Introduction
In recent years, diabetic encephalopathy has become
a common but often overlooked chronic diabetic complication in
clinical practice, and cognitive dysfunction is its main clinical
characteristic (1,2). According to epidemiological data,
60–70% of patients with DM have mild or moderate cognitive
impairment (MCI), which significantly increases the risk of
dementia and imposes a heavy burden on the families of patients and
society (3,4). Previous studies have shown that as
the disease progresses, patients with diabetes gradually exhibit
cognitive impairments, such as declining learning and memory
ability, and spatial cognitive ability (5,6).
It has been previously reported that the incidence of cognitive
impairment in patients with diabetes was twice that in a normal
control group (7). Similar
pathological changes have been observed in rats with type 2
diabetes and Alzheimer's disease (8).
At present, the pathogenesis of diabetic cognitive
dysfunction remains unclear; however, it is considered to be
related to numerous factors, such as abnormal glucose and lipid
metabolism, oxidative stress, cerebrovascular disease and neuronal
dysfunction (9,10). The hippocampus is an important
anatomical structure associated with learning and memory. Previous
studies have confirmed that diabetic hyperglycemia may lead to the
degeneration, dysfunction and even apoptosis of hippocampal cells,
and is one of most the important factors of diabetic cognitive
impairment (11,12). The endoplasmic reticulum (ER) is
involved in an apoptosis pathway that differs from the
mitochondrial and death receptor pathways; the core of this pathway
is ER stress (ERS). Notably, the role of ERS in the occurrence and
development of neurological diseases has attracted wide attention
(13,14).
Exenatide, a synthetic derivative of exendin-4, is a
novel glucose-lowering drug. Exenatide is a glucagon-like peptide
similar to GLP-1 that can bind to and activate the GLP-1 receptor
(GLP-1R), and exert biological effects through cyclic adenosine
phosphate or other signal transduction pathways. Previous studies
have shown that GLP-1 can reduce blood glucose in patients with
type 2 diabetes, improve dysfunctional lipid metabolism, increase
the sensitivity of target tissues and target organs to insulin, and
reduce insulin resistance (15,16). Moreover, the GLP-1R agonist
exenatide has been reported to ameliorate myocardial fibrosis in
diabetic rats (17,18). It has been demonstrated that GLP-1
and GLP-1Rs are widely expressed throughout the central nervous
system, suggesting that the GLP-1/GLP-1R pathway may be involved in
regulation of central nervous system function (17,18). The GLP-1/GLP-1 R pathway may also
be involved in the role of central insulin and in the regulation of
learning and memory functions (19). Furthermore, GLP-1 may exert
neurotrophic and neuroprotective effects that are closely related
to the plasticity and survival of neurons (20). GLP-1 and its analogs, such as
exendin-4, can activate GLP-1Rs, upregulate the levels of cAMP in
neurons, regulate the homeostasis of intracellular calcium ions,
reduce the excitotoxicity of nerve cells, inhibit neuronal
apoptosis, and promote the proliferation and differentiation of
nerve cells (21). In addition,
GLP-1 has been reported to inhibit the apoptosis of hippocampal
nerve cells, as well as learning and memory dysfunction induced by
microwave exposure in rats, and to protect nerve cells (22). Diabetic encephalopathy is a
complication of diabetes and cognitive impairment is its main
manifestation. In the present study, the effect of exenatide, a
GLP-1 analog, on cognitive function in diabetic rats was examined
and the possible mechanism was explored from an antiapoptotic point
of view.
Materials and methods
Experimental animals and grouping
A total of 80 male Sprague-Dawley rats (age, 8
weeks; weight, 260–300 g) supplied by the North China University of
Science and Technology (Tangshan, China) were used in laboratory
experiments. Rats were housed in constant temperature (22–25°C) and
humidity (50–60%) under a 12-h light-dark cycle with free access to
food and water. After 1 week of acclimation, the rat model was
established, and access to food and water was unrestricted. All
animal procedures complied with the Regulations of the Experimental
Animal Administration (approval no. LAEC-NCST-20200002). The
research was approved by the Animal Ethics Committee of North China
University of Science and Technology (approval no.
LAEC-NCST-20200002).
The experiments were divided into two stages. First,
to investigate the possible protective mechanism of exenatide (cat.
no. Pep03601; NJPeptide) in a diabetes mellitus (DM) model rat, 60
rats were divided into four groups (n=15 rats/group): The control
(CON) group, the DM group, the exenatide + control (Ex + CON) group
and the exenatide + DM (Ex + DM) group. DM modeling was performed
as described previously (23).
The rats in the DM and Ex + DM groups received streptozotocin (STZ;
30 mg/kg; cat. no. ab142155; Abcam) by intraperitoneal injection
and were offered a high-sugar and high-fat diet (basic feed 59%,
lard 18%, sucrose 20%, egg yolk 3%; Shanghai R&S Biotechnology
Co., Ltd.) 4 weeks before STZ injection. The CON and Ex + CON
groups were given the same amount of citric acid buffer (through
i.p. injection) and fed routinely. The blood glucose of the rats
was measured, and a blood glucose level >16.67 mmol/l was
considered to indicate successful modeling. After establishing the
model, the Ex + CON and Ex + DM group rats were treated with
exenatide (3 µg/kg, b.i.d.), whereas the CON and DM group rats were
treated with the same quantity of physiological saline (through
i.p. injection). The second stage of experiments was conducted to
determine whether the JNK/c-JUN pathway contributed to the
protective effects of exenatide in DM using a specific inhibitor of
JNK (SP600125; cat. no. ab120065; Abcam). The rats were divided
into four groups (n=5 rats/group): The DM group, the SP600125 + DM
(SP + DM) group (treated with SP600125, 10 mg/kg/d; through i.p.
injection), the Ex + DM group (treated with exenatide, 3 µg/kg,
b.i.d.), and the SP + DM + Ex group (treated with SP600125, 10
mg/kg/d and exenatide, 3 µg/kg/d through i.p. injection). DM
modeling was performed as aforementioned. After 16 weeks of
continuous subcutaneous injection of exenatide, body weight, were
measured. Blood was collected from the tail vein (5 µl) to measure
blood glucose with a Glucometer (Sannuo Biosensing Co., Ltd.) and
blood samples (5 ml) from the heart were taken to detect blood
lipids and insulin levels.
Morris water maze (MWM) test
Water was added to a pool (60 cm in height and 120
cm in diameter) to a depth of 30 cm and a platform was placed 1 cm
below the surface of the water. The experiment lasted for 6 days
following the 16 week treatment period. The first 5 days were
considered the training period, and the test was performed on day
6. During the training period, a quadrant was randomly selected as
the entry point, the rats were gently placed into the water facing
the pool wall, and the time from entering the water to climbing
onto the platform (i.e., the escape latency) was recorded. If the
platform was not found within 60 sec, the rat was led to the
platform and placed on the platform for 30 sec and the escape
latency was recorded as 60 sec. On day 6, the platform was removed,
and time spent in the platform quadrant and swimming speed were
recorded. The experimental animals, except for those used in
histological experiments, were sacrificed by cervical dislocation
after anesthesia (pentobarbital, 60 mg/kg).
