Introduction
Fibrosis is an inevitable outcome after chronic
damage to an organ or tissue, and is the main pathological basis
that ultimately causes organ dysfunction (1). According to a population-based
cohort study, >1/2 of patients with inflammatory bowel disease
have diseased intestinal segments that will progress to intestinal
fibrosis, ultimately leading to intestinal stenosis (2). In addition, due to repeated
flare-ups and an increased risk of bowel cancer, Crohn's disease is
a lifelong, incurable inflammatory bowel disease (3). During the course of intestinal
fibrosis development, fibrosis will gradually occur in the
intestinal wall (4), and can lead
to narrowing of the intestinal lumen and intestinal obstruction
(5,6). These patients will eventually
require surgery to relieve the obstruction caused by fibrotic
stenosis (7,8).
Compared with studies on liver, lung and skin
fibrosis, which have been a hotspot of research, it has only been
in recent years that intestinal fibrosis in inflammatory bowel
disease has gained increased attention from the academic community
(2). At present, the etiology of
inflammatory bowel disease remains largely unknown; however, the
generally accepted hypothesis is that under the influence of
certain genetic susceptibility and environmental factors, an
intestinal microecological imbalance can occur, which then causes
intestinal mucosal immune disorders (9). Early genetic to intestinal
microecology research, as well as mucosal immune-related studies,
have been key in understanding the underlying mechanism of
inflammatory bowel disease, and biologics targeting pathogenesis
have achieved a precise effect for disease control or remission
(10–12). However, the long-term outcome of
patients with inflammatory bowel disease has not been significantly
improved (13,14), and it remains difficult to
identify a solution based on the pathogenesis (15). Therefore, it is a reasonable to
focus on the final pathological state, which is fibrosis, and
develop strategies that can potentially alter the natural disease
course and long-term outcome of patients.
Extracellular matrix (ECM) is overproduced and
deposited during the process of intestinal fibrosis (16). There are a variety of growth
factors, such as transforming growth factor (TGF)-β, epidermal
growth factor and insulin like growth factor (17), in the ECM that can upregulate and
activate epithelial-mesenchymal transition (EMT) transcription
factors, thereby initiating the EMT process. At present, the
development of intestinal anti-fibrosis drugs is lacking (18). Therefore, it is of practical
significance to investigate novel drugs to delay or block the
development of fibrosis.
Atractylenolide III (ATL-III), the main bioactive
component of Atractylodes macrocephala, also exists in other
medicinal plants, such as Codonopsis and cocklebur (19). It has been shown to exert a
variety of pharmacological activities, including anti-allergic,
anti-inflammatory, gastroprotective and neuroprotective effects
(20,21). For example, ATL-III was found to
reduce depression and anxiety-like behaviors in rat depression
models (22). In addition, a
previous study revealed that ATL-III could reduce muscle wasting in
chronic kidney disease by activating oxidative stress (23), as well as effectively improve
bleomycin-induced lung injury and function in rats with pulmonary
fibrosis (24). Moreover, in
human breast cancer cell lines, MDA-MB-231 and MDA-MB-468, ATL-III
was shown to significantly block the cell migration and invasion
induced by TGF-β1, ultimately inhibiting the motility of metastatic
breast cancer cells in mice models, thereby indicating that ATL-III
inhibits the process of EMT in vitro and in vivo
(25). However, whether ATL-III
can also inhibit the EMT process of intestinal cells remains to be
verified.
It has been reported that ATL-III notably increases
the phosphorylation of AMP-activated protein kinase (AMPK) and the
expression of sirtuin 1, indicating that ATL-III may have a
beneficial effect on obesity and type 2 diabetes mellitus by
improving the energy metabolism of skeletal muscle (26). Apigenin inhibits the
proliferation, differentiation and function of renal fibroblasts
via AMPK activation, as well as reduces ERK1/2 phosphorylation,
suggesting that it may have a favorable therapeutic potential for
the treatment of renal fibrosis (27). Furthermore, wedelolactone, a major
coumarin ingredient of E. prostrata, can cause increases in
the expression levels of fibrosis markers (collagen I and α-smooth
muscle actin) and a decrease in that of the anti-fibrosis marker
(E-cadherin). Wedelolactone also activates AMPK and inhibits the
increase in TGF-β1 phosphorylation, thereby inhibiting the EMT of
alveolar epithelial cells (28).
Thus, it was suggested that activation of the AMPK signaling
pathway can inhibit the EMT process.
As IEC-6 cells can be passaged stably and have the
typical morphological and growth characteristics of normal
intestinal epithelial cells, they have been widely used in multiple
research studies related to intestines (29–31). In the present study, the effect of
ATL-III on the EMT process of a small intestine epithelial cell
line, IEC-6, was investigated, as well as its underlying mechanism.
The current findings could provide a theoretical basis for ATL-III
application to inhibit organ fibrosis in the future.
Materials and methods
Cell culture and reagents
IEC-6 (rat small intestinal epithelial) cells were
purchased from Merck-KGaA. ATL-III (purity, >98%) was purchased
from Chengdu Pufei De Biotech Co., Ltd. and diluted to 1, 10 or 20
µmol/l (32). TGF-β1 protein
(cat. no. TG1-M5218) was purchased from Acro Biosystems Co., Ltd.
Compound C (also known as dorsomorphin; cat. no. HY-13418A), a type
of selective AMPK inhibitor, was purchased from MedChemExpress.
IEC-6 cells were cultured in DMEM (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.), 10 U/ml insulin and 100 U/ml
penicillin-streptomycin (Invitrogen; Thermo Fisher Scientific,
Inc.) at 37°C in an incubator with 5% CO2. Cells were
divided into: Control, ATL-III, TGF-β1, TGF-β1 + ATL-III and TGF-β1
+ ATL-III + compound C groups.
MTT assay
IEC-6 cells (5×103 cells/well) were
seeded onto 96-well plates and cultured in an incubator with 5%
CO2 at 37°C. Cells were treated with different doses of
ATL-III (0, 1, 10 or 20 µmol/l) at 37°C for 24 h. MTT solution (200
µl) was added to each well and the incubation was continued at 37°C
for 4 h. Then, the medium was removed and DMSO was added. The
optical density was measured at 490 nm wavelengths using a
microplate reader (Thermo Fisher Scientific, Inc.).
Wound healing assay
IEC-6 cells were seeded onto 6-well plates and
incubated at 37°C overnight. Sterilized pipette tips (200 µl) were
used to scratch the cells. Next, plates were washed three times
with PBS to remove the cells, and serum-free medium with TGF-β1 (10
ng/ml), ATL-III (1, 10 or 20 µmol/l) and compound C (20 µmol/l) was
added for the continuing culture in the incubator at 37°C. At 0 and
24 h, images were captured under an inverted microscope
(magnification, ×100; Olympus Corporation) and evaluated using
ImageJ software (1.52v; National Institutes of Health). Cell
migration rate = wound area difference between 0 and 24 h/wound
area at 0 h.
