Oncotarget.
8:55453–55466. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cao J, Da Y, Li H, Peng Y and Hu X:
Upregulation of microRNA-451 attenuates myocardial I/R injury by
suppressing HMGB1. PLoS One. 15:e02356142020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Amato J, Cerofolini L, Brancaccio D,
Giuntini S, Iaccarino N, Zizza P, Iachettini S, Biroccio A,
Novellino E, Rosato A, et al: Insights into telomeric G-quadruplex
DNA recognition by HMGB1 protein. Nucleic Acids Res. 47:9950–9966.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kang R, Chen R, Zhang Q, Hou W, Wu S, Cao
L, Huang J, Yu Y, Fan XG, Yan Z, et al: HMGB1 in health and
disease. Mol Aspects Med. 40:1–116. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kang R, Livesey KM, Zeh HJ, Loze MT and
Tang D: HMGB1: A novel Beclin 1-binding protein active in
autophagy. Autophagy. 6:1209–1211. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pan B, Chen D, Huang J, Wang R, Feng B,
Song H and Chen L: HMGB1-mediated autophagy promotes docetaxel
resistance in human lung adenocarcinoma. Mol Cancer. 13:1652014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hou X, Yang C, Zhang L, Hu T, Sun D, Cao
H, Yang F, Guo G, Gong C, Zhang X, et al: Killing colon cancer
cells through PCD pathways by a novel hyaluronic acid-modified
shell-core nanoparticle loaded with RIP3 in combination with
chloroquine. Biomaterials. 124:195–210. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu L, Zhang J, Zhang X, Cheng P, Liu L,
Huang Q, Liu H, Ren S, Wei P, Wang C, et al: HMGB1: An important
regulator of myeloid differentiation and acute myeloid leukemia as
well as a promising therapeutic target. J Mol Med (Berl).
99:107–118. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang R, Xu Y, Wan W, Shou X, Qian J, You
Z, Liu B, Chang C, Zhou T, Lippincott-Schwartz J and Liu W:
Deacetylation of nuclear LC3 drives autophagy initiation under
starvation. Mol Cell. 57:456–466. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang F, Wang Y, Xue J, Ma Q, Zhang J, Chen
YF, Shang ZZ, Li QQ, Zhang SL and Zhao L: Effect of corilagin on
the miR-21/smad7/ERK signaling pathway in a schistosomiasis-induced
hepatic fibrosis mouse model. Parasitol Int. 65:308–315. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhou X, Xiong J, Lu S, Luo L, Chen ZL,
Yang F, Jin F, Wang Y, Ma Q, Luo YY, et al: Inhibitory effect of
corilagin on miR-21-regulated hepatic fibrosis signaling pathway.
Am J Chin Med. 47:1541–1569. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang Y, Chu X and Wei Q: MiR-451 promotes
cell apoptosis and inhibits autophagy in pediatric acute myeloid
leukemia by targeting HMGB1. J Environ Pathol Toxicol Oncol.
40:45–53. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pistritto G, Trisciuoglio D, Ceci C,
Garufi A and D'Orazi G: Apoptosis as anticancer mechanism: Function
and dysfunction of its modulators and targeted therapeutic
strategies. Aging (Albany NY). 8:603–619. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rai KR, Moore J, Wu J, Novick SC and
O'Brien SM: Effect of the addition of oblimersen (Bcl-2 antisense)
to fludarabine/cyclophosphamide for replased/refractory chronic
lymphocytic leukaemia (CLL) on survival in patients who achieve
CR/nPR: Five-year follow-up from a randomized phase III study. J
Clin Oncol. 26 (Suppl 15):S70082008. View Article : Google Scholar
|
|
41
|
Shangary S and Wang S: Small-molecule
inhibitors of the MDM2-p53 protein-protein interaction to
reactivate p53 function: A novel approach for cancer therapy. Annu
Rev Pharmacol Toxicol. 49:223–241. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hu H, Tian M, Ding C and Yu S: The C/EBP
homologous protein (CHOP) transcription factor functions in
endoplasmic reticulum stress-induced apoptosis and microbial
infection. Front Immunol. 9:30832018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Dikic I and Elazar Z: Mechanism and
medical implications of mammalian autophagy. Nat Rev Mol Cell Biol.
19:349–364. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Maiuri MC, Zalckvar E, Kimchi A and
Kroemer G: Self-eating and self-killing: Crosstalk between
autophagy and apoptosis. Nat Rev Mol Cell Biol. 8:741–752. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li Q, Yin Y, Zheng Y, Chen F and Jin P:
Inhibition of autophagy promoted high glucose/ROS-mediated
apoptosis in ADSCs. Stem Cell Res Ther. 9:2892018. View Article : Google Scholar : PubMed/NCBI
|