Hematoxylin and eosin (H&E)
staining
After the behavioral experiment, 5 rats in each
group were intraperitoneally injected with pentobarbital (60 mg/kg;
cat. no. H31020502; Shanghai Xinya Pharmaceutical Co., Ltd.) for
deep anesthesia. Cardiac perfusion with 4% paraformaldehyde (pH
7.4) was performed on rats. Subsequently, the brain was collected
and transferred to 4% paraformaldehyde and kept overnight at 4°C.
The brain tissue was washed with tap water, dehydrated with an
ethanol gradient, vitrified with dimethylbenzene and immersed in
wax. Brain tissues were sectioned (5 µm) and used for H&E
staining at 37°C. 5% eosin solution (5%) was added for 10 min,
after which samples were rinsed with purified water and redyed for
2 min. The sections were observed under a light microscope. Using
an a light microscope, the H&E-stained specimens in each field
of view at ×400 magnification were subjected to morphometric
analysis. The number of intact nerve cells in each field of view in
each part of the hippocampus was counted, and 10 fields of view
were counted for each specimen. The number of nerve cells in
different parts of the hippocampus is expressed as the number of
cells per field.
Reverse transcription-quantitative PCR
(RT-qPCR) detection
Trizol (cat. no. R0016; Beyotime Institute of
Biotechnology) was used to extract total RNA from hippocampal
tissues, and RT was performed at 50°C for 30 min (BeyoFast™
SYBR-Green One-Step qRT-PCR Kit; cat. no. D7268M; Beyotime
Institute of Biotechnology). According to the instructions of the
qPCR kit (cat. no. D7268M; Beyotime Institute of Biotechnology), a
20-µl reaction system was prepared. The primer sequences were
designed with the help of Primer 5.0 software (Primer Premier) as
follows: Caspase-3, forward, 5′-CCGAAACTCTTCATCATTCAGGC-3′,
reverse, 5′-GTTCCACTGTCTGTCTCAATACCG-3′; cytochrome c
(Cyt-c), forward, 5′-AAAAGGAGGCAAGCATAAGACTG-3′, reverse,
5′-CTTGTTGGCATCTGTGTAAGAGAATC-3′; and β-actin, forward,
5′-CAACCGTGAAAAGATGACCCAGAT-3′, reverse,
5′-CAACCGTGAAAAGATGACCCAGAT-3′. The reaction conditions were as
follows: Pre-denaturation at 95°C for 4 min; followed by 40 cycles
of denaturation at 95°C for 30 sec and then annealing/extension at
65°C for 30 sec. The fluorescence signal was detected at the end of
each annealing cycle, and the critical point of detection was set
as the cycle quantification value. Dissolution curve analysis was
subsequently performed under the following conditions: 95°C for 15
sec; 60°C for 20 sec; 95°C for 15 sec. The 2−∆∆ Cq
method was used to calculate the relative changes of gene
expression (24).
TUNEL staining
The level of apoptosis in each group was assessed
using an In Situ Cell Death Detection Kit (cat. no.
Roche-11684817910; Merck), according to the manufacturer's
instructions. A total of 10 high magnification fields
(magnification, ×400) were randomly counted for each section, which
were prepared as previously outlined, using a light microscope, and
the apoptotic index was calculated by dividing the number of
TUNEL-positive cells by the total number of cells counted. Each
sample was tested three times and the average was obtained.
Detection of Caspase-3 activity
The isolated brain tissue of rats were placed on an
ice plate and was homogenized in an ice bath. The homogenate was
transferred to a 1.5-ml centrifuge tube and was incubated in an ice
bath for 5 min. Subsequently, the homogenate was centrifuged at
13,000 × g (RCF) for 10–15 min at 4°C, and the supernatant was
collected for later use. Caspase-3 activity was tested using the
Caspase-3 kit (cat. no. C1116; Beyotime Institute of
Biotechnology). Notably, Caspase-3 can catalyze production of
yellow pNA from the substrate Ac-DEVD-pNA and yellow pNA has a
strong absorption peak near the wavelength of 405 nm; therefore,
Caspase-3 activity can be detected by measuring the absorbance
[optical density (OD)]. The calculation formula of Caspase-3
activity was as follows: Caspase-3 activity={[(OD sample-OD
blank)/OD blank] ×1,000}/(sample protein concentration × sampling
liquid).
Western blotting
Hippocampal tissue was sliced into fragments and
lysed with lysate to obtain total protein (cat. no. P0013E-1;
Beyotime Institute of Biotechnology) and the protein concentration
was analyzed using the Bradford method. Protein (50 µg) was loaded
and separated using a precast 12% SDS-PAGE gels. Subsequently, the
PVDF membrane was blocked in 5% skim milk powder solution at 37°C
for 2 h. Caspase-3 (1:1,000; cat. no. ab208161), JNK (1:1,000; cat.
no. ab179461), phosphorylated (p)-JNK (1:2,000; cat. no. ab219584),
c-JUN (1:1,000; cat. no. ab40766), p-c-JUN (1:1,000; cat. no.
ab227533), Cyt-c (1:1,000; cat. no. ab133504) and GAPDH (1:2,000;
cat. no. ab8245) antibodies (all from Abcam) were added to the
membrane and incubated at 4°C overnight. After washing the
membrane, horseradish peroxidase-labeled IgG secondary antibodies
(1:5,000; cat. no. A0208; Beyotime Institute of Biotechnology) were
added and incubated at 37°C for 2 h. After washing the membrane,
BeyoECL Moon (Beyotime Institute of Biotechnology) reagent was
added, and the protein bands were semi-quantified with LabWorks 4.5
software (Analytik Jena AG). The ratio of target protein band to
internal reference band (GAPDH) was calculated.
Statistical analysis
The experimental results are shown as the mean ± SD,
and data were analyzed using SPSS 23.0 software (IBM Corp.). The
results of the MWM test were analyzed by mixed ANOVA. The
differences in other parameters were analyzed by one-way ANOVA, and
Tukey's test was used for post-hoc comparisons. P<0.05 was
considered to indicate a statistically significant difference.