Transwell assay
Matrigel (BD Biosciences) was thawed overnight at
4°C and diluted with serum-free medium, following which the diluent
was inoculated into the upper chamber for 30 min at 37°C. IEC-6
cells (5×104) were seeded into the upper chamber
(Corning, Inc.) with 100 µl serum-free medium containing TGF-β1 (10
ng/ml), ATL-III (1, 10 or 20 µmol/l) and compound C (20 µmol/l).
The lower chamber was filled with 600 µl medium containing 10% FBS.
Following 24 h of incubation, the cells on the lower side were
fixed with 4% formaldehyde for 20 min at room temperature and
stained with 0.1% crystal violet solution for 20 min at room
temperature. Images were captured under an inverted microscope
(magnification, ×100; Olympus Corporation). Cell invasion rate =
the number of invasive cells/number of inoculated cells.
Western blot analysis
Upon IEC-6 cells reaching 60% confluence, TGF-β1 (10
ng/ml), ATL-III (1, 10 or 20 µmol/l) and compound C (20 µmol/l)
were added and incubated at 37°C for 48 h. Proteins were extracted
and homogenized in RIPA lysis buffer (Beyotime Institute of
Biotechnology). Proteins were determined using a BCA kit (Beyotime
Institute of Biotechnology), and then 30 µg samples/lane were
separated on 10% SDS-polyacrylamide gels, followed by transfer to a
PVDF membrane. The membrane was washed with TBS-0.01% Tween-20
(TBST) and then incubated in blocking fluid containing 5% non-fat
milk at 25°C for 1 h. Strips were cut and incubated with the
following primary antibodies at 4°C overnight: MMP9 (cat. no.
ab76003; 1:1,000), vimentin (cat. no. ab92547; 1:1,000), N-cadherin
(cat. no. ab18203; 1:1,000), E-cadherin (cat. no. ab40772; 10,000),
zonula occludens (ZO)-1 (cat. no. ab216880; 1:1,000),
phosphorylated (p)-AMPK (cat. no. ab133448; 1:1,000) and AMPK (cat.
no. ab32047; 1:1,000; all from Abcam). Next, strips were incubated
with HRP-conjugated goat anti-rabbit secondary antibody (cat. no.
ab6721; 1:3,000; Abcam) at room temperature for 1 h after washing
with TBST. An ECL chromogenic substrate (Beyotime Institute of
Biotechnology) was used for visualization and data were analyzed
using Image Lab v4.0 software (Bio-Rad Laboratories, Inc.).
Immunofluorescence assay
IEC-6 cells were seeded onto 6-well plates and
incubated. When cells grew to 90% confluence, TGF-β1 (10 ng/ml),
ATL-III (1, 10 or 20 µmol/l) and compound C (20 µmol/l) were added
at 37°C. After 48 h of incubation, cells were fixed with 4%
formaldehyde for 15 min at room temperature and washed with PBS.
Then, 0.2% Triton X-100 was added, and subsequently removed using
PBS after 15 min. Next, 5% BSA (Beijing Solarbio Science &
Technology Co., Ltd.) was added for 30 min at room temperature for
the blocking step, following which cells were incubated with
N-cadherin (cat. no. ab18203; 1:200) or E-cadherin antibody (cat.
no. ab40772; 1:500) at room temperature for 2 h. Then, FITC-labeled
goat anti-rabbit secondary antibody (cat. no. ab6717; 1:100; all
from Abcam) was applied at room temperature for 1 h. The nuclei
were stained with DAPI for 10 min at room temperature and images
were captured under a fluorescence microscope (magnification, ×200;
Nikon Corporation).
Statistical analysis
GraphPad Prism 8.0 statistical software (GraphPad
Software, Inc.) was utilized to analyze the experimental data. All
data are presented as the mean ± SD of three replicate experiments.
P<0.05 was considered to indicate a statistically significant
difference. A unpaired Student's t-test was used to evaluate
differences between two groups, while one-way ANOVA and Tukey's
post hoc test were used for multiple groups.
Results
ATL-III inhibits the invasion and
migration of IEC-6 cells stimulated with TGF-β1
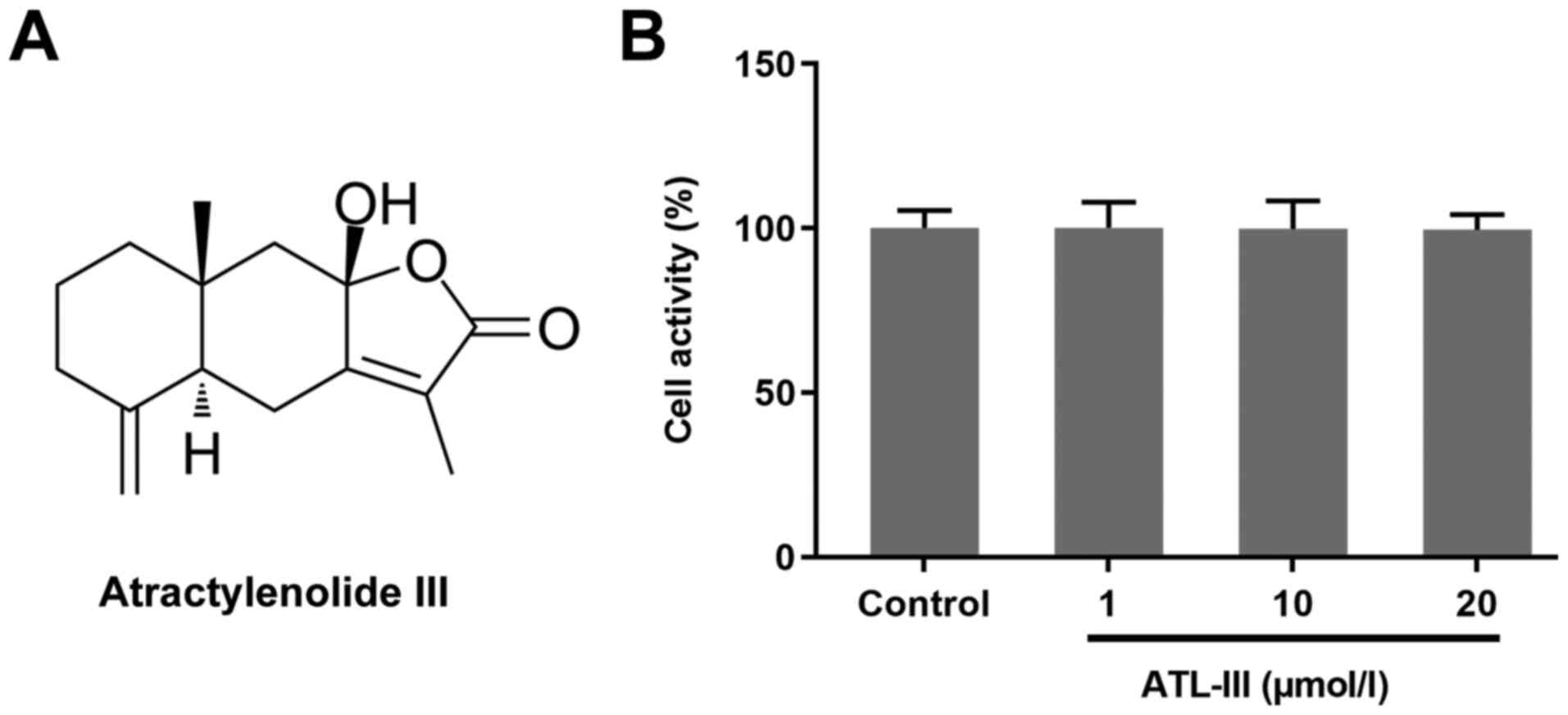
The chemical structure of ATL-III is shown in
Fig. 1A. An MTT assay was used to
detect the effect of ATL-III on the activity of IEC-6 cells. Cells
were treated with different doses of ATL-III [0 (control), 1, 10 or
20 µmol/l]. The addition of ATL-III had no significant effect on
normal cultured cells (Fig. 1B),
which suggests that ATL-III was not toxic to the normal cultured
cells if the concentration was ≤20 µmol/l.