Results
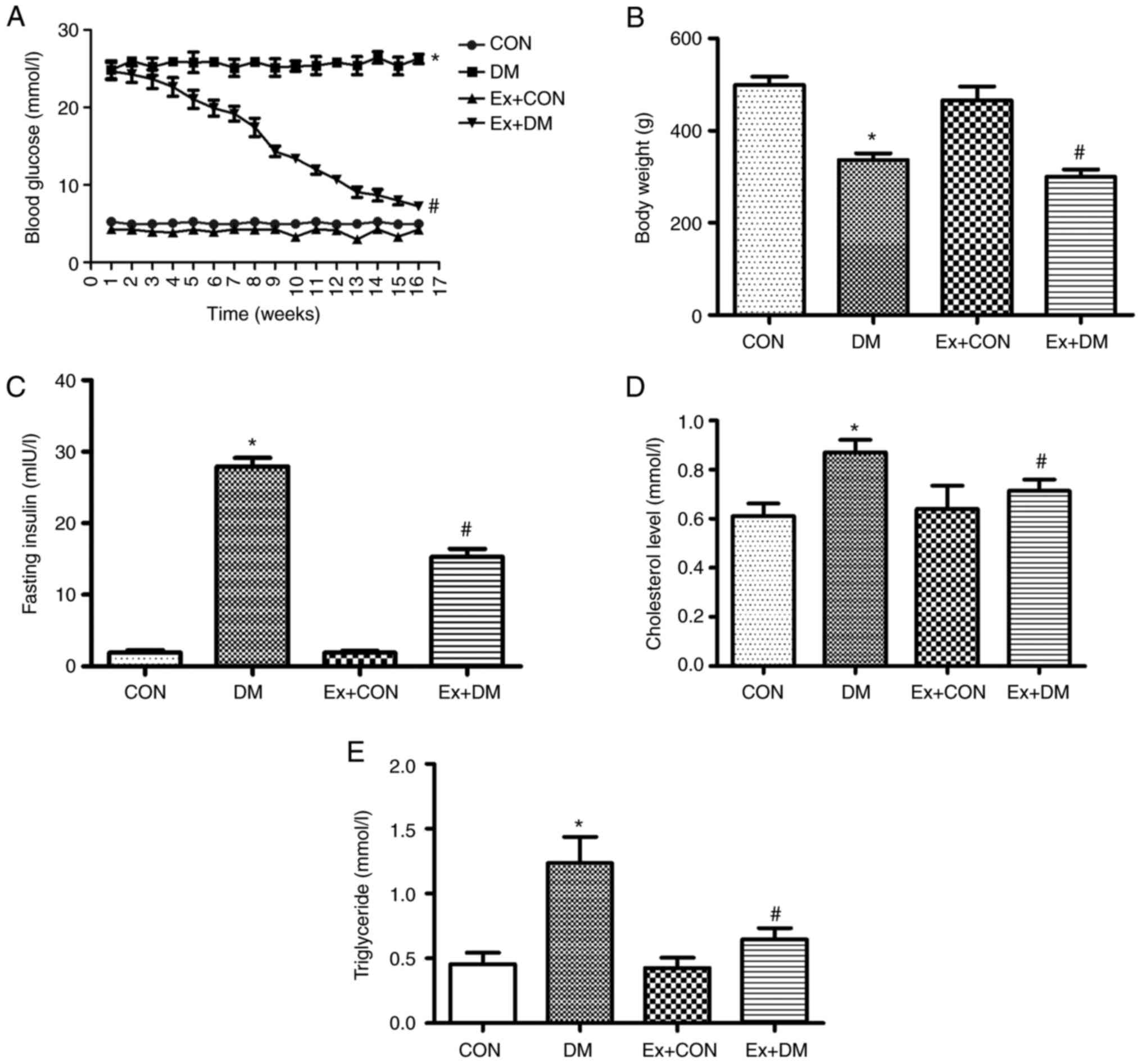
Effects of exenatide on blood glucose,
body weight and insulin in DM model rats
After the 16-week experiment, the blood glucose of
diabetic rats was significantly higher compared with that in the
CON group (Fig. 1A). Compared
with that in the DM group, the blood glucose of the Ex + DM group
exhibited a downward trend (P=0.0013; Fig. 1A). Blood glucose in the first week
was not baseline data; therefore, blood glucose in the control and
model groups were not the same. Compared with the body weight of
the CON rats, the weight of DM model rats was significantly
decreased (P=0.0019; Fig. 1B).
Following treatment with exenatide, the body weight of rats in the
Ex + DM group was significantly decreased compared with that of
rats in the DM group (P=0.0021; Fig.
1B). Moreover, rats in the DM group displayed higher insulin
levels compared with those in the CON group (P=0.0008), whereas the
insulin levels after exenatide treatment showed a significant
decrease (P=0.0014; Fig. 1C).
These results suggested that exenatide improved blood glucose
regulation and reduced body weight and fasting insulin levels.
Moreover, the rats in the DM group presented higher cholesterol
(P=0.0102; Fig. 1D) and
triglyceride (P=0.0022; Fig. 1E)
levels compared with those of the rats in the CON group; however,
the serum levels of cholesterol (P=0.0007; Fig. 1D) and triglyceride (P=0.0036;
Fig. 1E) were reduced following
exenatide treatment. The results showed that exenatide effectively
regulated lipid levels in the body, thereby reducing the incidence
of cardiovascular disease.
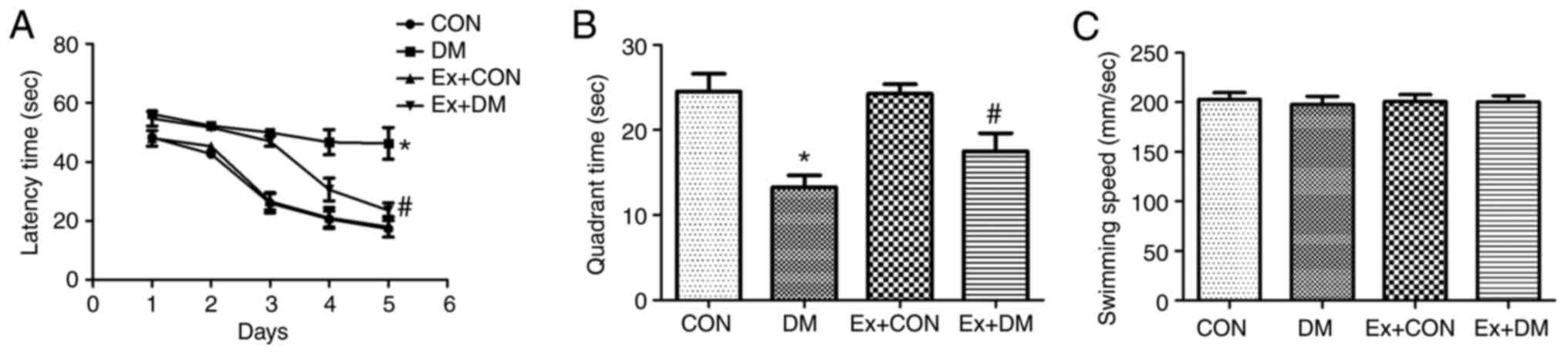
Effect of exenatide on learning and
memory ability in rats with DM
To investigate whether exenatide could ameliorate
the spatial memory impairment of rats with DM, the MWM hidden
platform task was performed to assess hippocampus-dependent
cognitive capacity. The effects of exenatide on learning and memory
ability during latency trials are shown in Fig. 2A. The DM model rats required more
time to find the hidden platform on days 1–5 (P=0.0019, P=0.0011,
P=0.0010, P=0.0009 and P=0.0007, respectively); however, the Ex +
DM rats displayed a significantly shorter latency time than the DM
rats (P=0.0053, P=0.0049, P=0.0051, P=0.0037 and P=0.0026,
respectively; Fig. 2A). In the
probe test, the platform was removed. In DM rats, the time spent in
the target quadrant was significantly decreased (P=0.0017; Fig. 2B); however, rats in the Ex + DM
group spent more time in the goal quadrant compared with those in
the DM group (P=0.0032; Fig. 2B).
Nevertheless, there was no significant difference in swimming speed
among the groups (P=0.0875; Fig.
2C). These findings are consistent with those of a previous
study (25) and indicated that
exenatide may be suitable for treatment of DM-induced memory
deficits.