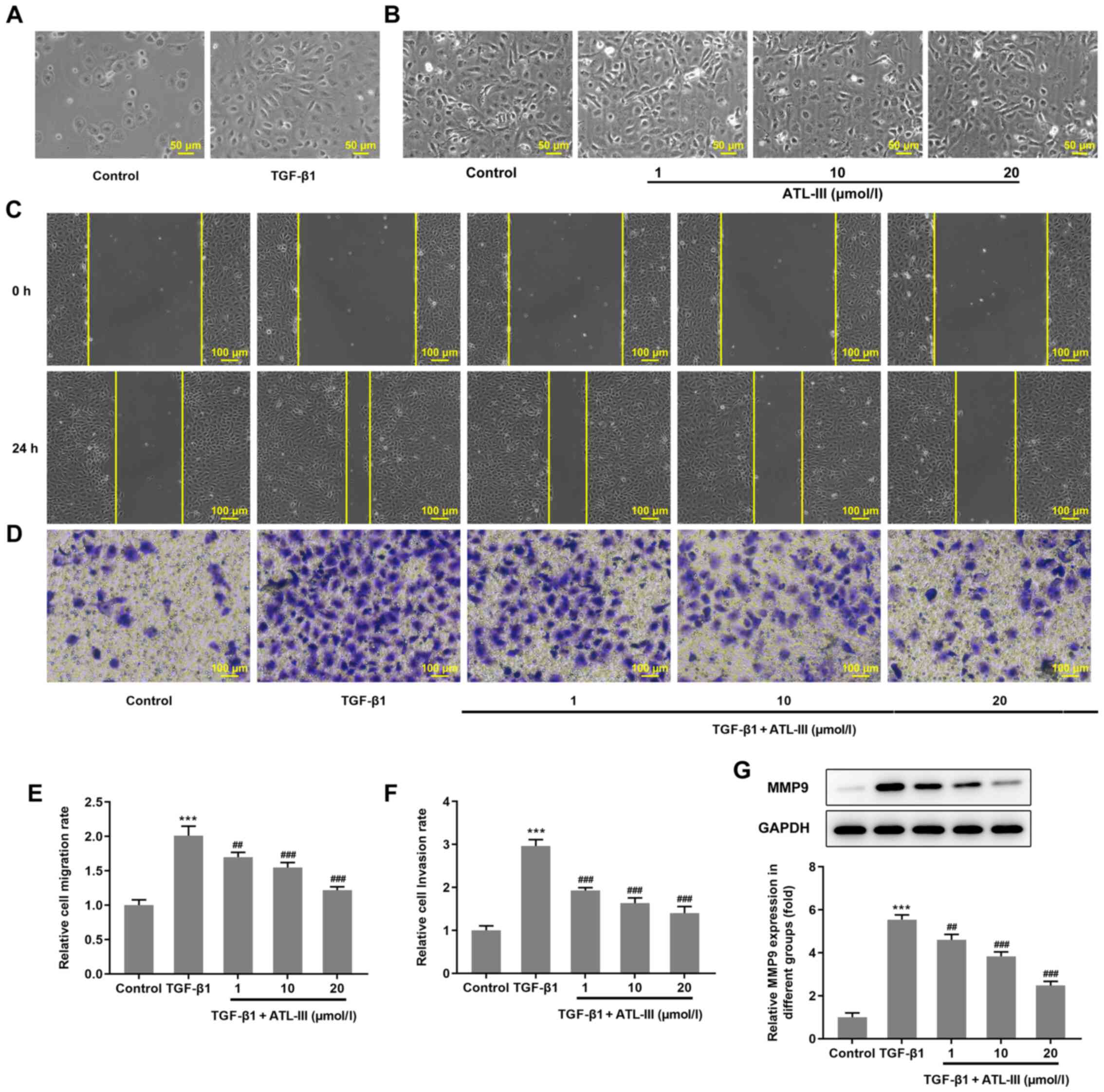
IEC-6 cells were treated with TGF-β1 or ATL-III for
48 h. Using a microscope it was observed that the cells in the
TGF-β1 group changed from a cubic shape to a spindle shape
(Fig. 2A), whereas the cells in
the ATL-III groups had no obvious alteration in morphology
(Fig. 2B). The migration of IEC-6
cells was detected using a wound healing assay. Compared with the 0
h group, after 24 h, the cells migrated to the wound area (Fig. 2C). The cell migration rate was
notably increased by TGF-β1 stimulation. However, when ATL-III was
added, the migration rate was decreased in a
concentration-dependent manner (Fig.
2E). Cell invasion was detected using a Transwell assay. The
trend observed for the invasive rate was the same as that of the
migration rate (Fig. 2D and F).
The expression levels of protein associated with migration were
detected via western blotting. The expression level of MMP9 was
increased when cells were treated with TGF-β1, while the addition
of ATL-III reduced this expression in a dose-dependent manner
(Fig. 2G). These findings
indicate that ATL-III can inhibit the invasion and migration of
IEC-6 cells induced by TGF-β1.
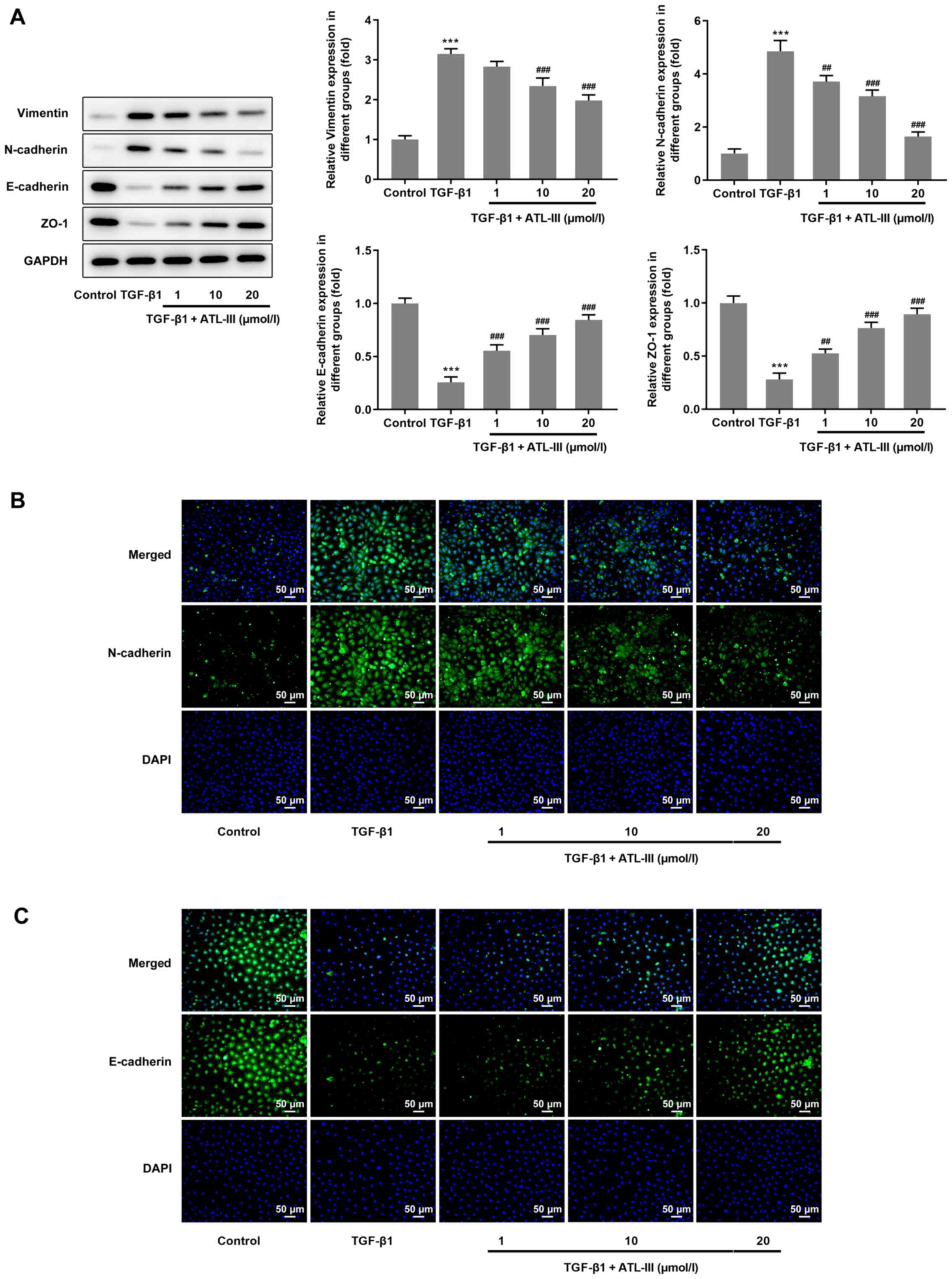
ATL-III inhibits the EMT of IEC-6
cells induced by TGF-β1
The expression levels of proteins associated with
EMT were detected via western blotting. The expression levels of
vimentin and N-cadherin were increased when cells were treated with
TGF-β1, but the addition of ATL-III reduced this expression in a
dose-dependent manner. However, opposite trends were observed with
regards to the expression levels of E-cadherin and ZO-1. After
stimulation with TGF-β1, the expression levels of E-cadherin and
ZO-1 were decreased, but when ATL-III was added, these were
increased in a dose-dependent manner (Fig. 3A). Furthermore, an
immunofluorescence assay was used for verification. The results for
N-cadherin and E-cadherin expression from the fluorescent images
matched the results from the western blot analysis (Fig. 3B and C).
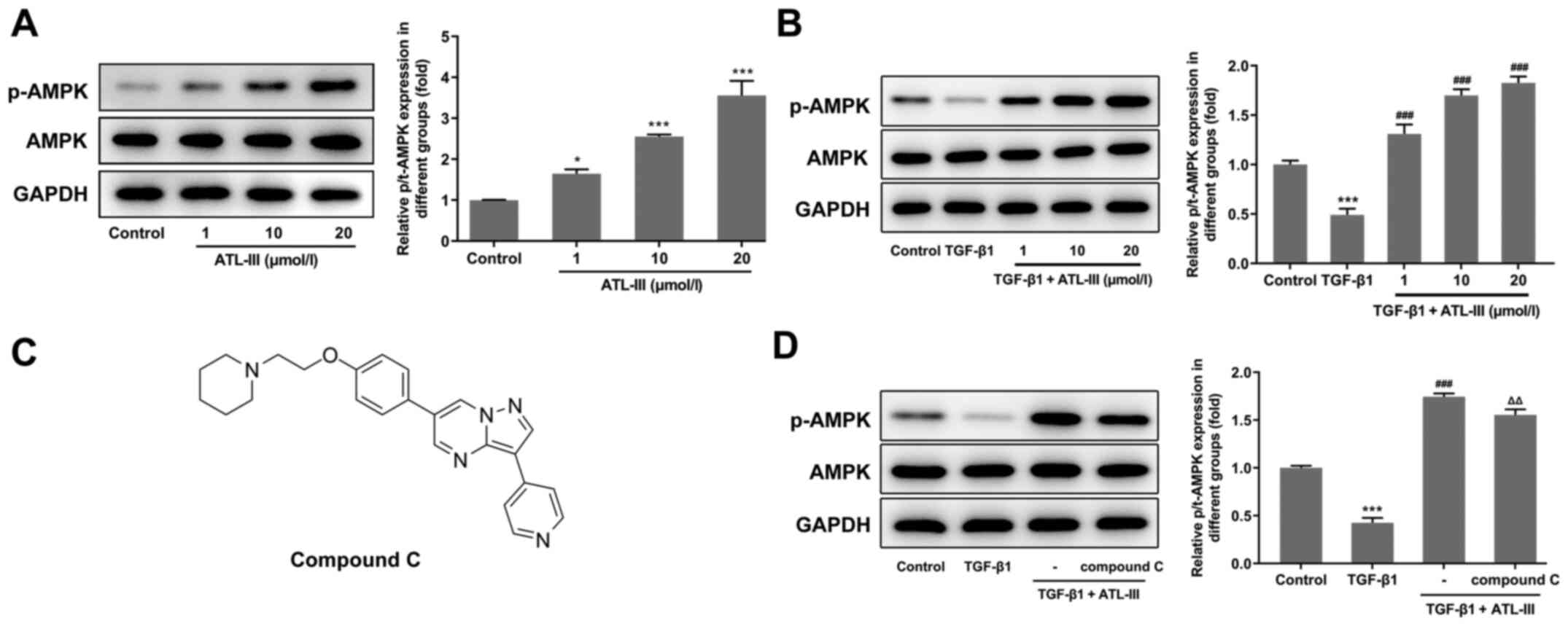
ATL-III inhibits the invasion,
migration and EMT process of IEC-6 cells induced by TGF-β1 through
activating the AMPK signaling pathway
To investigate the effect of ATL-III on the AMPK
signaling pathway, the expression levels of p-AMPK and AMPK were
detected using western blotting. The expression level of p-AMPK was
increased in a concentration-dependent manner after treatment with
ATL-III (Fig. 4A). When IEC-6
cells were stimulated by TGF-β1, the expression level of p-AMPK was
decreased compared with the control group. However, after the
addition of ATL-III, p-AMPK expression was upregulated (Fig. 4B). This indicates that ATL-III can
activate the AMPK signaling pathway in IEC-6 cells.