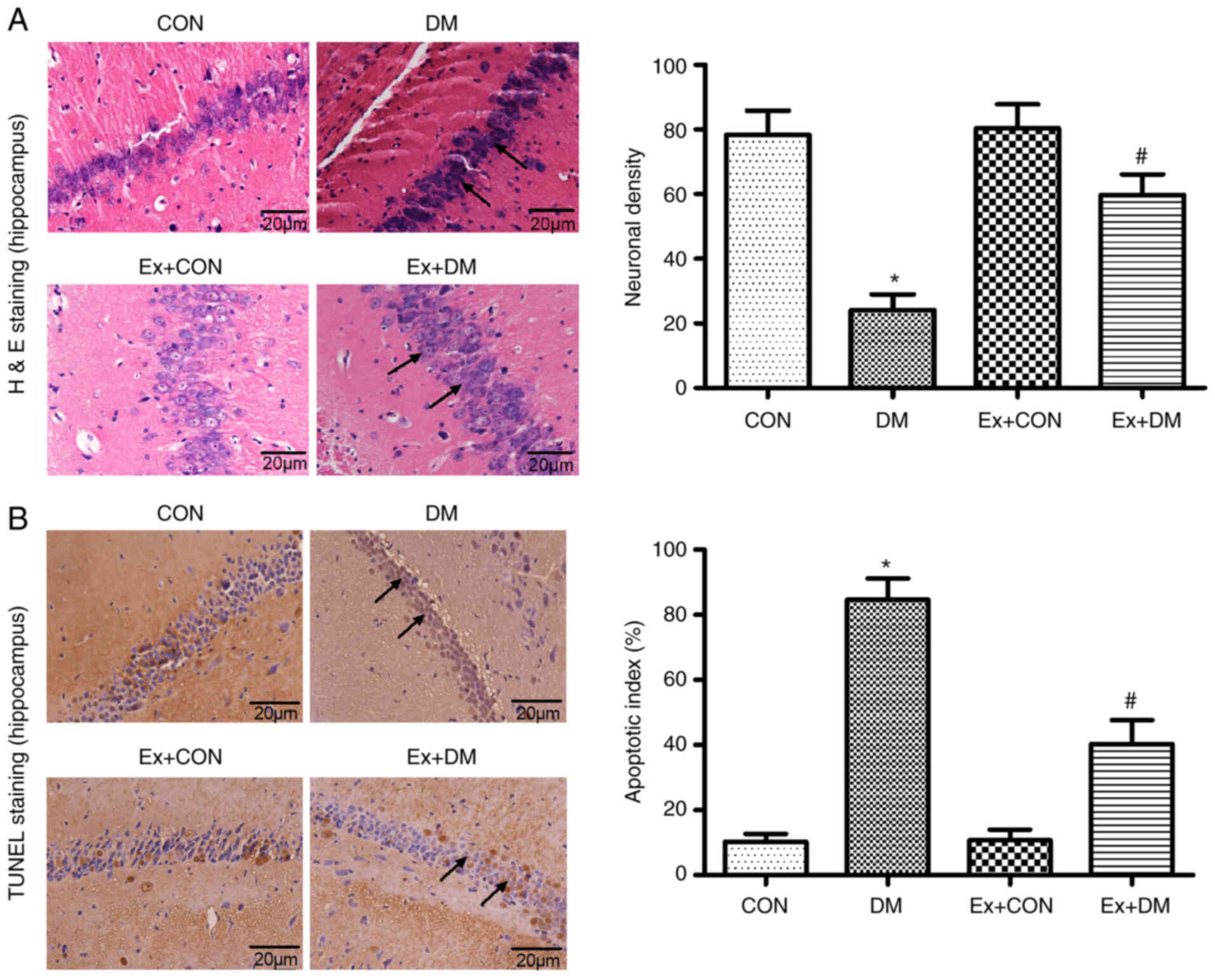
Effect of exenatide on hippocampal
neuron morphology in rats with DM
10 high magnification fields were selected for each
section, the number of positive cells was counted and the average
value was calculated. Morphological changes in hippocampal neurons
were observed by H&E staining among the groups. Karyopyknosis
and nucleoli disappearance were observed in the hippocampal neurons
of rats in the DM group (P=0.0012; Fig. 3A). In the DM model rats treated
with exenatide, the hippocampal neuron structure was relatively
intact, the number of neurons was higher than that in rats in the
DM group, and the cytoplasmic shrinkage was recovered (Fig. 3A). These results suggested that
exenatide exhibited protective effects on the morphology of
hippocampal neurons.
Effect of exenatide on apoptosis of
hippocampal neurons
The anti-apoptotic effects of exenatide on diabetic
rats was investigated. The ratio of apoptotic cells to total cell
number was calculated following TUNEL staining, and the results
revealed that there were more apoptotic cells in the DM group
compared with in the CON group (P=0.0025); however, neuronal
apoptosis was significantly alleviated after exenatide
administration (P=0.0017; Fig.
3B). The slight difference between the CON group and the Ex +
CON group was not statistically significant (P=0.0981).
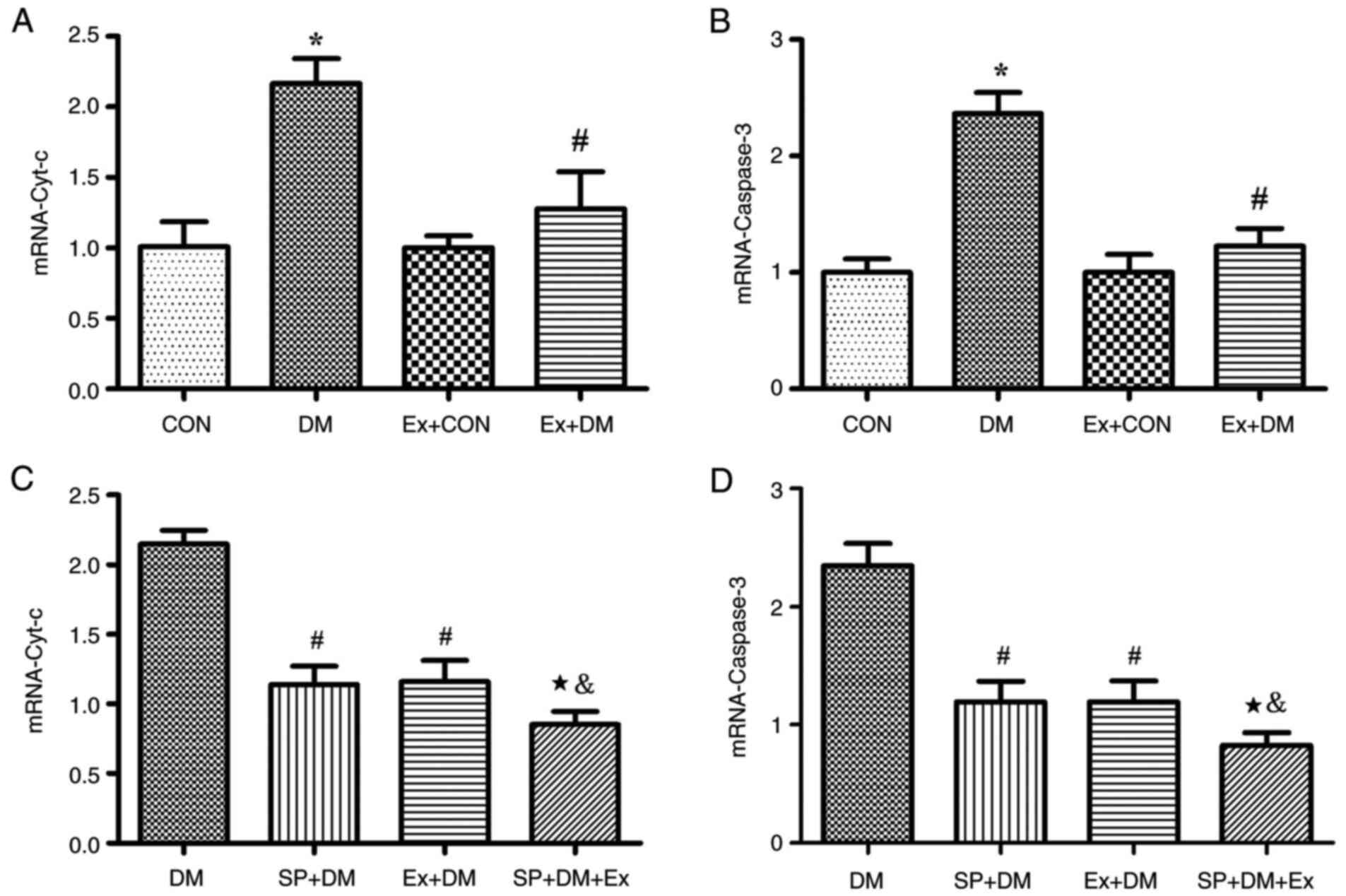