Next, the cells were divided into four groups
(control, TGF-β1, TGF-β1 + ATL-III and TGF-β1 + ATL-III + compound
C). The concentration of ATL-III used was 20 µmol/l. Western
blotting was used to detect the expression levels of AMPK signaling
pathway-related proteins after pretreatment with compound C
(Fig. 4C). When cells were
stimulated with TGF-β1, p-AMPK expression was decreased compared
with the control group. After ATL-III was added, this expression
rose rapidly. However, when compound C was added, the expression
level was decreased. Simultaneously, the expression level of AMPK
showed no fluctuation (Fig.
4D).
Wound healing and Transwell assays were used to
detect the invasion and migration of cells. The cell invasive and
migratory rates were both increased when compound C was added
compared with the TGF-β1 + ATL-III group (Fig. 5A-D). In addition, the expression
levels of MMP9, vimentin, N-cadherin, E-cadherin and ZO-1 were
detected via western blotting. The expression levels of MMP9,
vimentin and N-cadherin were increased, while those of E-cadherin
and ZO-1 were decreased after the addition of compound C compared
with the TGF-β1 + ATL-III group (Fig.
5E). These results were in line with the aforementioned wound
healing and Transwell assay results. Moreover, an
immunofluorescence assay was used to detect the expression levels
of N-cadherin and E-cadherin. It was found that the N-cadherin
expression was inhibited in the TGF-β1 + ATL-III group, while the
addition of compound C increased the expression levels of
N-cadherin (Fig. 5F). E-cadherin
expression was found to be promoted in the TGF-β1 + ATL-III group,
while the addition of compound C declined E-cadherin expression
(Fig. 5G). Thus, it was suggested
that the AMPK pathway plays a role in the EMT process. Overall,
ATL-III may inhibit the invasion, migration and EMT process of
IEC-6 cells induced by TGF-β1 by activating the AMPK signaling
pathway.
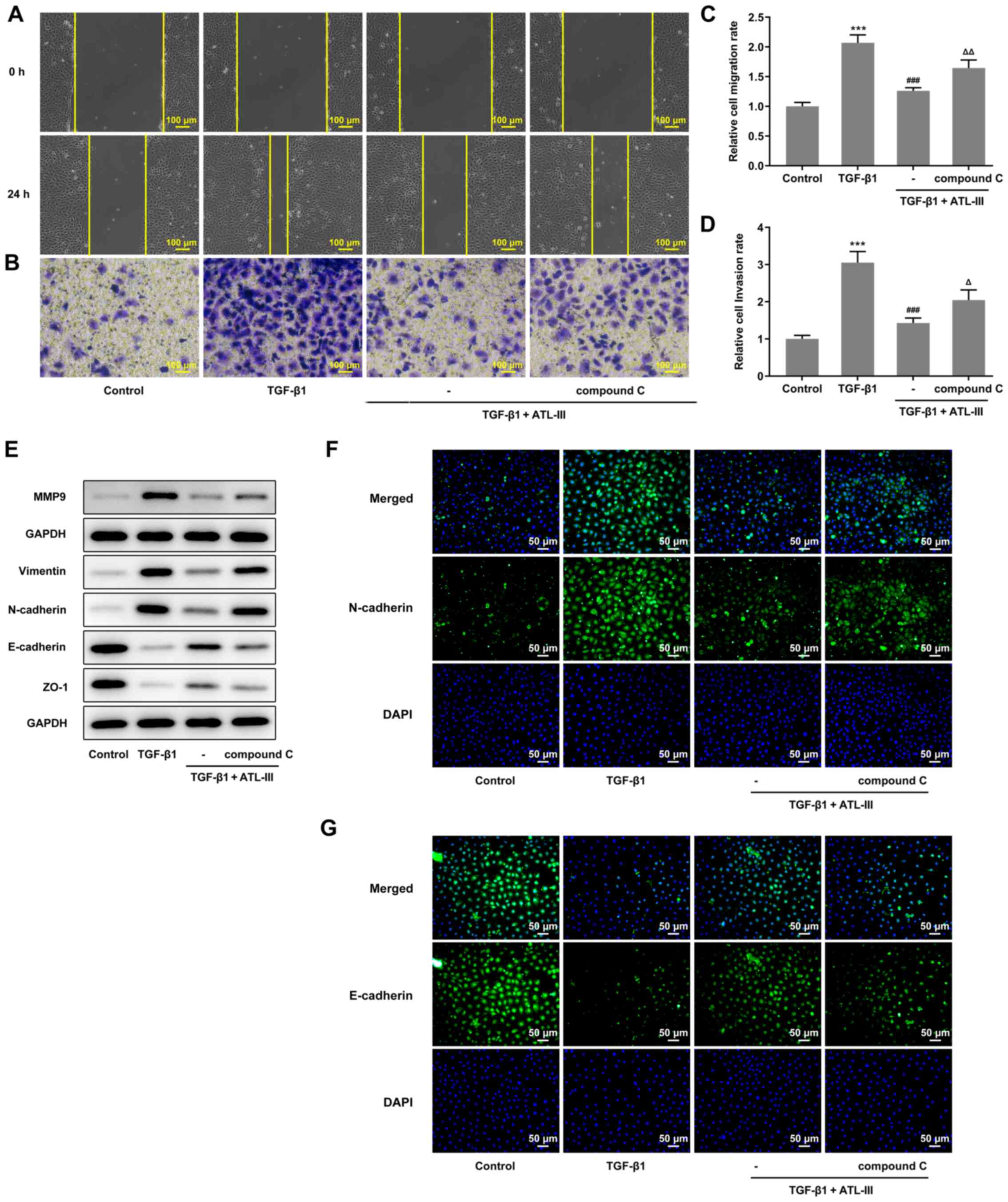 | Figure 5.ATL-III inhibits the invasion,
migration and EMT process of IEC-6 cells induced by TGF-β1 by
activating the AMPK signaling pathway. (A) Cell migration, as
determined using a wound healing assay. (B) Cells invasion was
detected using a Transwell assay. Magnification, ×100. Histograms
of cell (C) migration and (D) invasion rates. (E) Expression levels
of MMP9 and proteins associated with EMT, as detected via western
blotting. (F) N-cadherin and (G) E-cadherin expression was detected
using an immunofluorescence assay. Magnification, ×200.
***P<0.001 vs. control group; ###P<0.001 vs.
TGF-β1 group; △P<0.05, △△P<0.01 vs. TGF-β1 + ATL-III (20
µmol/l) group; n≥3. ATL-III, atractylenolide III; EMT,
epithelial-mesenchymal transition; AMPK, AMP-activated protein
kinase; ZO-1, zonula occludens-1. |
Discussion
The concept of EMT was first proposed 40 years ago
(33). Later studies have
reported that EMT was closely associated with tumor epithelial cell
invasion and organ fibrosis (34–36). Over the past 10 years, it was
discovered that EMT is also an important mechanism in the process
of intestinal fibrosis (37).
Intestinal fibrosis can cause serious disease. Most patients can
only rely on conservative treatment and fibrosis will remain
throughout the patient's lifetime. In severe cases, fibrosis can be
relieved through surgery, but patients have a high probability of
recurrence (38). In view of the
fact that clinical studies have reported that there are effective
treatments for organ fibrosis, including lung and skin (39–41), it is a reasonable idea to
similarly improve intestinal fibrosis using anti-fibrotic drugs. In
the present study, the effect of ATL-III on the EMT process of
intestinal cells was examined. First, normal IEC-6 cells were
treated with ATL-III, and it was found that ATL-III had no effect
on cell activity, as determined using an MTT assay.
Next, TGF-β was used to stimulate IEC-6 cells. TGF-β
belongs to the family of growth factors and is a multifunctional
polypeptide cytokine. TGF-β is expressed in a variety of cell types
and organs in mammals, and is associated with ECM (42). In the intestine, both immune and
non-immune cells can secrete TGF-β. In animal experiments, it has
been shown that the overexpression of TGF-β can cause mice to
develop intestinal fibrosis and obstruction (43). In addition, blocking TGF-β/Smad
signal transduction can protect mice from colonic fibrosis
(44). There are three subtypes
of TGF-β, of which TGF-β1 is the most highly expressed (45). These findings indicate that TGF-β1
is associated with fibrosis, and thus, in the present study it was
used to stimulate IEC-6 cells to differentiate into fibroblasts.
After 48 h of induction, using a microscope it was observed that
IEC-6 cells become fibroblasts.
When cells undergo EMT, they have a strong ability
to migrate and invade (30). In
the current study, wound healing and Transwell assays were used to
detect the migration and invasion of cells. The present results
demonstrated that the migratory and invasive rates of
TGF-β1-stimulated cells were increased compared with normal cells.
After the addition of the different doses of ATL-III, these rates
were gradually decreased. Moreover, western blotting was used to
detect the expression level of MMP9. It has been shown that MMP9
expression is correlated with metastatic potential (46). In the present study, the
expression level of MMP9 in cells treated with TGF-β was notably
increased, but was decreased after the addition of ATL-III.
E-cadherin and N-cadherin are two important
cadherins expressed in epithelial and mesenchymal cells,
respectively. The change in phenotype of cells expressing
E-cadherin to N-cadherin is an important mechanism in EMT (47). ZO-1 plays an important role in
maintaining the integrity of the tightly linked structure and
function of cells (48). Vimentin
is positively expressed in mesenchymal cells, but negatively
expressed in epithelial cells (49). The current results of the
immunofluorescence and western blotting assays were consistent with
this previous study, as it was demonstrated that E-cadherin and
ZO-1 expression was decreased in cells treated with TGF-β, while
N-cadherin and vimentin expression was increased. Moreover,
treatment with ATL-III could reduce the differences observed
between the control group and the stimulated cells group. This
indicates that ATL-III can inhibit TGF-β, thereby blocking the EMT
process of IEC-6 cells.
AMPK is a recognized cell bioenergy sensor and
metabolic master switch (50). A
decrease in AMPK activity is associated with diabetes, obesity and
aging, which are risk factors for organ fibrosis (51,52). In preclinical studies, AMPK
activators have been shown to play a protective role against lung
injury and reduce the subsequent development of fibrosis (53,54). Specifically, pharmacological
activation of AMPK in lung myofibroblasts of patients with
idiopathic pulmonary fibrosis caused low fibrotic activity
(55). Using a mouse lung
fibrosis model, it was found that the use of metformin (an AMPK
activator) to promote the inactivation and apoptosis of
myofibroblasts could reverse the established lung fibrosis
(56). Thus, the inhibitory
effect of AMPK activation on EMT has been previously shown. In the
present study, western blotting was used to detect p-AMPK
expression, which was found to increase when the dose of ATL-III
was enhanced. Moreover, after pretreatment with compound C, p-AMPK
expression was decreased. This suggests that ATL-III activates the
AMPK signaling pathway in IEC-6 cells. Wound healing and Transwell
assays were used to investigate cell migration and invasion after
compound C treatment. It was identified that compound C reduced the
inhibitory effect of ATL-III on the invasion and migration of
stimulated cells. The detection of MMP9 expression using western
blotting also verified this result. Finally, an immunofluorescence
assay and western blotting were used to detect the expression
levels of EMT-related proteins. The results demonstrated that when
the AMPK pathway was inhibited, the EMT inhibitory effect of
ATL-III on the cells stimulated by TGF-β was reduced. Thus, it was
indicated that the AMPK pathway may be essential for ATL-III to
inhibit the EMT process of cells stimulated by TGF-β. To the best
of our knowledge, the present study was the first to demonstrate
that ATL-III could inhibit the EMT process of intestinal cells. At
present, compared with the drug treatment for other types of organ
fibrosis, the development of intestinal anti-fibrosis drugs is
lacking. Therefore, the development of drugs to inhibit or relieve
intestinal fibrosis is of great importance. However, in vivo
experiments will also need to be performed to verify the findings
in the future.
In conclusion, the present study identified the
inhibitory effect of ATL-III on in vitro intestinal
fibrosis, and the current findings provide a novel idea for future
intestinal fibrosis drug research.
Acknowledgements
Not applicable.
Funding
This work was supported by Guizhou Science and Technology
Department and Guizhou University Joint Fund Project [grant no. LH
[(2017)].