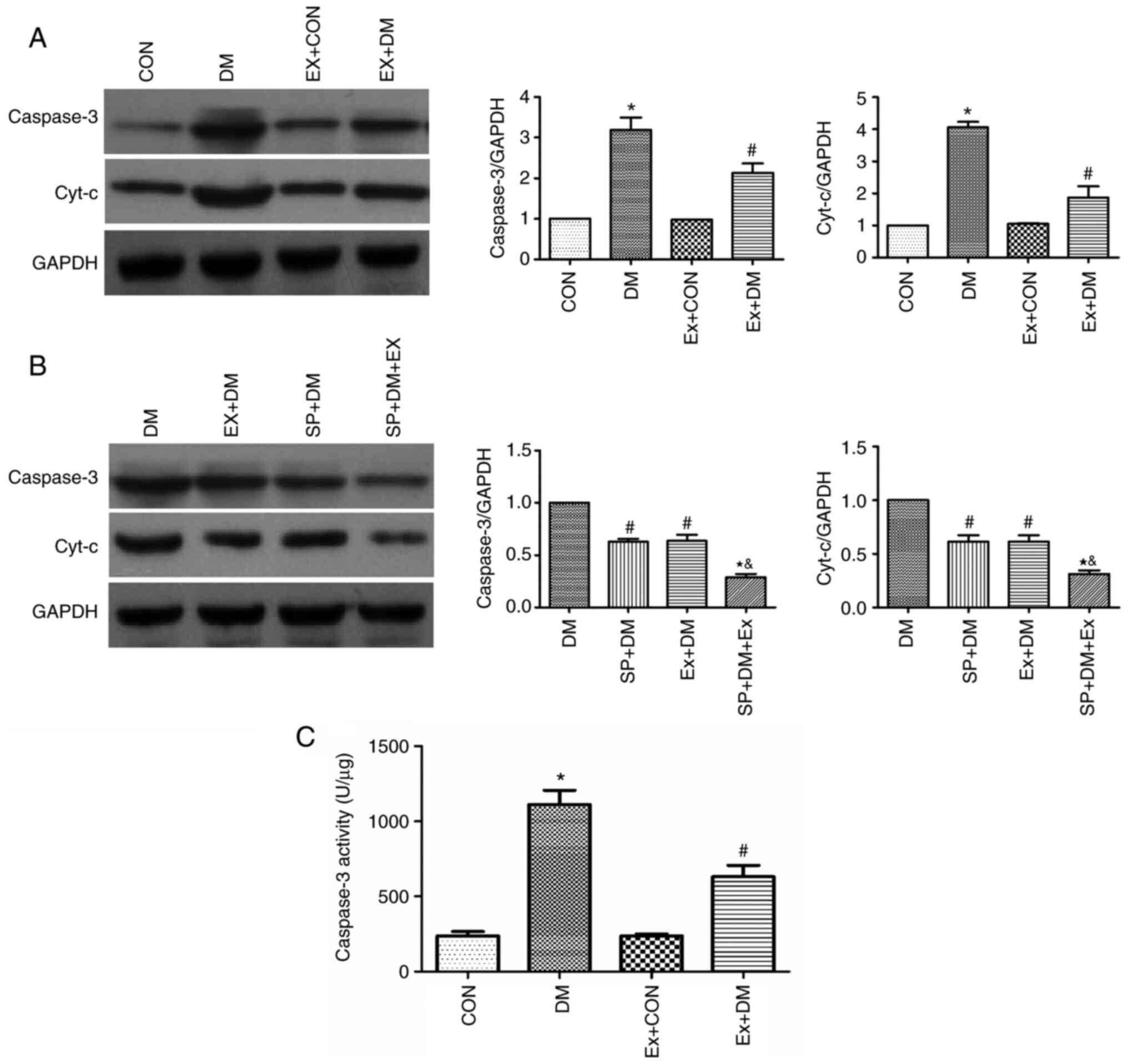
Moreover, the results of RT-qPCR revealed that the
mRNA expression levels of Cyt-c (P=0.0011; Fig. 4A) and Caspase-3 (P=0.0018;
Fig. 4B) were higher in the DM
group compared with those in the CON group. Exenatide lowered the
mRNA expression of Cyt-c and Caspase-3. Furthermore, western
blotting was performed to detect the expression levels of the
apoptosis-related proteins Caspase-3 and Cyt-c in hippocampal
tissues. As shown in Fig. 5A,
compared with those in the DM group, the expression levels of
Caspase-3 and Cyt-c were significantly decreased in the Ex + DM
group. mRNA (P=0.0008, P=0.0014; Fig.
4C and D) and protein (P=0.0021, P=0.0013; Fig. 5B) levels of Cyt-c and Caspase-3
were significantly decreased in diabetic rats treated with
SP600125. The results suggested that exenatide could inhibit
neuronal apoptosis induced by diabetes.
Effect of exenatide on Caspase-3
activity in diabetic rats
Caspase-3 activity in the DM group was significantly
higher compared with that in the CON group (P=0.0016; Fig. 5C). These results indicated that
diabetes could lead to enhanced Caspase-3 activity. By contrast,
Caspase-3 activity was significantly decreased in diabetic rats
after exenatide administration (P=0.0020; Fig. 5C), indicating that exenatide could
significantly inhibit the diabetes-induced increase in Caspase-3
activity.