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MH and WJ designed and performed the experiments. CL
and MY participated in experiments and analyzed the data. YR made
substantial contributions to conception, design and wrote the
manuscript. All authors read and approved the final manuscript. YR
and MH confirmed the authenticity of the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rockey DC, Bell PD and Hill JA: Fibrosis -
a common pathway to organ injury and failure. N Engl J Med.
372:1138–1149. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rieder F, Fiocchi C and Rogler G:
Mechanisms, management, and treatment of fibrosis in patients with
inflammatory bowel diseases. Gastroenterology. 152:340–350.e6.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sicilia B, Arias L, Hontoria G, García N,
Badia E and Gomollón F: Are steroids still useful in
immunosuppressed patients with inflammatory bowel disease? A
retrospective, population-based study. Front Med (Lausanne).
8:6516852021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rutgeerts P, Geboes K, Vantrappen G,
Kerremans R, Coenegrachts JL and Coremans G: Natural history of
recurrent Crohn's disease at the ileocolonic anastomosis after
curative surgery. Gut. 25:665–672. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Freeman HJ: Natural history and clinical
behavior of Crohn's disease extending beyond two decades. J Clin
Gastroenterol. 37:216–219. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mao R, Chen BL, He Y, Cui Y, Zeng ZR and
Chen MH: Factors associated with progression to surgery in Crohn's
disease patients with endoscopic stricture. Endoscopy. 46:956–962.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Froehlich F, Juillerat P, Mottet C, Pittet
V, Felley C, Vader JP, Gonvers JJ and Michetti P: Fibrostenotic
Crohn's disease. Digestion. 76:113–115. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Masaki T, Kishiki T, Kojima K, Asou N,
Beniya A and Matsuoka H: Recent trends (2016–2017) in the treatment
of inflammatory bowel disease. Ann Gastroenterol Surg. 2:282–288.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kelsen JR, Russo P and Sullivan KE:
Early-Onset Inflammatory Bowel Disease. Immunol Allergy Clin North
Am. 39:63–79. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mentella MC, Scaldaferri F, Pizzoferrato
M, Gasbarrini A and Miggiano GA: Nutrition, IBD and Gut Microbiota:
A Review. Nutrients. 12:122020. View Article : Google Scholar
|
|
11
|
Ahluwalia B, Moraes L, Magnusson MK and
Öhman L: Immunopathogenesis of inflammatory bowel disease and
mechanisms of biological therapies. Scand J Gastroenterol.
53:379–389. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chapman TP, Gomes CF, Louis E, Colombel JF
and Satsangi J: De-escalation of immunomodulator and biological
therapy in inflammatory bowel disease. Lancet Gastroenterol
Hepatol. 5:63–79. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fuller MK: Pediatric inflammatory bowel
disease: Special considerations. Surg Clin North Am. 99:1177–1183.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Colombel JF, D'haens G, Lee WJ, Petersson
J and Panaccione R: Outcomes and strategies to support a
treat-to-target approach in inflammatory bowel disease: A
Systematic Review. J Crohn's Colitis. 14:254–266. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cosnes J, Nion-Larmurier I, Beaugerie L,
Afchain P, Tiret E and Gendre JP: Impact of the increasing use of
immunosuppressants in Crohn's disease on the need for intestinal
surgery. Gut. 54:237–241. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Speca S, Giusti I, Rieder F and Latella G:
Cellular and molecular mechanisms of intestinal fibrosis. World J
Gastroenterol. 18:3635–3661. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rieder F and Fiocchi C: Intestinal
fibrosis in IBD - a dynamic, multifactorial process. Nat Rev
Gastroenterol Hepatol. 6:228–235. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Holvoet T, Devriese S, Castermans K,
Boland S, Leysen D, Vandewynckel YP, Devisscher L, Van den Bossche
L, Van Welden S, Dullaers M, et al: Treatment of intestinal
fibrosis in experimental inflammatory bowel disease by the
pleiotropic actions of a local rho kinase inhibitor.
Gastroenterology. 153:1054–1067. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang N, Liu C, Sun TM, Ran XK, Kang TG
and Dou DQ: Two new compounds from Atractylodes macrocephala
with neuroprotective activity. J Asian Nat Prod Res. 19:35–41.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao H, Ji ZH, Liu C and Yu XY:
Neuroprotection and mechanisms of atractylenolide III in preventing
learning and memory impairment induced by chronic high-dose
homocysteine administration in rats. Neuroscience. 290:485–491.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kang TH, Han NR, Kim HM and Jeong HJ:
Blockade of IL-6 secretion pathway by the sesquiterpenoid
atractylenolide III. J Nat Prod. 74:223–227. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhou Y, Huang S, Wu F, Zheng Q, Zhang F,
Luo Y and Jian X: Atractylenolide III reduces depressive- and
anxiogenic-like behaviors in rat depression models. Neurosci Lett.
759:1360502021. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang M, Hu R, Wang Y, Liu L, You H, Zhang
J, Wu X, Pei T, Wang F, Lu L, et al: Atractylenolide III attenuates
muscle wasting in chronic kidney disease via the oxidative
stress-mediated PI3K/AKT/mTOR pathway. Oxid Med Cell Longev.
2019:18754712019.PubMed/NCBI
|
|
24
|
Huai B and Ding J: Atractylenolide III
attenuates bleomycin-induced experimental pulmonary fibrosis and
oxidative stress in rat model via Nrf2/NQO1/HO-1 pathway
activation. Immunopharmacol Immunotoxicol. 42:436–444. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fu J, Ke X, Tan S, Liu T, Wang S, Ma J and
Lu H: The natural compound codonolactone attenuates TGF-β1-mediated
epithelial-to-mesenchymal transition and motility of breast cancer
cells. Oncol Rep. 35:117–126. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Song MY, Jung HW, Kang SY and Park YK:
Atractylenolide III enhances energy metabolism by increasing the
SIRT-1 and PGC1α expression with AMPK phosphorylation in C2C12
mouse skeletal muscle cells. Biol Pharm Bull. 40:339–344. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li N, Wang Z, Sun T, Lei Y, Liu X and Li
Z: Apigenin alleviates renal fibroblast activation through AMPK and
ERK signaling pathways in vitro. Curr Pharm Biotechnol.
21:1107–1118. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang JY, Tao LJ, Liu B, You XY, Zhang CF,
Xie HF and Li RS: Wedelolactone attenuates pulmonary fibrosis
partly through activating AMPK and regulating Raf-MAPKs signaling
pathway. Front Pharmacol. 10:1512019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shi J, Sun S, Liao Y, Tang J, Xu X, Qin B,
Qin C, Peng L, Luo M, Bai L, et al: Advanced oxidation protein
products induce G1 phase arrest in intestinal epithelial cells via
a RAGE/CD36-JNK-p27kip1 mediated pathway. Redox Biol.
25:1011962019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang BL, Zhu P, Li YR, Xu MM, Wang H, Qiao
LC, Xu HX and Chen HJ: Total flavone of Abelmoschus manihot
suppresses epithelial-mesenchymal transition via interfering
transforming growth factor-β1 signaling in Crohn's disease
intestinal fibrosis. World J Gastroenterol. 24:3414–3425. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang HJ, Zhang YN, Zhou H, Guan L, Li Y
and Sun MJ: IL-17A promotes initiation and development of
intestinal fibrosis through EMT. Dig Dis Sci. 63:2898–2909. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gong WX, Zhou YZ, Qin XM and Du GH:
Involvement of mitochondrial apoptotic pathway and MAPKs/NF-κB
inflammatory pathway in the neuroprotective effect of
atractylenolide III in corticosterone-induced PC12 cells. Chin J
Nat Med. 17:264–274. 2019.PubMed/NCBI
|
|
33
|
Greenburg G and Hay ED: Epithelia
suspended in collagen gels can lose polarity and express
characteristics of migrating mesenchymal cells. J Cell Biol.
95:333–339. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
López-Novoa JM and Nieto MA: Inflammation
and EMT: An alliance towards organ fibrosis and cancer progression.
EMBO Mol Med. 1:303–314. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang E, Liu S, Xu Z, Huang S, Tan X, Sun
C and Lu L: Pituitary tumor-transforming gene 1 (PTTG1) is
overexpressed in oral squamous cell carcinoma (OSCC) and promotes
migration, invasion and epithelial-mesenchymal transition (EMT) in
SCC15 cells. Tumour Biol. 35:8801–8811. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Aiello NM, Maddipati R, Norgard RJ, Balli
D, Li J, Yuan S, Yamazoe T, Black T, Sahmoud A, Furth EE, et al:
EMT subtype influences epithelial plasticity and mode of cell
migration. Dev Cell. 45:681–695.e4. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Flier SN, Tanjore H, Kokkotou EG, Sugimoto
H, Zeisberg M and Kalluri R: Identification of epithelial to
mesenchymal transition as a novel source of fibroblasts in
intestinal fibrosis. J Biol Chem. 285:20202–20212. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mege D and Panis Y: Unmet therapeutic
needs: Focus on intestinal fibrosis surgical approach: Resection,
strictureplasty and others. Dig Dis. 35:38–44. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Noble PW, Albera C, Bradford WZ, Costabel
U, Glassberg MK, Kardatzke D, King TE Jr, Lancaster L, Sahn SA,
Szwarcberg J, et al CAPACITY Study Group, : Pirfenidone in patients
with idiopathic pulmonary fibrosis (CAPACITY): Two randomised
trials. Lancet. 377:1760–1769. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kuhn A, Haust M, Ruland V, Weber R, Verde
P, Felder G, Ohmann C, Gensch K and Ruzicka T: Effect of bosentan
on skin fibrosis in patients with systemic sclerosis: A
prospective, open-label, non-comparative trial. Rheumatology
(Oxford). 49:1336–1345. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
King TE Jr, Brown KK, Raghu G, du Bois RM,
Lynch DA, Martinez F, Valeyre D, Leconte I, Morganti A, Roux S, et
al: BUILD-3: A randomized, controlled trial of bosentan in
idiopathic pulmonary fibrosis. Am J Respir Crit Care Med.
184:92–99. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Munger JS, Harpel JG, Gleizes PE, Mazzieri
R, Nunes I and Rifkin DB: Latent transforming growth factor-beta:
Structural features and mechanisms of activation. Kidney Int.
51:1376–1382. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Vallance BA, Gunawan MI, Hewlett B, Bercik
P, Van Kampen C, Galeazzi F, Sime PJ, Gauldie J and Collins SM:
TGF-beta1 gene transfer to the mouse colon leads to intestinal
fibrosis. Am J Physiol Gastrointest Liver Physiol. 289:G116–G128.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Salminen A and Kaarniranta K:
AMP-activated protein kinase (AMPK) controls the aging process via
an integrated signaling network. Ageing Res Rev. 11:230–241. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Morikawa M, Derynck R and Miyazono K:
TGF-β and the TGF-β family: Context-dependent roles in cell and
tissue physiology. Cold Spring Harb Perspect Biol. 8:82016.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Baruch RR, Melinscak H, Lo J, Liu Y, Yeung
O and Hurta RA: Altered matrix metalloproteinase expression
associated with oncogene-mediated cellular transformation and
metastasis formation. Cell Biol Int. 25:411–420. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wong SHM, Fang CM, Chuah LH, Leong CO and
Ngai SC: E-cadherin: Its dysregulation in carcinogenesis and
clinical implications. Crit Rev Oncol Hematol. 121:11–22. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hisada M, Hiranuma M, Nakashima M, Goda N,
Tenno T and Hiroaki H: High dose of baicalin or baicalein can
reduce tight junction integrity by partly targeting the first PDZ
domain of zonula occludens-1 (ZO-1). Eur J Pharmacol.
887:1734362020. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang N: Vimentin and tumor diagnosis.
Zhonghua Bing Li Xue Za Zhi. 19:122–124. 1990.(In Chinese).
PubMed/NCBI
|
|
50
|
Hardie DG, Ross FA and Hawley SA: AMPK: A
nutrient and energy sensor that maintains energy homeostasis. Nat
Rev Mol Cell Biol. 13:251–262. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hecker L, Logsdon NJ, Kurundkar D,
Kurundkar A, Bernard K, Hock T, Meldrum E, Sanders YY and
Thannickal VJ: Reversal of persistent fibrosis in aging by
targeting Nox4-Nrf2 redox imbalance. Sci Transl Med. 6:231ra472014.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Burkewitz K, Zhang Y and Mair WB: AMPK at
the nexus of energetics and aging. Cell Metab. 20:10–25. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Park CS, Bang BR, Kwon HS, Moon KA, Kim
TB, Lee KY, Moon HB and Cho YS: Metformin reduces airway
inflammation and remodeling via activation of AMP-activated protein
kinase. Biochem Pharmacol. 84:1660–1670. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Liu Z, Bone N, Jiang S, Park DW, Tadie JM,
Deshane J, Rodriguez CA, Pittet JF, Abraham E and Zmijewski JW:
AMP-activated protein kinase and glycogen synthase kinase 3β
modulate the severity of sepsis-induced lung injury. Mol Med.
21:937–950. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Sato N, Takasaka N, Yoshida M, Tsubouchi
K, Minagawa S, Araya J, Saito N, Fujita Y, Kurita Y, Kobayashi K,
et al: Metformin attenuates lung fibrosis development via NOX4
suppression. Respir Res. 17:1072016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Rangarajan S, Bone NB, Zmijewska AA, Jiang
S, Park DW, Bernard K, Locy ML, Ravi S, Deshane J, Mannon RB, et
al: Metformin reverses established lung fibrosis in a bleomycin
model. Nat Med. 24:1121–1127. 2018. View Article : Google Scholar : PubMed/NCBI
|