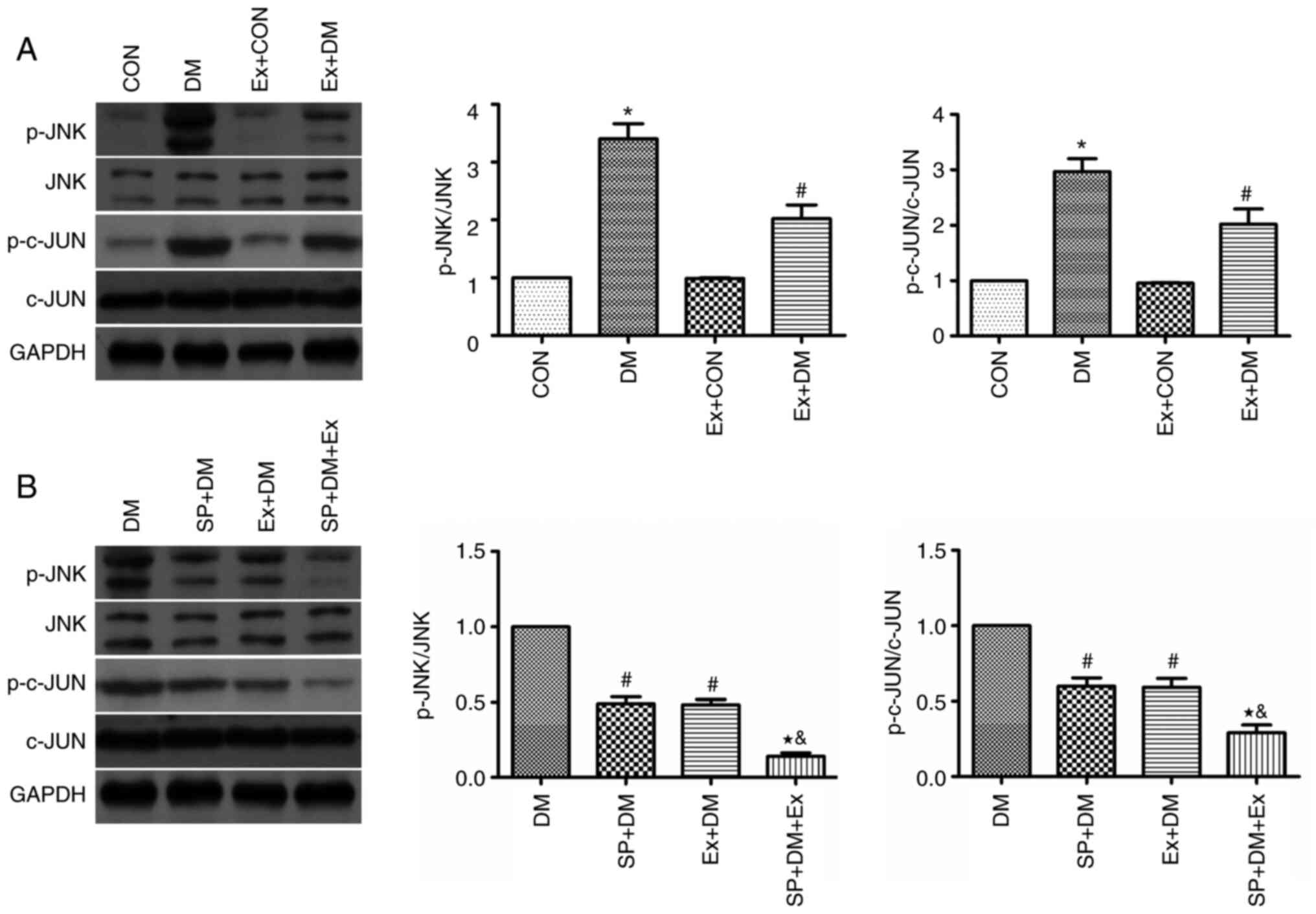
Effects of exenatide on the JNK/c-JUN
signaling pathway
The protein expression levels of JNK, p-JNK, c-JUN
and p-c-JUN were detected in hippocampal tissues by western
blotting. The protein expression levels of p-JNK and p-c-JUN were
significantly higher in the DM group compared with those in the CON
group (P=0.0048 and P=0.0011, respectively; Fig. 6A). Conversely, the expression
levels of these proteins were significantly reduced in the Ex + DM
group (P=0.0051 and P=0.0043, respectively; Fig. 6A). The findings indicated that
exenatide may inhibit neuronal apoptosis via the JNK/c-JUN
signaling pathway.
To further explore the protective effect of
exenatide on the ERS signaling pathway, rats were treated with a
specific inhibitor of JNK (SP600125) 30 min before modeling and the
changes in p-JNK and p-c-JUN expression were detected. Compared
with those in the DM group, the protein expression levels of p-JNK
and p-c-JUN were significantly decreased in the SP + DM group
(P=0.0023 and P=0.0082, respectively; Fig. 6B). Notably, p-JNK and p-c-JUN were
expressed at similar levels in the Ex+DM and SP+DM groups,
suggesting that exenatide and SP600125 have similar inhibitory
effects on the JNK pathway. This conclusion was further verified in
the SP + DM + Ex group, as p-JNK and p-c-JUN were expressed at half
the level of the Ex+DM and SP+DM group. These results indicated
that exenatide may reduce neuronal apoptosis in rats with diabetes
by inhibiting JNK/c-JUN signaling.
Discussion
Diabetic encephalopathy is characterized by acquired
cognitive and behavioral deficits, and includes pathological,
morphological, electrophysiological, neurobiochemical,
neuropsychological and behavioral changes. Studies have shown that
diabetes can cause MCI, which is a state between normal cognitive
function and dementia (26,27). The earliest and largest
cross-sectional study of the association between diabetes and
dementia revealed that the relative risk values of diabetes with
Alzheimer's disease and vascular dementia were as high as 1.3-2.2
and 2.0-3.4, respectively (28,29).
In patients with diabetes, insufficient insulin
secretion and insulin resistance can lead to hyperglycemia.
Increased blood sugar levels can accelerate the onset of
Alzheimer's disease, with diabetes being closely associated with
Alzheimer's disease (30).
Epidemiological studies have revealed that elderly individuals with
diabetes have twice the risk of developing cognitive dysfunction
and other dementia symptoms compared with the risk among the
general population, and diabetes is closely associated with
Alzheimer's disease (31,32). Cognitive impairment associated
with diabetes causes a great inconvenience for patients; therefore,
it is of great clinical significance to carry out research with the
aim of preventing cognitive impairment in diabetes. In the
treatment of type 2 diabetes, exenatide is effective in controlling
blood glucose, improving the function of pancreatic β cells,
increasing insulin sensitivity, reducing body mass in patients with
obesity, and regulating inflammatory mediators and lipids, thereby
reducing the incidence of cardiovascular disease, hypoglycemia and
pancreatitis (33). In a rat
model of Alzheimer's disease, GLP-1 reduced endogenous β-amyloid
levels and inhibited the Aβ-induced apoptosis of nerve cells
(34). GLP-1 and its long-acting
counterpart, exendin-4, have also been reported to inhibit
glutamate-induced apoptosis of hippocampal neurons; however, the
specific mechanism by which GLP-1 interferes with neuronal
apoptosis remains unclear (35).
JNK is a member of the MAPK family and serves an
important role in cell apoptosis, growth, differentiation,
embryonic development and immune responses. The JNK signaling
pathway can be activated by various cellular stimuli, such as
ultraviolet radiation, heat shock or inflammatory stimuli, through
the MAPKKK-MAPKK-MAPK (JNK) cascade system (36,37). c-JUN is a member of the HEG family
of fast-reacting genes and can serve a role in the activated
protein-1 (AP-1) complex to regulate cell proliferation,
differentiation and apoptosis (38). Under the action of JNK, Ser63 and
Ser73 in the N-terminal active region of JUN are phosphorylated,
self-translocation occurs, and the activity is upregulated to
promote the activity of AP-1; activated JUN further promotes the
expression of a variety of pro-apoptotic proteins (39). The results of the present study
revealed that, in STZ-induced diabetic rats with learning and
memory impairment, JNK and c-JUN proteins in the hippocampus were
phosphorylated, and exenatide could reduce the expression levels of
p-JNK and p-c-JUN, thus suggesting that the high glucose
environment in the body may activate the JNK/c-JUN signaling
pathway.
The metabolic dysfunction caused by the imbalance of
the endocrine system and the activation of a large number of
inflammatory mediators under stress can lead to the opening of the
mitochondrial permeability pores to release Cyt-c and activate
Caspase-3 to initiate the apoptosis program (40). Caspase-3 is an apoptotic factor
that can be activated by an upstream initiator. After activation,
Caspase-3 forms an apoptotic protein that acts on specific
substrates to cause morphological and biochemical changes in cells,
leading to apoptosis. Regulating blood sugar, inhibiting the
opening of mitochondrial permeability transition pores, and
reducing the release of Cyt-c, thereby blocking the apoptotic
pathway at the level of Caspase-3, can effectively protect
hippocampal neurons (41). The
results of the present study revealed that the expression levels of
Cyt-c and Caspase-3 in the hippocampus of rats with diabetic
cognitive dysfunction were significantly increased, suggesting that
the high-glycemic environment in the body may upregulate Cyt-c and
Caspase-3 expression levels, thereby inducing hippocampal neuron
apoptosis. By contrast, exenatide could inhibit the process and
reduce the expression levels of Cyt-c and Caspase-3. Currently,
exenatide is used to regulate blood sugar in patients with type 2
diabetes, improve insulin sensitivity and effectively control body
weight; however, the application of exenatide to reverse cognitive
impairment in diabetes is not very common. Therefore, more
experiments are required to confirm the effectiveness and safety of
exenatide for cognitive impairment. Moreover, exenatide treatment
of type 2 diabetes often results in adverse gastrointestinal
reactions, such as nausea, loss of appetite and vomiting (42). Therefore, further investigation is
required to improve the safety of exenatide therapy in clinical
practice.
In conclusion, in the process of cognitive
dysfunction caused by diabetes, along with activation of the
JNK/c-JUN signaling pathway, the expression levels of Cyt-c and
Caspase-3 are significantly increased. Therefore, it was
hypothesized that regulation of the JNK/c-JUN signaling pathway may
lead to the upregulation of the expression of apoptotic proteins,
which may be one of the factors involved in hippocampal neuronal
apoptosis leading to diabetic cognitive impairment. Notably,
exenatide inhibited the phosphorylation of JNK and c-JUN, and
downregulated the expression levels of Cyt-c and Caspase-3. These
findings indicated that the suppression of apoptosis-induced
diabetic cognitive impairment by exenatide may be achieved through
its inhibition of the JNK/c-JUN signaling pathway.
Acknowledgements
Not applicable.
Funding
The experiment was funded by Hebei Provincial Health Commission
(grant no. 20200130) and the Startup Foundation for Advanced
Talents (grant no. 2019GC18).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GW and MY confirm the authenticity of all the raw
data. GW and MY designed the experiments. ZZ and BR made
substantial contributions to acquisition of data. JL and WY made
contributions to the analysis of data. DS, LW and XZ performed the
experiments. All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of North China University of Science and Technology
(approval no. LAEC-NCST-20200002).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that there are no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
DM
|
diabetes mellitus
|
|
ER
|
endoplasmic reticulum
|
|
ERS
|
ER stress
|
|
MCI
|
mild or moderate cognitive
impairment
|
|
MWM
|
Morris water maze
|
References
|
1
|
Prieto-Gómez B, Díaz-Vázquez M and
Pérez-Torres D: Hippocampal electrophysiological changes during the
elicited metabolic syndrome in Wistar rats. Metabol Open.
5:1000272020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Meng Y, Wang W, Kang J, Wang X and Sun L:
Role of the PI3K/AKT signalling pathway in apoptotic cell death in
the cerebral cortex of streptozotocin-induced diabetic rats. Exp
Ther Med. 13:2417–2422. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Moran C, Beare R, Wang W, Callisaya M and
Srikanth V; Alzheimer's Disease Neuroimaging Initiative (ADNI), :
Type 2 diabetes mellitus, brain atrophy, and cognitive decline.
Neurology. 92:e823–e830. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chornenkyy Y, Wang WX, Wei A and Nelson
PT: Alzheimer's disease and type 2 diabetes mellitus are distinct
diseases with potential overlapping metabolic dysfunction upstream
of observed cognitive decline. Brain Pathol. 29:3–17. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hassing LB, Grant MD, Hofer SM, Pedersen
NL, Nilsson SE, Berg S, McClearn G and Johansson B: Type 2 diabetes
mellitus contributes to cognitive decline in old age: A
longitudinal population-based study. J Int Neuropsychol Soc.
10:599–607. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mayeda ER, Haan MN, Yaffe K, Kanaya AM and
Neuhaus J: Does type 2 diabetes increase rate of cognitive decline
in older Mexican Americans? Alzheimer Dis Assoc Disord. 29:206–212.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yanagawa M, Umegaki H, Makino T, Nakashima
H and Kuzuya M: Neuropsychological differences in Alzheimer's
disease patients with or without type 2 diabetes mellitus. Geriatr
Gerontol Int. 16:1232–1235. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Blalock EM, Phelps JT, Pancani T, Searcy
JL, Anderson KL, Gant JC, Popovic J, Avdiushko MG, Cohen DA, Chen
KC, et al: Effects of long-term pioglitazone treatment on
peripheral and central markers of aging. PLoS One. 5:e104052010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Duarte JM: Metabolic alterations
associated to brain dysfunction in diabetes. Aging Dis. 6:304–321.
2015.PubMed/NCBI
|
|
10
|
Calvo-Ochoa E and Arias C: Cellular and
metabolic alterations in the hippocampus caused by insulin
signalling dysfunction and its association with cognitive
impairment during aging and Alzheimer's disease: Studies in animal
models. Diabetes Metab Res Rev. 31:1–13. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ma P, Mao XY, Li XL, Ma Y, Qiao YD, Liu
ZQ, Zhou HH and Cao YG: Baicalin alleviates diabetes-associated
cognitive deficits via modulation of mitogen-activated protein
kinase signaling, brain-derived neurotrophic factor and apoptosis.
Mol Med Rep. 12:6377–6383. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sowndhararajan K, Deepa P, Kim M, Park SJ
and Kim S: Neuroprotective and cognitive enhancement potentials of
baicalin: A review. Brain Sci. 8:1042018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kaneko M: Molecular pharmacological
studies on the protection mechanism against endoplasmic reticulum
stress-induced neurodegenerative disease. Yakugaku Zasshi.
132:1437–1442. 2012.(In Japanese). View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nomura Y: Pharmacological studies on
neurodegenerative diseases focusing on refolding and degradation of
unfolded proteins in the endoplasmic reticulum. Yakugaku Zasshi.
134:537–543. 2014.(In Japanese). View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Deacon CF, Mannucci E and Ahrén B:
Glycaemic efficacy of glucagon-like peptide-1 receptor agonists and
dipeptidyl peptidase-4 inhibitors as add-on therapy to metformin in
subjects with type 2 diabetes-a review and meta analysis. Diabetes
Obes Metab. 14:762–767. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Calanna S, Christensen M, Holst JJ,
Laferrère B, Gluud LL, Vilsbøll T and Knop FK: Secretion of
glucose-dependent insulinotropic polypeptide in patients with type
2 diabetes: Systematic review and meta-analysis of clinical
studies. Diabetes Care. 36:3346–3352. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen WJY, Diamant M, de Boer K, Harms HJ,
Robbers LFHJ, van Rossum AC, Kramer MHH, Lammertsma AA and Knaapen
P: Effects of exenatide on cardiac function, perfusion, and
energetics in type 2 diabetic patients with cardiomyopathy: A
randomized controlled trial against insulin glargine. Cardiovasc
Diabetol. 16:672017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ravassa S, Zudaire A and Díez J: GLP-1 and
cardioprotection: from bench to bedside. Cardiovasc Res.
94:316–323. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang XH, Li L, Hölscher C, Pan YF, Chen XR
and Qi JS: Val8-glucagon-like peptide-1 protects against
Aβ1-40-induced impairment of hippocampal late-phase long-term
potentiation and spatial learning in rats. Neuroscience.
170:1239–1248. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Himeno T, Kamiya H, Naruse K, Harada N,
Ozaki N, Seino Y, Shibata T, Kondo M, Kato J, Okawa T, et al:
Beneficial effects of exendin-4 on experimental polyneuropathy in
diabetic mice. Diabetes. 60:2397–2406. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Linnemann AK, Neuman JC, Battiola TJ,
Wisinski JA, Kimple ME and Davis DB: Glucagon-like peptide-1
regulates cholecystokinin production in β-cells to protect from
apoptosis. Mol Endocrinol. 29:978–987. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jin J, Kang HM, Jung J, Jeong JW and Park
C: Related expressional change of HIF-1α to the neuroprotective
activity of exendin-4 in transient global ischemia. Neuroreport.
25:65–70. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Islam MS and Loots du T: Experimental
rodent models of type 2 diabetes: A review. Methods Find Exp Clin
Pharmacol. 31:249–261. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen S, Liu AR, An FM, Yao WB and Gao XD:
Amelioration of neurodegenerative changes in cellular and rat
models of diabetes-related Alzheimer's disease by exendin-4. Age
(Dordr). 34:1211–1224. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dong Y, Kua ZJ, Khoo EY, Koo EH and
Merchant RA: The utility of brief cognitive tests for patients with
type 2 diabetes mellitus: A systematic review. J Am Med Dir Assoc.
17:889–895. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sadanand S, Balachandar R and Bharath S:
Memory and executive functions in persons with type 2 diabetes: A
meta-analysis. Diabetes Metab Res Rev. 32:132–142. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dar TA, Sheikh IA, Ganie SA, Ali R, Singh
LR, Gan SH, Kamal MA and Zargar MA: Molecular linkages between
diabetes and Alzheimer's disease: Current scenario and future
prospects. CNS Neurol Disord Drug Targets. 13:290–298. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kubis-Kubiak AM, Rorbach-Dolata A and
Piwowar A: Crucial players in Alzheimer's disease and diabetes
mellitus: Friends or foes? Mech Ageing Dev. 181:7–21. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang SB and Jia JP: Oxymatrine attenuates
diabetes-associated cognitive deficits in rats. Acta Pharmacol Sin.
35:331–338. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu J, Feng L, Ma D, Zhang M, Gu J, Wang
S, Fu Q, Song Y, Lan Z, Qu R and Ma S: Neuroprotective effect of
paeonol on cognition deficits of diabetic encephalopathy in
streptozotocin-induced diabetic rat. Neurosci Lett. 549:63–68.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu YW, Zhu X, Lu Q, Wang JY, Li W, Wei YQ
and Yin XX: Total saponins from Rhizoma Anemarrhenae ameliorate
diabetes-associated cognitive decline in rats: Involvement of
amyloid-beta decrease in brain. J Ethnopharmaco. 139:194–200. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
DeFronzo RA, Triplitt C, Qu Y, Lewis MS,
Maggs D and Glass LC: Effects of exenatide plus rosiglitazone on
beta-cell function and insulin sensitivity in subjects with type 2
diabetes on metformin. Diabetes Care. 33:951–957. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang ZJ, Han YF, Zhao F, Yang GZ, Yuan L,
Cai HY, Yang JT, Holscher C, Qi JS and Wu MN: A dual GLP-1 and Gcg
receptor agonist rescues spatial memory and synaptic plasticity in
APP/PS1 transgenic mice. Horm Behav. 118:1046402020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Perry T and Greig NH: Enhancing central
nervous system endogenous GLP-1 receptor pathways for intervention
in Alzheimer's disease. Curr Alzheimer Res. 2:377–385. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee HN, Shin SA, Choo GS, Kim HJ, Park YS,
Kim BS, Kim SK, Cho SD, Nam JS, Choi CS, et al: Anti-inflammatory
effect of quercetin and galangin in LPS-stimulated RAW264.7
macrophages and DNCB-induced atopic dermatitis animal models. Int J
Mol Med. 41:888–898. 2018.PubMed/NCBI
|
|
37
|
Harrison JC, Zyla TR, Bardes ES and Lew
DJ: Stress-specific activation mechanisms for the ‘cell integrity’
MAPK pathway. J Biol Chem. 279:2616–2622. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Huang HM and Liu JC: c-Jun blocks cell
differentiation but not growth inhibition or apoptosis of chronic
myelogenous leukemia cells induced by STI571 and by histone
deacetylase inhibitors. J Cell Physiol. 218:568–574. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li L, Feng Z and Porter AG: JNK-dependent
phosphorylation of c-Jun on serine 63 mediates nitric oxide-induced
apoptosis of neuroblastoma cells. J Biol Chem. 279:4058–4065. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jorda A, Aldasoro M, Aldasoro C,
Guerra-Ojeda S, Iradi A, Vila JM, Campos-Campos J and Valles SL:
Action of low doses of Aspirin in Inflammation and Oxidative stress
induced by aβ1-42 on astrocytes in primary culture. Int
J Med Sci. 17:834–843. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cheng FR, Cui HX, Fang JL, Yuan K and Guo
Y: Ameliorative effect and mechanism of the purified
anthraquinone-glycoside preparation from rheum palmatum L. on type
2 diabetes mellitus. Molecules. 24:14542019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Vilsbøll T, Christensen M, Junker AE, Knop
FK and Gluud LL: Effects of glucagon-like peptide-1 receptor
agonists on weight loss: Systematic review and meta-analyses of
randomised controlled trials. BMJ. 344:d77712012. View Article : Google Scholar : PubMed/NCBI
|